

Novel Antioxidant Peptides Identified from Arthrospira platensis Hydrolysates Prepared by a Marine Bacterium Pseudoalteromonas sp. JS4-1 Extracellular Protease

Abstract
:1. Introduction
2. Results
2.1. Screening of Proteases for Enzymatic Hydrolysis
2.2. Optimization of Enzymatic Hydrolyzing Parameters
2.3. Antioxidant Activities of Size-Exclusion Liquid Chromatography Fractions of Phycobiliprotein Hydrolysis Products
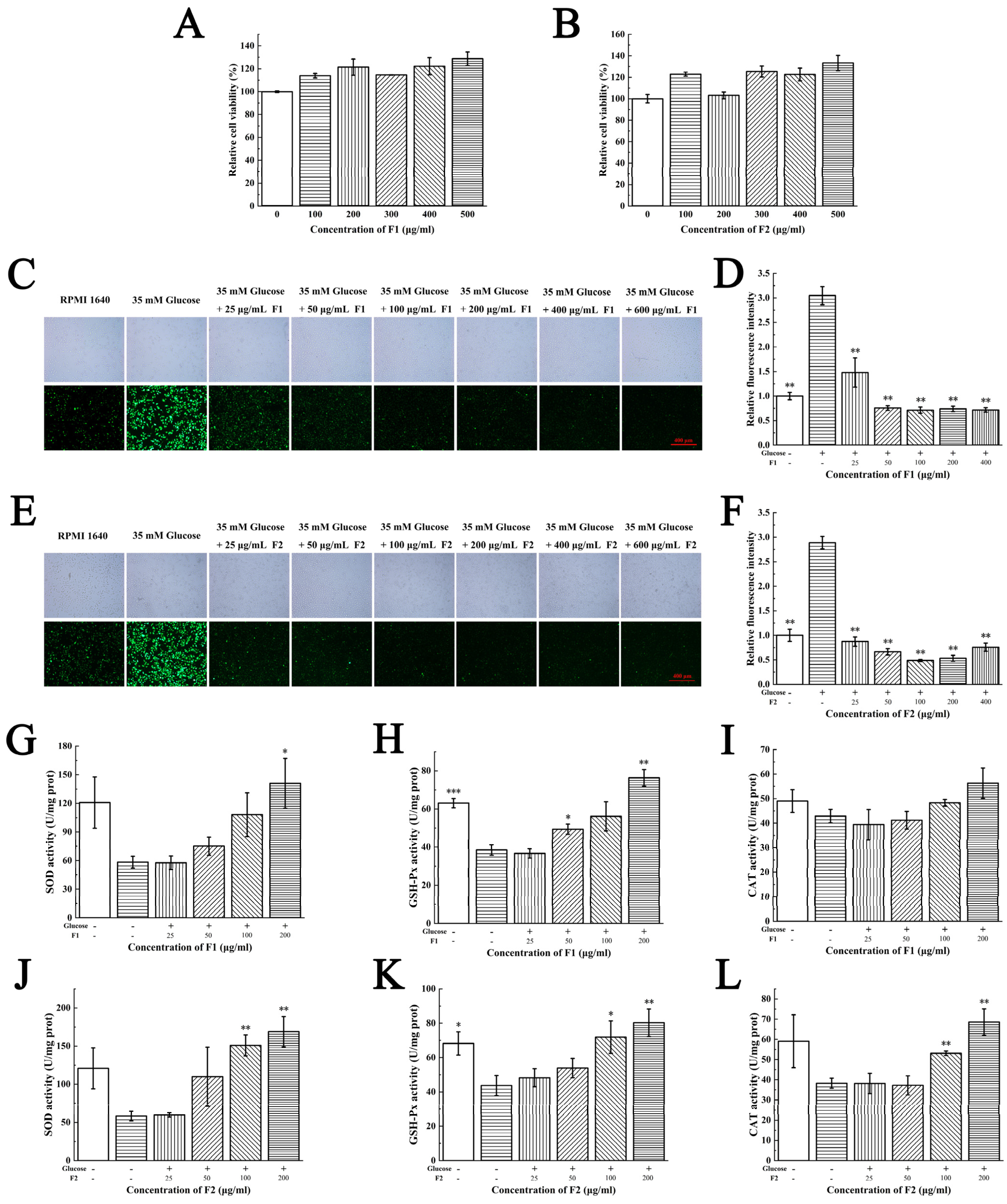
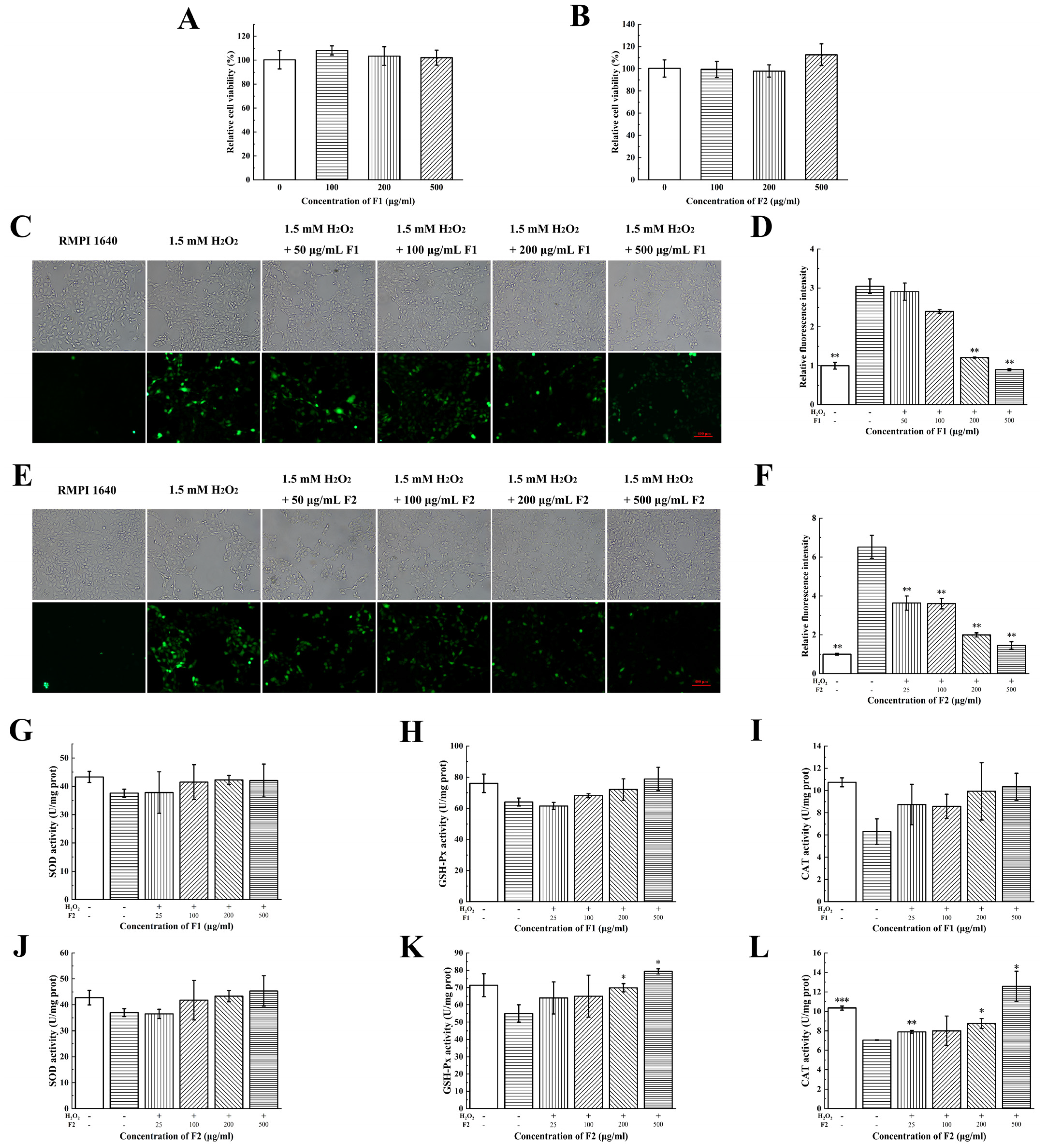
2.4. Antioxidant Activities of F1 and F2 at Cellular Levels
2.5. Effects of F2 on Resistance to Oxidative Stress in C. elegans
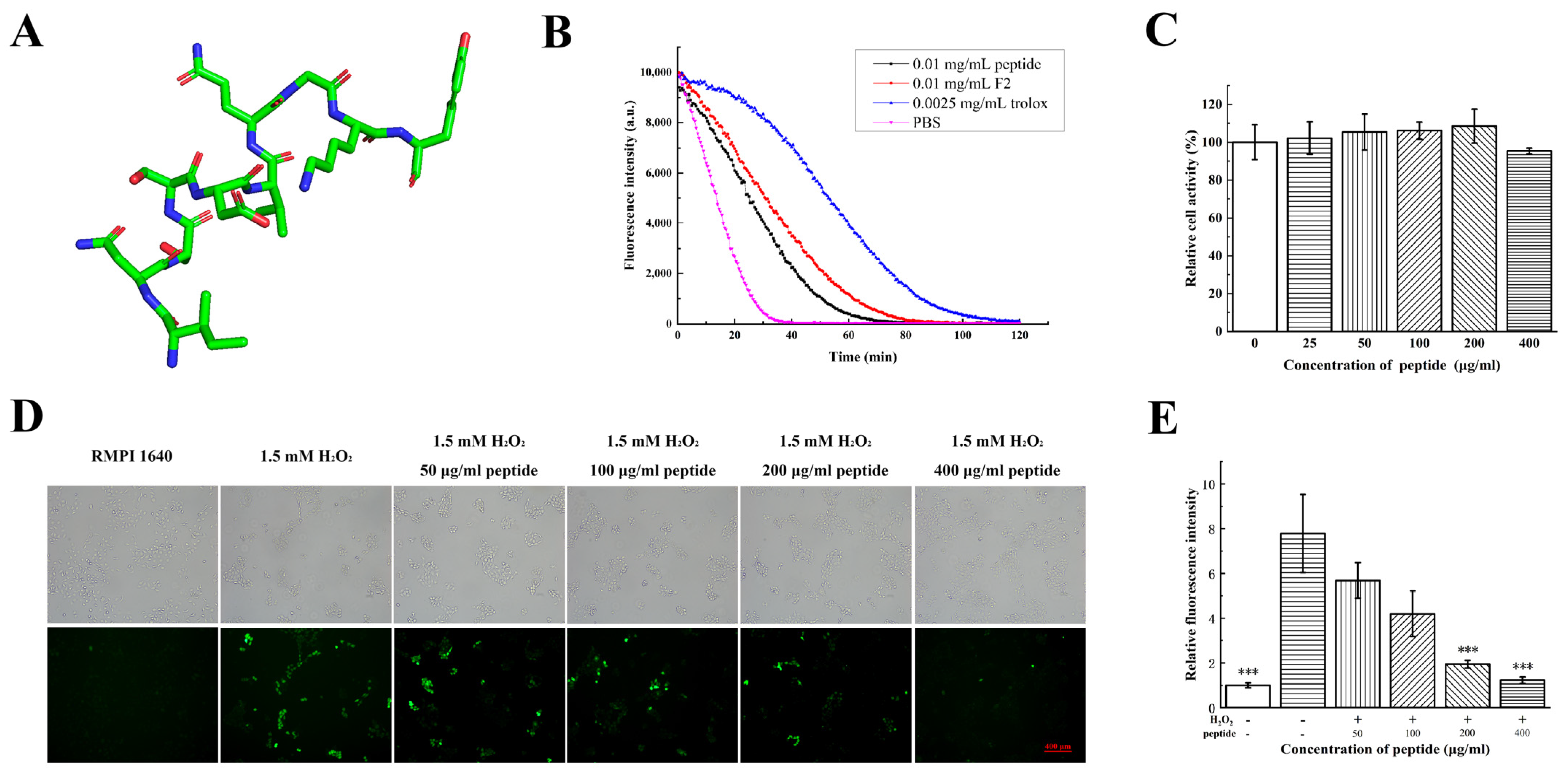
2.6. Antioxidant Activities of Synthesized Peptide INSSDVQGKY
3. Discussion
4. Materials and Methods
4.1. Strains and Reagents
4.2. Extraction of Phycobiliprotein and Screening of Proteases
4.2.1. Extraction of Phycobiliprotein from A. platensis
4.2.2. Preparation of Crude Bacterial Extracellular Protease
4.2.3. Substrate Immersing Zymography
4.3. Antioxidant Activities of Hydrolysates
4.3.1. Optimization of Hydrolysis Conditions
4.3.2. Purification of Hydrolysates
4.3.3. Determination of Antioxidant Activities
4.3.4. Protective Effects on Oxidative Damage of Plasmid DNA
4.3.5. Determination of Intracellular ROS Levels and the Activities of Intracellular Oxidative Enzymes
4.3.6. Antioxidant Activities Evaluation Using C. elegans Model
Maintenance of C. elegans and Lifespan Assay
Determination of ROS Levels and Activities of Antioxidant Enzymes in C. elegans
Oxidative Stress Experiment
4.3.7. Identification and Solid Phase Synthesis of APs
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lupatini, A.L.; Colla, L.M.; Canan, C.; Colla, E. Potential application of microalga Spirulina platensis as a protein source. J. Sci. Food Agric. 2017, 97, 724–732. [Google Scholar] [CrossRef] [PubMed]
- Romay, C.; González, R.; Ledón, N.; Remirez, D.; Rimbau, V. C-phycocyanin: A biliprotein with antioxidant, anti-inflammatory and neuroprotective effects. Curr. Protein Pept. Sci. 2003, 4, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Marthandam Asokan, S.; Wang, T.; Su, W.T.; Lin, W.T. Antidiabetic Effects of a Short Peptide of Potato Protein Hydrolysate in STZ-Induced Diabetic Mice. Nutrients 2019, 11, 779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, Q.; Chen, L.; Li, X.; Lu, X.; Xu, Q. Roles of Dietary Bioactive Peptides in Redox Balance and Metabolic Disorders. Oxid. Med. Cell Longev. 2021, 2021, 5582245. [Google Scholar] [CrossRef] [PubMed]
- Amigo, L.; Hernández-Ledesma, B. Current Evidence on the Bioavailability of Food Bioactive Peptides. Molecules 2020, 25, 4479. [Google Scholar] [CrossRef]
- Wang, K.; Han, L.; Hong, H.; Pan, J.; Liu, H.; Luo, Y. Purification and identification of novel antioxidant peptides from silver carp muscle hydrolysate after simulated gastrointestinal digestion and transepithelial transport. Food Chem. 2021, 342, 128275. [Google Scholar] [CrossRef]
- Chalamaiah, M.; Dinesh Kumar, B.; Hemalatha, R.; Jyothirmayi, T. Fish protein hydrolysates: Proximate composition, amino acid composition, antioxidant activities and applications: A review. Food Chem. 2012, 135, 3020–3038. [Google Scholar] [CrossRef]
- Lu, X.; Zhang, L.; Sun, Q.; Song, G.; Huang, J. Extraction, identification and structure-activity relationship of antioxidant peptides from sesame (Sesamum indicum L.) protein hydrolysate. Food Res. Int. 2019, 116, 707–716. [Google Scholar] [CrossRef]
- Olena, Z.; Yang, Y.; TingTing, Y.; XiaoTao, Y.; HaiLian, R.; Xun, X.; Dong, X.; CuiLing, W.; HaiLun, H. Simultaneous preparation of antioxidant peptides and lipids from microalgae by pretreatment with bacterial proteases. Bioresour. Technol. 2022, 348, 126759. [Google Scholar] [CrossRef]
- Xu, F.; Cha, Q.Q.; Zhang, Y.Z.; Chen, X.L. Degradation and Utilization of Alginate by Marine Pseudoalteromonas: A Review. Appl. Environ. Microbiol. 2021, 87, e0036821. [Google Scholar] [CrossRef]
- Kou, X.; Gao, J.; Xue, Z.; Zhang, Z.; Wang, H.; Wang, X. Purification and identification of antioxidant peptides from chickpea (Cicer arietinum L.) albumin hydrolysates. LWT—Food Sci. Technol. 2013, 50, 591–598. [Google Scholar] [CrossRef]
- Hou, H.; Wang, J.; Wang, J.; Tang, W.; Shaikh, A.S.; Li, Y.; Fu, J.; Lu, L.; Wang, F.; Sun, F.; et al. A Review of Bioactive Peptides: Chemical Modification, Structural Characterization and Therapeutic Applications. J. Biomed. Nanotechnol. 2020, 16, 1687–1718. [Google Scholar] [CrossRef] [PubMed]
- Cai, S.; Lu, C.; Liu, Z.; Wang, W.; Lu, S.; Sun, Z.; Wang, G. Derivatives of gecko cathelicidin-related antioxidant peptide facilitate skin wound healing. Eur. J. Pharmacol. 2021, 890, 173649. [Google Scholar] [CrossRef] [PubMed]
- Hseu, Y.C.; Vudhya Gowrisankar, Y.; Wang, L.W.; Zhang, Y.Z.; Chen, X.Z.; Huang, P.J.; Yen, H.R.; Yang, H.L. The in vitro and in vivo depigmenting activity of pterostilbene through induction of autophagy in melanocytes and inhibition of UVA-irradiated α-MSH in keratinocytes via Nrf2-mediated antioxidant pathways. Redox Biol. 2021, 44, 102007. [Google Scholar] [CrossRef]
- Wu, R.; Wu, C.; Liu, D.; Yang, X.; Huang, J.; Zhang, J.; Liao, B.; He, H. Antioxidant and anti-freezing peptides from salmon collagen hydrolysate prepared by bacterial extracellular protease. Food Chem. 2018, 248, 346–352. [Google Scholar] [CrossRef]
- Zou, J.; Fei, Q.; Xiao, H.; Wang, H.; Liu, K.; Liu, M.; Zhang, H.; Xiao, X.; Wang, K.; Wang, N. VEGF-A promotes angiogenesis after acute myocardial infarction through increasing ROS production and enhancing ER stress-mediated autophagy. J. Cell Physiol. 2019, 234, 17690–17703. [Google Scholar] [CrossRef]
- Nensat, C.; Songjang, W.; Tohtong, R.; Suthiphongchai, T.; Phimsen, S.; Rattanasinganchan, P.; Metheenukul, P.; Kumphune, S.; Jiraviriyakul, A. Porcine placenta extract improves high-glucose-induced angiogenesis impairment. BMC Complement. Med. Ther. 2021, 21, 66. [Google Scholar] [CrossRef]
- Rezabakhsh, A.; Rahbarghazi, R.; Malekinejad, H.; Fathi, F.; Montaseri, A.; Garjani, A. Quercetin alleviates high glucose-induced damage on human umbilical vein endothelial cells by promoting autophagy. Phytomedicine 2019, 56, 183–193. [Google Scholar] [CrossRef]
- Son, D.H.; Yang, D.J.; Sun, J.S.; Kim, S.K.; Kang, N.; Kang, J.Y.; Choi, Y.H.; Lee, J.H.; Moh, S.H.; Shin, D.M.; et al. A Novel Peptide, Nicotinyl-Isoleucine-Valine-Histidine (NA-IVH), Promotes Antioxidant Gene Expression and Wound Healing in HaCaT Cells. Mar. Drugs 2018, 16, 262. [Google Scholar] [CrossRef] [Green Version]
- Dong, H.; Liu, M.; Wang, L.; Liu, Y.; Lu, X.; Stagos, D.; Lin, X.; Liu, M. Bromophenol Bis (2,3,6-Tribromo-4,5-dihydroxybenzyl) Ether Protects HaCaT Skin Cells from Oxidative Damage via Nrf2-Mediated Pathways. Antioxidants 2021, 10, 1436. [Google Scholar] [CrossRef]
- Wang, G.; Hao, M.; Liu, Q.; Jiang, Y.; Huang, H.; Yang, G.; Wang, C. Protective effect of recombinant Lactobacillus plantarum against H(2)O(2)-induced oxidative stress in HUVEC cells. J. Zhejiang Univ. Sci. B 2021, 22, 348–365. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Xiao, J.; Xi, Y.; Zhang, X.; Zhong, Q.; Zheng, H.; Cao, Y.; Chen, Y. Rosmarinic acid improved antioxidant properties and healthspan via the IIS and MAPK pathways in Caenorhabditis elegans. Biofactors 2019, 45, 774–787. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H.; Shiozaki, T.; Kobatake, E.; Hosoya, T.; Moriya, T.; Sakai, F.; Taru, H.; Miyazaki, T. Effects and mechanisms of prolongevity induced by Lactobacillus gasseri SBT2055 in Caenorhabditis elegans. Aging Cell 2016, 15, 227–236. [Google Scholar] [CrossRef] [Green Version]
- Ayuda-Durán, B.; González-Manzano, S.; González-Paramás, A.M.; Santos-Buelga, C. Caernohabditis elegans as a Model Organism to Evaluate the Antioxidant Effects of Phytochemicals. Molecules 2020, 25, 3194. [Google Scholar] [CrossRef] [PubMed]
- Tsikas, D. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: Analytical and biological challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Meng, X.; Zhang, J. Multi- and trans-generational effects of N-butylpyridium chloride on reproduction, lifespan, and pro/antioxidant status in Caenorhabditis elegans. Sci. Total Environ. 2021, 778, 146371. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Hu, Y.; Xue, M.; Dun, Y.; Li, S.; Peng, N.; Liang, Y.; Zhao, S. Purification and Identification of Antioxidant Peptides from Enzymatic Hydrolysate of Spirulina platensis. J. Microbiol. Biotechnol. 2016, 26, 1216–1223. [Google Scholar] [CrossRef] [Green Version]
- Pagels, F.; Guedes, A.C.; Amaro, H.M.; Kijjoa, A.; Vasconcelos, V. Phycobiliproteins from cyanobacteria: Chemistry and biotechnological applications. Biotechnol. Adv. 2019, 37, 422–443. [Google Scholar] [CrossRef]
- Gómez-Estaca, J.; Bravo, L.; Gómez-Guillén, M.C.; Alemán, A.; Montero, P. Antioxidant properties of tuna-skin and bovine-hide gelatin films induced by the addition of oregano and rosemary extracts. Food Chem. 2009, 112, 18–25. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.Y.; Je, J.Y.; Kim, S.K. Purification and characterization of antioxidant peptide from hoki (Johnius belengerii) frame protein by gastrointestinal digestion. J. Nutr. Biochem. 2007, 18, 31–38. [Google Scholar] [CrossRef]
- Wu, R.; Chen, L.; Liu, D.; Huang, J.; Zhang, J.; Xiao, X.; Lei, M.; Chen, Y.; He, H. Preparation of Antioxidant Peptides from Salmon Byproducts with Bacterial Extracellular Proteases. Mar. Drugs 2017, 15, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fontoura, R.; Daroit, D.J.; Corrêa, A.P.F.; Moresco, K.S.; Santi, L.; Beys-da-Silva, W.O.; Yates, J.R., 3rd; Moreira, J.C.F.; Brandelli, A. Characterization of a novel antioxidant peptide from feather keratin hydrolysates. New Biotechnol. 2019, 49, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Huang, K.; Wu, Y. In Vivo/In Vitro Properties of Novel Antioxidant Peptide from Pinctada fucata. J. Microbiol. Biotechnol. 2021, 31, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Jia, Z.; Trush, M.A. Defining ROS in Biology and Medicine. React. Oxyg Species 2016, 1, 9–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants Maintain Cellular Redox Homeostasis by Elimination of Reactive Oxygen Species. Cell Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef] [PubMed]
- Christofidou-Solomidou, M.; Muzykantov, V.R. Antioxidant strategies in respiratory medicine. Treat Respir. Med. 2006, 5, 47–78. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Zhan, J.; Hu, L.; Yuan, C.; Ying, X.; Hu, Y. Identification of novel antioxidant peptide from porcine plasma hydrolysate and its effect in in vitro digestion/HepG2 cells model. J. Food Biochem. 2022, 46, e13853. [Google Scholar] [CrossRef]
- Park, Y.R.; Park, C.I.; Soh, Y. Antioxidant and Anti-Inflammatory Effects of NCW Peptide from Clam Worm (Marphysa sanguinea). J. Microbiol. Biotechnol. 2020, 30, 1387–1394. [Google Scholar] [CrossRef]
- Walker, J.M. The bicinchoninic acid (BCA) assay for protein quantitation. Methods Mol. Biol. 1994, 32, 5–8. [Google Scholar]
- Liu, D.; Yang, X.; Huang, J.; Wu, R.; Wu, C.; He, H.; Li, H. In situ demonstration and characteristic analysis of the protease components from marine bacteria using substrate immersing zymography. Appl. Biochem. Biotechnol. 2015, 175, 489–501. [Google Scholar] [CrossRef]
- Leach, A.A. The determination of the substitution achieved at the alpha-amino, epsilon-amino and imidazole groups of proteins with special reference to derivatives of gelatin. Biochem. J. 1966, 98, 506–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dávalos, A.; Gómez-Cordovés, C.; Bartolomé, B. Extending applicability of the oxygen radical absorbance capacity (ORAC-fluorescein) assay. J. Agric. Food Chem. 2004, 52, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Shimada, K.; Fujikawa, K.; Yahara, K.; Nakamura, T. Antioxidative properties of xanthan on the autoxidation of soybean oil in cyclodextrin emulsion. J. Agric. Food Chem. 1992, 40, 945–948. [Google Scholar] [CrossRef]
- Wang, B.; Li, Z.R.; Chi, C.F.; Zhang, Q.H.; Luo, H.Y. Preparation and evaluation of antioxidant peptides from ethanol-soluble proteins hydrolysate of Sphyrna lewini muscle. Peptides 2012, 36, 240–250. [Google Scholar] [CrossRef]
- Alugoju, P.; Janardhanshetty, S.S.; Subaramanian, S.; Periyasamy, L.; Dyavaiah, M. Quercetin Protects Yeast Saccharomyces cerevisiae pep4 Mutant from Oxidative and Apoptotic Stress and Extends Chronological Lifespan. Curr. Microbiol. 2018, 75, 519–530. [Google Scholar] [CrossRef]
- Lin, C.; Su, Z.; Luo, J.; Jiang, L.; Shen, S.; Zheng, W.; Gu, W.; Cao, Y.; Chen, Y. Polysaccharide extracted from the leaves of Cyclocarya paliurus (Batal.) Iljinskaja enhanced stress resistance in Caenorhabditis elegans via skn-1 and hsf-1. Int. J. Biol. Macromol. 2020, 143, 243–254. [Google Scholar] [CrossRef]
- Sakamoto, T.; Imai, H. Hydrogen peroxide produced by superoxide dismutase SOD-2 activates sperm in Caenorhabditis elegans. J. Biol. Chem. 2017, 292, 14804–14813. [Google Scholar] [CrossRef] [Green Version]
- Yasuda, K.; Kubo, Y.; Murata, H.; Sakamoto, K. Cortisol promotes stress tolerance via DAF-16 in Caenorhabditis elegans. Biochem. Biophys. Rep. 2021, 26, 100961. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Amino Acid Sequence | Mass (Da) | Source (Position) | Counts (%) |
|---|---|---|---|
| INSSDVQGKY | 1109.535 | Allophycocyanin beta-subunit [Arthrospira platensis qy3] (9–18) | 17.83 |
| VGGSVPREY | 962.482 | Elongation factor G [Arthrospira platensis] (531–539) | 9.12 |
| SSYPNRPVP | 1015.509 | Photosystem II lipoprotein Psb27 [Arthrospira platensis C1] (102–110) | 8.81 |
| GIFDTMNH | 933.401 | ATP-dependent Clp protease proteolytic subunit [Arthrospira platensis C1] (74–81) | 6.91 |
| YTPDYTPK | 983.460 | Ribulose bisphosphate carboxylase large chain [Arthrospira platensis C1] (26–33) | 3.34 |
| IDPSHGTGF | 929.424 | Phospho-2-dehydro-3-deoxyheptonate aldolase [Arthrospira platensis] (277–285) | 3.33 |
| TPEPKPEPKPEPKPEP | 1795.936 | CAAD domain-containing protein [Arthrospira platensis C1] (52–67) | 3.25 |
| YEQMPEPKY | 1183.522 | NAD(P)H-quinone oxidoreductase subunit K [Arthrospira platensis C1] (105–113) | 3.14 |
| DVDWSDYQKQ | 1282.547 | Ferredoxin-NADP reductase [Arthrospira platensis] (380–389) | 2.78 |
| VGGSVPKEY | 934.476 | Elongation factor G [Arthrospira platensis C1] (496–504) | 2.76 |
| TDVPANHPY | 1012.461 | S-layer domain protein [Arthrospira platensis C1] (283–291) | 2.42 |
| QGTLEKYI | 950.507 | Putative adenylate cyclase [Arthrospira platensis C1] (375–382) | 2.40 |
| AGIDEINRT | 987.499 | Phycocyanin alpha-subunit [Arthrospira platensis qy3] (113–121) | 2.32 |
| SESPNLILMDIQMP | 1586.768 | Multi-sensor hybrid histidine kinase [Arthrospira platensis C1] (1700–1713) | 2.32 |
| APYDESEVVFH | 1291.572 | Rod linker polypeptide CpcI [Arthrospira platensis] (107–117) | 2.20 |
| IDTIPTGGK | 900.492 | 50S ribosomal protein L13 [Arthrospira platensis C1] (144–152) | 1.96 |
| LIAGGTGPM | 815.421 | Phycocyanin alpha-subunit [Arthrospira platensis qy3] (99–107) | 1.92 |
| DVDWSDYQK | 1154.488 | Ferredoxin-NADP reductase [Arthrospira platensis] (380–388) | 1.90 |
| LPEEPMTGK | 1000.490 | Transcriptional regulator AbrB family [Arthrospira platensis C1] (9–17) | 1.80 |
| VINSSDVQGKY | 1208.604 | Allophycocyanin beta-subunit [Arthrospira platensis qy3] (8–18) | 1.68 |
| LVTQQPLGGKAQ | 1238.698 | DNA-directed RNA polymerase subunit beta [Arthrospira platensis] (992–1003) | 1.52 |
| KELTKKSPNSP | 1227.682 | NYN domain-containing protein [Arthrospira platensis] (398–408) | 1.37 |
| IETKEIPVPT | 1125.628 | Assimilatory sulfite reductase (ferredoxin) [Arthrospira platensis C1] (592–601) | 1.34 |
| VDYQEQPREY | 1325.589 | Major membrane protein I [Arthrospira platensis C1] (81–90) | 1.30 |
| GANYEDEWK | 1110.462 | Transketolase [Arthrospira platensis] (308–316) | 1.30 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, C.; Chen, G.; Rao, H.; Xiao, X.; Chen, Y.; Wu, C.; Bian, F.; He, H. Novel Antioxidant Peptides Identified from Arthrospira platensis Hydrolysates Prepared by a Marine Bacterium Pseudoalteromonas sp. JS4-1 Extracellular Protease. Mar. Drugs 2023, 21, 133. https://doi.org/10.3390/md21020133
Liu C, Chen G, Rao H, Xiao X, Chen Y, Wu C, Bian F, He H. Novel Antioxidant Peptides Identified from Arthrospira platensis Hydrolysates Prepared by a Marine Bacterium Pseudoalteromonas sp. JS4-1 Extracellular Protease. Marine Drugs. 2023; 21(2):133. https://doi.org/10.3390/md21020133
Chicago/Turabian StyleLiu, Congling, Gong Chen, Hailian Rao, Xun Xiao, Yidan Chen, Cuiling Wu, Fei Bian, and Hailun He. 2023. "Novel Antioxidant Peptides Identified from Arthrospira platensis Hydrolysates Prepared by a Marine Bacterium Pseudoalteromonas sp. JS4-1 Extracellular Protease" Marine Drugs 21, no. 2: 133. https://doi.org/10.3390/md21020133
APA StyleLiu, C., Chen, G., Rao, H., Xiao, X., Chen, Y., Wu, C., Bian, F., & He, H. (2023). Novel Antioxidant Peptides Identified from Arthrospira platensis Hydrolysates Prepared by a Marine Bacterium Pseudoalteromonas sp. JS4-1 Extracellular Protease. Marine Drugs, 21(2), 133. https://doi.org/10.3390/md21020133