
Transcriptome Sequencing of the Diatom Asterionellopsis thurstonii and In Silico Identification of Enzymes Potentially Involved in the Synthesis of Bioactive Molecules

 , ,
, ,  ,
, 

Abstract
:1. Introduction
2. Results
2.1. Genetic Characterization of the Strains
2.2. Transcriptome Assembly and Funtional Annotation
2.3. Differential Expression Analysis
2.3.1. Differential Expression Analysis of Strain A4
2.3.2. Differential Expression Analysis of Strain FE355
2.4. DEGs Involved in Anti-Cancer and Immunomodulatory Responses
3. Discussion
4. Materials and Methods
4.1. Cell Culturing and Harvesting
4.2. DNA Extraction and Genetic Characterization of the Strains
4.3. RNA Extraction
4.4. Sequencing, Assembly, Annotation and Differential Expression Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Saide, A.; Martínez, K.A.; Ianora, A.; Lauritano, C. Unlocking the Health Potential of Microalgae as Sustainable Sources of Bioactive Compounds. Int. J. Mol. Sci. 2021, 22, 4383. [Google Scholar] [CrossRef] [PubMed]
- Hargraves, P.E. Identifying Marine Phytoplankton. Eos Trans. AGU 1998, 79, 99. [Google Scholar] [CrossRef] [Green Version]
- Martínez, K.A.; Saide, A.; Crespo, G.; Martín, J.; Romano, G.; Reyes, F.; Lauritano, C.; Ianora, A. Promising Antiproliferative Compound From the Green Microalga Dunaliella Tertiolecta Against Human Cancer Cells. Front. Mar. Sci. 2022, 9, 778108. [Google Scholar] [CrossRef]
- Ingebrigtsen, R.A.; Hansen, E.; Andersen, J.H.; Eilertsen, H.C. Light and Temperature Effects on Bioactivity in Diatoms. J. Appl. Phycol. 2016, 28, 939–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuño, K.; Villarruel-López, A.; Puebla-Pérez, A.M.; Romero-Velarde, E.; Puebla-Mora, A.G.; Ascencio, F. Effects of the Marine Microalgae Isochrysis galbana and Nannochloropsis oculata in Diabetic Rats. J. Funct. Foods 2013, 5, 106–115. [Google Scholar] [CrossRef]
- Montuori, E.; de Pascale, D.; Lauritano, C. Recent Discoveries on Marine Organism Immunomodulatory Activities. Mar. Drugs 2022, 20, 422. [Google Scholar] [CrossRef] [PubMed]
- Elagoz, A.M.; Ambrosino, L.; Lauritano, C. De Novo Transcriptome of the Diatom Cylindrotheca Closterium Identifies Genes Involved in the Metabolism of Anti-Inflammatory Compounds. Sci. Rep. 2020, 10, 4138. [Google Scholar] [CrossRef] [Green Version]
- Vingiani, G.M.; Štālberga, D.; De Luca, P.; Ianora, A.; De Luca, D.; Lauritano, C. De Novo Transcriptome of the Non-Saxitoxin Producing Alexandrium Tamutum Reveals New Insights on Harmful Dinoflagellates. Mar. Drugs 2020, 18, 386. [Google Scholar] [CrossRef]
- Riccio, G.; Martinez, K.A.; Ianora, A.; Lauritano, C. De Novo Transcriptome of the Flagellate Isochrysis galbana Identifies Genes Involved in the Metabolism of Antiproliferative Metabolites. Biology 2022, 11, 771. [Google Scholar] [CrossRef]
- Lauritano, C.; Ferrante, M.I.; Rogato, A. Marine Natural Products from Microalgae: An -Omics Overview. Mar. Drugs 2019, 17, 269. [Google Scholar] [CrossRef]
- Sasso, S.; Pohnert, G.; Lohr, M.; Mittag, M.; Hertweck, C. Microalgae in the Postgenomic Era: A Blooming Reservoir for New Natural Products. FEMS Microbiol. Rev. 2012, 36, 761–785. [Google Scholar] [CrossRef] [PubMed]
- Osuna-Cruz, C.M.; Bilcke, G.; Vancaester, E.; De Decker, S.; Bones, A.M.; Winge, P.; Poulsen, N.; Bulankova, P.; Verhelst, B.; Audoor, S.; et al. The Seminavis Robusta Genome Provides Insights into the Evolutionary Adaptations of Benthic Diatoms. Nat. Commun. 2020, 11, 3320. [Google Scholar] [CrossRef] [PubMed]
- Kamikawa, R.; Mochizuki, T.; Sakamoto, M.; Tanizawa, Y.; Nakayama, T.; Onuma, R.; Cenci, U.; Moog, D.; Speak, S.; Sarkozi, K.; et al. Genome Evolution of a Nonparasitic Secondary Heterotroph, the Diatom Nitzschia putrida. Sci. Adv. 2022, 8, eabi5075. [Google Scholar] [CrossRef] [PubMed]
- Ribalet, F.; Wichard, T.; Pohnert, G.; Ianora, A.; Miralto, A.; Casotti, R. Age and Nutrient Limitation Enhance Polyunsaturated Aldehyde Production in Marine Diatoms. Phytochemistry 2007, 68, 2059–2067. [Google Scholar] [CrossRef]
- Miralto, A.; Barone, G.; Romano, G.; Poulet, S.A.; Ianora, A.; Russo, G.L.; Buttino, I.; Mazzarella, G.; Laabir, M.; Cabrini, M.; et al. The Insidious Effect of Diatoms on Copepod Reproduction. Nature 1999, 402, 173–176. [Google Scholar] [CrossRef]
- Romano, S.; Jackson, S.; Patry, S.; Dobson, A. Extending the “One Strain Many Compounds” (OSMAC) Principle to Marine Microorganisms. Mar. Drugs 2018, 16, 244. [Google Scholar] [CrossRef] [Green Version]
- Lauritano, C.; Andersen, J.H.; Hansen, E.; Albrigtsen, M.; Escalera, L.; Esposito, F.; Helland, K.; Hanssen, K.Ø.; Romano, G.; Ianora, A. Bioactivity Screening of Microalgae for Antioxidant, Anti-Inflammatory, Anticancer, Anti-Diabetes, and Antibacterial Activities. Front. Mar. Sci. 2016, 3, 68. [Google Scholar] [CrossRef] [Green Version]
- Franco, A.d.O.d.R.; They, N.H.; Canani, L.G.d.C.; Maggioni, R.; Odebrecht, C. Asterionellopsis tropicalis (Bacillariophyceae): A New Tropical Species Found in Diatom Accumulations. J. Phycol. 2016, 52, 888–895. [Google Scholar] [CrossRef]
- Tomas, C.R.; Hasle, G.R. (Eds.) Identifying Marine Phytoplankton; Academic Press: San Diego, CA, USA, 1997; ISBN 978-0-12-693018-4. [Google Scholar]
- Srichandan, S.; Baliarsingh, S.K.; Prakash, S.; Lotliker, A.A.; Parida, C.; Sahu, K.C. Seasonal Dynamics of Phytoplankton in Response to Environmental Variables in Contrasting Coastal Ecosystems. Environ. Sci Pollut. Res. 2019, 26, 12025–12041. [Google Scholar] [CrossRef]
- Kaczmarska, I.; Mather, L.; Luddington, I.A.; Muise, F.; Ehrman, J.M. Cryptic Diversity in a Cosmopolitan Diatom Known as Asterionellopsis Glacialis (Fragilariaceae): Implications for Ecology, Biogeography, and Taxonomy. Am. J. Bot. 2014, 101, 267–286. [Google Scholar] [CrossRef]
- Keeling, P.J.; Burki, F.; Wilcox, H.M.; Allam, B.; Allen, E.E.; Amaral-Zettler, L.A.; Armbrust, E.V.; Archibald, J.M.; Bharti, A.K.; Bell, C.J.; et al. The Marine Microbial Eukaryote Transcriptome Sequencing Project (MMETSP): Illuminating the Functional Diversity of Eukaryotic Life in the Oceans through Transcriptome Sequencing. PLoS Biol. 2014, 12, e1001889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez Andrade, K.; Lauritano, C.; Romano, G.; Ianora, A. Marine Microalgae with Anti-Cancer Properties. Mar. Drugs 2018, 16, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wichard, T.; Poulet, S.A.; Halsband-Lenk, C.; Albaina, A.; Harris, R.; Liu, D.; Pohnert, G. Survey of the Chemical Defence Potential of Diatoms: Screening of Fifty Species for α,β,γ,δ-Unsaturated Aldehydes. J. Chem. Ecol. 2005, 31, 949–958. [Google Scholar] [CrossRef] [PubMed]
- Ianora, A.; Bentley, M.G.; Caldwell, G.S.; Casotti, R.; Cembella, A.D.; Engström-Öst, J.; Halsband, C.; Sonnenschein, E.; Legrand, C.; Llewellyn, C.A.; et al. The Relevance of Marine Chemical Ecology to Plankton and Ecosystem Function: An Emerging Field. Mar. Drugs 2011, 9, 1625–1648. [Google Scholar] [CrossRef] [Green Version]
- Lauritano, C.; Carotenuto, Y.; Vitiello, V.; Buttino, I.; Romano, G.; Hwang, J.-S.; Ianora, A. Effects of the Oxylipin-Producing Diatom Skeletonema marinoi on Gene Expression Levels of the Calanoid Copepod Calanus sinicus. Mar. Genom. 2015, 24, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Orefice, I.; Di Dato, V.; Sardo, A.; Lauritano, C.; Romano, G. Lipid Mediators in Marine Diatoms. Aquat. Ecol. 2022, 56, 377–397. [Google Scholar] [CrossRef]
- Ruocco, N.; Costantini, S.; Zupo, V.; Lauritano, C.; Caramiello, D.; Ianora, A.; Budillon, A.; Romano, G.; Nuzzo, G.; D’Ippolito, G.; et al. Toxigenic Effects of Two Benthic Diatoms upon Grazing Activity of the Sea Urchin: Morphological, Metabolomic and de Novo Transcriptomic Analysis. Sci. Rep. 2018, 8, 5622. [Google Scholar] [CrossRef] [Green Version]
- Rörig, L.R.; Honorato, M.C.; Itokazu, A.G.; de Macedo, C.X.; Deschamps, F.; Lins, J.V.H.; Maraschin, M.; Ramlov, F.; Gressler, P.D.; Pereira Filho, J. Ecophysiological and Biochemical Variation of the Surf Zone Diatom Asterionellopsis glacialis Sensu Lato from Santa Catarina, Southern Brazil. Braz. J. Ocean. 2017, 65, 695–708. [Google Scholar] [CrossRef] [Green Version]
- Viso, A.-C.; Marty, J.-C. Fatty Acids from 28 Marine Microalgae. Phytochemistry 1993, 34, 1521–1533. [Google Scholar] [CrossRef]
- Shibl, A.A.; Isaac, A.; Ochsenkühn, M.A.; Cárdenas, A.; Fei, C.; Behringer, G.; Arnoux, M.; Drou, N.; Santos, M.P.; Gunsalus, K.C.; et al. Diatom Modulation of Select Bacteria through Use of Two Unique Secondary Metabolites. Proc. Natl. Acad. Sci. USA 2020, 117, 27445–27455. [Google Scholar] [CrossRef]
- Lauritano, C.; Martín, J.; de la Cruz, M.; Reyes, F.; Romano, G.; Ianora, A. First Identification of Marine Diatoms with Anti-Tuberculosis Activity. Sci. Rep. 2018, 8, 2284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Ferrante, N.; Leachman, R.D.; Angelini, P.; Donnelly, P.V.; Francis, G.; Almazan, A.; Segni, G.; Franzblau, C.; Jordan, R.E. Ehlers-Danlos Type V (X-Linked Form): A Lysyl Oxidase Deficiency. Birth Defects Orig. Artic. Ser. 1975, 11, 31–37. [Google Scholar] [PubMed]
- Frangópulos, M.; Guisande, C.; deBlas, E.; Maneiro, I. Toxin Production and Competitive Abilities under Phosphorus Limitation of Alexandrium Species. Harmful Algae 2004, 3, 131–139. [Google Scholar] [CrossRef]
- Yang, I.; Beszteri, S.; Tillmann, U.; Cembella, A.; John, U. Growth- and Nutrient-Dependent Gene Expression in the Toxigenic Marine Dinoflagellate Alexandrium minutum. Harmful Algae 2011, 12, 55–69. [Google Scholar] [CrossRef] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-Length Transcriptome Assembly from RNA-Seq Data without a Reference Genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.; Niu, B.; Gao, Y.; Fu, L.; Li, W. CD-HIT Suite: A Web Server for Clustering and Comparing Biological Sequences. Bioinformatics 2010, 26, 680–682. [Google Scholar] [CrossRef] [PubMed]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Near-Optimal Probabilistic RNA-Seq Quantification. Nat. Biotechnol. 2016, 34, 525–527. [Google Scholar] [CrossRef]
- Smith-Unna, R.; Boursnell, C.; Patro, R.; Hibberd, J.M.; Kelly, S. TransRate: Reference-Free Quality Assessment of de Novo Transcriptome Assemblies. Genome Res. 2016, 26, 1134–1144. [Google Scholar] [CrossRef] [Green Version]
- Törönen, P.; Medlar, A.; Holm, L. PANNZER2: A Rapid Functional Annotation Web Server. Nucleic Acids Res. 2018, 46, W84–W88. [Google Scholar] [CrossRef] [Green Version]
- Tarazona, S.; García-Alcalde, F.; Dopazo, J.; Ferrer, A.; Conesa, A. Differential Expression in RNA-Seq: A Matter of Depth. Genome Res. 2011, 21, 2213–2223. [Google Scholar] [CrossRef]
- Tarazona, S.; Furió-Tarí, P.; Turrà, D.; Pietro, A.D.; Nueda, M.J.; Ferrer, A.; Conesa, A. Data Quality Aware Analysis of Differential Expression in RNA-Seq with NOISeq R/Bioc Package. Nucleic Acids Res. 2015, 43, e140. [Google Scholar] [CrossRef] [Green Version]
- Liu, K.K.-C.; Sakya, S.M.; O’Donnell, C.J.; Flick, A.C.; Ding, H.X. Synthetic Approaches to the 2010 New Drugs. Bioorg. Med. Chem. 2012, 20, 1155–1174. [Google Scholar] [CrossRef]
- Zhang, K.; Li, J.; Zhou, Z.; Huang, R.; Lin, S. Roles of Alkaline Phosphatase PhoA in Algal Metabolic Regulation under Phosphorus-Replete Conditions. J. Phycol. 2021, 57, 703–707. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Liu, H.; Wu, B. Structure-Based Drug Design of Catechol-O-Methyltransferase Inhibitors for CNS Disorders: Structure-Based Design of COMT Inhibitors. Br. J. Clin. Pharm. 2014, 77, 410–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Lai, L.; Liu, Y.; Yang, B.; Wang, Y. Expression and Characterization of a Novel Glycerophosphodiester Phosphodiesterase from Pyrococcus furiosus DSM 3638 That Possesses Lysophospholipase D Activity. Int. J. Mol. Sci. 2016, 17, 831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raetz, C.R.H. Molecular Genetics of Membrane Phospholipid Synthesis. Annu. Rev. Genet. 1986, 20, 253–291. [Google Scholar] [CrossRef]
- Abd El Baky, H.H.; El Baz, F.K.; El Baroty, G.S.; Asker, M.S.; Ibrahim, E.A. Phospholipids of Some Marine Microalgae: Identification, Antivirus, Anticancer and Antimicrobial Bioactivities. Der Pharm. Chem. 2014, 6, 9–18. [Google Scholar]
- Watanabe, H.; Shiratori, T.; Shoji, H.; Miyatake, S.; Okazaki, Y.; Ikuta, K.; Sato, T.; Saito, T. A Novel Acyl-CoA Thioesterase Enhances Its Enzymatic Activity by Direct Binding with HIV Nef. Biochem. Biophys. Res. Commun. 1997, 238, 234–239. [Google Scholar] [CrossRef]
- Göthel, S.F.; Marahiel, M.A. Peptidyl-Prolyl Cis-Trans Isomerases, a Superfamily of Ubiquitous Folding Catalysts. Cell. Mol. Life Sci. 1999, 55, 423–436. [Google Scholar] [CrossRef] [PubMed]
- Shaw, P.E. Peptidyl-prolyl Isomerases: A New Twist to Transcription. EMBO Rep. 2002, 3, 521–526. [Google Scholar] [CrossRef] [Green Version]
- Perrucci, G.L.; Gowran, A.; Zanobini, M.; Capogrossi, M.C.; Pompilio, G.; Nigro, P. Peptidyl-Prolyl Isomerases: A Full Cast of Critical Actors in Cardiovascular Diseases. Cardiovasc. Res. 2015, 106, 353–364. [Google Scholar] [CrossRef]
- Hitosugi, T.; Zhou, L.; Elf, S.; Fan, J.; Kang, H.-B.; Seo, J.H.; Shan, C.; Dai, Q.; Zhang, L.; Xie, J.; et al. Phosphoglycerate Mutase 1 Coordinates Glycolysis and Biosynthesis to Promote Tumor Growth. Cancer Cell 2012, 22, 585–600. [Google Scholar] [CrossRef] [Green Version]
- Vetting, M.W.; Hegde, S.S.; Fajardo, J.E.; Fiser, A.; Roderick, S.L.; Takiff, H.E.; Blanchard, J.S. Pentapeptide Repeat Proteins. Biochemistry 2006, 45, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Kieselbach, T.; Mant, A.; Robinson, C.; Schroder, W.P. Characterisation of an Arabidopsis CDNA Encoding a Thylakoid Lumen Protein Related to a Novel ‘pentapeptide Repeat’ Family of Proteins. FEBS Lett. 1998, 428, 241–244. [Google Scholar] [CrossRef] [Green Version]
- Simard, J.; Sanchez, R.; Durocher, F.; Rhéaume, E.; Turgeon, C.; Labrie, Y.; Luu-The, V.; Mebarki, F.; Morel, Y.; de Launoit, Y.; et al. Structure-Function Relationships and Molecular Genetics of the 3β-Hydroxysteroid Dehydrogenase Gene Family. J. Steroid Biochem. Mol. Biol. 1995, 55, 489–505. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Ji, Y.; Wang, Y.; Zhang, X.; Yu, Y. Cloning and Characterization of a Novel β-Ketoacyl-ACP Reductase from Comamonas testosteroni. Chem.-Biol. Interact. 2015, 234, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Maltsev, Y.; Maltseva, K. Fatty Acids of Microalgae: Diversity and Applications. Rev. Environ. Sci. Biotechnol. 2021, 20, 515–547. [Google Scholar] [CrossRef]
- Bateman, A.; Coggill, P.; Finn, R.D. DUFs: Families in Search of Function. Acta Cryst. F Struct. Biol. Cryst. Commun. 2010, 66, 1148–1152. [Google Scholar] [CrossRef] [Green Version]
- Griffith, L.C. CaMKII, an Enzyme on the Move: Regulation of Temporospatial Localization. Mol. Interv. 2003, 3, 386–403. [Google Scholar] [CrossRef] [PubMed]
- Eraso, J.M.; Markillie, L.M.; Mitchell, H.D.; Taylor, R.C.; Orr, G.; Margolin, W. The Highly Conserved MraZ Protein Is a Transcriptional Regulator in Escherichia coli. J. Bacteriol. 2014, 196, 2053–2066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gudkov, A.T. The L7/L12 Ribosomal Domain of the Ribosome: Structural and Functional Studies. FEBS Lett. 1997, 407, 253–256. [Google Scholar] [CrossRef]
- Raabe, K.; Honys, D.; Michailidis, C. The Role of Eukaryotic Initiation Factor 3 in Plant Translation Regulation. Plant Physiol. Biochem. 2019, 145, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Secco, D.; Wang, C.; Arpat, B.A.; Wang, Z.; Poirier, Y.; Tyerman, S.D.; Wu, P.; Shou, H.; Whelan, J. The Emerging Importance of the SPX Domain-containing Proteins in Phosphate Homeostasis. New Phytol. 2012, 193, 842–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dreyfuss, B.W.; Thornber, J.P. Assembly of the Light-Harvesting Complexes (LHCs) of Photosystem II (Monomeric LHC IIb Complexes Are Intermediates in the Formation of Oligomeric LHC IIb Complexes). Plant Physiol. 1994, 106, 829–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, M.; Kroth, P.G. Diatom Fucoxanthin Chlorophyll a/c-Binding Protein (FCP) and Land Plant Light-Harvesting Proteins Use a Similar Pathway for Thylakoid Membrane Insertion. J. Biol. Chem. 2001, 276, 7985–7991. [Google Scholar] [CrossRef] [Green Version]
- Vedalankar, P.; Tripathy, B.C. Evolution of Light-Independent Protochlorophyllide Oxidoreductase. Protoplasma 2019, 256, 293–312. [Google Scholar] [CrossRef]
- Wallace, J.C.; Jitrapakdee, S.; Chapman-Smith, A. Pyruvate Carboxylase. Int. J. Biochem. Cell Biol. 1998, 30, 1–5. [Google Scholar] [CrossRef]
- Horner, D.S.; Foster, P.G.; Embley, T.M. Iron Hydrogenases and the Evolution of Anaerobic Eukaryotes. Mol. Biol. Evol. 2000, 17, 1695–1709. [Google Scholar] [CrossRef] [Green Version]
- Batool, T.; Makky, E.A.; Jalal, M.; Yusoff, M.M. A Comprehensive Review on L-Asparaginase and Its Applications. Appl. Biochem. Biotechnol. 2016, 178, 900–923. [Google Scholar] [CrossRef] [Green Version]
- Ebrahiminezhad, A.; Rasoul-Amini, S.; Ghoshoon, M.B.; Ghasemi, Y. Chlorella Vulgaris, a Novel Microalgal Source for L-Asparaginase Production. Biocatal. Agric. Biotechnol. 2014, 3, 214–217. [Google Scholar] [CrossRef]
- Kumar, K.; Kaur, J.; Walia, S.; Pathak, T.; Aggarwal, D. L-Asparaginase: An Effective Agent in the Treatment of Acute Lymphoblastic Leukemia. Leuk. Lymphoma 2014, 55, 256–262. [Google Scholar] [CrossRef]
- Pieters, R.; Hunger, S.P.; Boos, J.; Rizzari, C.; Silverman, L.; Baruchel, A.; Goekbuget, N.; Schrappe, M.; Pui, C.-H. L-Asparaginase Treatment in Acute Lymphoblastic Leukemia: A Focus on Erwinia Asparaginase. Cancer 2011, 117, 238–249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarkson, B.; Krakoff, I.; Burchenal, J.; Karnofsky, D.; Golbey, R.; Dowling, M.; Oettgen, H.; Lipton, A. Clinical Results of Treatment with E. Coli L-Asparaginase in Adults with Leukemia, Lymphoma, and Solid Tumors. Cancer 1970, 25, 279–305. [Google Scholar] [CrossRef] [PubMed]
- Kobrinsky, N.L.; Sposto, R.; Shah, N.R.; Anderson, J.R.; DeLaat, C.; Morse, M.; Warkentin, P.; Gilchrist, G.S.; Cohen, M.D.; Shina, D.; et al. Outcomes of Treatment of Children and Adolescents with Recurrent Non-Hodgkin’s Lymphoma and Hodgkin’s Disease with Dexamethasone, Etoposide, Cisplatin, Cytarabine, and L-Asparaginase, Maintenance Chemotherapy, and Transplantation: Children’s Cancer Group Study CCG-5912. J. Clin. Oncol. 2001, 19, 2390–2396. [Google Scholar] [CrossRef]
- Abd El Baky, H.H.; El Baroty, G.S. Optimization of Growth Conditions for Purification and Production of L-Asparaginase by Spirulina maxima. Evid. Based Complement. Altern. Med. 2016, 2016, 1785938. [Google Scholar] [CrossRef] [Green Version]
- Lauritano, C.; De Luca, D.; Ferrarini, A.; Avanzato, C.; Minio, A.; Esposito, F.; Ianora, A. De Novo Transcriptome of the Cosmopolitan Dinoflagellate Amphidinium Carterae to Identify Enzymes with Biotechnological Potential. Sci. Rep. 2017, 7, 11701. [Google Scholar] [CrossRef] [Green Version]
- Gold, V. (Ed.) The IUPAC Compendium of Chemical Terminology: The Gold Book, 4th ed.; International Union of Pure and Applied Chemistry (IUPAC): Research Triangle Park, NC, USA, 2019. [Google Scholar]
- Vingiani, G.M.; De Luca, P.; Ianora, A.; Dobson, A.D.W.; Lauritano, C. Microalgal Enzymes with Biotechnological Applications. Mar. Drugs 2019, 17, 459. [Google Scholar] [CrossRef] [Green Version]
- Kornfuehrer, T.; Eustáquio, A.S. Diversification of Polyketide Structures via Synthase Engineering. Med. Chem. Commun. 2019, 10, 1256–1272. [Google Scholar] [CrossRef]
- Ridley, C.P.; Khosla, C. Polyketides. In Encyclopedia of Microbiology; Elsevier: Amsterdam, The Netherlands, 2009; pp. 472–481. ISBN 978-0-12-373944-5. [Google Scholar]
- Peebles, C.A.M.; Sander, G.W.; Hughes, E.H.; Peacock, R.; Shanks, J.V.; San, K.-Y. The Expression of 1-Deoxy-d-Xylulose Synthase and Geraniol-10-Hydroxylase or Anthranilate Synthase Increases Terpenoid Indole Alkaloid Accumulation in Catharanthus roseus Hairy Roots. Metab. Eng. 2011, 13, 234–240. [Google Scholar] [CrossRef] [PubMed]
- Yam-Puc, J.C.; Zhang, L.; Zhang, Y.; Toellner, K.-M. Role of B-Cell Receptors for B-Cell Development and Antigen-Induced Differentiation. F1000Research 2018, 7, 429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Yin, W.; Jing, Y.; Kang, D.; Yang, L.; Cheng, J.; Yu, Z.; Peng, Z.; Li, X.; Wen, Y.; et al. The Coordination Between B Cell Receptor Signaling and the Actin Cytoskeleton During B Cell Activation. Front. Immunol. 2019, 9, 3096. [Google Scholar] [CrossRef] [PubMed]
- Tolar, P. Cytoskeletal Control of B Cell Responses to Antigens. Nat. Rev. Immunol. 2017, 17, 621–634. [Google Scholar] [CrossRef] [PubMed]
- Hoyo, J.; Guaus, E.; Torrent-Burgués, J. Monogalactosyldiacylglycerol and Digalactosyldiacylglycerol Role, Physical States, Applications and Biomimetic Monolayer Films. Eur. Phys. J. E 2016, 39, 39. [Google Scholar] [CrossRef] [Green Version]
- Riccio, G.; De Luca, D.; Lauritano, C. Monogalactosyldiacylglycerol and Sulfolipid Synthesis in Microalgae. Mar. Drugs 2020, 18, 237. [Google Scholar] [CrossRef]
- Boudière, L.; Michaud, M.; Petroutsos, D.; Rébeillé, F.; Falconet, D.; Bastien, O.; Roy, S.; Finazzi, G.; Rolland, N.; Jouhet, J.; et al. Glycerolipids in Photosynthesis: Composition, Synthesis and Trafficking. Biochim. Biophys. Acta Bioenerg. 2014, 1837, 470–480. [Google Scholar] [CrossRef] [PubMed]
- Plyavnik, N.; Shtil, A.; Serebrennikova, G. Ether Lipids as Anticancer Agents: Focus on Non-Phosphorus Cationic Glycerolipids. MRMC 2006, 6, 533–542. [Google Scholar] [CrossRef]
- Shelest, E.; Heimerl, N.; Fichtner, M.; Sasso, S. Multimodular Type I Polyketide Synthases in Algae Evolve by Module Duplications and Displacement of AT Domains in Trans. BMC Genom. 2015, 16, 1015. [Google Scholar] [CrossRef] [Green Version]
- Huang, A.; Li, Y.; Duan, J.; Guo, S.; Cai, X.; Zhang, X.; Long, H.; Ren, W.; Xie, Z. Metabolomic, Proteomic and Lactylated Proteomic Analyses Indicate Lactate Plays Important Roles in Maintaining Energy and C:N Homeostasis in Phaeodactylum Tricornutum. Biotechnol. Biofuels 2022, 15, 61. [Google Scholar] [CrossRef]
- Di Dato, V.; Orefice, I.; Amato, A.; Fontanarosa, C.; Amoresano, A.; Cutignano, A.; Ianora, A.; Romano, G. Animal-like Prostaglandins in Marine Microalgae. ISME J. 2017, 11, 1722–1726. [Google Scholar] [CrossRef] [Green Version]
- Guillard, R.R.L. Culture of Phytoplankton for Feeding Marine Invertebrates. In Culture of Marine Invertebrate Animals; Smith, W.L., Chanley, M.H., Eds.; Springer: Boston, MA, USA, 1975; pp. 29–60. ISBN 978-1-4615-8716-3. [Google Scholar]
- Aceto, S.; Caputo, P.; Cozzolino, S.; Gaudio, L.; Moretti, A. Phylogeny and Evolution of Orchis and Allied Genera Based on ITS DNA Variation: Morphological Gaps and Molecular Continuity. Mol. Phylogenet. Evol. 1999, 13, 67–76. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the Sensitivity of Progressive Multiple Sequence Alignment through Sequence Weighting, Position-Specific Gap Penalties and Weight Matrix Choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest 2: More Models, New Heuristics and Parallel Computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akaike, H. A New Look at the Statistical Model Identification. IEEE Trans. Autom. Contr. 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, T.; Liu, Y.; Yan, H.; You, Q.; Yi, X.; Du, Z.; Xu, W.; Su, Z. AgriGO v2.0: A GO Analysis Toolkit for the Agricultural Community, 2017 Update. Nucleic Acids Res. 2017, 45, W122–W129. [Google Scholar] [CrossRef] [PubMed]



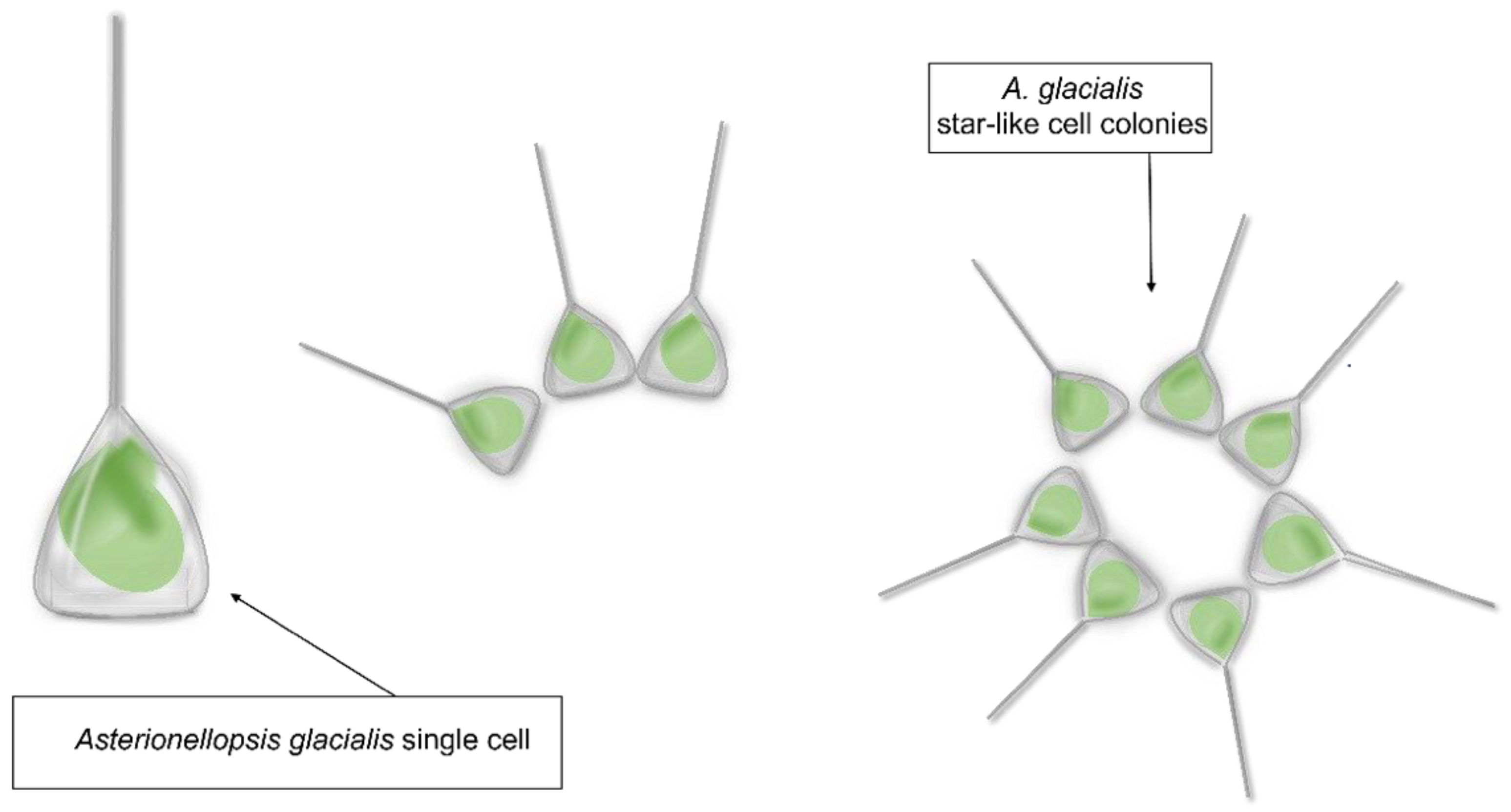{kind=link}
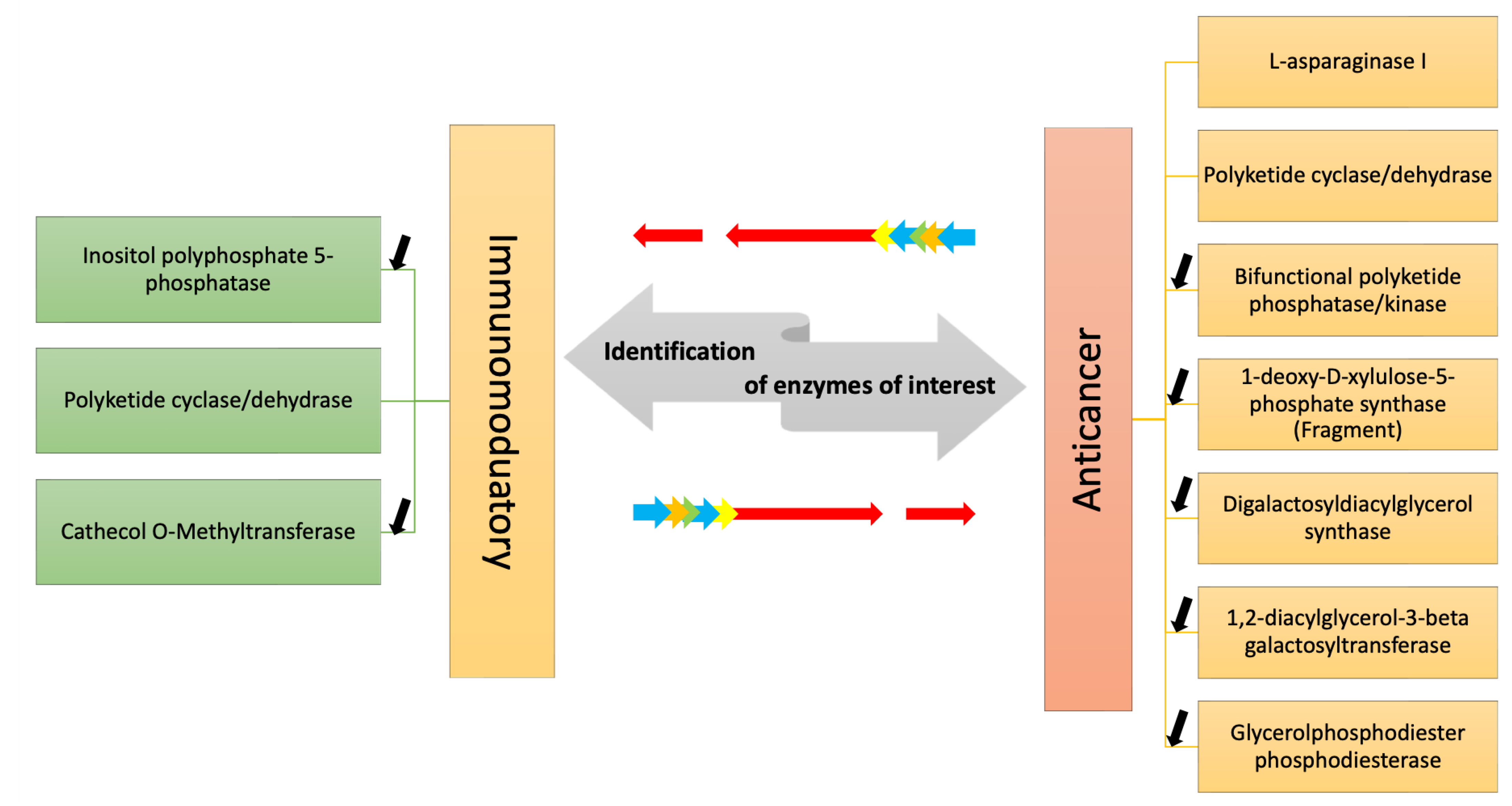{kind=link}
| Trinity (Raw Assembly) | CD-HIT | KALLISTO | BLAST | |
|---|---|---|---|---|
| FE355 | 44,430 | 42,571 | 42,527 | 40,493 |
| A4 | 46,835 | 44,637 | 44,586 | 41,698 |
| Parameters | FE355 Assembly | A4 Assembly |
|---|---|---|
| Total transcripts | 40,493 | 41,698 |
| Percent GC | 41.46 | 41.37 |
| Contig N50 | 1090 | 1127 |
| Median contig length | 558 | 597 |
| Average contig length | 771.53 | 802.21 |
| Total assembled bases | 31,241,667 | 33,450,597 |
| Comparison | Total DE Transcripts | Up-Regulated | Down-Regulated |
|---|---|---|---|
| A4_ phosphate starvation_vs._A4_Control | 3669 | 2276 | 1393 |
| FE355_ phosphate starvation_vs._FE355_Control | 3358 | 2206 | 1152 |
| A4_Control_vs._FE355_Control | 1628 | 441 | 1187 |
| Up-Regulated Genes | log2FC | Down-Regulated Genes | log2FC |
|---|---|---|---|
| O-methyltransferase family 3 | +8.41 | Peptidyl-prolyl cis–trans isomerase | −6.96 |
| Alkaline phosphatase | +7.99 | Phosphoglycerate mutase-like protein | −6.40 |
| Catechol O-methyltransferase domain-containing protein 1 | +7.91 | Pentapeptide repeat-containing protein | −6.24 |
| Glycerophosphodiester phosphodiesterase | +7.62 | Thylakoid lumenal 17.4 kDa protein, chloroplastic | −6.15 |
| Nucleotide-sugar transporter-domain-containing protein | +7.03 | 3-beta hydroxysteroid dehydrogenase/isomerase family | −5.54 |
| Acyl-CoA thioesterase | +6.62 | 3-oxoacyl-ACP synthase (Fragment) | −5.59 |
| Nucleotide-sugar transporter-domain-containing protein | +6.49 | DUF1995 domain-containing protein (Fragment) | −5.46 |
| Glycerophosphodiester phosphodiesterase (Fragment) | +6.44 | Peptidyl-prolyl cis–trans isomerase, chloroplastic-like isoform X2 | −5.39 |
| Solute carrier family 35, member C2 | +6.25 | Calcium/calmodulin dependent protein kinase II association-domain protein | −5.29 |
| DNA degradation protein EddB | +6.20 | Ternary protein-Dna Complex1 (Fragment) | −5.17 |
| Up-Regulated | log2FC | Down-Regulated | log2FC |
|---|---|---|---|
| Catechol O-methyltransferase domain-containing protein 1 | +8.50 | Fucoxanthin chlorophyll a/c protein | −7.87 |
| Glycerophosphodiester phosphodiesterase (Fragment) | +8.01 | Light-harvesting complex protein LHCC4 (fragment) | −7.19 |
| ATP-binding cassette | +7.96 | Light-harvesting complex I polypeptide | −7.02 |
| Glycerophosphodiester phosphodiesterase | +7.70 | Protochlorophyllide oxidoreductase | −6.86 |
| SPX domain-containing protein | +6.12 | Chloroplast light harvesting protein isoform 7 | −6.84 |
| Transcriptional regulator MraZ | +5.99 | Pyruvate carboxylase (fragment) | −6.54 |
| Extracellular nuclease, putative | +5.89 | Iron-only hydrogenase group A | −6.51 |
| Translation initiation factor IF-3 | +5.87 | Light harvesting complex protein 10 | −6.40 |
| Ribosomal protein l12 | +5.67 | Chloroplast light harvesting protein isoform 5 | −6.39 |
| Cell division cycle 2, cofactor of APC complex | +5.61 | Fucoxanthin-chlorophyll a/c light-harvesting protein (fragment) | −6.34 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montuori, E.; Martinez, K.A.; De Luca, D.; Ianora, A.; Lauritano, C. Transcriptome Sequencing of the Diatom Asterionellopsis thurstonii and In Silico Identification of Enzymes Potentially Involved in the Synthesis of Bioactive Molecules. Mar. Drugs 2023, 21, 126. https://doi.org/10.3390/md21020126
Montuori E, Martinez KA, De Luca D, Ianora A, Lauritano C. Transcriptome Sequencing of the Diatom Asterionellopsis thurstonii and In Silico Identification of Enzymes Potentially Involved in the Synthesis of Bioactive Molecules. Marine Drugs. 2023; 21(2):126. https://doi.org/10.3390/md21020126
Chicago/Turabian StyleMontuori, Eleonora, Kevin A. Martinez, Daniele De Luca, Adrianna Ianora, and Chiara Lauritano. 2023. "Transcriptome Sequencing of the Diatom Asterionellopsis thurstonii and In Silico Identification of Enzymes Potentially Involved in the Synthesis of Bioactive Molecules" Marine Drugs 21, no. 2: 126. https://doi.org/10.3390/md21020126
APA StyleMontuori, E., Martinez, K. A., De Luca, D., Ianora, A., & Lauritano, C. (2023). Transcriptome Sequencing of the Diatom Asterionellopsis thurstonii and In Silico Identification of Enzymes Potentially Involved in the Synthesis of Bioactive Molecules. Marine Drugs, 21(2), 126. https://doi.org/10.3390/md21020126