
Application of Marine Natural Products against Alzheimer’s Disease: Past, Present and Future


Abstract
:
1. Introduction
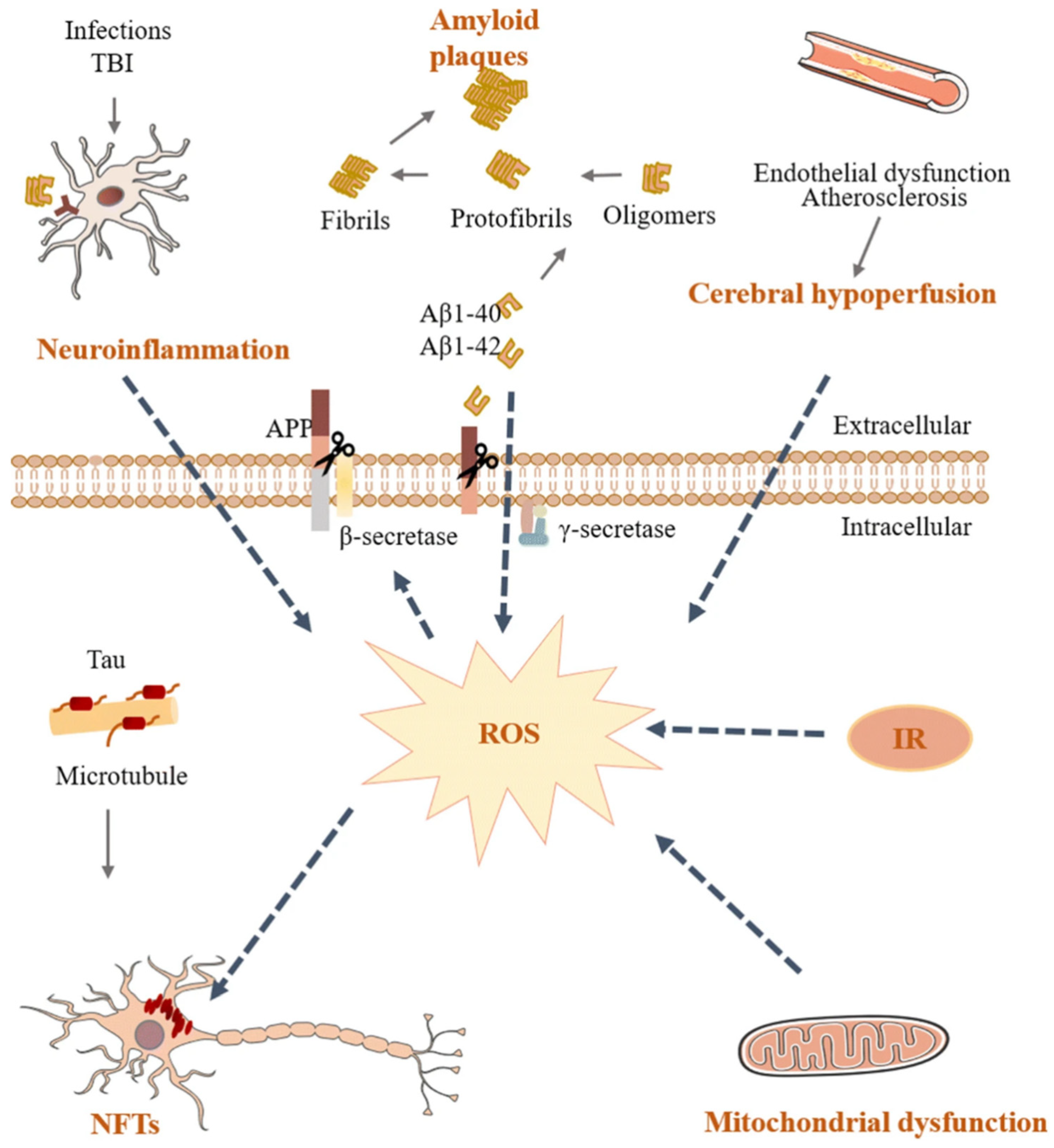
2. Pathogenesis of Alzheimer’s Disease
2.1. Amyloid Cascade Hypothesis
2.2. Neuroinflammation Hypothesis
2.3. Ca2+ Hypothesis
2.4. Tau Hypothesis
2.5. Cholinergic Hypothesis
2.6. Glutamate Hypothesis
3. Currently Approved Drugs for Alzheimer’s Disease
3.1. Acetylcholinesterase in-jibtor (AChEI)
3.1.1. Tacrine
3.1.2. Donepezil
3.1.3. Rivastigmine
3.1.4. Galantamine (GAL)
3.2. NMDAR Antagonists
3.3. Aducanumab
3.4. Sodium Oligomanne Capsules (GV-971)
3.5. Combination Drug Therapy
4. Anti-AD Marine Natural Products
4.1. Polysaccharides
4.2. Polyphenols
4.3. Sterols
4.4. Carotenoids
4.5. Diterpenoids
4.6. Alkaloids
4.7. Amino Sulfonates
4.8. Marine Toxins
4.9. Macrolide-Type Compound
5. Challenges and Opportunities in Developing Marine Natural Products for Alzheimer’s Disease
6. Remarks and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- 2021 Alzheimer’s Disease Facts and Figures—2021—Alzheimer’s & Dementia—Wiley Online Library. Available online: https://alz-journals.onlinelibrary.wiley.com/doi/10.1002/alz.12328 (accessed on 7 December 2022).
- Yan, X.; Hu, Y.; Wang, B.; Wang, S.; Zhang, X. Metabolic Dysregulation Contributes to the Progression of Alzheimer’s Disease. Front. Neurosci. 2020, 14, 530219. [Google Scholar] [CrossRef]
- Ballard, C.; Gauthier, S.; Corbett, A.; Brayne, C.; Aarsland, D.; Jones, E. Alzheimer’s Disease. Lancet 2011, 377, 1019–1031. [Google Scholar] [CrossRef] [PubMed]
- Cabrera, M.J.A.; Pérez, R.M.; Ravelo, A.G.; Rodríguez, D.H.; Davison, G.P. Patogenia y tratamientos actuales de la enfermedad de Alzheimer. Rev. Cuba. Farm. 2014, 48, 508–518. Available online: https://www.scienceopen.com/document?vid=85802991-f11a-4e69-8f79-6cc467306d4d. (accessed on 20 August 2022).
- Albert, M.S.; DeKosky, S.T.; Dickson, D.; Dubois, B.; Feldman, H.H.; Fox, N.C.; Gamst, A.; Holtzman, D.M.; Jagust, W.J.; Petersen, R.C.; et al. The diagnosis of mild cognitive impairment due to Alzheimer’s disease: Recommendations from the National Institute on Aging-Alzheimer’s Association workgroups on diagnostic guidelines for Alzheimer’s disease. Alzheimers Dement. 2011, 7, 270–279. [Google Scholar] [CrossRef] [Green Version]
- Neuropsychological Assessment, 5th Edition—ScienceOpen. Available online: https://www.scienceopen.com/document?vid=37ea5eff-64eb-42b1-95b4-b5de983e1424 (accessed on 26 September 2022).
- Alzheimer’s Disease Clinical and Research Update for Health Care Practitioners. Available online: https://www.hindawi.com/journals/jar/2013/207178/ (accessed on 26 September 2022).
- McKhann, G.M.; Knopman, D.S.; Chertkow, H.; Hyman, B.T.; Jack, C.R.J.; Kawas, C.H.; Klunk, W.E.; Koroshetz, W.J.; Manly, J.J.; Mayeux, R.; et al. The diagnosis of dementia due to Alzheimer’s disease: Recommendations from the National Institute on Aging-Alzheimer’s association workgroups on diagnostic guidelines for Alzheimer’s disease. Alzheimers Dement. 2011, 7, 263–269. [Google Scholar] [CrossRef] [Green Version]
- The Neuropsychological Profile of Alzheimer Disease. Available online: https://perspectivesinmedicine.cshlp.org/content/2/4/a006171 (accessed on 26 September 2022).
- Achterberg, W.P.; Pieper, M.J.; van Dalen-Kok, A.H.; De Waal, M.W.; Husebo, B.S.; Lautenbacher, S.; Kunz, M.; Scherder, E.J.; Corbett, A. Pain Management in Patients with Dementia. Clin. Interv. Aging 2013, 8, 1471–1482. [Google Scholar] [CrossRef] [Green Version]
- Jin, X.P.; Shen, M.; Vasculocardiology, D.O. Research Progress of Continuing Nursing Care for Patients with Hypertension after Discharging from Hospital. Chin. J. New Clin. Med. 2017, 9, 933–936. Available online: http://en.cnki.com.cn/Article_en/CJFDTotal-ZYLN201709033.htm (accessed on 26 September 2022).
- Ghoran, S.H.; Kijjoa, A. Marine-Derived Compounds with Anti-Alzheimer’s Disease Activities. Mar. Drugs 2021, 19, 410. [Google Scholar] [CrossRef]
- Hosseini, S.F.; Rezaei, M.; McClements, D.J. Bioactive Functional Ingredients from Aquatic Origin: A Review of Recent Progress in Marine-Derived Nutraceuticals. Crit. Rev. Food Sci. Nutr. 2020, 62, 1242–1269. [Google Scholar] [CrossRef]
- Bălașa, A.F.; Chircov, C.; Grumezescu, A.M. Marine Biocompounds for Neuroprotection—A Review. Mar. Drugs 2020, 18, 290. [Google Scholar] [CrossRef]
- Altmann, K.-H. Drugs from the Oceans: Marine Natural Products as Leads for Drug Discovery. Chim. Int. J. Chem. 2017, 71, 646–652. [Google Scholar] [CrossRef] [PubMed]
- Khotimchenko, Y.S.; Silachev, D.N.; Katanaev, V.L. Marine Natural Products from the Russian Pacific as Sources of Drugs for Neurodegenerative Diseases. Mar. Drugs 2022, 20, 708. [Google Scholar] [CrossRef] [PubMed]
- Wattmo, C.; Wallin, Å.K. Early-versus Late-Onset Alzheimer’s Disease in Clinical Practice: Cognitive and Global Outcomes over 3 Years. Alzheimers Res. Ther. 2017, 9, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finneran, D.J.; Nash, K.R. Neuroinflammation and fractalkine signaling in Alzheimer’s disease. J. Neuroinflamm. 2019, 16, 30. [Google Scholar] [CrossRef]
- Despres, C.; Byrne, C.; Qi, H.; Cantrelle, F.-X.; Huvent, I.; Chambraud, B.; Baulieu, E.-E.; Jacquot, Y.; Landrieu, I.; Lippens, G.; et al. Identification of the Tau phosphorylation pattern that drives its aggregation. Proc. Natl. Acad. Sci. USA 2017, 114, 9080–9085. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Gao, J.; Zhu, M.; Liu, K.; Zhang, H.-L. Gut Microbiota and Dysbiosis in Alzheimer’s Disease: Implications for Pathogenesis and Treatment. Mol. Neurobiol. 2020, 57, 5026–5043. [Google Scholar] [CrossRef]
- Tian, Y.; Meng, L.; Zhang, Z. What is strain in neurodegenerative diseases? Cell. Mol. Life Sci. 2020, 77, 665–676. [Google Scholar] [CrossRef]
- Tiwari, S.; Atluri, V.; Kaushik, A.; Yndart, A.; Nair, M. Alzheimer’s disease: Pathogenesis, diagnostics, and therapeutics. Int. J. Nanomed. 2019, 14, 5541–5554. [Google Scholar] [CrossRef] [Green Version]
- Hardy, J.A.; Higgins, G.A. Alzheimer’s disease: The amyloid cascade hypothesis. Science 1992, 256, 184–185. [Google Scholar] [CrossRef]
- Silva, M.; Seijas, P.; Otero, P. Exploitation of Marine Molecules to Manage Alzheimer’s Disease. Mar. Drugs 2021, 19, 373. [Google Scholar] [CrossRef]
- Kinney, J.W.; BeMiller, S.M.; Murtishaw, A.S.; Leisgang, A.M.; Salazar, A.M.; Lamb, B.T. Inflammation as a central mechanism in Alzheimer’s disease. Alzheimers Dement. Transl. Res. Clin. Interv. 2018, 4, 575–590. [Google Scholar] [CrossRef] [PubMed]
- Pascoal, T.A.; Benedet, A.L.; Ashton, N.J.; Kang, M.S.; Therriault, J.; Chamoun, M.; Savard, M.; Lussier, F.Z.; Tissot, C.; Karikari, T.K.; et al. Microglial activation and tau propagate jointly across Braak stages. Nat. Med. 2021, 27, 1592–1599. [Google Scholar] [CrossRef] [PubMed]
- Zhong, W.; Wu, A.; Berglund, K.; Gu, X.; Jiang, M.Q.; Talati, J.; Zhao, J.; Wei, L.; Yu, S.P. Pathogenesis of sporadic Alzheimer’s disease by deficiency of NMDA receptor subunit GluN3A. Alzheimers Dement. 2021, 18, 222–239. [Google Scholar] [CrossRef]
- Naseri, N.N.; Wang, H.; Guo, J.; Sharma, M.; Luo, W. The complexity of tau in Alzheimer’s disease. Neurosci. Lett. 2019, 705, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Van der Jeugd, A.; Hochgräfe, K.; Ahmed, T.; Decker, J.M.; Sydow, A.; Hofmann, A.; Wu, D.; Messing, L.; Balschun, D.; D’Hooge, R.; et al. Cognitive defects are reversible in inducible mice expressing pro-aggregant full-length human Tau. Acta Neuropathol. 2012, 123, 787–805. [Google Scholar] [CrossRef] [Green Version]
- Ferreira-Vieira, T.H.; Guimaraes, I.M.; Silva, F.R.; Ribeiro, F.M. Alzheimer’s disease: Targeting the Cholinergic System. Curr. Neuropharmacol. 2016, 14, 101–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenberg, P.B.; Nowrangi, M.A.; Lyketsos, C.G. Neuropsychiatric symptoms in Alzheimer’s disease: What might be associated brain circuits? Mol. Asp. Med. 2015, 43–44, 25–37. [Google Scholar] [CrossRef] [Green Version]
- Martorana, A.; Koch, G. “Is Dopamine Involved in Alzheimer’s Disease?”. Front. Aging Neurosci. 2014, 6, 252. [Google Scholar] [CrossRef] [Green Version]
- Nitsch, R.M.; Deng, M.; Tennis, M.; Schoenfeld, D.; Growdon, J.H. The selective muscarinic M1 agonist AF102B decreases levels of total Abeta in cerebrospinal fluid of patients with Alzheimer’s disease. Ann. Neurol. 2000, 48, 913–918. [Google Scholar] [CrossRef]
- Jeon, J.; Dencker, D.; Wörtwein, G.; Woldbye, D.P.D.; Cui, Y.; Davis, A.A.; Levey, A.I.; Schütz, G.; Sager, T.N.; Mørk, A.; et al. A Subpopulation of Neuronal M4 Muscarinic Acetylcholine Receptors Plays a Critical Role in Modulating Dopamine-Dependent Behaviors. J. Neurosci. 2010, 30, 2396–2405. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Chang, L.; Song, Y.; Li, H.; Wu, Y. The Role of NMDA Receptors in Alzheimer’s Disease. Front. Neurosci. 2019, 13, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Role of Glutamate and NMDA Receptors in Alzheimer’s Disease—IOS Press. Available online: https://content.iospress.com/articles/journal-of-alzheimers-disease/jad160763 (accessed on 26 September 2022).
- Breijyeh, Z.; Karaman, R. Comprehensive Review on Alzheimer’s Disease: Causes and Treatment. Molecules 2020, 25, 5789. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K. Cholinesterase Inhibitors as Alzheimer’s Therapeutics (Review). Mol. Med. Rep. 2019, 20, 1479–1487. [Google Scholar] [CrossRef] [Green Version]
- Green Valley Obtains IND Approval from U.S. FDA for Sodium Oligomannate’s International, Phase III Clinical Study—PR Newswire APAC. Available online: https://en.prnasia.com/story/278549-0.shtml (accessed on 3 November 2022).
- Carney, G.; Bassett, K.; Wright, J.M.; Maclure, M.; McGuire, N.; Dormuth, C.R. Comparison of cholinesterase inhibitor safety in real-world practice. Alzheimers Dement. Transl. Res. Clin. Interv. 2019, 5, 732–739. [Google Scholar] [CrossRef] [PubMed]
- Prince, M.; Bryce, R.; Albanese, E.; Wimo, A.; Ribeiro, W.; Ferri, C.P. The global prevalence of dementia: A systematic review and metaanalysis. Alzheimers Dement. 2013, 9, 63–75.e2. [Google Scholar] [CrossRef]
- Khoury, R.; Rajamanickam, J.; Grossberg, G.T. An update on the safety of current therapies for Alzheimer’s disease: Focus on rivastigmine. Ther. Adv. Drug Saf. 2018, 9, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Syed, Y.Y. Sodium Oligomannate: First Approval. Drugs 2020, 80, 441–444. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Gupta, V.; Sharma, S. Donepezil. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Birks, J.S.; Evans, J.G. Rivastigmine for Alzheimer’s disease. Cochrane Database Syst. Rev. 2015, 4, 1465–1858, CD001191. [Google Scholar] [CrossRef]
- Drugs to Treat Alzheimer’s Disease. J. Psychosoc. Nurs. Ment. Health Serv. 2018, 56, 5–6. [CrossRef]
- Léveillé, F.; El Gaamouch, F.; Gouix, E.; Lecocq, M.; Lobner, D.; Nicole, O.; Buisson, A. Neuronal viability is controlled by a functional relation between synaptic and extrasynaptic NMDA receptors. FASEB J. 2008, 22, 4258–4271. [Google Scholar] [CrossRef] [Green Version]
- Arndt, J.W.; Qian, F.; Smith, B.A.; Quan, C.; Kilambi, K.P.; Bush, M.W.; Walz, T.; Pepinsky, R.B.; Bussière, T.; Hamann, S.; et al. Structural and kinetic basis for the selectivity of aducanumab for aggregated forms of amyloid-β. Sci. Rep. 2018, 8, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biogen. A Phase 3 Multicenter, Randomized, Double-Blind, Placebo-Controlled, Parallel-Group Study to Evaluate the Efficacy and Safety of Aducanumab (BIIB037) in Subjects With Early Alzheimer’s Disease. 2021; NCT02484547. Available online: https://clinicaltrials.gov/ct2/show/NCT02484547 (accessed on 9 August 2021).
- Xiao, S.; Chan, P.; Wang, T.; Hong, Z.; Wang, S.; Kuang, W.; He, J.; Pan, X.; Zhou, Y.; Ji, Y.; et al. A 36-week multicenter, randomized, double-blind, placebo-controlled, parallel-group, phase 3 clinical trial of sodium oligomannate for mild-to-moderate Alzheimer’s dementia. Alzheimers Res. Ther. 2021, 13, 62. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sun, G.; Feng, T.; Zhang, J.; Huang, X.; Wang, T.; Geng, M. Sodium Oligomannate Therapeutically Remodels Gut Microbiota and Suppresses Gut Bacterial Amino Acids-Shaped Neuroinflammation to Inhibit Alzheimer’s Disease Progression. Cell Res. 2019, 29, 787–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, L.N.; Geng, M.Y.; Mu, L.; Xin, X.L.; Yang, N.; Zuo, P.P. Effects of Acidic Oligose on Differentially Expressed Genes in the Mice Model of Alzheimer’s Disease by Microarray. Acta Pharm. Sin. 2005, 40, 1105–1109. Available online: https://en.cnki.com.cn/Article_en/CJFDTotal-YXXB200512009.htm (accessed on 20 August 2022).
- Jiang, R.W.; Du, X.G.; Zhang, X.; Wang, X.; Hu, D.Y.; Meng, T.; Chen, Y.L.; Geng, M.Y.; Shen, J.K. Synthesis and Bioassay of β-(1,4)-D-Mannans as Potential Agents against Alzheimer’s Disease. Acta Pharmacol. Sin. 2013, 34, 1585–1591. [Google Scholar] [CrossRef] [Green Version]
- Cummings, J.L.; Tong, G.; Ballard, C. Treatment Combinations for Alzheimer’s Disease: Current and Future Pharmacotherapy Options. J. Alzheimers Dis. 2019, 67, 779–794. [Google Scholar] [CrossRef] [Green Version]
- Amat-Ur-Rasool, H.; Ahmed, M.; Hasnain, S.; Carter, W. Anti-Cholinesterase Combination Drug Therapy as a Potential Treatment for Alzheimer’s Disease. Brain Sci. 2021, 11, 184. [Google Scholar] [CrossRef]
- Karthikeyan, A.; Joseph, A.; Nair, B.G. Promising bioactive compounds from the marine environment and their potential effects on various diseases. J. Genet. Eng. Biotechnol. 2022, 20, 14. [Google Scholar] [CrossRef]
- Zhang, L.; Hao, J.; Zheng, Y.; Su, R.; Liao, Y.; Gong, X.; Liu, L.; Wang, X. Fucoidan Protects Dopaminergic Neurons by Enhancing the Mitochondrial Function in a Rotenone-induced Rat Model of Parkinson’s Disease. Aging Dis. 2018, 9, 590–604. [Google Scholar] [CrossRef] [Green Version]
- Hao, C.; Wang, W.; Wang, S.; Zhang, L.; Guo, Y. An Overview of the Protective Effects of Chitosan and Acetylated Chitosan Oligosaccharides against Neuronal Disorders. Mar. Drugs 2017, 15, 89. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Xi, Y.; Wang, Q.; Liu, J.; Li, P.; Meng, X.; Liu, K.; Chen, W.; Liu, X.; Liu, Z. Mannan oligosaccharide attenuates cognitive and behavioral disorders in the 5xFAD Alzheimer’s disease mouse model via regulating the gut microbiota-brain axis. Brain Behav. Immun. 2021, 95, 330–343. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.-H.; Lee, N.; Youn, K.; Jo, M.; Kim, H.-R.; Lee, D.-S.; Ho, C.-T.; Jun, M. Dieckol Ameliorates Aβ Production via PI3K/Akt/GSK-3β Regulated APP Processing in SweAPP N2a Cell. Mar. Drugs 2021, 19, 152. [Google Scholar] [CrossRef] [PubMed]
- Mouzat, K.; Chudinova, A.; Polge, A.; Kantar, J.; Camu, W.; Raoul, C.; Lumbroso, S. Regulation of Brain Cholesterol: What Role Do Liver X Receptors Play in Neurodegenerative Diseases? Int. J. Mol. Sci. 2019, 20, 3858. [Google Scholar] [CrossRef]
- Hannan, A.; Dash, R.; Sohag, A.A.M.; Moon, I.S. Deciphering Molecular Mechanism of the Neuropharmacological Action of Fucosterol through Integrated System Pharmacology and In Silico Analysis. Mar. Drugs 2019, 17, 639. [Google Scholar] [CrossRef] [Green Version]
- Torregrosa-Crespo, J.; Montero, Z.; Fuentes, J.L.; García-Galbis, M.R.; Garbayo, I.; Vílchez, C.; Martínez-Espinosa, R.M. Exploring the Valuable Carotenoids for the Large-Scale Production by Marine Microorganisms. Mar. Drugs 2018, 16, 203. [Google Scholar] [CrossRef] [Green Version]
- Sathasivam, R.; Ki, J.-S. A Review of the Biological Activities of Microalgal Carotenoids and Their Potential Use in Healthcare and Cosmetic Industries. Mar. Drugs 2018, 16, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Li, L.; Wang, Z.; Cui, Y.; Tan, X.; Yuan, T.; Liu, Q.; Liu, Z.; Liu, X. Supplementation of lycopene attenuates lipopolysaccharide-induced amyloidogenesis and cognitive impairments via mediating neuroinflammation and oxidative stress. J. Nutr. Biochem. 2018, 56, 16–25. [Google Scholar] [CrossRef]
- Gegunde, S.; Alfonso, A.; Alonso, E.; Alvariño, R.; Botana, L.M. Gracilin-Derivatives as Lead Compounds for Anti-inflammatory Effects. Cell. Mol. Neurobiol. 2020, 40, 603–615. [Google Scholar] [CrossRef]
- Palem, J.R.; Mudit, M.; Hsia, S.-C.V.; El Sayed, K.A. Discovery and preliminary structure-activity relationship of the marine natural product manzamines as herpes simplex virus type-1 inhibitors. Z. Nat. C 2017, 72, 49–54. [Google Scholar] [CrossRef]
- Hu, Y.; Chen, S.; Yang, F.; Dong, S. Marine Indole Alkaloids—Isolation, Structure and Bioactivities. Mar. Drugs 2021, 19, 658. [Google Scholar] [CrossRef]
- Caltagirone, C.; Ferrannini, L.; Marchionni, N.; Nappi, G.; Scapagnini, G.; Trabucchi, M. The potential protective effect of tramiprosate (homotaurine) against Alzheimer’s disease: A review. Aging Clin. Exp. Res. 2012, 24, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Emartorana, A.; Lorenzo, F.E.; Manenti, G.; Esemprini, R.; Koch, G. Homotaurine Induces Measurable Changes of Short Latency Afferent Inhibition in a Group of Mild Cognitive Impairment Individuals. Front. Aging Neurosci. 2014, 6, 254. [Google Scholar] [CrossRef]
- Otero, P.; Alfonso, A.; Vieytes, M.R.; Cabado, A.G.; Vieites, J.M.; Botana, L.M. Effects of environmental regimens on the toxin profile ofAlexandrium ostenfeldii. Environ. Toxicol. Chem. 2010, 29, 301–310. [Google Scholar] [CrossRef]
- Russo, P.; Kisialiou, A.; Lamonaca, P.; Moroni, R.; Prinzi, G.; Fini, M. New Drugs from Marine Organisms in Alzheimer’s Disease. Mar. Drugs 2015, 14, 5. [Google Scholar] [CrossRef] [Green Version]
- Natural Polysaccharide: Modification and Application. Available online: http://qikan.cqvip.com/Qikan/Article/Detail?id=7104613483 (accessed on 26 September 2022).
- Apostolova, E.; Lukova, P.; Baldzhieva, A.; Katsarov, P.; Nikolova, M.; Iliev, I.; Peychev, L.; Trica, B.; Oancea, F.; Delattre, C.; et al. Immunomodulatory and Anti-Inflammatory Effects of Fucoidan: A Review. Polymers 2020, 12, 2338. [Google Scholar] [CrossRef]
- Wang, Y.; Xing, M.; Cao, Q.; Ji, A.; Liang, H.; Song, S. Biological Activities of Fucoidan and the Factors Mediating Its Therapeutic Effects: A Review of Recent Studies. Mar. Drugs 2019, 17, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aleissa, M.S.; Alkahtani, S.; Eldaim, M.A.A.; Ahmed, A.M.; Bungău, S.G.; Almutairi, B.; Bin-Jumah, M.; AlKahtane, A.A.; Alyousif, M.S.; Abdel-Daim, M.M. Fucoidan Ameliorates Oxidative Stress, Inflammation, DNA Damage, and Hepatorenal Injuries in Diabetic Rats Intoxicated with Aflatoxin B1. Oxidative Med. Cell. Longev. 2020, 2020, 9316751. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.; Gao, S.; Terakawa, S. Inhibitory effects of fucoidan on NMDA receptors and l-type Ca2+ channels regulating the Ca2+ responses in rat neurons. Pharm. Biol. 2019, 57, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, C.-H.; Lu, C.-H.; Kuo, Y.-Y.; Lin, G.-B.; Chao, C.-Y. The protective effect of non-invasive low intensity pulsed electric field and fucoidan in preventing oxidative stress-induced motor neuron death via ROCK/Akt pathway. PLoS ONE 2019, 14, e0214100. [Google Scholar] [CrossRef]
- Ji, H.L.; Park, J.E.; Ji, S.H. Fucoidan Stimulates Glucose Uptake via the PI3K/AMPK Pathway and Increases Insulin Sensitivity in 3T3-L1 Adipocytes. Korean Soc. Life Sci. 2021, 31, 1–9. [Google Scholar] [CrossRef]
- Wei, H.; Gao, Z.; Zheng, L.; Zhang, C.; Liu, Z.; Yang, Y.; Teng, H.; Hou, L.; Yin, Y.; Zou, X. Protective Effects of Fucoidan on Aβ25–35 and d-Gal-Induced Neurotoxicity in PC12 Cells and d-Gal-Induced Cognitive Dysfunction in Mice. Mar. Drugs 2017, 15, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Wang, J.; Zhang, Q.; Zhang, H. The effect of different substitute groups and molecular weights of fucoidan on neuroprotective and anticomplement activity. Int. J. Biol. Macromol. 2018, 113, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Tsvetkov, Y.E.; Paulovičová, E.; Paulovičová, L.; Farkaš, P.; Nifantiev, N.E. Synthesis of Biotin-Tagged Chitosan Oligosaccharides and Assessment of Their Immunomodulatory Activity. Front. Chem. 2020, 8, 554732. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.N.R. A Review of Chitin and Chitosan Applications. React. Funct. Polym. 2000, 46, 1–27. [Google Scholar] [CrossRef]
- Jeon, Y.-J.; Park, P.-J.; Kim, S.-K. Antimicrobial Effect of Chitooligosaccharides Produced by Bioreactor. Carbohydr. Polym. 2001, 44, 71–76. [Google Scholar] [CrossRef]
- Liaqat, F.; Eltem, R. Chitooligosaccharides and their biological activities: A comprehensive review. Carbohydr. Polym. 2018, 184, 243–259. [Google Scholar] [CrossRef]
- Khodagholi, F.; Eftekharzadeh, B.; Maghsoudi, N.; Rezaei, P.F. Retracted Article: Chitosan prevents oxidative stress-induced amyloid β formation and cytotoxicity in NT2 neurons: Involvement of transcription factors Nrf2 and NF-κB. Mol. Cell. Biochem. 2009, 337, 39–51. [Google Scholar] [CrossRef]
- Yoon, N.Y.; Ngo, D.-N.; Kim, S.-K. Acetylcholinesterase inhibitory activity of novel chitooligosaccharide derivatives. Carbohydr. Polym. 2009, 78, 869–872. [Google Scholar] [CrossRef]
- Zhu, L.; Li, R.; Jiao, S.; Wei, J.; Yan, Y.; Wang, Z.A.; Li, J.; Du, Y. Blood-Brain Barrier Permeable Chitosan Oligosaccharides Interfere with β-amyloid Aggregation and Alleviate β-Amyloid Protein Mediated Neurotoxicity and Neuroinflammation in a Dose- and Degree of Polymerization-Dependent Manner. Mar. Drugs 2020, 18, 488. [Google Scholar] [CrossRef]
- Xue, Y.; Wang, N.; Zeng, Z.; Huang, J.; Xiang, Z.; Guan, Y.-Q. Neuroprotective effect of chitosan nanoparticle gene delivery system grafted with acteoside (ACT) in Parkinson’s disease models. J. Mater. Sci. Technol. 2020, 43, 197–207. [Google Scholar] [CrossRef]
- Faustino, M.; Durão, J.; Pereira, C.F.; Pintado, M.E.; Carvalho, A.P. Mannans and mannan oligosaccharides (MOS) from Saccharomyces cerevisiae—A sustainable source of functional ingredients. Carbohydr. Polym. 2021, 272, 118467. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Singh, G.; Arya, S.K. Mannans: An overview of properties and application in food products. Int. J. Biol. Macromol. 2018, 119, 79–95. [Google Scholar] [CrossRef] [PubMed]
- Crispi, S.; Filosa, S. Novel perspectives for neurodegeneration prevention: Effects of bioactive polyphenols. Neural Regen. Res. 2021, 16, 1411–1412. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Meng, P.; Zhang, B.; Kang, H.; Wen, H.; Schluesener, H.; Cao, Z.; Zhang, Z. Computer-aided identification of protein targets of four polyphenols in Alzheimer’s disease (AD) and validation in a mouse AD model. J. Biomed. Res. 2019, 33, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Sang, V.T.; Hung, N.D.; Se-Kwon, K. Pharmaceutical properties of marine polyphenols: An overview. Acta Pharm. Sci. 2019, 57, 217. [Google Scholar] [CrossRef] [Green Version]
- Ying, S.-W.; Futter, M.; Rosenblum, K.; Webber, M.J.; Hunt, S.P.; Bliss, T.V.P.; Bramham, C.R. Brain-Derived Neurotrophic Factor Induces Long-Term Potentiation in Intact Adult Hippocampus: Requirement for ERK Activation Coupled to CREB and Upregulation of Arc Synthesis. J. Neurosci. 2002, 22, 1532–1540. [Google Scholar] [CrossRef] [Green Version]
- Um, M.Y.; Lim, D.W.; Son, H.J.; Cho, S.; Lee, C. Phlorotannin-rich fraction from Ishige foliacea brown seaweed prevents the scopolamine-induced memory impairment via regulation of ERK-CREB-BDNF pathway. J. Funct. Foods 2018, 40, 110–116. [Google Scholar] [CrossRef]
- Zambrano, P.; Suwalsky, M.; Jemiola-Rzeminska, M.; Strzalka, K.; Sepúlveda, B.; Gallardo, M.J.; Aguilar, L.F. The acetylcholinesterase (AChE) inhibitor and anti-Alzheimer drug donepezil interacts with human erythrocytes. Biochim. Biophys. Acta Biomembr. 2019, 1861, 1078–1085. [Google Scholar] [CrossRef]
- Hannan, A.; Dash, R.; Haque, N.; Mohibbullah; Sohag, A.; Rahman, A.; Uddin, J.; Alam, M.; Moon, I.S. Neuroprotective Potentials of Marine Algae and Their Bioactive Metabolites: Pharmacological Insights and Therapeutic Advances. Mar. Drugs 2020, 10, 347. [Google Scholar] [CrossRef]
- Minter, M.R.; Taylor, J.M.; Crack, P.J. The contribution of neuroinflammation to amyloid toxicity in Alzheimer’s disease. J. Neurochem. 2016, 136, 457–474. [Google Scholar] [CrossRef] [Green Version]
- Rahman, A.; Dash, R.; Sohag, A.; Alam, M.; Rhim, H.; Ha, H.; Moon, I.; Uddin, J.; Hannan, A. Prospects of Marine Sterols against Pathobiology of Alzheimer’s Disease: Pharmacological Insights and Technological Advances. Mar. Drugs 2021, 19, 167. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Lu, Y.; Lin, X.; Yang, B.; Yang, X.; Liu, Y. Brominated aliphatic hydrocarbons and sterols from the sponge Xestospongia testudinaria with their bioactivities. Chem. Phys. Lipids 2011, 164, 703–706. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.H.; Gan, S.Y.; Tan, S.C.; Gany, S.A.; Ying, T.; Gray, A.I.; Igoli, J.; Chan, E.W.L.; Phang, S.M. Fucosterol inhibits the cholinesterase activities and reduces the release of pro-inflammatory mediators in lipopolysaccharide and amyloid-induced microglial cells. J. Appl. Phycol. 2018, 30, 3261–3270. [Google Scholar] [CrossRef]
- Castro-Silva, E.S.; Bello, M.; Hernández-Rodríguez, M.; Correa-Basurto, J.; Murillo-Álvarez, J.I.; Rosales-Hernández, M.C.; Muñoz-Ochoa, M. In vitro and in silico evaluation of fucosterol from Sargassum horridum as potential human acetylcholinesterase inhibitor. J. Biomol. Struct. Dyn. 2019, 37, 3259–3268. [Google Scholar] [CrossRef] [PubMed]
- Olasehinde, T.A.; Olaniran, A.O.; Okoh, A.I. Macroalgae as a Valuable Source of Naturally Occurring Bioactive Compounds for the Treatment of Alzheimer’s Disease. Mar. Drugs 2019, 17, 609. [Google Scholar] [CrossRef] [Green Version]
- Galasso, C.; Corinaldesi, C.; Sansone, C. Carotenoids from Marine Organisms: Biological Functions and Industrial Applications. Antioxidants 2017, 6, 96. [Google Scholar] [CrossRef] [Green Version]
- Lin, J.; Huang, L.; Yu, J.; Xiang, S.; Wang, J.; Zhang, J.; Yan, X.; Cui, W.; He, S.; Wang, Q. Fucoxanthin, A Marine Carotenoid, Reverses Scopolamine-Induced Cognitive Impairments in Mice and Inhibits Acetylcholinesterase in Vitro. Mar. Drugs 2016, 14, 67. [Google Scholar] [CrossRef] [Green Version]
- Xiang, S.; Liu, F.; Lin, J.; Chen, H.; Huang, C.; Chen, L.; Zhou, Y.; Ye, L.; Zhang, K.; Jin, J.; et al. Fucoxanthin Inhibits β-Amyloid Assembly and Attenuates β-Amyloid Oligomer-Induced Cognitive Impairments. J. Agric. Food Chem. 2017, 65, 4092–4102. [Google Scholar] [CrossRef]
- Manochkumar, J.; Doss, C.G.P.; El-Seedi, H.R.; Efferth, T.; Ramamoorthy, S. The neuroprotective potential of carotenoids in vitro and in vivo. Phytomedicine 2021, 91, 153676. [Google Scholar] [CrossRef]
- Ohgami, K.; Shiratori, K.; Kotake, S.; Nishida, T.; Mizuki, N.; Yazawa, K.; Ohno, S. Effects of Astaxanthin on Lipopolysaccharide-Induced Inflammation In Vitro and In Vivo. Investig. Opthalmol. Vis. Sci. 2003, 44, 2694–2701. [Google Scholar] [CrossRef] [Green Version]
- Bonito, M.C.; Cicala, C.; Marcotullio, M.C.; Maione, F.; Mascolo, N. Biological Activity of Bicyclic and Tricyclic Diterpenoids from Salvia Species of Immediate Pharmacological and Pharmaceutical Interest. Nat. Prod. Commun. 2011, 6, 1205–1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leirós, M.; Alonso, E.; Rateb, M.E.; Houssen, W.E.; Ebel, R.; Jaspars, M.; Alfonso, A.; Botana, L.M. Gracilins: Spongionella-derived promising compounds for Alzheimer disease. Neuropharmacology 2015, 93, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.-Y.; Choi, H. Natural products from marine organisms with neuroprotective activity in the experimental models of Alzheimer’s disease, Parkinson’s disease and ischemic brain stroke: Their molecular targets and action mechanisms. Arch. Pharmacal Res. 2015, 38, 139–170. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, Q.; Zheng, D.-K.; Zhang, D.; Xie, Z.; Hu, J.-W.; Xie, X.-H.; Li, J.; Jiang, S.-P. Abietane diterpenoids with neuroprotective activities from Phlegmariurus carinatus. Nat. Prod. Res. 2022, 36, 6006–6011. [Google Scholar] [CrossRef]
- Abbasov, M.E.; Alvariño, R.; Chaheine, C.M.; Alonso, E.; Sánchez, J.A.; Conner, M.L.; Alfonso, A.; Jaspars, M.; Botana, L.M.; Romo, D. Simplified immunosuppressive and neuroprotective agents based on gracilin A. Nat. Chem. 2019, 11, 342–350. [Google Scholar] [CrossRef]
- Alvariño, R.; Alonso, E.; Abbasov, M.E.; Chaheine, C.M.; Conner, M.L.; Romo, D.; Alfonso, A.; Botana, L.M. Gracilin A Derivatives Target Early Events in Alzheimer’s Disease: In Vitro Effects on Neuroinflammation and Oxidative Stress. ACS Chem. Neurosci. 2019, 10, 4102–4111. [Google Scholar] [CrossRef]
- Kabir, T.; Uddin, S.; Jeandet, P.; Emran, T.; Mitra, S.; Albadrani, G.; Sayed, A.; Abdel-Daim, M.; Simal-Gandara, J. Anti-Alzheimer’s Molecules Derived from Marine Life: Understanding Molecular Mechanisms and Therapeutic Potential. Mar. Drugs 2021, 19, 251. [Google Scholar] [CrossRef]
- Li, H.; Liu, X.; Li, X.; Hu, Z.; Wang, L. Novel Harziane Diterpenes from Deep-Sea Sediment Fungus Trichoderma sp. SCSIOW21 and Their Potential Anti-Inflammatory Effects. Mar. Drugs 2021, 19, 689. [Google Scholar] [CrossRef]
- Althagbi, H.I.; Alarif, W.M.; Al-Footy, K.O.; Abdel-Lateff, A. Marine-Derived Macrocyclic Alkaloids (MDMAs): Chemical and Biological Diversity. Mar. Drugs 2020, 18, 368. [Google Scholar] [CrossRef]
- Karan, D.; Dubey, S.; Pirisi, L.; Nagel, A.; Pina, I.; Choo, Y.-M.; Hamann, M.T. The Marine Natural Product Manzamine A Inhibits Cervical Cancer by Targeting the SIX1 Protein. J. Nat. Prod. 2020, 83, 286–295. [Google Scholar] [CrossRef]
- Yousaf, M.; Hammond, N.L.; Peng, J.; Wahyuono, S.; McIntosh, K.A.; Charman, W.N.; Mayer, A.A.M.S.; Hamann, M.T. New Manzamine Alkaloids from an Indo-Pacific Sponge. Pharmacokinetics, Oral Availability, and the Significant Activity of Several Manzamines against HIV-I, AIDS Opportunistic Infections, and Inflammatory Diseases. J. Med. Chem. 2004, 47, 3512–3517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, K.V.; Donia, M.S.; Peng, J.; Garcia-Palomero, E.; Alonso, D.; Martinez, A.; Medina, M.; Franzblau, S.G.; Tekwani, B.L.; Khan, S.I.; et al. Manzamine B and E and Ircinal A Related Alkaloids from an Indonesian Acanthostrongylophora Sponge and Their Activity against Infectious, Tropical Parasitic, and Alzheimer’s Diseases. J. Nat. Prod. 2006, 69, 1034–1040. [Google Scholar] [CrossRef] [Green Version]
- Lima, E.; Medeiros, J. Marine Organisms as Alkaloid Biosynthesizers of Potential Anti-Alzheimer Agents. Mar. Drugs 2022, 20, 75. [Google Scholar] [CrossRef] [PubMed]
- Miguel-Gordo, M.; Gegunde, S.; Calabro, K.; Jennings, L.K.; Alfonso, A.; Genta-Jouve, G.; Vacelet, J.; Botana, L.M.; Thomas, O.P. Bromotryptamine and Bromotyramine Derivatives from the Tropical Southwestern Pacific Sponge Narrabeena nigra. Mar. Drugs 2019, 17, 319. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Shi, J.-S.; Li, D.-D.; Zheng, C.-Q. Potential neuroprotection by Dendrobium nobile Lindl alkaloid in Alzheimer’s disease models. Neural Regen. Res. 2022, 17, 972–977. [Google Scholar] [CrossRef] [PubMed]
- Aisen, P.S.; Saumier, D.; Briand, R.; Laurin, J.; Gervais, F.; Tremblay, P.; Garceau, D. A Phase II study targeting amyloid-with 3APS in mild-to-moderate Alzheimer disease. Neurology 2006, 67, 1757–1763. [Google Scholar] [CrossRef] [PubMed]
- Jakaria, M.; Azam, S.; Haque, M.E.; Jo, S.-H.; Uddin, M.S.; Kim, I.-S.; Choi, D.-K. Taurine and its analogs in neurological disorders: Focus on therapeutic potential and molecular mechanisms. Redox Biol. 2019, 24, 101223. [Google Scholar] [CrossRef]
- Shivaraj, M.C.; Marcy, G.; Low, G.; Ryu, J.R.; Zhao, X.; Rosales, F.J.; Goh, E.L.K. Taurine Induces Proliferation of Neural Stem Cells and Synapse Development in the Developing Mouse Brain. PLoS ONE 2012, 7, e42935. [Google Scholar] [CrossRef]
- Mehdinia, A.; Fumani, N.S.; Kayyal, T.B.; Ghaderiardakani, F. Homotaurine of Marine Macroalgae of the Persian Gulf as a Potential Treatment Agent for Alzheimer. Mar. Sci. 2018, 9, 1–8. [Google Scholar]
- Chekan, J.R.; Fallon, T.R.; Moore, B.S. Biosynthesis of marine toxins. Curr. Opin. Chem. Biol. 2020, 59, 119–129. [Google Scholar] [CrossRef]
- Kem, W.R.; Andrud, K.; Bruno, G.; Xing, H.; Soti, F.; Talley, T.T.; Taylor, P. Interactions of Nereistoxin and Its Analogs with Vertebrate Nicotinic Acetylcholine Receptors and Molluscan ACh Binding Proteins. Mar. Drugs 2022, 20, 49. [Google Scholar] [CrossRef]
- Touzet, N.; Franco, J.M.; Raine, R. Morphogenetic diversity and biotoxin composition of Alexandrium (Dinophyceae) in Irish coastal waters. Harmful Algae 2008, 7, 782–797. [Google Scholar] [CrossRef]
- Hu, T.; Burton, I.W.; Cembella, A.D.; Curtis, J.M.; Quilliam, M.A.; Walter, J.A.; Wright, J.L.C. Characterization of Spirolides A, C, and 13-Desmethyl C, New Marine Toxins Isolated from Toxic Plankton and Contaminated Shellfish. J. Nat. Prod. 2001, 64, 308–312. [Google Scholar] [CrossRef]
- Boente-Juncal, A.; Méndez, A.G.; Vale, C.; Vieytes, M.R.; Botana, L.M. In Vitro Effects of Chronic Spirolide Treatment on Human Neuronal Stem Cell Differentiation and Cholinergic System Development. ACS Chem. Neurosci. 2018, 9, 1441–1452. [Google Scholar] [CrossRef]
- Alonso, E.; Vale, C.; Vieytes, M.R.; Laferla, F.M.; Giménez-Llort, L.; Botana, L.M. 13-Desmethyl spirolide-C is neuroprotective and reduces intracellular Aβ and hyperphosphorylated tau in vitro. Neurochem. Int. 2011, 59, 1056–1065. [Google Scholar] [CrossRef]
- Botelho, M.J.; Milinovic, J.; Bandarra, N.M.; Vale, C. Alzheimer’s Disease and Toxins Produced by Marine Dinoflagellates: An Issue to Explore. Mar. Drugs 2022, 20, 253. [Google Scholar] [CrossRef]
- Mahmood, N.A.; Carmichael, W.W. Anatoxin-a(s), an anticholinesterase from the cyanobacterium Anabaena flos-aquae NRC-525-17. Toxicon 1987, 25, 1221–1227. [Google Scholar] [CrossRef]
- Castaneda, A.; Ferraz, R.; Vieira, M.; Cardoso, I.; Vasconcelos, V.; Martins, R. Bridging Cyanobacteria to Neurodegenerative Diseases: A New Potential Source of Bioactive Compounds against Alzheimer’s Disease. Mar. Drugs 2021, 19, 343. [Google Scholar] [CrossRef]
- Aldholmi, M.; Marchand, P.; Ourliac-Garnier, I.; Le Pape, P.; Ganesan, A. A Decade of Antifungal Leads from Natural Products: 2010–2019. Pharmaceuticals 2019, 12, 182. [Google Scholar] [CrossRef] [Green Version]
- Alvariño, R.; Alonso, E.; Lacret, R.; Oves-Costales, D.; Genilloud, O.; Reyes, F.; Alfonso, A.; Botana, L.M. Caniferolide A, a Macrolide from Streptomyces caniferus, Attenuates Neuroinflammation, Oxidative Stress, Amyloid-Beta, and Tau Pathology in Vitro. Mol. Pharm. 2019, 16, 1456–1466. [Google Scholar] [CrossRef]
- Sun, M.-K.; Nelson, T.J.; Alkon, D.L. Towards universal therapeutics for memory disorders. Trends Pharmacol. Sci. 2015, 36, 384–394. [Google Scholar] [CrossRef]
- Hongpaisan, J.; Sun, M.-K.; Alkon, D.L. PKC Activation Prevents Synaptic Loss, A Elevation, and Cognitive Deficits in Alzheimer’s Disease Transgenic Mice. J. Neurosci. 2011, 31, 630–643. [Google Scholar] [CrossRef] [Green Version]
- Neurotrope Announces Positive Top-Line Results From Its Phase 2a Study of Bryostatin-1 in Alzheimer’s Disease. Available online: https://www.prnewswire.com/news-releases/neurotrope-announces-positive-top-line-results-from-its-phase-2a-study-of-bryostatin-1-in-alzheimers-disease-300038367.html (accessed on 3 November 2022).
- Farlow, M.R.; Thompson, R.E.; Wei, L.-J.; Tuchman, A.J.; Grenier, E.; Crockford, D.; Wilke, S.; Benison, J.; Alkon, D.L. A Randomized, Double-Blind, Placebo-Controlled, Phase II Study Assessing Safety, Tolerability, and Efficacy of Bryostatin in the Treatment of Moderately Severe to Severe Alzheimer’s Disease. J. Alzheimers Dis. 2019, 67, 555–570. [Google Scholar] [CrossRef] [Green Version]
- Weidner, W.S.; Barbarino, P. P4–443: The State Of The Art Of Dementia Research: New Frontiers. Alzheimers Dement. 2019, 15, P1473. [Google Scholar] [CrossRef]
- Cacabelos, R. How plausible is an Alzheimer’s disease vaccine? Expert Opin. Drug Discov. 2019, 15, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Reich, N.; Hölscher, C. Acylated Ghrelin as a Multi-Targeted Therapy for Alzheimer’s and Parkinson’s Disease. Front. Neurosci. 2020, 14, 614828. [Google Scholar] [CrossRef]
- Cabrera-Pardo, J.R.; Fuentealba, J.; Gavilán, J.; Cajas, D.; Becerra, J.; Napiórkowska, M. Exploring the Multi–Target Neuroprotective Chemical Space of Benzofuran Scaffolds: A New Strategy in Drug Development for Alzheimer’s Disease. Front. Pharmacol. 2020, 10, 1679. [Google Scholar] [CrossRef]
- Bolea, I.; Gella, A.; Unzeta, M. Propargylamine-derived multitarget-directed ligands: Fighting Alzheimer’s disease with monoamine oxidase inhibitors. J. Neural Transm. 2013, 120, 893–902. [Google Scholar] [CrossRef]
- Xuan, Z.; Gu, X.; Yan, S.; Xie, Y.; Zhou, Y.; Zhang, H.; Jin, H.; Hu, S.; Mak, M.S.H.; Zhou, D.; et al. Dimeric Tacrine(10)-hupyridone as a Multitarget-Directed Ligand To Treat Alzheimer’s Disease. ACS Chem. Neurosci. 2021, 12, 2462–2477. [Google Scholar] [CrossRef]
- Anisha, G.S.; Padmakumari, S.; Patel, A.K.; Pandey, A.; Singhania, R.R. Fucoidan from Marine Macroalgae: Biological Actions and Applications in Regenerative Medicine, Drug Delivery Systems and Food Industry. Bioengineering 2022, 9, 472. [Google Scholar] [CrossRef]
- Vargas, M.; Gonzalez-Martinez, C. Recent Patents on Food Applications of Chitosan. Recent Pat. Food Nutr. Agric. 2010, 2, 121–128. [Google Scholar] [CrossRef]
- Wang, S.-H.; Huang, C.-Y.; Chen, C.-Y.; Chang, C.-C.; Huang, C.-Y.; Dong, C.-D.; Chang, J.-S. Isolation and purification of brown algae fucoidan from Sargassum siliquosum and the analysis of anti-lipogenesis activity. Biochem. Eng. J. 2021, 165, 107798. [Google Scholar] [CrossRef]
- Lourenço-Lopes, C.; Garcia-Oliveira, P.; Carpena, M.; Fraga-Corral, M.; Jimenez-Lopez, C.; Pereira, A.G.; Prieto, M.A.; Simal-Gandara, J. Scientific Approaches on Extraction, Purification and Stability for the Commercialization of Fucoxanthin Recovered from Brown Algae. Foods 2020, 9, 1113. [Google Scholar] [CrossRef]
- Priyadarsani, S.; Patel, A.S.; Kar, A.; Dash, S. Process optimization for the supercritical carbondioxide extraction of lycopene from ripe grapefruit (Citrus paradisi) endocarp. Sci. Rep. 2021, 11, 10273. [Google Scholar] [CrossRef]
- Obluchinskaya, E.; Pozharitskaya, O.; Zakharova, L.; Daurtseva, A.; Flisyuk, E.; Shikov, A. Efficacy of Natural Deep Eutectic Solvents for Extraction of Hydrophilic and Lipophilic Compounds from Fucus vesiculosus. Molecules 2021, 26, 4198. [Google Scholar] [CrossRef]
- Trombino, F.C.A.C.R.S.; Curcio, F.; Cassano, R. Nano- and Micro-Technologies Applied to Food Nutritional Ingredients. Curr. Drug Deliv. 2021, 18, 670–678. [Google Scholar] [CrossRef]
- ElKhoury, K.; Sanchez-Gonzalez, L.; Lavrador, P.; Almeida, R.; Gaspar, V.; Kahn, C.; Cleymand, F.; Arab-Tehrany, E.; Mano, J.F. Gelatin Methacryloyl (GelMA) Nanocomposite Hydrogels Embedding Bioactive Naringin Liposomes. Polymers 2020, 12, 2944. [Google Scholar] [CrossRef]
- Zhang, D.; Zhong, D.; Ouyang, J.; He, J.; Qi, Y.; Chen, W.; Zhang, X.; Tao, W.; Zhou, M. Microalgae-based oral microcarriers for gut microbiota homeostasis and intestinal protection in cancer radiotherapy. Nat. Commun. 2022, 13, 1413. [Google Scholar] [CrossRef]
- Zhong, D.; Zhang, D.; Chen, W.; He, J.; Ren, C.; Zhang, X.; Kong, N.; Tao, W.; Zhou, M. Orally deliverable strategy based on microalgal biomass for intestinal disease treatment. Sci. Adv. 2021, 7, eabi9265. [Google Scholar] [CrossRef]
- Jeromin, A.; Bowser, R. Biomarkers in Neurodegenerative Diseases. Nat. Med. 2017, 15, 491–528. [Google Scholar] [CrossRef]
- Ehrenberg, A.J.; Khatun, A.; Coomans, E.; Betts, M.J.; Capraro, F.; Thijssen, E.H.; Senkevich, K.; Bharucha, T.; Jafarpour, M.; Young, P.N.E.; et al. Relevance of biomarkers across different neurodegenerative diseases. Alzheimers Res. Ther. 2020, 12, 56. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, I.; Rauter, A.P.; Bandarra, N.M. Marine Sources of DHA-Rich Phospholipids with Anti-Alzheimer Effect. Mar. Drugs 2022, 20, 662. [Google Scholar] [CrossRef] [PubMed]
- da Silva, A.P.G. Fighting coronaviruses with natural polyphenols. Biocatal. Agric. Biotechnol. 2021, 37, 102179. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drug’s Name | Chemical Structures | Pharmacological Mechanism | Ref. |
|---|---|---|---|
| Tacrine |  | Inhibition of cholinesterase (both AChE and BChE) | [40] |
| Donepezile HCl |  | Inhibition of AChE | [41] |
| Rivastigmine |  | Reversible double inhibitors of AChE and BChE | [42] |
| Galantamine |  | Weak competitive and reversible cholinesterase inhibitors | [38] |
| Memantine |  | Antagonist of the NMDAR | [37] |
| sodium oligomannate | n = 1~9; m = 0, 1, 2; m’ = 0,1 | Directly combine with Aβ to reduce Aβ in brain deposition | [43] |
| Family | Compound | Origin | Pharmacological Activity | Ref. |
|---|---|---|---|---|
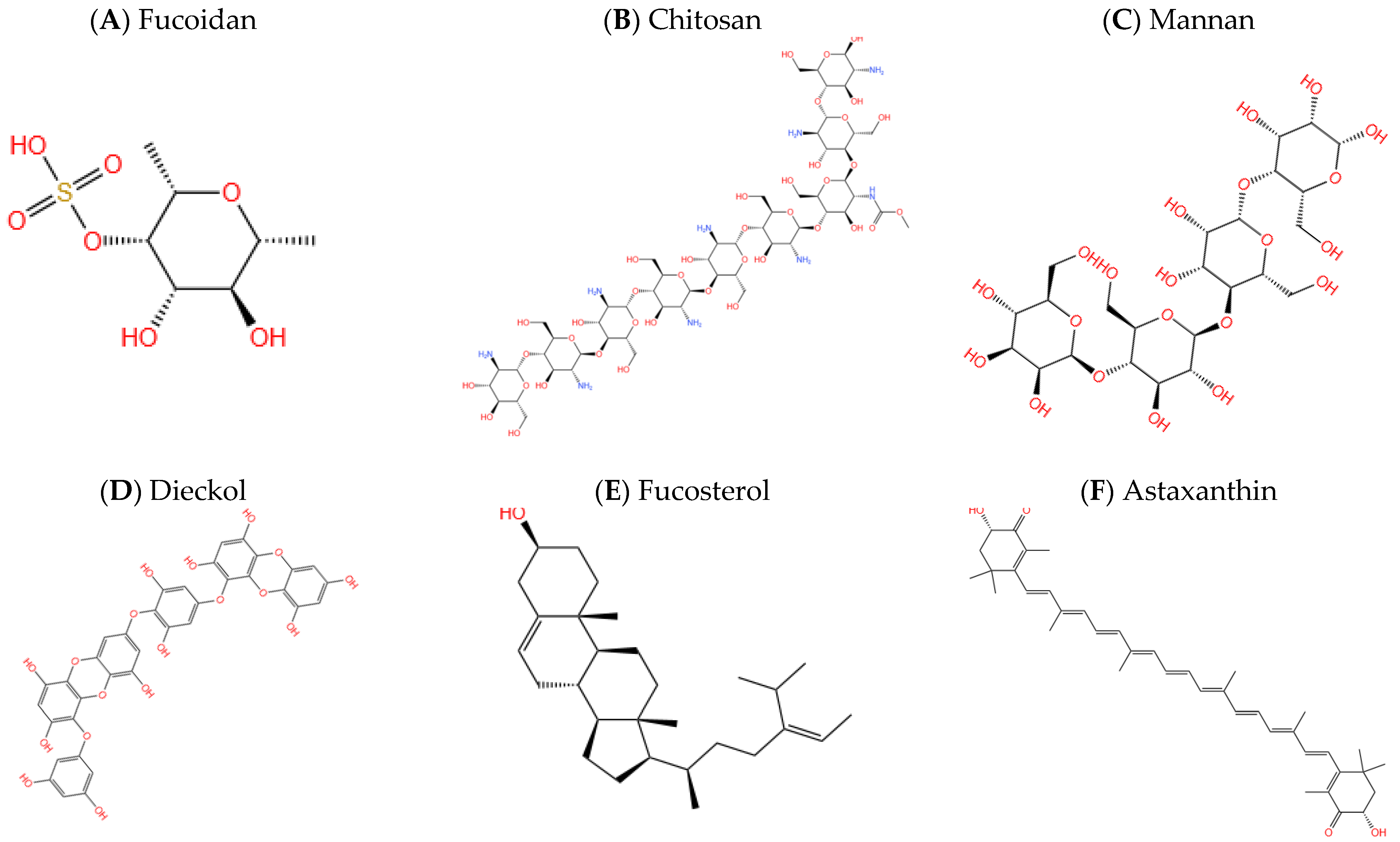
| Polysaccharide | Fucoidan | Brown seaweeds | Block caspase-9 and caspase-3 enzymes. | [57] |
| Chitosan | Crustaceans | Inhibition of the enzyme acetylcholinesterase. | [58] | |
| Mannan | Codium fragile | Inhibition of the enzyme β-secretase (Bace1) | [59] | |
| Polyphenol | Dieckol | Brown seaweeds (Ecklonia cava, Ecklonia stolonifera) | Inhibition of the enzymes acetylcholinesterase and Butyrylcholinesterase | [60] |
| Sterol | Fucosterol | brown alarge (Panida australis) | Anti-inflammatory and anti-BACE1 | [61,62] |
| Carotenoid | Fucoxanthin | brown algae (Sargassum siliquastrum) | Anti-inflammatory and antioxidant | [63] |
| Astaxanthin | green algae (Haematococcus pluvialis) | Decrease the production of NF-κB transcription factors and inflammatory cytokines | [64] | |
| Lycopene | Red seaweeds | Anti-inflammatory and antioxidant | [65] | |
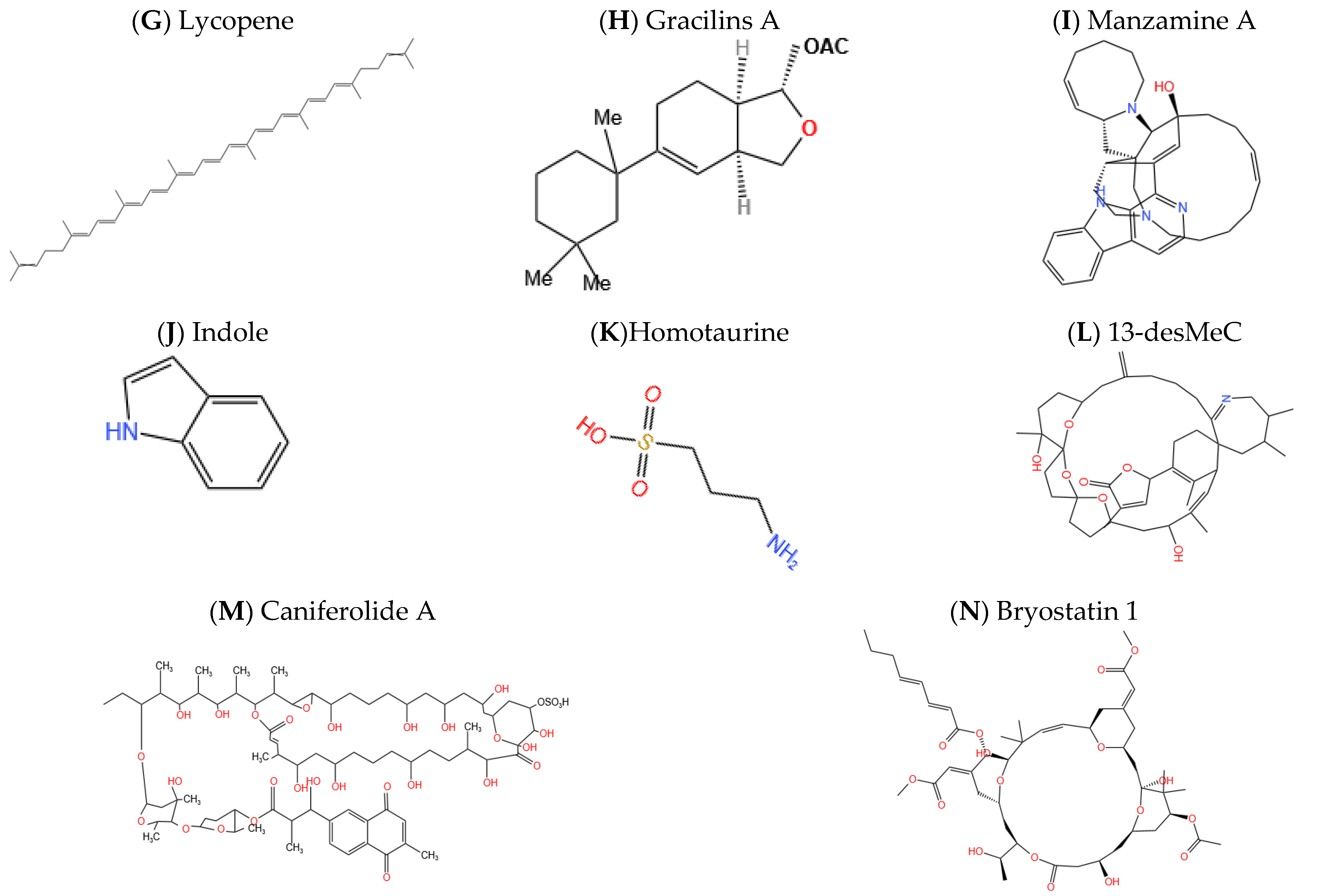
| Diterpenoid | Gracilins | Marine sponges (Spongionella gracilis) | Inhibition of the enzyme β-secretase or BACE-1.Anti-inflammatory and antioxidant properties Reduction in hyperphosphorylation of tau protein | [66] |
| Alkaloid | Manzamine | Marine sponges (Acanthostrongylophora) | GSK-3 inhibition and reduction in hyperphosphorylation of tau protein | [67] |
| Indole | Streptomyces sp. | Inhibition Aβ plaque production by activating the nuclear factor Nrf2 | [68] | |
| Amino sulfonate | Homotaurine | Red seaweeds | Aβ lowering and prevention of the formation of a toxic soluble amyloid oligomer | [69,70] |
| Marine Toxin | Spirolides | Alexandrium ostenfeldii | Acetylcholinesterase inhibition, and restraint the formation of amyloid-β | [71] |
| Macrolide | Caniferolide A | Phylum Actinobacteria | Anti-inflammatory and antioxidant action Blockade of the BACE-1 enzyme. | [72] |
| Parameters 1 | Fucoidan | Mannan | Fucosterol | Astaxanthin | Lycopene | Manzamine A | Indole | Homotaurin | 13-desMeC |
|---|---|---|---|---|---|---|---|---|---|
| MW (130~725) | 242.05 | 666.22 | 412.37 | 596.39 | 536.44 | 548.35 | 117.06 | 139.03 | 691.44 |
| LogS (−4~0.5) | −0.042 | 0.762 | −6.887 | −7.226 | −7.642 | −3.86 | −2.151 | −0.14 | −5.162 |
| LogP (0~3) | −2.003 | −4.868 | 7.447 | 8.045 | 11.072 | 5.459 | 2.292 | −2.745 | 5.652 |
| Pgp-inh | 0 | 0 | 0.679 | 1 | 0.998 | 0.999 | 0.001 | 0.001 | 0.998 |
| Pgp-sub | 0.006 | 0.894 | 0.001 | 0.011 | 0.758 | 0.056 | 0.012 | 0.002 | 0.724 |
| HIA | 0.927 | 1 | 0.004 | 0.019 | 0.02 | 0.034 | 0.005 | 0.925 | 0.027 |
| F (30%) | 0.85 | 1 | 0.224 | 0.001 | 0.113 | 0.003 | 0.468 | 0.835 | 0.534 |
| Caco-2 | −5.53 | −6.294 | −4.624 | −5.196 | −5.708 | −5.131 | −4.259 | −5.902 | −4.813 |
| BBB | 0.678 | 0.45 | 0.818 | 0.001 | 0.001 | 0.99 | 0.737 | 0.936 | 0.25 |
| PPB | 18.86% | 5.85% | 98.64% | 101.00% | 99.44% | 96.16% | 86.32% | 10.54% | 97.60% |
| Fu | 72.18% | 54.83% | 1.78% | 2.56% | 5.42% | 3.27% | 16.50% | 87.62% | 1.54% |
| CYP1A2-inh | 0.003 | 0 | 0.058 | 0.015 | 0.266 | 0.28 | 0.975 | 0.005 | 0.005 |
| CYP1A2-sub | 0.171 | 0.002 | 0.436 | 0.156 | 0.521 | 0.786 | 0.805 | 0.237 | 0.817 |
| CL | 3.024 | 0.28 | 13.304 | 0.719 | −0.286 | 7.851 | 11.189 | 3.782 | 18.565 |
| T1/2 | 0.302 | 0.543 | 0.016 | 0.067 | 0.137 | 0.008 | 0.794 | 0.525 | 0.015 |
| hERG | 0.029 | 0.025 | 0.011 | 0.235 | 0.852 | 0.863 | 0.029 | 0.035 | 0.679 |
| Ames | 0.16 | 0.072 | 0.023 | 0.369 | 0.393 | 0.154 | 0.311 | 0.056 | 0.02 |
| ROA | 0.943 | 0.089 | 0.022 | 0.111 | 0.218 | 0.891 | 0.844 | 0.346 | 0.993 |
| FDAMDD | 0.027 | 0 | 0.638 | 0.974 | 0.948 | 0.932 | 0.219 | 0.014 | 0.942 |
| BCF | 0.444 | 0.105 | 3.317 | 1.465 | 2.187 | 1.081 | 0.934 | 0.212 | 1.755 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, D.; Jin, Y.; Hou, X.; Zhu, Y.; Chen, D.; Tai, J.; Chen, Q.; Shi, C.; Ye, J.; Wu, M.; et al. Application of Marine Natural Products against Alzheimer’s Disease: Past, Present and Future. Mar. Drugs 2023, 21, 43. https://doi.org/10.3390/md21010043
Hu D, Jin Y, Hou X, Zhu Y, Chen D, Tai J, Chen Q, Shi C, Ye J, Wu M, et al. Application of Marine Natural Products against Alzheimer’s Disease: Past, Present and Future. Marine Drugs. 2023; 21(1):43. https://doi.org/10.3390/md21010043
Chicago/Turabian StyleHu, Di, Yating Jin, Xiangqi Hou, Yinlong Zhu, Danting Chen, Jingjing Tai, Qianqian Chen, Cui Shi, Jing Ye, Mengxu Wu, and et al. 2023. "Application of Marine Natural Products against Alzheimer’s Disease: Past, Present and Future" Marine Drugs 21, no. 1: 43. https://doi.org/10.3390/md21010043
APA StyleHu, D., Jin, Y., Hou, X., Zhu, Y., Chen, D., Tai, J., Chen, Q., Shi, C., Ye, J., Wu, M., Zhang, H., & Lu, Y. (2023). Application of Marine Natural Products against Alzheimer’s Disease: Past, Present and Future. Marine Drugs, 21(1), 43. https://doi.org/10.3390/md21010043