
Fuc-S—A New Ultrasonic Degraded Sulfated α-l-Fucooligosaccharide—Alleviates DSS-Inflicted Colitis through Reshaping Gut Microbiota and Modulating Host–Microbe Tryptophan Metabolism
 ,
,
Abstract
:1. Introduction
2. Results
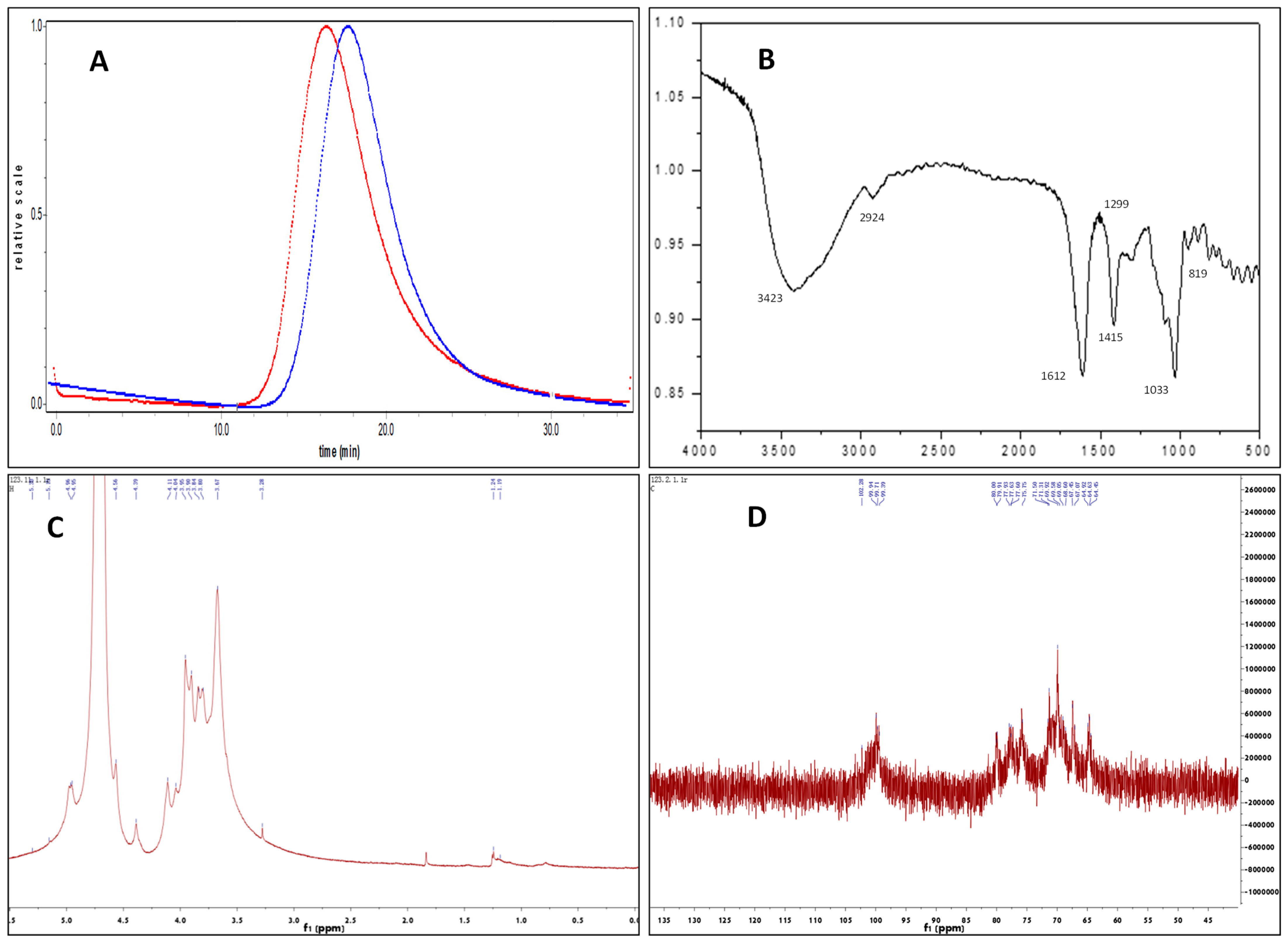
2.1. Structural Characterization of Fuc-S
2.1.1. Mw and Chemical Composition Analysis
2.1.2. FTIR Spectral Analysis
2.1.3. Methylation Analysis
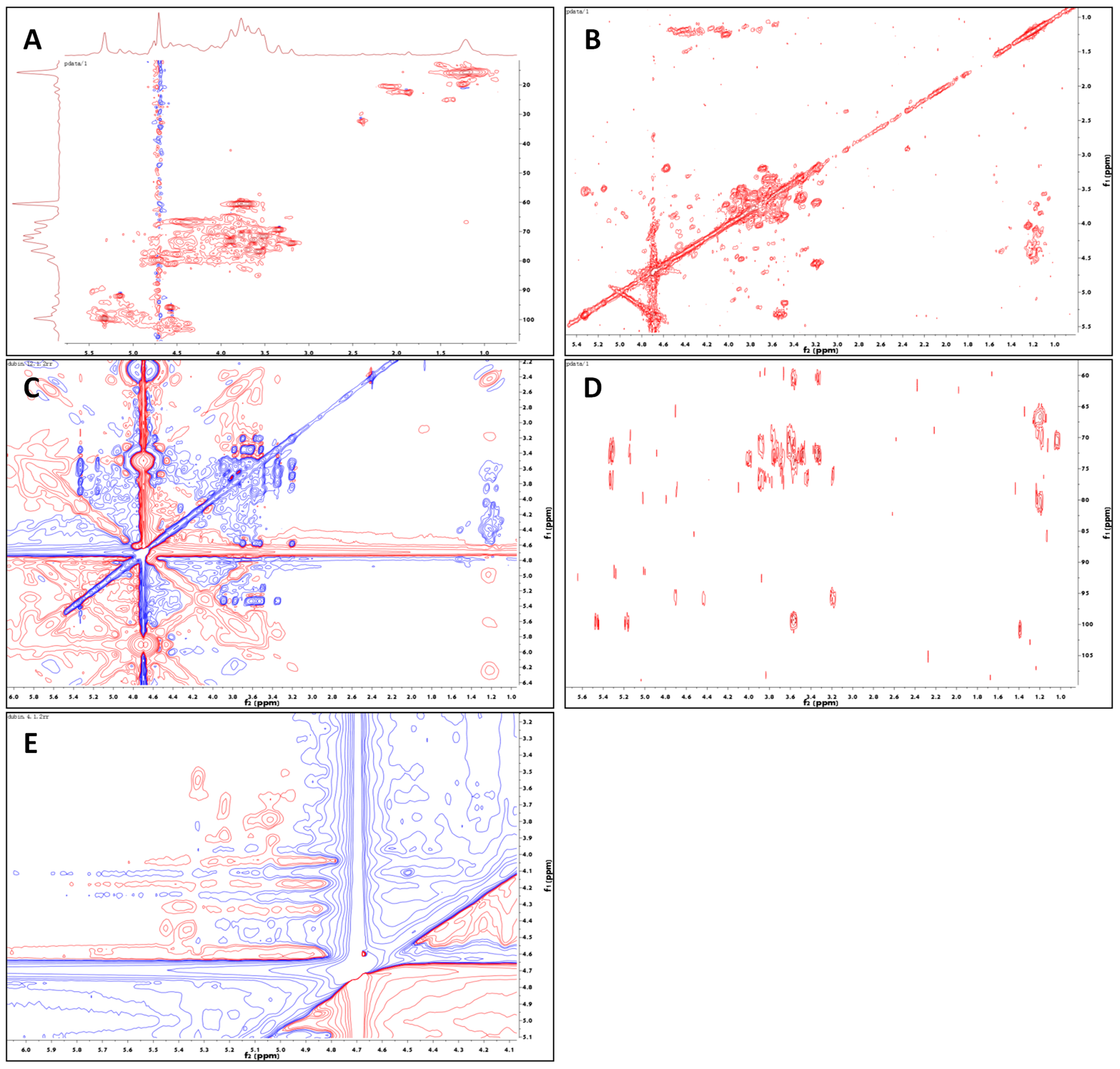
2.1.4. NMR Analysis
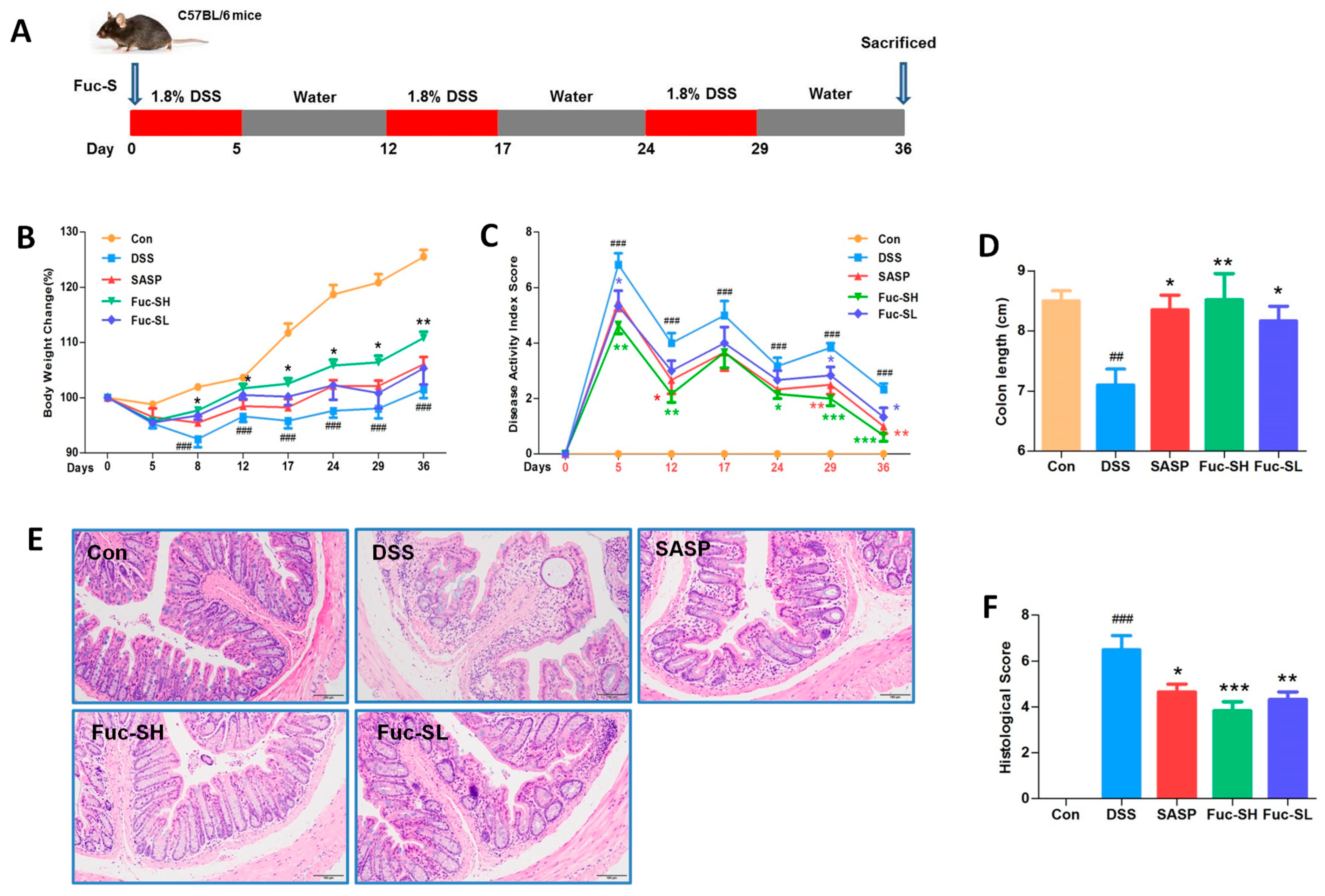
2.2. Anti-Colitis Effects of Fuc-S on DSS-Induced Chronic Colitis in Mice
2.2.1. Fuc-S Attenuates DSS-induced Chronic Colitis in Mice
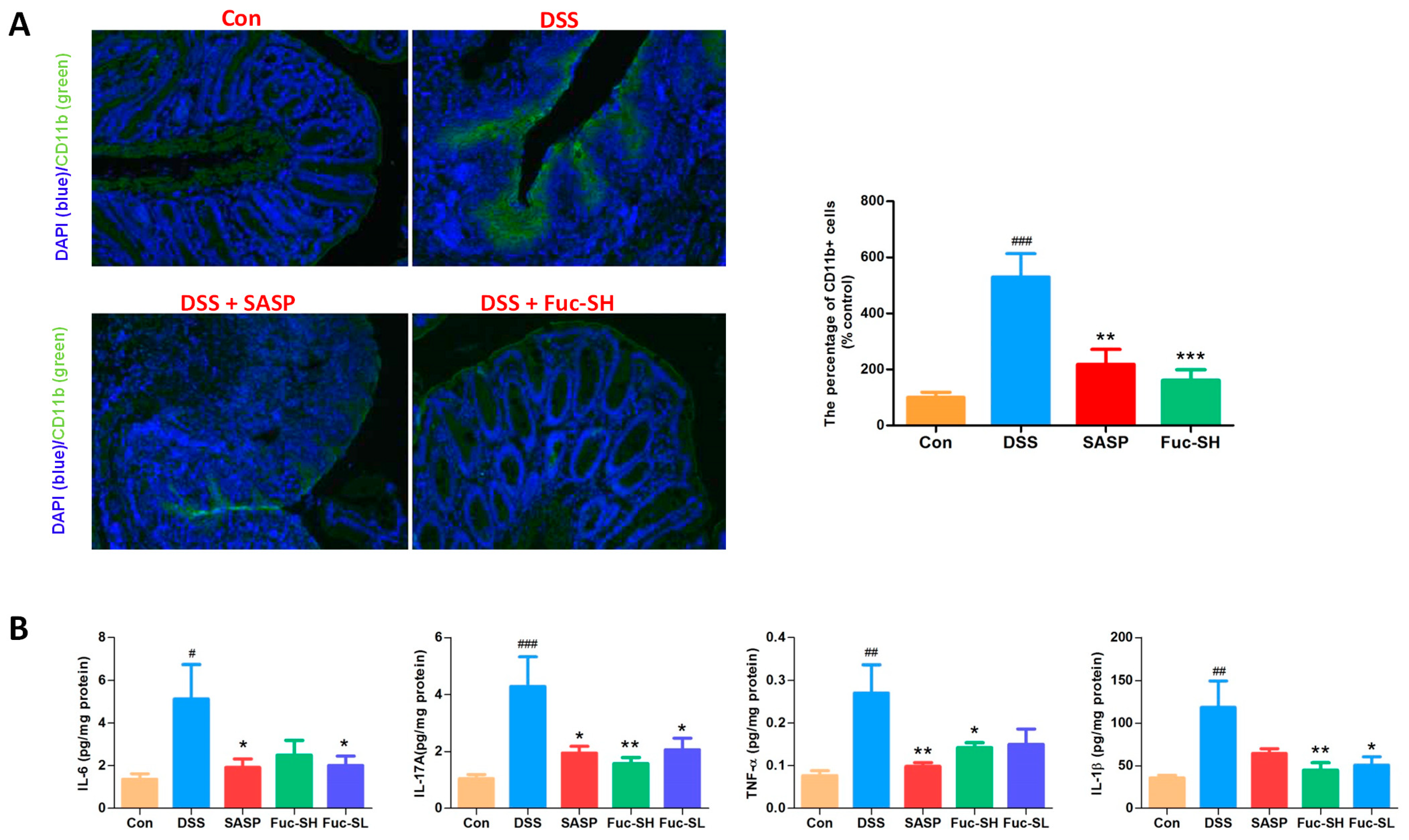
2.2.2. Fuc-S Attenuates Inflammatory Cell Infiltration and Pro-Inflammatory Cytokine Production in the Colon Tissues of DSS-Induced Chronic Colitic Mice
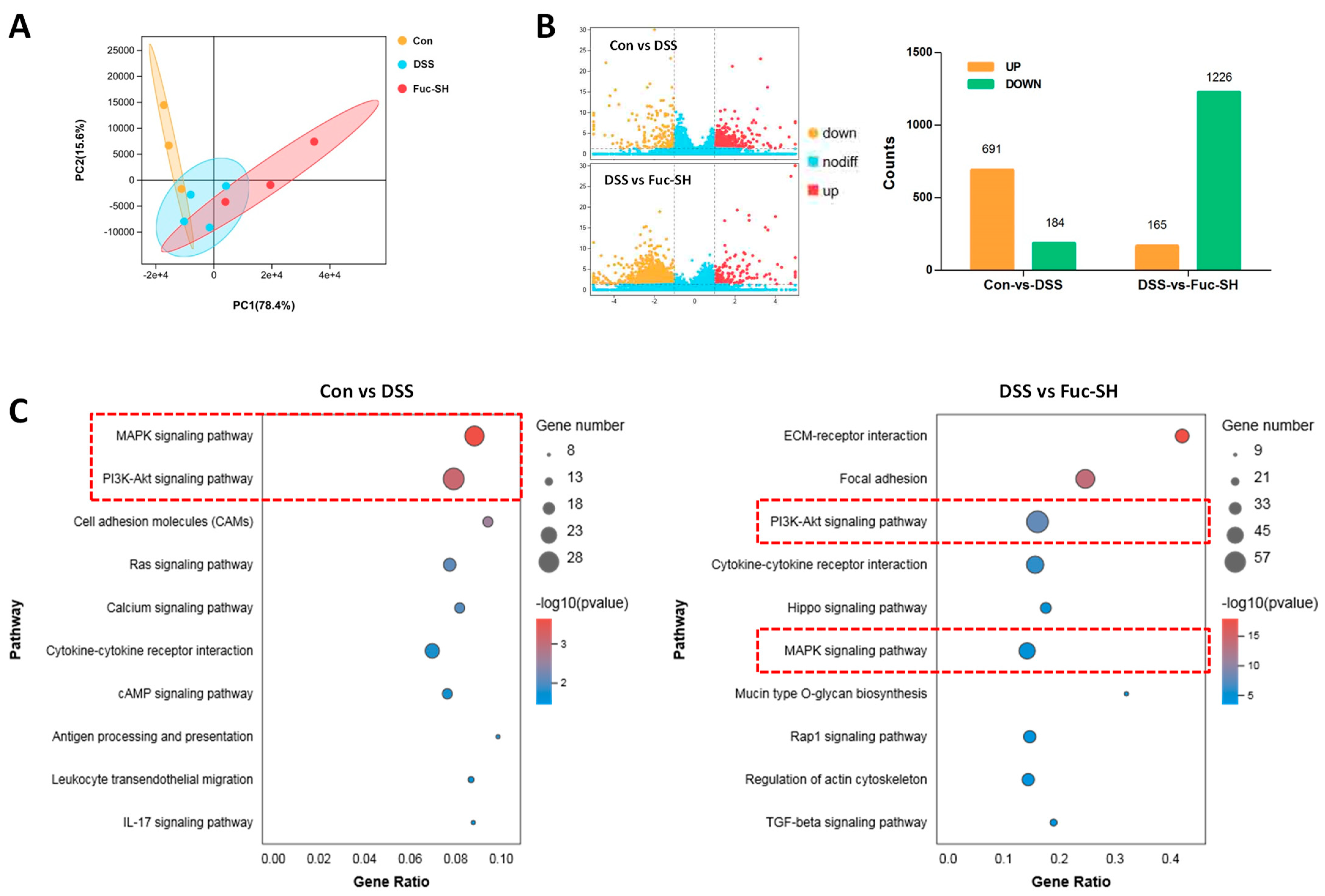
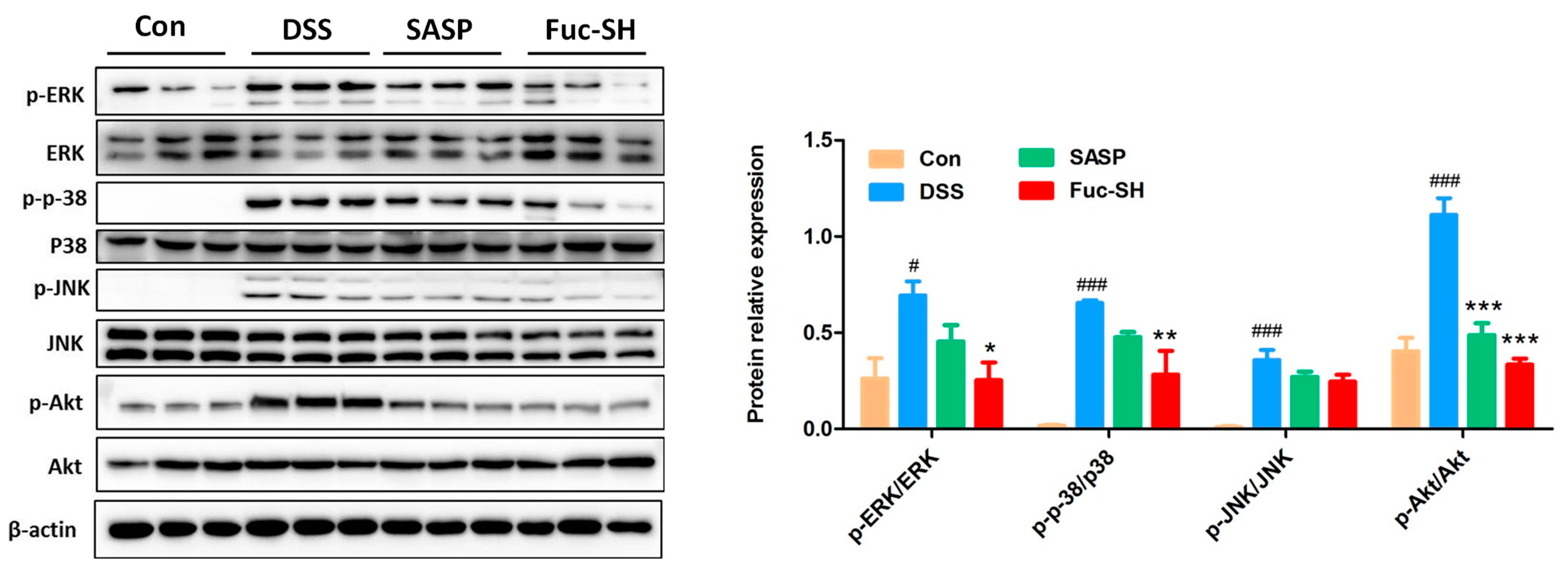
2.2.3. Transcriptomics Analysis Showed That Fuc-S Attenuates Colonic Inflammation by Regulating the PI3K-Akt and MAPK Signaling Pathways
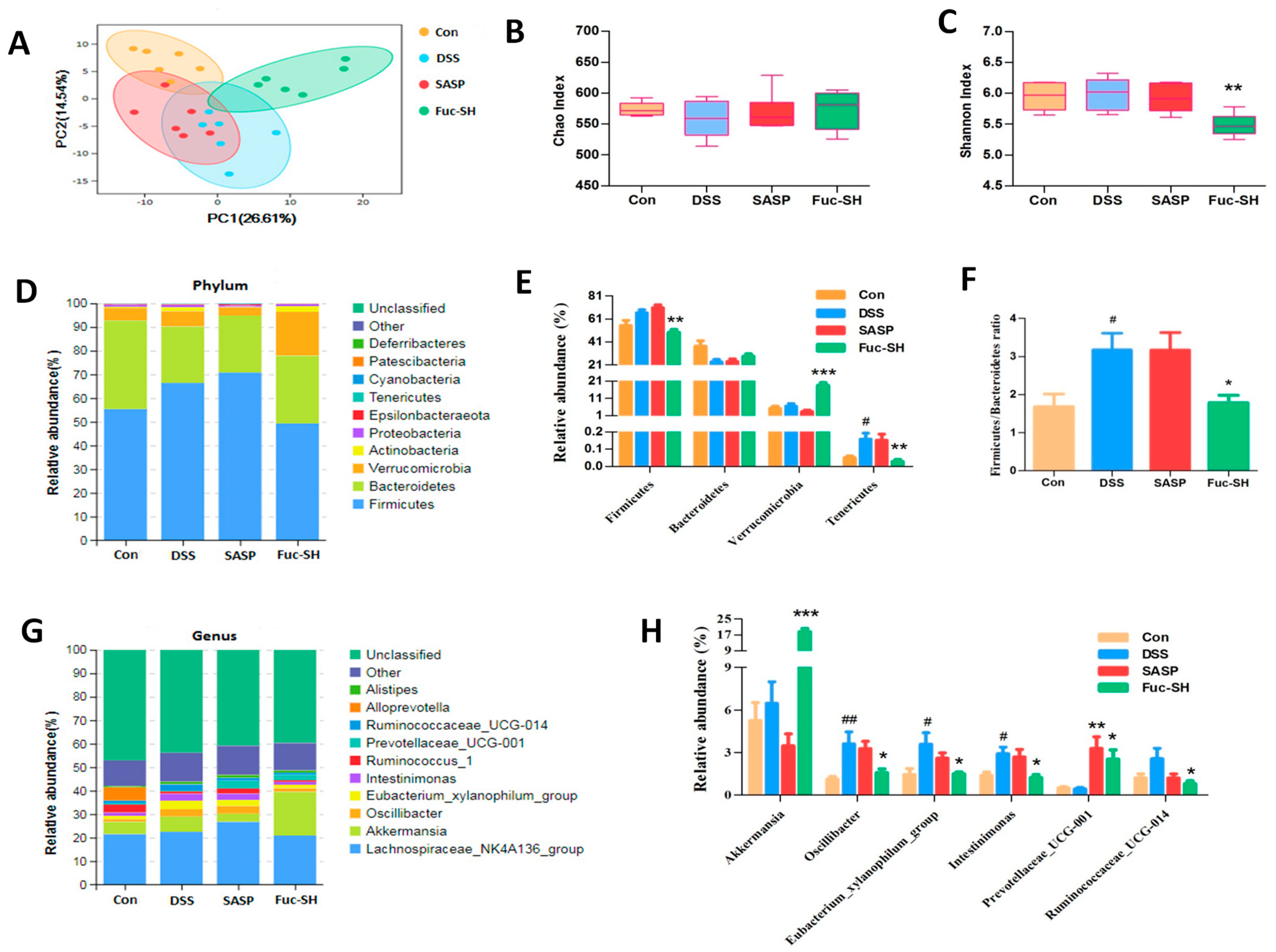
2.2.4. Fuc-S Regulates the Intestinal Microecology of DSS-Induced Chronic Colitic Mice
2.2.5. Fuc-S Modulates Host–Microbe Tryptophan Metabolism in DSS-Induced Chronic Colitis
3. Discussion
4. Materials and Methods
4.1. Preparation of Fuc-S
4.2. Molecular Weight Determination
4.3. Monosaccharide Composition Determination
4.4. Methylation Analysis
4.5. FTIR and NMR Spectroscopy
4.6. Animals and Experimental Design
4.7. Histopathological Evaluation and Immunofluorescence (IF) Staining
4.8. Luminex Detection and Western Blot Analysis
4.9. Transcriptomics by RNA-Sequencing Analysis
4.10. Microbiota Analysis
4.11. Metabolic Analysis of Tryptophan Metabolism
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hufnagel, B.; Muellner, V.; Hlatky, K.; Tallian, C.; Vielnascher, R.; Guebitz, G.M.; Wirth, M.; Gabor, F. Chemically modified inulin for intestinal drug delivery—A new dual bioactivity concept for inflammatory bowel disease treatment. Carbohydr. Polym. 2021, 252, 117091. [Google Scholar] [CrossRef]
- Peng, J.; Zheng, T.T.; Li, X.; Liang, Y.; Wang, L.J.; Huang, Y.C.; Xiao, H.T. Plant-derived alkaloids: The promising disease-modifying agents for inflammatory bowel disease. Front. Pharmacol. 2019, 10, 351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Xiao, H.T.; Wang, H.S.; Mu, H.X.; Zhao, L.; Du, J.; Yang, D.; Wang, D.; Bian, Z.X.; Lin, S.H. Halofuginone reduces the inflammatory responses of DSS-induced colitis through metabolic reprogramming. Mol. Biosyst. 2016, 12, 2296–2303. [Google Scholar] [CrossRef]
- Mei, Y.; Wang, Z.; Zhang, Y.; Wan, T.; Xue, J.; He, W.; Luo, Y.; Xu, Y.; Bai, X.; Wang, Q.; et al. FA-97, a new synthetic caffeic acid phenethyl ester derivative, ameliorates DSS-induced colitis against oxidative stress by activating Nrf2/HO-1 pathway. Front. Immunol. 2020, 10, 2969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Sanromán, A.; Esplugues, J.V.; Domènech, E. Pharmacology and safety of tofacitinib in ulcerative colitis. Gastroenterol. Hepatol. 2021, 44, 39–48, (In English, Spanish). [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.T.; Peng, J.; Wen, B.; Hu, D.D.; Hu, X.P.; Shen, X.C.; Liu, Z.G.; He, Z.D.; Bian, Z.X. Indigo naturalis suppresses colonic oxidative stress and Th1/Th17 responses of DSS-induced colitis in mice. Oxid. Med. Cell. Longev. 2019, 2019, 9480945. [Google Scholar] [CrossRef] [PubMed]
- Bibi, S.; de Sousa Moraes, L.F.; Lebow, N.; Zhu, M.J. Dietary green pea protects against DSS-induced colitis in mice challenged with high-fat diet. Nutrient 2017, 9, 509. [Google Scholar] [CrossRef] [Green Version]
- Durchschein, F.; Petritsch, W.; Hammer, H.F. Diet therapy for inflammatory bowel diseases: The established and the new. World J. Gastroenterol. 2016, 22, 2179–2194. [Google Scholar] [CrossRef]
- Mijan, M.A.; Lim, B.O. Diets, functional foods, and nutraceuticals as alternative therapies for inflammatory bowel disease: Present status and future trends. World J. Gastroenterol. 2018, 24, 2673–2685. [Google Scholar] [CrossRef]
- Vargas-Robles, H.; Castro-Ochoa, K.F.; Citalán-Madrid, A.F.; Schnoor, M. Beneficial effects of nutritional supplements on intestinal epithelial barrier functions in experimental colitis models in vivo. World J. Gastroenterol. 2019, 25, 4181–4198. [Google Scholar] [CrossRef]
- Wędrychowicz, A.; Zając, A.; Tomasik, P. Advances in nutritional therapy in inflammatory bowel diseases: Review. World J. Gastroenterol. 2016, 22, 1045–1066. [Google Scholar] [CrossRef]
- Wong, C.; Harris, P.J.; Ferguson, L.R. Potential benefits of dietary fibre intervention in inflammatory bowel disease. Int. J. Mol. Sci. 2016, 17, 919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apostolova, E.; Lukova, P.; Baldzhieva, A.; Katsarov, P.; Nikolova, M.; Iliev, I.; Peychev, L.; Trica, B.; Oancea, F.; Delattre, C.; et al. Immunomodulatory and Anti-Inflammatory Effects of Fucoidan: A Review. Polymers 2020, 12, 2338. [Google Scholar] [CrossRef]
- Besednova, N.N.; Zaporozhets, T.S.; Kuznetsova, T.A.; Makarenkova, I.D.; Kryzhanovsky, S.P.; Fedyanina, L.N.; Ermakova, S.P. Extracts and marine algae polysaccharides in therapy and prevention of inflammatory diseases of the intestine. Mar. Drugs 2020, 18, 289. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.E.; Kim, H.; Seo, C.; Park, T.; Lee, K.B.; Yoo, S.Y.; Hong, S.C.; Kim, J.T.; Lee, J. Marine polysaccharides: Therapeutic efficacy and biomedical applications. Arch. Pharm. Res. 2017, 40, 1006–1020. [Google Scholar] [CrossRef] [PubMed]
- Mabate, B.; Daub, C.D.; Malgas, S.; Edkins, A.L.; Pletschke, B.I. Fucoidan structure and its impact on glucose metabolism: Implications for diabetes and cancer therapy. Mar. Drugs 2021, 19, 30. [Google Scholar] [CrossRef]
- Usoltseva, R.V.; Malyarenko, O.S.; Anastyuk, S.D.; Shevchenko, N.M.; Silchenko, A.S.; Zvyagintseva, T.N.; Isakov, V.V.; Thinh, P.D.; Khanh, H.H.N.; Hang, C.T.T.; et al. The structure of fucoidan from Sargassum oligocystum and radiosensitizing activity of galactofucans from some algae of genus Sargassum. Int. J. Biol. Macromol. 2021, 183, 1427–1435. [Google Scholar] [CrossRef]
- Liu, X.; Xi, X.; Jia, A.; Zhang, M.; Cui, T.; Bai, X.; Shi, Y.; Liu, C. A fucoidan from Sargassum fusiforme with novel structure and its regulatory effects on intestinal microbiota in high-fat diet-fed mice. Food Chem. 2021, 358, 129908. [Google Scholar] [CrossRef]
- Chen, X.; Siu, K.C.; Cheung, Y.C.; Wu, J.Y. Structure and properties of a (1→3)-β-D-glucan from ultrasound-degraded exopolysaccharides of a medicinal fungus. Carbohydr. Polym. 2014, 106, 270–275. [Google Scholar] [CrossRef]
- Park, S.B.; Chun, K.R.; Kim, J.K.; Suk, K.; Jung, Y.M.; Lee, W.H. The differential effect of high and low molecular weight fucoidans on the severity of collagen-induced arthritis in mice. Phytother. Res. 2010, 24, 1384–1391. [Google Scholar] [CrossRef]
- Wang, Z.M.; Cheung, Y.C.; Leung, P.H.; Wu, J.Y. Ultrasonic treatment for improved solution properties of a high-molecular weight exopolysaccharide produced by a medicinal fungus. Bioresour. Technol. 2010, 101, 5517–5522. [Google Scholar] [CrossRef]
- Li, C.; Huang, X.; Peng, Q.; Shan, Y.; Xue, F. Physicochemical properties of peanut protein isolate-glucomannan conjugates prepared by ultrasonic treatment. Ultrason. Sonochem. 2014, 21, 1722–1727. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.Y.; Ma, L.Y.; Siu, K.C.; Wu, J.Y. Effects of ultrasonication on the conformational, microstructural, and antioxidant properties of konjac glucomannan. Appl. Sci. 2019, 9, 461. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Q.; Li, W.; Liang, S.; Zhang, H.; Zhang, Y. Effects of ultrasonic treatment on the molecular weight and anti-inflammatory activity of oxidized konjac glucomannan. CYTA J. Food 2019, 17, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Tang, Q.; Zhang, J.; Xia, Y.; Yang, Y.; Wu, D.; Fan, H.; Cui, S.W. Triple helix conformation of β-d-glucan from Ganoderma lucidum and effect of molecular weight on its immunostimulatory activity. Int. J. Biol. Macromol. 2018, 114, 1064–1070. [Google Scholar] [CrossRef]
- Cavallero, G.J.; Ferreira, M.L.; Casabuono, A.C.; Ramírez, S.A.; Vullo, D.L.; Couto, A.S. Structural characterization and metal biosorptive activity of the major polysaccharide produced by Pseudomonas veronii 2E. Carbohydr. Polym. 2020, 245, 116458. [Google Scholar] [CrossRef]
- Gao, X.; Qi, J.; Ho, C.T.; Li, B.; Mu, J.; Zhang, Y.; Hu, H.; Mo, W.; Chen, Z.; Xie, Y. Structural characterization and immunomodulatory activity of a water-soluble polysaccharide from Ganoderma leucocontextum fruiting bodies. Carbohydr. Polym. 2020, 249, 116874. [Google Scholar] [CrossRef]
- Chen, Y.; Mao, W.; Gao, Y.; Teng, X.; Zhu, W.; Chen, Y.; Zhao, C.; Li, N.; Wang, C.; Yan, M.; et al. Structural elucidation of an extracellular polysaccharide produced by the marine fungus Aspergillus versicolor. Carbohydr. Polym. 2013, 93, 478–483. [Google Scholar] [CrossRef]
- Yang, Y.; Hu, T.; Li, J.; Xin, M.; Zhao, X. Structural characterization and effect on leukopenia of fucoidan from Durvillaea antarctica. Carbohydr. Polym. 2021, 256, 117529. [Google Scholar] [CrossRef]
- Kuang, M.T.; Li, J.Y.; Yang, X.B.; Yang, L.; Xu, J.Y.; Yan, S.; Lv, Y.F.; Ren, F.C.; Hu, J.M.; Zhou, J. Structural characterization and hypoglycemic effect via stimulating glucagon-like peptide-1 secretion of two polysaccharides from Dendrobium officinale. Carbohydr. Polym. 2020, 241, 116326. [Google Scholar] [CrossRef]
- Zhu, M.; Huang, R.; Wen, P.; Song, Y.; He, B.; Tan, J.; Hao, H.; Wang, H. Structural characterization and immunological activity of pectin polysaccharide from kiwano (Cucumis metuliferus) peels. Carbohydr. Polym. 2021, 254, 117371. [Google Scholar] [CrossRef] [PubMed]
- Alboofetileh, M.; Rezaei, M.; Tabarsa, M.; Rittà, M.; Donalisio, M.; Mariatti, F.; You, S.; Lembo, D.; Cravotto, G. Effect of different non-conventional extraction methods on the antibacterial and antiviral activity of fucoidans extracted from Nizamuddinia zanardinii. Int. J. Biol. Macromol. 2019, 124, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Wei, Y.; Liang, L.; Huang, L.; Yu, G.; Li, Q. A novel low-molecular-mass pumpkin polysaccharide: Structural characterization, antioxidant activity, and hypoglycemic potential. Carbohydr. Polym. 2021, 251, 117090. [Google Scholar] [CrossRef]
- Chen, H.; Zeng, J.; Wang, B.; Cheng, Z.; Xu, J.; Gao, W.; Chen, K. Structural characterization and antioxidant activities of Bletilla striata polysaccharide extracted by different methods. Carbohydr. Polym. 2021, 266, 118149. [Google Scholar] [CrossRef] [PubMed]
- Qu, Y.; Li, C.; Zhang, C.; Zeng, R.; Fu, C. Optimization of infrared-assisted extraction of Bletilla striata polysaccharides based on response surface methodology and their antioxidant activities. Carbohydr. Polym. 2016, 148, 345–353. [Google Scholar] [CrossRef]
- Bulyhina, T.V.; Zdorovenko, E.L.; Varbanets, L.D.; Shashkov, A.S.; Kadykova, A.A.; Knirel, Y.A.; Lushchak, O.V. Structure of O-polysaccharide and lipid A of pantoea agglomerans 8488. Biomolecule 2020, 10, 804. [Google Scholar] [CrossRef]
- He, W.; Sun, H.; Su, L.; Zhou, D.; Zhang, X.; Shanggui, D.; Chen, Y. Structure and anticoagulant activity of a sulfated fucan from the sea cucumber Acaudina leucoprocta. Int. J. Biol. Macromol. 2020, 164, 87–94. [Google Scholar] [CrossRef]
- Usoltseva, R.V.; Shevchenko, N.M.; Malyarenko, O.S.; Anastyuk, S.D.; Kasprik, A.E.; Zvyagintsev, N.V.; Ermakova, S.P. Fucoidans from brown algae Laminaria longipes and Saccharina cichorioides: Structural characteristics, anticancer and radiosensitizing activity in vitro. Carbohydr. Polym. 2019, 221, 157–165. [Google Scholar] [CrossRef]
- Li, Y.H.; Xiao, H.T.; Hu, D.D.; Fatima, S.; Lin, C.Y.; Mu, H.X.; Lee, N.P.; Bian, Z.X. Berberine ameliorates chronic relapsing dextran sulfate sodium-induced colitis in C57BL/6 mice by suppressing Th17 responses. Pharmacol. Res. 2016, 110, 227–239. [Google Scholar] [CrossRef]
- Xiao, H.T.; Lin, C.Y.; Ho, D.H.; Peng, J.; Chen, Y.; Tsang, S.W.; Wong, M.; Zhang, X.J.; Zhang, M.; Bian, Z.X. Inhibitory effect of the gallotannin corilagin on dextran sulfate sodium-induced murine ulcerative colitis. J. Nat. Prod. 2013, 76, 2120–2125. [Google Scholar] [CrossRef]
- Du, S.Y.; Huang, H.F.; Li, X.Q.; Zhai, L.X.; Zhu, Q.C.; Zheng, K.; Song, X.; Xu, C.S.; Li, C.Y.; Li, Y.; et al. Anti-inflammatory properties of uvaol on DSS-induced colitis and LPS-stimulated macrophages. Chin. Med. 2020, 15, 43. [Google Scholar] [CrossRef]
- Zuo, T.; Ng, S.C. The gut microbiota in the pathogenesis and therapeutics of inflammatory bowel disease. Front. Microbiol. 2018, 9, 2247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamas, B.; Richard, M.L.; Leducq, V.; Pham, H.P.; Michel, M.L.; Da Costa, G.; Bridonneau, C.; Jegou, S.; Hoffmann, T.W.; Natividad, J.M.; et al. CARD9 impacts colitis by altering gut microbiota metabolism of tryptophan into aryl hydrocarbon receptor ligands. Nat. Med. 2016, 22, 598–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monteleone, I.; Rizzo, A.; Sarra, M.; Sica, G.; Sileri, P.; Biancone, L.; MacDonald, T.T.; Pallone, F.; Monteleone, G. Aryl hydrocarbon receptor-induced signals up-regulate IL-22 production and inhibit inflammation in the gastrointestinal tract. Gastroenterology 2011, 141, 237–248. [Google Scholar] [CrossRef]
- Nikolaus, S.; Schulte, B.; Al-Massad, N.; Thieme, F.; Schulte, D.M.; Bethge, J.; Rehman, A.; Tran, F.; Aden, K.; Häsler, R.; et al. Increased tryptophan metabolism is associated with activity of inflammatory bowel diseases. Gastroenterology 2017, 153, 1504–1516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, J.H.; Lee, Y.K.; Shon, W.J.; Kim, B.; Jeon, C.O.; Cho, J.Y.; Morse, H.C., 3rd; Choi, E.Y.; Shin, D.M. Gut microorganisms and their metabolites modulate the severity of acute colitis in a tryptophan metabolism-dependent manner. Eur. J. Nutr. 2020, 59, 3591–3601. [Google Scholar] [CrossRef] [PubMed]
- Sofia, M.A.; Ciorba, M.A.; Meckel, K.; Lim, C.K.; Guillemin, G.J.; Weber, C.R.; Bissonnette, M.; Pekow, J.R. Tryptophan metabolism through the kynurenine pathway is associated with endoscopic inflammation in ulcerative colitis. Inflamm. Bowel Dis. 2018, 24, 1471–1480. [Google Scholar] [CrossRef]
- Zayed, A.; Ulber, R. Fucoidan production: Approval key challenges and opportunities. Carbohydr. Polym. 2019, 211, 289–297. [Google Scholar] [CrossRef]
- van Weelden, G.; Bobiński, M.; Okła, K.; van Weelden, W.J.; Romano, A.; Pijnenborg, J.M.A. Fucoidan structure and activity in relation to anti-cancer mechanisms. Mar. Drugs 2019, 17, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zayed, A.; El-Aasr, M.; Ibrahim, A.S.; Ulber, R. Fucoidan characterization: Determination of purity and physicochemical and chemical properties. Mar. Drugs 2020, 18, 571. [Google Scholar] [CrossRef]
- Koh, H.S.A.; Lu, J.; Zhou, W. Structure characterization and antioxidant activity of fucoidan isolated from Undaria pinnatifida grown in New Zealand. Carbohydr. Polym. 2019, 212, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Ermakova, S.; Sokolova, R.; Kim, S.M.; Um, B.H.; Isakov, V.; Zvyagintseva, T. Fucoidans from brown seaweeds Sargassum hornery, Eclonia cava, Costaria costata: Structural characteristics and anticancer activity. Appl. Biochem. Biotechnol. 2011, 164, 841–850. [Google Scholar] [CrossRef] [PubMed]
- Broom, O.J.; Widjaya, B.; Troelsen, J.; Olsen, J.; Nielsen, O.H. Mitogen activated protein kinases: A role in inflammatory bowel disease? Clin. Exp. Immunol. 2009, 158, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Lertnimitphun, P.; Jiang, Y.; Kim, N.; Fu, W.; Zheng, C.; Tan, H.; Zhou, H.; Zhang, X.; Pei, W.; Lu, Y.; et al. Safranal alleviates dextran sulfate sodium-induced colitis and suppresses macrophage-mediated inflammation. Front. Pharmacol. 2019, 10, 1281. [Google Scholar] [CrossRef] [PubMed]
- Roy, P.K.; Rashid, F.; Bragg, J.; Ibdah, J.A. Role of the JNK signal transduction pathway in inflammatory bowel disease. World J. Gastroenterol. 2008, 14, 200–202. [Google Scholar] [CrossRef]
- Huang, X.L.; Xu, J.; Zhang, X.H.; Qiu, B.Y.; Peng, L.; Zhang, M.; Gan, H.T. PI3K/Akt signaling pathway is involved in the pathogenesis of ulcerative colitis. Inflamm. Res. 2011, 60, 727–734. [Google Scholar] [CrossRef]
- Khan, M.W.; Keshavarzian, A.; Gounaris, E.; Melson, J.E.; Cheon, E.C.; Blatner, N.R.; Chen, Z.E.; Tsai, F.N.; Lee, G.; Ryu, H.; et al. PI3K/AKT signaling is essential for communication between tissue-infiltrating mast cells, macrophages, and epithelial cells in colitis-induced cancer. Clin. Cancer Res. 2013, 19, 2342–2354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.; Son, Y.S.; Lee, M.O.; Ryu, J.W.; Park, K.; Kwon, O.; Jung, K.B.; Kim, K.; Ryu, T.Y.; Baek, A.; et al. Low-dose interleukin-2 alleviates dextran sodium sulfate-induced colitis in mice by recovering intestinal integrity and inhibiting AKT-dependent pathways. Theranostics 2020, 10, 5048–5063. [Google Scholar] [CrossRef]
- Davenport, M.; Poles, J.; Leung, J.M.; Wolff, M.J.; Abidi, W.M.; Ullman, T.; Mayer, L.; Cho, I.; Loke, P. Metabolic alterations to the mucosal microbiota in inflammatory bowel disease. Inflamm. Bowel Dis. 2014, 20, 723–731. [Google Scholar] [CrossRef] [Green Version]
- Boopathi, S.; Liu, D.; Jia, A.Q. Molecular trafficking between bacteria determines the shape of gut microbial community. Gut Microbes 2021, 13, 1959841. [Google Scholar] [CrossRef]
- Derrien, M.; Van Baarlen, P.; Hooiveld, G.; Norin, E.; Müller, M.; de Vos, W.M. Modulation of mucosal immune response, tolerance, and proliferation in mice colonized by the mucin-degrader Akkermansia muciniphila. Front. Microbiol. 2011, 2, 166. [Google Scholar] [CrossRef] [Green Version]
- Plovier, H.; Everard, A.; Druart, C.; Depommier, C.; Van Hul, M.; Geurts, L.; Chilloux, J.; Ottman, N.; Duparc, T.; Lichtenstein, L.; et al. A purified membrane protein from Akkermansia muciniphila or the pasteurized bacterium improves metabolism in obese and diabetic mice. Nat. Med. 2017, 23, 107–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, N.R.; Lee, J.C.; Lee, H.Y.; Kim, M.S.; Whon, T.W.; Lee, M.S.; Bae, J. An increase in the Akkermansia spp. population induced by metformin treatment improves glucose homeostasis in diet-induced obese mice. Gut 2014, 63, 727–735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bian, X.; Wu, W.; Yang, L.; Lv, L.; Wang, Q.; Li, Y.; Ye, J.; Fang, D.; Wu, J.; Jiang, X.; et al. Administration of Akkermansia muciniphila ameliorates dextran sulfate sodium-induced ulcerative colitis in mice. Front. Microbiol. 2019, 10, 2259. [Google Scholar] [CrossRef] [Green Version]
- Png, C.W.; Lindén, S.K.; Gilshenan, K.S.; Zoetendal, E.G.; McSweeney, C.S.; Sly, L.I.; McGuckin, M.A.; Florin, T.H. Mucolytic bacteria with increased prevalence in IBD mucosa augment in vitro utilization of mucin by other bacteria. Am. J. Gastroenterol. 2010, 105, 2420–2428. [Google Scholar] [CrossRef] [PubMed]
- Hall, A.B.; Yassour, M.; Sauk, J.; Garner, A.; Jiang, X.; Arthur, T.; Lagoudas, G.K.; Vatanen, T.; Fornelos, N.; Wilson, R.; et al. A novel Ruminococcus gnavus clade enriched in inflammatory bowel disease patients. Genome Med. 2017, 9, 103. [Google Scholar] [CrossRef]
- Mak, J.W.Y.; Ng, S.C. Epidemiology of fibrostenosing inflammatory bowel disease. J. Dig. Dis. 2020, 21, 332–335. [Google Scholar] [CrossRef]
- Lavelle, A.; Sokol, H. Gut microbiota-derived metabolites as key actors in inflammatory bowel disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 223–237. [Google Scholar] [CrossRef]
- Li, Y.; Hu, N.; Yang, D.; Oxenkrug, G.; Yang, Q. Regulating the balance between the kynurenine and serotonin pathways of tryptophan metabolism. FEBS J. 2017, 284, 948–966. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.F.; Tang, Q.J.; Feng, J.; Liu, J.; Tang, C.H.; Yan, M.Q.; Zhou, S.; Liu, L.P.; Zhou, J.; Zang, J.S. Efects of molecular weight on intestinal anti-inflammatory activities of β-D-glucan from Ganoderma lucidum. Front. Nutr. 2022, 9, 1028727. [Google Scholar] [CrossRef]
- Wu, Q.; Luo, M.; Yao, X.; Yu, L. Purification, structural characterization, and antioxidant activity of the COP-W1 polysaccharide from Codonopsis tangshen Oliv. Carbohydr. Polym. 2020, 236, 116020. [Google Scholar] [CrossRef]
- Rong, Y.; Yang, R.; Yang, Y.; Wen, Y.; Liu, S.; Li, C.; Hu, Z.; Cheng, X.; Li, W. Structural characterization of an active polysaccharide of longan and evaluation of immunological activity. Carbohydr. Polym. 2019, 213, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Wang, J.Y.; Huang, H.F.; Zheng, T.T.; Li, J.; Wang, L.J.; Ma, X.C.; Xiao, H.T. Adiponectin deficiency suppresses rhabdomyosarcoma associated with gut microbiota regulation. BioMed Res. Int. 2021, 2021, 8010694. [Google Scholar] [CrossRef]
- Mao, Y.H.; Song, A.X.; Li, L.Q.; Yang, Y.; Yao, Z.P.; Wu, J.Y. A high-molecular weight exopolysaccharide from the Cs-HK1 fungus: Ultrasonic degradation, characterization and in vitro fecal fermentation. Carbohydr. Polym. 2020, 246, 116636. [Google Scholar] [CrossRef] [PubMed]
- Du, B.; Zeng, H.; Yang, Y.; Bian, Z.; Xu, B. Anti-inflammatory activity of polysaccharide from Schizophyllum commune as affected by ultrasonication. Int. J. Biol. Macromol. 2016, 91, 100–105. [Google Scholar] [CrossRef]
- Du, B.; Jeepipalli, S.P.K.; Xu, B. Critical review on alterations in physiochemical properties and molecular structure of natural polysaccharides upon ultrasonication. Ultrason. Sonochem. 2022, 90, 106170. [Google Scholar] [CrossRef] [PubMed]
- Suprunchuk, V. Ultrasonic-treated fucoidan as a promising therapeutic agent. Polym. Med. 2021, 51, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Nishida, A.; Inoue, R.; Inatomi, O.; Bamba, S.; Naito, Y.; Andoh, A. Gut microbiota in the pathogenesis of inflammatory bowel disease. Clin. J. Gastroenterol. 2018, 11, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Sun, Q.; Cheng, L.; Zeng, X.; Zhang, X.; Wu, Z.; Weng, P. The modulatory effect of plant polysaccharides on gut flora and the implication for neurodegenerative diseases from the perspective of the microbiota-gut-brain axis. Int. J. Biol. Macromol. 2020, 164, 1484–1492. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Retention Time (min) | Permethylated Alditol Acetate | Linkages | Molar Ratio | Mass Fragments (m/z) |
|---|---|---|---|---|
| 12.975 | 2,4-Me2-Fuc | →3)-Fuc-(1→ | 0.43 | 263, 233, 201, 173, 155, 129, 111, 101, 87, 71, 57, 43 |
| 11.439 | 2,3-Me2-Fuc | →4)-Fuc-(1→ | 0.15 | 283, 233, 204, 162, 142, 131, 118, 99, 87, 71, 43 |
| 15.868 | 2,6-Me2-Glc | →3, 4)-Glc-(1→ | 0.11 | 305, 253, 231, 203, 185, 143, 129, 118, 87, 71, 43 |
| 13.449 | 3-Me-Glc | →2, 4, 6)-Glc-(1→ | 0.10 | 261, 231, 201, 142, 127, 118, 99, 85, 71, 43 |
| 14.393 | 2-Me-Gal | →3, 4, 6)-Gal-(1→ | 0.07 | 333, 267, 202, 183, 160, 139, 127, 118, 85, 57, 43 |
| 13.817 | 2,3,4-Me3-Gal | →6)-Gal-(1→ | 0.06 | 292, 249, 233, 203, 189, 173, 162, 129, 102, 87, 71, 43 |
| 15.254 | 2,4,6-Me3-Man | →3)-Man-(1→ | 0.03 | 277, 183, 162, 146, 131, 118, 84, 59, 43 |
| 17.324 | 2,3,6-Me3-Glc | →4)-GlcUA-(1→ | 0.02 | 277, 233, 187, 162, 142, 131, 118, 99, 87, 71, 43 |
| 18.548 | 2,4-Me2-Gal | →3, 6)-GalUA-(1→ | 0.01 | 305, 234, 189, 160, 139, 129, 118, 99, 87, 71, 43 |
| 11.002 | 3,4-Me2-Xyl | →2)-Xyl-(1→ | 0.01 | 191, 175, 162, 118, 99, 87, 71, 59, 43 |
| 12.214 | 3,4- Me2-Rha | →2)-Rha-(1→ | 0.02 | 234, 190, 159, 129, 111, 99, 71, 43 |
| Structure No. | Glycosyl Residues | Chemical Shift (ppm) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| H1/C1 | H2/C2 | H3/C3 | H4/C4 | H5/C5 | H6/C6 | C=O | CH3 | ||
| A | →3)-α-L-Fucp-(1→ | 5.03/98.27 | 3.78/68.94 | 3.94/76.06 | 4.36/79.66 | 4.08/66.68 | 1.18/15.59 | ||
| B | →4)-α-L-Fucp-(1→ | 5.22/99.79 | 3.72/68.39 | 3.89/73.06 | 3.79/77.75 | 4.02/80.02 | 1.24/19.71 | ||
| C | →3,4)-α-D-Glcp-(1→ | 5.15/91.89 | 3.50/71.45 | 3.89/76.29 | 4.10/77.35 | 3.58/70.06 | 3.76/60.31 | ||
| D | →6)-α-D-Glap-(1→ | 5.32/99.62 | 3.54/71.46 | 3.91/72.62 | 3.77/71.26 | 3.59/72.62 | 3.30/69.24 | -/179.2 | 1.88/22.57 |
| E | →2,4,6)-α-D-Glcp-(1→ | 4.96/100.32 | 3.88/77.34 | 4.50/80.70 | 3.58/76.75 | 3.75/71.21 | 3.32/69.24 | ||
| F | →3,4,6)-β-D-Glap-(1→ | 4.57/95.88 | 3.20/73.92 | 3.69/76.13 | 3.80/76.82 | 3.58/72.74 | 3.51/71.50 | -/180.10 | 2.42/32.41 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiao, H.; Feng, J.; Peng, J.; Wu, P.; Chang, Y.; Li, X.; Wu, J.; Huang, H.; Deng, H.; Qiu, M.; et al. Fuc-S—A New Ultrasonic Degraded Sulfated α-l-Fucooligosaccharide—Alleviates DSS-Inflicted Colitis through Reshaping Gut Microbiota and Modulating Host–Microbe Tryptophan Metabolism. Mar. Drugs 2023, 21, 16. https://doi.org/10.3390/md21010016
Xiao H, Feng J, Peng J, Wu P, Chang Y, Li X, Wu J, Huang H, Deng H, Qiu M, et al. Fuc-S—A New Ultrasonic Degraded Sulfated α-l-Fucooligosaccharide—Alleviates DSS-Inflicted Colitis through Reshaping Gut Microbiota and Modulating Host–Microbe Tryptophan Metabolism. Marine Drugs. 2023; 21(1):16. https://doi.org/10.3390/md21010016
Chicago/Turabian StyleXiao, Haitao, Jinxiu Feng, Jiao Peng, Peigen Wu, Yaoyao Chang, Xianqian Li, Jinhui Wu, Haifeng Huang, Huan Deng, Miao Qiu, and et al. 2023. "Fuc-S—A New Ultrasonic Degraded Sulfated α-l-Fucooligosaccharide—Alleviates DSS-Inflicted Colitis through Reshaping Gut Microbiota and Modulating Host–Microbe Tryptophan Metabolism" Marine Drugs 21, no. 1: 16. https://doi.org/10.3390/md21010016
APA StyleXiao, H., Feng, J., Peng, J., Wu, P., Chang, Y., Li, X., Wu, J., Huang, H., Deng, H., Qiu, M., Yang, Y., & Du, B. (2023). Fuc-S—A New Ultrasonic Degraded Sulfated α-l-Fucooligosaccharide—Alleviates DSS-Inflicted Colitis through Reshaping Gut Microbiota and Modulating Host–Microbe Tryptophan Metabolism. Marine Drugs, 21(1), 16. https://doi.org/10.3390/md21010016