

Large-Scale Cultivation of Magnetotactic Bacteria and the Optimism for Sustainable and Cheap Approaches in Nanotechnology
 , , , and
, , , and
Abstract
:1. Introduction
2. Bioreactor Cultivation Strategies
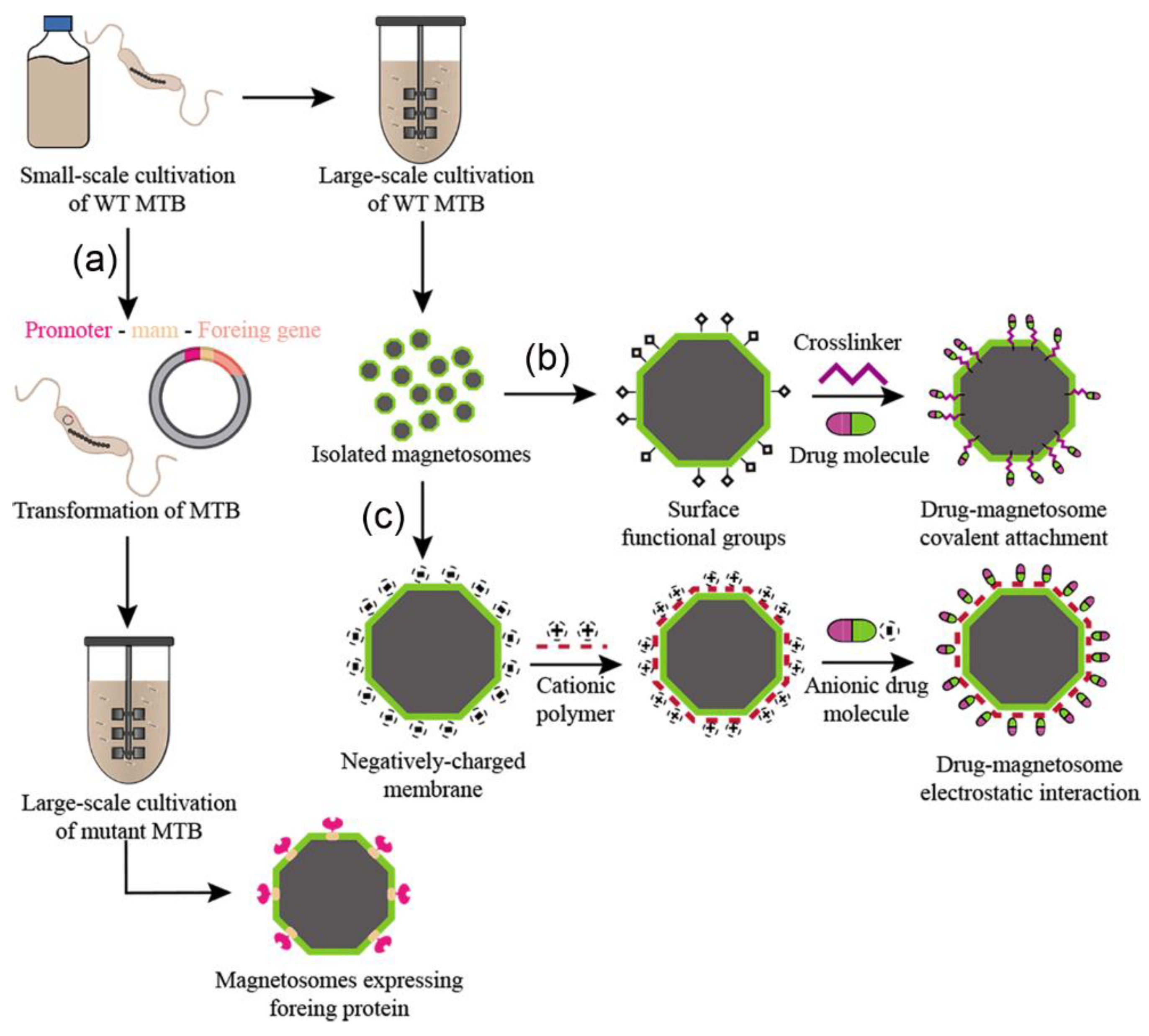
3. BMN Purification Methods
4. Functionalization Methods
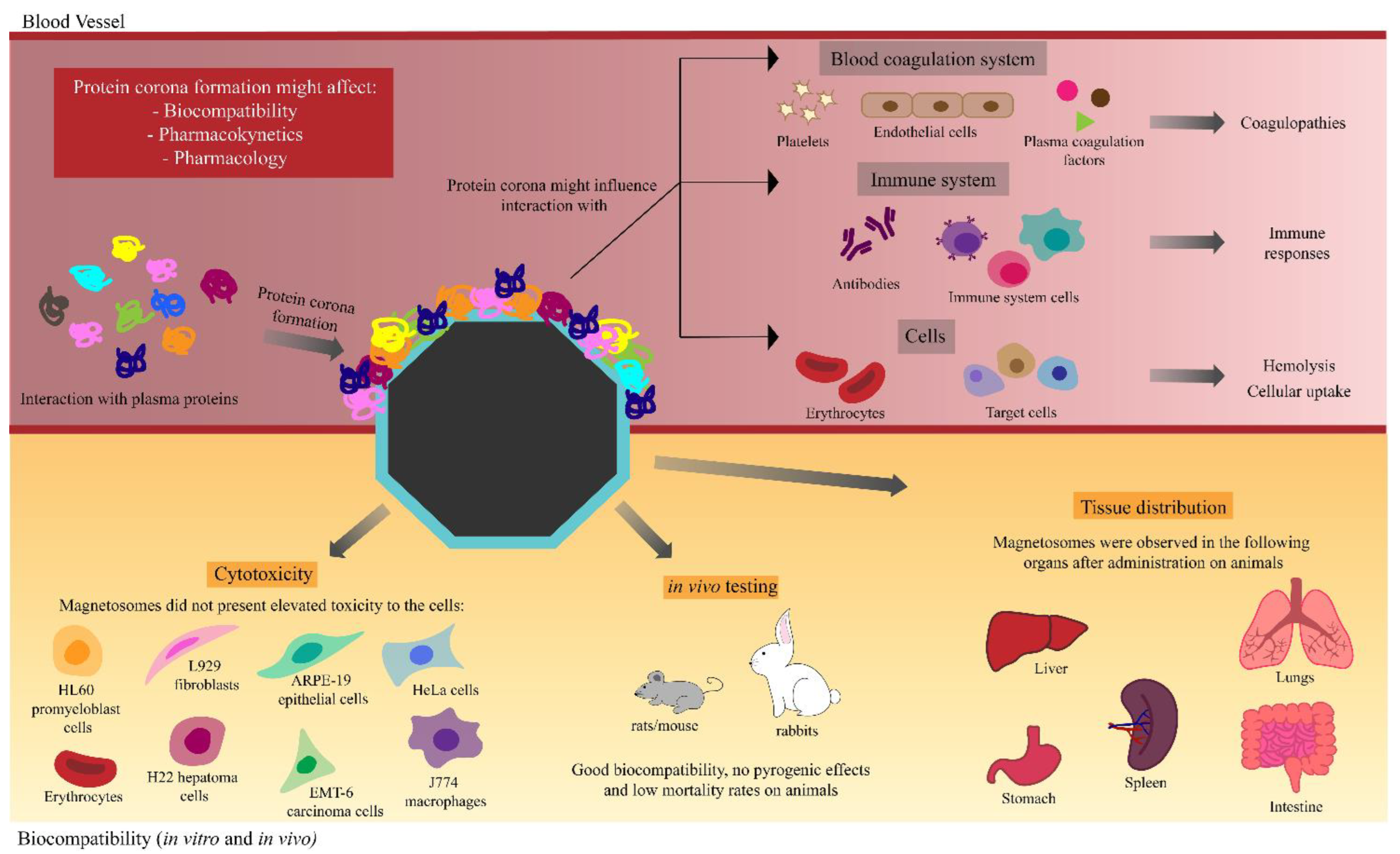
5. Biomedical Applications
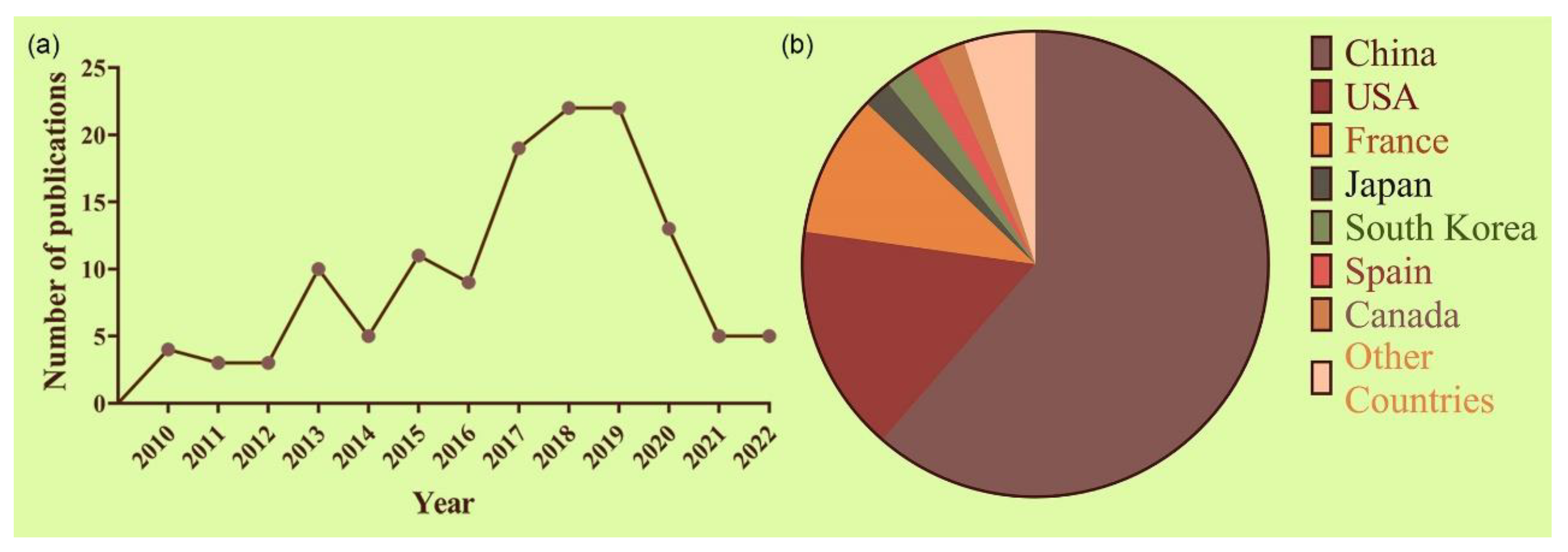
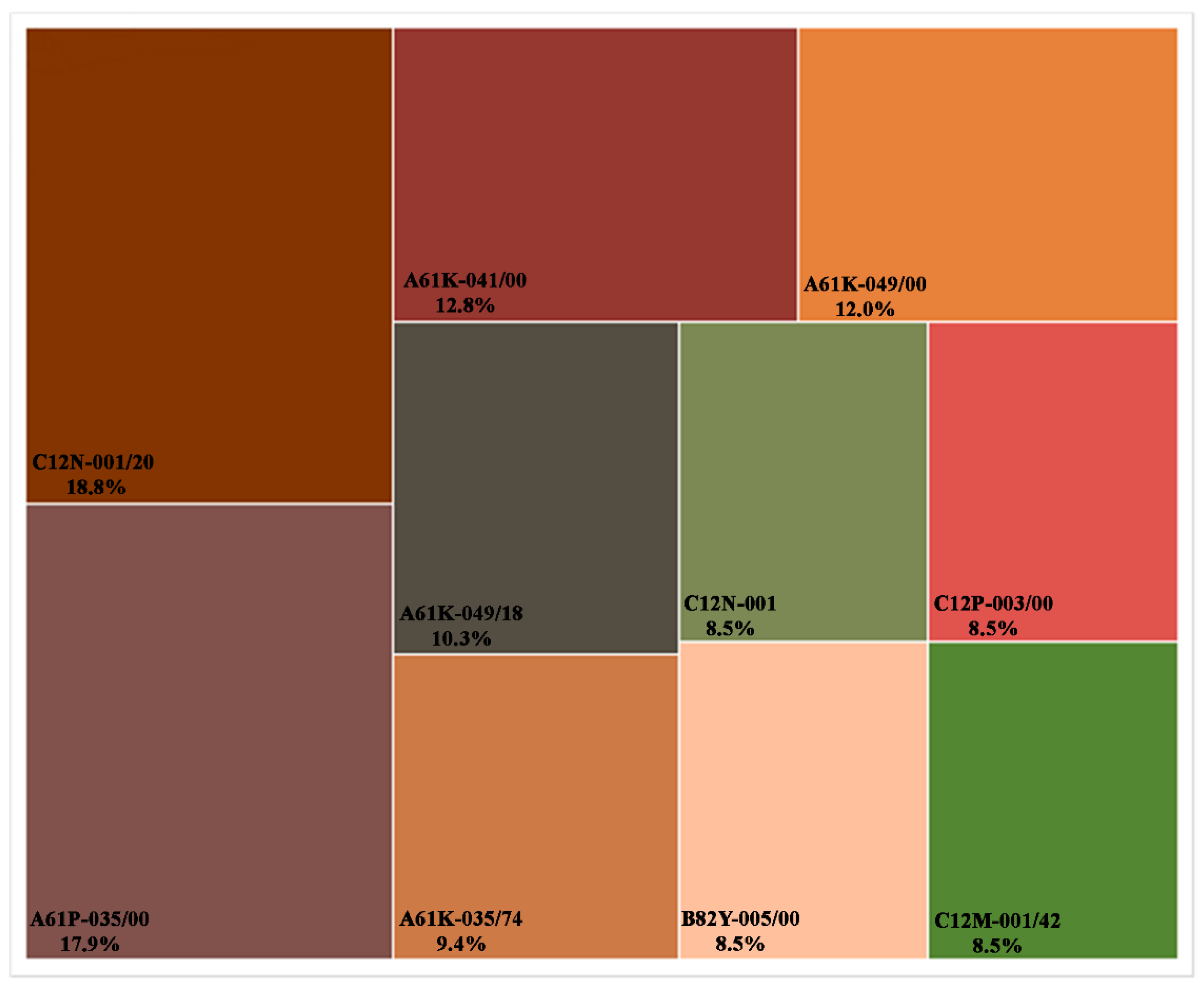
6. Technological Outputs
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blakemore, R. Magnetotactic bacteria. Science 1975, 190, 377–379. [Google Scholar] [CrossRef] [PubMed]
- Abreu, F.; Acosta-Avalos, D. Biology and Physics of Magnetotactic Bacteria. In Microorganisms, 1st ed.; Blumenberg, M., Shaaban, M., Elgaml, A., Eds.; IntechOpen: London, UK, 2018; pp. 1–19. [Google Scholar] [CrossRef] [Green Version]
- Vargas, G.; Cypriano, J.; Correa, T.; Leão, P.; Bazylinski, D.A.; Abreu, F. Applications of magnetotactic bacteria, magnetosomes and magnetosome crystals in biotechnology and nanotechnology: Mini-review. Molecules 2018, 23, 2438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazylinski, D.A.; Frankel, R.B. Magnetosome formation in prokaryotes. Nat. Rev. Microbiol. 2004, 2, 217–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunin-Borkowski, R.E.; McCartney, M.R.; Frankel, R.B.; Bazylinski, D.A.; Pósfai, M.; Buseck, P.R. Magnetic Microstructure of Magnetotactic Bacteria by Electron Holography. Science 1998, 282, 1868–1870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blakemore, R.P.; Short, K.A.; Bazylinski, D.A.; Rosenblatt, C.; Frankel, R.B. Microaerobic Conditions Are Required for Magnetite Formation within Aquaspirillum magnetotacticum. Geomicrobiol. J. 1984, 4, 53–71. [Google Scholar] [CrossRef] [Green Version]
- Zhulin, I.B.; Bespalov, V.A.; Johnson, M.S.; Taylor, B.L. Oxygen taxis and proton motive force in Azospirillum brasilense. J. Bacteriol. 1996, 178, 5199–5204. [Google Scholar] [CrossRef] [Green Version]
- Lin, W.; Pan, Y.; Bazylinski, D.A. Diversity and ecology of and biomineralization by magnetotactic bacteria. Environ. Microbiol. Rep. 2017, 9, 345–356. [Google Scholar] [CrossRef]
- Matsunaga, T.; Kamiya, S. Use of magnetic particles isolated from magnetotactic bacteria for enzyme immobilization. Appl. Microbiol. Biotechnol. 1987, 26, 328–332. [Google Scholar] [CrossRef]
- Gorby, Y.A.; Beveridge, T.J.; Blakemore, R.P. Characterization of the bacterial magnetosome membrane. J. Bacteriol. 1988, 170, 834–841. [Google Scholar] [CrossRef]
- Guo, L.; Huang, J.; Zhang, X.; Li, Y.; Zheng, L. Bacterial magnetic nanoparticles as drug carriers. J. Mater. Chem. 2008, 18, 5993–5997. [Google Scholar] [CrossRef]
- Rosenfeldt, S.; Mickoleit, F.; Jörke, C.; Clement, J.H.; Markert, S.; Jérôme, V.; Schwarzinger, S.; Freitag, R.; Schüler, D.; Uebe, R.; et al. Towards standardized purification of bacterial magnetic nanoparticles for future in vivo applications. Acta Biomater. 2021, 120, 293–303. [Google Scholar] [CrossRef]
- Yan, L.; Da, H.; Zhang, S.; López, V.M.; Wang, W. Bacterial magnetosome and its potential application. Microbiol. Res. 2017, 203, 19–28. [Google Scholar] [CrossRef]
- Tay, A.; McCausland, H.; Komeili, A.; Di Carlo, D. Nano and Microtechnologies for the Study of Magnetotactic Bacteria. Adv. Funct. Mater. 2019, 29, 1904178. [Google Scholar] [CrossRef] [Green Version]
- Peigneux, A.; Valverde-Tercedor, C.; López-Moreno, R.; Pérez-González, T.; Fernández-Vivas, M.A.; Jiménez-López, C. Learning from magnetotactic bacteria: A review on the synthesis of biomimetic nanoparticles mediated by magnetosome-associated proteins. J. Struct. Biol. 2016, 196, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Arakaki, A.; Masuda, F.; Amemiya, Y.; Tanaka, T.; Matsunaga, T. Control of the morphology and size of magnetite particles with peptides mimicking the Mms6 protein from magnetotactic bacteria. J. Colloid Interface Sci. 2010, 343, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Prozorov, T.; Bazylinski, D.A.; Mallapragada, S.K.; Prozorov, R. Novel magnetic nanomaterials inspired by magnetotactic bacteria: Topical review. Mater. Sci. Eng. R Rep. 2013, 74, 133–172. [Google Scholar] [CrossRef]
- Nudelman, H.; Zarivach, R. Structure prediction of magnetosome-associated proteins. Front. Microbiol. 2014, 5, 9. [Google Scholar] [CrossRef]
- Valverde-Tercedor, C.; Abadía-Molina, F.; Martinez-Bueno, M.; Pineda-Molina, E.; Chen, L.; Oestreicher, Z.; Lower, B.H.; Lower, S.K.; Bazylinski, D.A.; Jimenez-Lopez, C. Subcellular localization of the magnetosome protein MamC in the marine magnetotactic bacterium Magnetococcus marinus strain MC-1 using immunoelectron microscopy. Arch. Microbiol. 2014, 196, 481–488. [Google Scholar] [CrossRef]
- Valverde-Tercedor, C.; Montalbán-López, M.; Perez-Gonzalez, T.; Sanchez-Quesada, M.S.; Prozorov, T.; Pineda-Molina, E.; Fernandez-Vivas, M.A.; Rodriguez-Navarro, A.B.; Trubitsyn, D.; Bazylinski, D.A.; et al. Size control of in vitro synthesized magnetite crystals by the MamC protein of Magnetococcus marinus strain MC-1. Appl. Microb. Biotechnol. 2015, 99, 5109–5121. [Google Scholar] [CrossRef]
- Nudelman, H.; Valverde-Tercedor, C.; Kolusheva, S.; Pérez-González, T.; Widdrat, M.; Grimberg, N.; Zarivach, R. Structure–function studies of the magnetite-biomineralizing magnetosome-associated protein MamC. J. Struct. Biol. 2016, 194, 244–252. [Google Scholar] [CrossRef]
- Bazylinski, D.A.; Frankel, R.B.; Heywook, B.R.; Mann, S.; King, J.W.; Donaghay, P.L.; Hanson, A.K. Controlled biomineralization of magnetite (Fe3O4) and greigite (Fe3S4) in a magnetotactic bacterium. Appl. Environ. Microbiol. 1995, 61, 3232–3239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araujo, A.; Abreu, F.; Silva, K.; Bazylinski, D.; Lins, U. Magnetotactic Bacteria as Potential Sources of Bioproducts. Mar. Drugs 2015, 13, 389–430. [Google Scholar] [CrossRef] [PubMed]
- Mickoleit, F.; Borkner, C.B.; Toro-Nahuelpan, M.; Herold, H.M.; Maier, D.S.; Plitzko, J.M.; Scheibel, T.; Schüler, D. In Vivo Coating of Bacterial Magnetic Nanoparticles by Magnetosome Expression of Spider Silk-Inspired Peptides. Biomacromolecules 2018, 19, 962–972. [Google Scholar] [CrossRef] [PubMed]
- Cypriano, J.; Bahri, M.; Dembelé, K.; Baaziz, W.; Leão, P.; Bazylinski, D.A.; Abreu, F.; Ersen, O.; Farina, M.; Werckmann, J. Insight on thermal stability of magnetite magnetosomes: Implications for the fossil record and biotechnology. Sci. Rep. 2020, 10, 6706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazylinski, D.A.; Garratt-Reed, A.J.; Frankel, R.B. Electron Microscopic Studies of Magnetosomes in Magnetotactic Bacteria. Microsc. Res. Tech. 1994, 27, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Faivre, D.; Schüler, D. Magnetotactic Bacteria and Magnetosomes. Chem. Rev. 2008, 108, 4875–4898. [Google Scholar] [CrossRef] [PubMed]
- Uebe, R.; Schüler, D. Magnetosome biogenesis in magnetotactic bacteria. Nat. Rev. Microbiol. 2016, 14, 621–637. [Google Scholar] [CrossRef]
- Barber-Zucker, S.; Zarivach, R. A Look into the Biochemistry of Magnetosome Biosynthesis in Magnetotactic Bacteria. ACS Chem. Biol. 2017, 12, 13–22. [Google Scholar] [CrossRef]
- Jogler, C.; Schüler, D. Genomics, Genetics, and Cell Biology of Magnetosome Formation. Ann. Rev. Microbiol. 2009, 63, 501–521. [Google Scholar] [CrossRef]
- Lefèvre, C.T.; Bazylinski, D.A. Ecology, diversity, and evolution of magnetotactic bacteria. Microbiol. Mol. Biol. Rev. 2013, 77, 497–526. [Google Scholar] [CrossRef] [Green Version]
- Pósfai, M.; Lefevre, C.T.; Trubitsyn, D.; Bazylinski, D.A.; Frankel, R.B. Phylogenetic significance of composition and crystal morphology of magnetosome minerals. Front. Microbiol. 2013, 4, 344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, W.; Bazylinski, D.A.; Xiao, T.; Wu, L.F.; Pan, Y. Life with compass: Diversity and biogeography of magnetotactic bacteria. Environ. Microbiol. 2014, 16, 2646–2658. [Google Scholar] [CrossRef] [PubMed]
- Koziaeva, V.V.; Rusakova, S.A.; Slobodova, N.V.; Uzun, M.; Kolganova, T.V.; Skryabin, K.G.; Grouzdev, D.S. Magnetospirillum kuznetsovii sp. nov., a novel magnetotactic bacterium isolated from a lake in the Moscow region. Int. J. Syst. Evol. Microbiol. 2019, 69, 1953–1959. [Google Scholar] [CrossRef] [PubMed]
- Bazylinski, D.A.; Williams, T.J.; Lefèvre, C.T.; Trubitsyn, D.; Fang, J.; Beveridge, T.J.; Moskowitz, B.M.; Ward, B.; Schübbe, S.; Dubbels, B.L.; et al. Magnetovibrio blakemorei gen. nov., sp. nov., a magnetotactic bacterium (Alphaproteobacteria: Rhodospirillaceae) isolated from a salt marsh. Int. J. Syst. Evol. Microbiol. 2013, 63, 1824–1833. [Google Scholar] [CrossRef] [Green Version]
- Sakaguchi, T.; Arakaki, A.; Matsunaga, T. Desulfovibrio magneticus sp nov., a novel sulfate-reducing bacterium that produces intracellular single-domain-sized magnetite particles. Int. J. Syst. Evol. Microbiol. 2002, 52, 215–221. [Google Scholar] [CrossRef] [Green Version]
- Lefèvre, C.T.; Trubitsyn, D.; Abreu, F.; Kolinko, S.; de Almeida, L.G.P.; de Vasconcelos, A.T.R.; Lins, U.; Schüler, D.; Ginet, N.; Pignol, D.; et al. Monophyletic origin of magnetotaxis and the first magnetosomes. Environ. Microbiol. 2013, 15, 2267–2274. [Google Scholar] [CrossRef]
- Chariaou, M.; Rahn-Lee, L.; Kind, J.; García-Rubio, I.; Komeili, A.; Gehring, A.U. Anisotropy of Bullet-Shaped Magnetite Nanoparticles in the Magnetotactic Bacteria Desulfovibrio magneticus sp. Strain RS-1. Biophys. J. 2015, 108, 1268–1274. [Google Scholar] [CrossRef] [Green Version]
- Descamps, E.C.T.; Monteil, C.L.; Menguy, N.; Ginet, N.; Pignol, D.; Bazylinski, D.A.; Lefèvre, C.T. Desulfamplus magnetovallimortis gen. Nov., sp. nov., a magnetotactic bacterium from a brackish desert spring able to biomineralize greigite and magnetite, that represents a novel lineage in the Desulfobacteraceae. Syst. Appl. Microbiol. 2017, 40, 280–289. [Google Scholar] [CrossRef]
- Araujo, A.C.V.; Morillo, V.; Cypriano, J.; Teixeira, L.C.R.S.; Leão, P.; Lyra, S.; de Almeida, L.G.; Bazylinski, D.A.; de Vasconcellos, A.T.R.; Abreu, F.; et al. Combined genomic and structural analyses of a cultured magnetotactic bacterium reveals its niche adaptation to a dynamic environment. BMC Genom. 2016, 17, 363–375. [Google Scholar] [CrossRef]
- Kolinko, S.; Jogler, C.; Katzmann, E.; Wanner, G.; Peplies, J.; Schüler, D. Single-cell analysis reveals a novel uncultivated magnetotactic bacterium within the candidate division OP3. Environ. Microbiol. 2012, 14, 1709–1721. [Google Scholar] [CrossRef]
- Kolinko, S.; Richter, M.; Glöckner, F.O.; Brachmann, A.; Schüler, D. Single-cell genomics of uncultivated deep-branching magnetotactic bacteria reveals a conserved set of magnetosome genes. Environ. Microbiol. 2015, 18, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Jogler, C.; Niebler, M.; Lin, W.; Kube, M.; Wanner, G.; Kolinko, S.; Stief, P.; Beck, A.J.; de Beer, D.; Petersen, N.; et al. Cultivation-independent characterization of “Candidatus Magnetobacterium bavaricum” via ultrastructural, geochemical, ecological and metagenomic methods. Environ. Microbiol. 2010, 12, 2466–2478. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Zhang, H.; Zhang, X.; Wang, Q.; Tian, J.; Li, Y.; Li, J. Rapid separation and immunoassay for low levels of Salmonella in foods using magnetosome-antibody complex and real-time fluorescence quantitative PCR. J. Sep. Sci. 2010, 33, 3437–3443. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Zhang, W.; Zhao, X.; Roberts, A.P.; Paterson, G.A.; Bazylinski, D.A.; Pan, Y. Genomic expansion of magnetotactic bacteria reveals an early common origin of magnetotaxis with lineage-specific evolution. ISME J. 2018, 12, 1508–1519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abreu, F.; Cantão, M.E.; Nicolás, M.F.; Barcellos, F.G.; Morillo, V.; Almeida, L.G.P.; do Nascimento, F.F.; Lefèvre, C.T.; Bazylinski, D.A.; de Vasconcelos, A.T.R.; et al. Common ancestry of iron oxide- and iron-sulfide-based biomineralization in magnetotactic bacteria. ISME J. 2011, 5, 1634–1640. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Katzmann, E.; Borg, S.; Schüler, D. The periplasmic nitrate reductase Nap is required for anaerobic growth and involved in redox control of magnetite biomineralization in Magnetospirillum gryphiswaldense. J. Bacteriol. 2012, 194, 4847–4856. [Google Scholar] [CrossRef] [Green Version]
- Popa, R.; Fang, W.; Nealson, K.H.; Souza-Egipsy, V.; Berquo, T.S.; Banerje, S.K.; Penn, L.R. Effect of oxidative stress on the growth of magnetic particles in Magnetospirillum magneticum. Int. Microbiol. 2009, 12, 49–57. [Google Scholar] [CrossRef]
- Moisescu, C.; Ardelean, I.I.; Benning, L.G. The effect and role of environmental conditions on magnetosome synthesis. Front. Microbiol. 2014, 5, 49. [Google Scholar] [CrossRef]
- Silva, K.T.; Leão, P.E.; Abreu, F.; López, J.A.; Gutarra, M.L.; Farina, M.; Bazylinski, D.A.; Freire, D.M.G.; Lins, U. Optimization of magnetosome production and growth by the magnetotactic vibrio Magnetovibrio blakemorei strain MV-1 through a statistics-based experimental design. Appl. Environ. Microbiol. 2013, 79, 2823–2827. [Google Scholar] [CrossRef]
- Mandernack, K.W.; Bazylinski, D.A.; Shanks, W.C.; Bullen, T.D. Oxygen and Iron Isotope Studies of Magnetite Produced by Magnetotactic Bacteria. Science 1999, 285, 1892–1896. [Google Scholar] [CrossRef]
- Bazylinski, D.A.; Frankel, R.B.; Jannasch, H.W. Anaerobic magnetite production by marine, magnetotactic bacterium. Nature 1988, 334, 518–519. [Google Scholar] [CrossRef] [Green Version]
- Simmons, S.L.; Sievert, S.M.; Frankel, R.B.; Bazylinski, D.A.; Edwards, K.J. Spatiotemporal Distribution of Marine Magnetotactic Bacteria in a Seasonally Stratified Coastal Salt Pond. Appl. Environ. Microbiol. 2004, 70, 6230–6239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frankel, R.B.; Williams, T.J.; Bazylinski, D.A. Magneto-Aerotaxis. In Magnetoreception and Magnetosomes in Bacteria, 1st ed.; Schüler, D., Ed.; Springer: Heidelberg, Germany, 2007; pp. 1–24. ISBN 978-3-540-37468-8. [Google Scholar]
- Lefèvre, C.T.; Wu, L.F. Evolution of the bacterial organelle responsible for magnetotaxis. Trends Microbiol. 2013, 21, 534–543. [Google Scholar] [CrossRef] [PubMed]
- Abreu, F.; Morillo, V.; Trubitsyn, D.; Bazylinski, D.A. Magnetotaxis in prokaryotes. In eLS; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2020; pp. 1–14. [Google Scholar] [CrossRef]
- Scheffel, A.; Gärdes, A.; Grünberg, K.; Wanner, G.; Schüler, D. The major magnetosome proteins MamGFDC are not essential for magnetite biomineralization in Magnetospirillum gryphiswaldense but regulate the size of magnetosome crystals. J. Bacteriol. 2008, 190, 377–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murat, D.; Quinlan, A.; Vali, H.; Komeili, A. Comprehensive genetic dissection of the magnetosome gene island reveals the step-wise assembly of a prokaryotic organelle. Proc. Natl. Acad. Sci. USA 2010, 107, 5593–5598. [Google Scholar] [CrossRef] [Green Version]
- Heyen, U.; Schüler, D. Growth and magnetosome formation by microaerophilic Magnetospirillum strains in an oxygen-controlled fermentor. Appl. Microbiol. Biotechnol. 2003, 61, 536–544. [Google Scholar] [CrossRef]
- Correa, T.; Godoy, M.G.; Bazylinski, D.A.; Abreu, F. Continuous Production of Biogenic Magnetite Nanoparticles by the Marine Bacterium Magnetovibrio blakemorei Strain MV-1T with a Nitrous Oxide Injection Strategy. Mar. Drugs 2022, 20, 724. [Google Scholar] [CrossRef]
- Yang, C.D.; Takeyama, H.; Tanaka, T.; Matsunaga, T. Effects of growth medium composition, iron sources and atmospheric oxygen concentrations on production of luciferase-bacterial magnetic particle complex by a recombinant Magnetospirillum magneticum AMB-1. Enzym. Microb. Technol. 2001, 29, 13–19. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.; Jiang, W.; Li, Y.; Li, J. Semicontinuous culture of Magnetospirillum gryphiswaldense MSR-1 cells in an autofermentor by nutrient-balanced and isosmotic feeding strategies. Appl. Environ. Microbiol. 2011, 77, 5851–5856. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Castané, A.; Li, H.; Thomas, O.R.T.; Overton, T.W. Development of a simple intensified fermentation strategy for growth of Magnetospirillum gryphiswaldense MSR-1: Physiological responses to changing environmental conditions. New Biotechnol. 2018, 46, 22–30. [Google Scholar] [CrossRef]
- Liu, Y.; Li, G.R.; Guo, F.F.; Jiang, W.; Li, Y.; Li, L.J. Large-scale production of magnetosomes by chemostat culture of Magnetospirillum gryphiswaldense at high cell density. Microb. Cell Factories 2010, 9, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Ding, Y.; Jiang, W.; Tian, J.; Li, Y.; Li, J. A mutation upstream of an ATPase gene significantly increases magnetosome production in Magnetospirillum gryphiswaldense. Appl. Microbiol. Biotechnol. 2008, 81, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.B.; Zhao, F.; Tang, T.; Jiang, W.; Tian, J.S.; Li, Y.; Li, J.L. High-yield growth and magnetosome formation by Magnetospirillum gryphiswaldense MSR-1 in an oxygen-controlled fermentor supplied solely with air. Appl. Microbiol. Biotechnol. 2008, 79, 389–397. [Google Scholar] [CrossRef] [PubMed]
- Berny, C.; Le Fèvre, R.; Guyot, F.; Blondeau, K.; Guizonne, C.; Rousseau, E.; Bayan, N.; Alphandéry, E. A Method for producing highly pure magnetosomes in large quantity for medical applications using Magnetospirillum gryphiswaldense MSR-1 magnetotactic bacteria amplified in minimal growth media. Front. Bioeng. Biotechnol. 2020, 8, 16. [Google Scholar] [CrossRef] [Green Version]
- Ke, L.; Chen, Y.; Liu, P.; Liu, S.; Wu, D.; Yuan, Y.; Wu, Y.; Gao, M. Characteristics and optimised fermentation of a novel magnetotactic bacterium, Magnetospirillum sp. ME-1. FEMS Microbiol. Lett. 2018, 365, fny052. [Google Scholar] [CrossRef]
- Ali, I.; Peng, C.; Khan, Z.M.; Naz, I. Yield cultivation of magnetotactic bacteria and magnetosomes: A review. J. Basic Microbiol. 2017, 57, 643–652. [Google Scholar] [CrossRef]
- Li, J.; Pan, Y. Environmental factors affect magnetite magnetosome synthesis in Magnetospirillum magneticum AMB-1: Implications for biologically controlled mineralization. Geomicrobiol. J. 2012, 29, 362–373. [Google Scholar] [CrossRef]
- Olszewska-Widdrat, A.; Schiro, G.; Reichel, V.E.; Faivre, D. Reducing conditions favor magnetosome production in Magnetospirillum magneticum AMB-1. Front. Microbiol. 2019, 10, 582. [Google Scholar] [CrossRef]
- World Health Organization. The International Pharmacopoeia, 9th ed.; World Health Organization: Geneva, Switzerland, 2019.
- Burda, C.; Chen, X.; Narayanan, R.; El-Sayed, M.A. Chemistry and properties of nanocrystals of different shapes. Chem. Rev. 2005, 105, 1025–1102. [Google Scholar] [CrossRef]
- Trahms, L. Biomedical applications of magnetic nanoparticles. In Lectures notes in physics: Coloidal Magnetic Fluids; Odenbach, S., Ed.; Springer: Heidelberg, Germany, 2009; Volume 763, pp. 327–358. ISBN 978-3-540-85387-9. [Google Scholar]
- Sparks, N.H.C.; Mann, S.; Bazylinski, D.A.; Lovely, D.R.; Jannasch, H.W.; Frankel, R.B. Structure and morphology of magnetite anaerobically-produced by a marine magnetotactic bacterium and a dissimilatory iron-reducing bacterium. Earth Planet. Sci. Lett. 1990, 98, 14–22. [Google Scholar] [CrossRef] [Green Version]
- Cypriano, J.; Werckmann, J.; Vargas, G.; dos Santos, A.L.; Silva, K.T.; Leão, P.; Almeida, F.P.; Bazylinski, D.A.; Farina, M.; Ulysses, L.; et al. Uptake and persistence of bacterial magnetite magnetosomes in a mammalian cell line: Implications for medical and biotechnological applications. PLoS ONE 2019, 14, e0215657. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, T.; Hirabe, H.; Takahashi, M.; Kuhara, M.; Takeyama, H.; Matsunaga, T. Magnetic cell separation using nano-sized bacterial magnetic particles with reconstructed magnetosome membrane. Biotechnol. Bioeng. 2008, 101, 470–477. [Google Scholar] [CrossRef] [PubMed]
- de Carvalho, J.C.; Medeiros, A.B.P.; Letti, L.A.J.; Kirnev, P.C.S.; Soccol, C.R. Cell disruption and isolation of intracellular products. In Current Developments in Biotechnology and Bioengineering: Production, Isolation and Purification of Industrial Products, 1st ed.; Pandey, A., Negi, S., Soccol, C.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 807–822. ISBN 978-0-444-63662-1. [Google Scholar]
- Xu, J.; Ma, S.; Zhang, W.; Jia, L.; Zheng, H.; Bo, P.; Bai, X.; Sun, H.; Qi, L.; Zhang, T.; et al. In vitro magnetosome remineralization for silver-magnetite hybrid magnetosome biosynthesis and used for healing of the infected wound. J. Nanobiotechnology 2022, 20, 364. [Google Scholar] [CrossRef]
- Mamat, U.; Wilke, K.; Bramhill, D.; Schromm, A.B.; Lindner, B.; Kohl, T.A.; Corchero, J.L.; Villaverde, A.; Schaffer, L.; Head, S.R.; et al. Detoxifying Escherichia coli for endotoxin-free production of recombinant proteins. Microb. Cell Factories 2015, 14, 57. [Google Scholar] [CrossRef] [Green Version]
- Abreu, F.; Leão, P.; Vargas, G.; Cypriano, J.; Figueiredo, V.; Enrich-Prast, A.; Bazylinski, D.A.; Lins, U. Culture-independent characterization of a novel magnetotactic member affiliated to the Beta class of the Proteobacteria phylum from an acidic lagoon. Environ. Microbiol. 2018, 20, 2615–2624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, F.; Liu, Y.; Chen, Y.; Tang, T.; Jiang, W.; Li, J. A novel rapid and continuous procedure for large-scale purification of magnetosomes from Magnetospirillum gryphiswaldense. Appl. Microbiol. Biotechnol. 2011, 90, 1277–1283. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.-S.; Wang, D.; Zhou, C.; Zhang, S. Preparation and anti-tumor efficiency evaluation of bacterial magnetosome -anti-4-1BB antibody complex: Bacterial magnetosome as antibody carriers isolated from Magnetospirillum gryphiswaldense. Biotechnol. Appl. Biochem. 2019, 66, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Fischer, A.; Schmitz, M.; Aichmayer, B.; Fratzl, P.; Faivre, D. Structural purity of magnetite nanoparticles in magnetotactic bacteria. J. R. Soc. Interface 2011, 8, 1011–1018. [Google Scholar] [CrossRef] [PubMed]
- Sangeetha, J.; Thomas, S.; Arutchelvi, J.; Doble, M.; Philip, J. Functionalization of Iron Oxide Nanoparticles with Biosurfactants and Biocompatibility Studies. J. Biomed. Nanotechnol. 2013, 9, 751–764. [Google Scholar] [CrossRef]
- Liu, Y.; Dai, Q.; Wang, S.; Deng, Q.; Wu, W.; Chen, A. Preparation and in vitro antitumor effects of cytosine arabinoside-loaded genipin-poly-L-glutamic acid-modified bacterial magnetosomes. Int. J. Nanomed. 2015, 10, 1387–1396. [Google Scholar] [CrossRef] [Green Version]
- Mandawala, C.; Chebbi, I.; Durand-Dubief, M.; Le Fèvre, R.; Hamdous, Y.; Guyot, F.; Alphandéry, E. Biocompatible and stable magnetosome minerals coated with poly-l-lysine, citric acid, oleic acid, and carboxy-methyl-dextran for application in the magnetic hyperthermia treatment of tumors. J. Mater. Chem. B 2017, 5, 7644–7660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, Q.; Ma, Y.; Wang, S.; Kankala, R.K.; Liu, Y. Investigation of Various Cross-Linking Methods for the Immobilization of Cytosine Arabinoside on Bacterial Magnetosomes. J. Nanomater. 2017, 2017, 6738484. [Google Scholar] [CrossRef] [Green Version]
- Muhamad, M.Z.; Koh, K.S.; Chong, V.H. Gluteraldehyde-induced colitis: A rare cause of lower gastrointestinal bleeding. Am. J. Emerg. Med. 2013, 32, 685.e1–685.e2. [Google Scholar] [CrossRef] [PubMed]
- Beauchamp, R.O.; St Clai, M.B.G.; Fennell, T.R.; Clarke, D.; Morgan, K.T.; Karl, F.W. A Critical Review of the Toxicology of Glutaraldehyde. Crit. Rev. Toxicol. 1992, 22, 143–174. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Gao, L.; Wang, B. ppiPre: Predicting protein-protein interactions by combining heterogeneous features. BMC Syst. Biol. 2013, 7 (Suppl. S2), S8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pujana, M.A.; Pérez-Álvarez, L.; Iturbe, L.C.C.; Katime, I. Water soluble folate-chitosan nanogels crosslinked by genipin. Carbohydr. Polym. 2014, 101, 113–120. [Google Scholar] [CrossRef]
- Correa, T.; Bazylinski, A.; Garcia, F.; Abreu, F. A rapid and simple preparation of amphotericin B-loaded bacterial magnetite nanoparticles. RSC Adv. 2021, 11, 28000–28007. [Google Scholar] [CrossRef]
- Mickoleit, F.; Lanzloth, C.; Schüler, D. A Versatile Toolkit for Controllable and Highly Selective Multifunctionalization of Bacterial Magnetic Nanoparticles. Small 2020, 16, 1906922. [Google Scholar] [CrossRef]
- Borg, S.; Hofmann, J.; Pollithy, A.; Lang, C.; Schüler, D. New Vectors for Chromosomal Integration Enable High-Level Constitutive or Inducible Magnetosome Expression of Fusion Proteins in Magnetospirillum gryphiswaldense. Appl. Environ. Microbiol. 2014, 80, 2609–2616. [Google Scholar] [CrossRef] [Green Version]
- Mickoleit, F.; Schüler, D. Generation of Multifunctional Magnetic Nanoparticles with Amplified Catalytic Activities by Genetic Expression of Enzyme Arrays on Bacterial Magnetosomes. Adv. Biosyst. 2017, 2, 1700109. [Google Scholar] [CrossRef]
- Sun, J.; Li, Y.; Liang, X.-J.; Wang, P.C. Bacterial Magnetosome: A Novel Biogenetic Magnetic Targeted Drug Carrier with Potential Multifunctions. J. Nanomater. 2011, 2011, 469031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, Z.; Yang, X.; Xu, J.; Lai, W.; Wang, Z.; Hu, Z.; Tian, J.; Geng, L.; Fang, Q. Tumor detection using magnetosome nanoparticles functionalized with a newly screened EGFR/HER2 targeting peptide. Biomaterials 2017, 115, 53–64. [Google Scholar] [CrossRef]
- Schwarz, S.; Fernandes, F.; Sanroman, L.; Hodenius, M.; Lang, C.; Himmelreich, U.; Schmitz-Rode, T.; Schueler, D.; Hoehn, M.; Zenke, M.; et al. Synthetic and biogenic magnetite nanoparticles for tracking of stem cells and dendritic cells. J. Magn. Magn. Mater. 2009, 321, 1533–1538. [Google Scholar] [CrossRef]
- Orlando, T.; Mannucci, S.; Fantechi, E.; Conti, G.; Tambalo, S.; Busato, A.; Innocenti, C.; Ghin, L.; Bassi, R.; Arosio, P.; et al. Characterization of magnetic nanoparticles from Magnetospirillum Gryphiswaldense as potential theranostics tools. Contrast Media Mol. Imaging 2015, 11, 139–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Usov, N.A.; Gubanova, E.M. Application of Magnetosomes in Magnetic Hyperthermia. Nanomaterials 2020, 10, 1320. [Google Scholar] [CrossRef] [PubMed]
- Ginet, N.; Pardoux, R.; Adryanczyk, G.; Garcia, D.; Brutesco, C.; Pignol, D. Single-Step Production of a Recyclable Nanobiocatalyst for Organophosphate Pesticides Biodegradation Using Functionalized Bacterial Magnetosomes. PLoS ONE 2011, 6, e21442. [Google Scholar] [CrossRef] [Green Version]
- Wacker, R.; Ceyhan, B.; Alhorn, P.; Schueler, D.; Lang, C.; Niemeyer, C.M. Magneto Immuno-PCR: A novel immunoassay based on biogenic magnetosome nanoparticles. Biochem. Biophys. Res. Commun. 2007, 357, 391–396. [Google Scholar] [CrossRef]
- Li, J.; Pan, Y.; Liu, Q.; Yu-Zhang, K.; Menguy, N.; Che, R.; Qin, H.; Lin, W.; Wu, W.; Petersen, N.; et al. Biomineralization, crystallography and magnetic properties of bullet-shaped magnetite magnetosomes in giant rod magnetotactic bacteria. Earth Planet Sci. Lett. 2010, 293, 368–376. [Google Scholar] [CrossRef]
- He, J.; Tian, J.; Xu, J.; Wang, K.; Li, J.; Gee, S.J.; Hammock, B.D.; Li, Q.X.; Xu, T. Strong and oriented conjugation of nanobodies onto magnetosomes for the development of a rapid immunomagnetic assay for the environmental detection of tetrabromobisphenol-A. Anal. Bioanal. Chem. 2018, 410, 6633–6642. [Google Scholar] [CrossRef]
- Ivanov, A.V.; Safenkova, I.V.; Zherdev, A.V.; Dzantiev, B.B. Recombinase polymerase amplification combined with a magnetic nanoparticle-based immunoassay for fluorometric determination of troponin T. Microchim. Acta 2019, 186, 549. [Google Scholar] [CrossRef]
- Tanaka, M.; Arakaki, A.; Staniland, S.S.; Matsunaga, T. Simultaneously Discrete Biomineralization of Magnetite and Tellurium Nanocrystals in Magnetotactic Bacteria. Appl. Environ. Microbiol. 2010, 76, 5526. [Google Scholar] [CrossRef] [Green Version]
- Arakaki, A.; Takeyama, H.; Tanaka, T.; Matsunaga, T. Cadmium Recovery by a Sulfate-Reducing Magnetotactic Bacterium, Desulfovibrio magneticus RS-1, Using Magnetic Separation. Appl. Biochem. Biotechnol. 2002, 98, 833–840. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Brown, R.; Hondow, N.; Arakaki, A.; Matsunaga, T.; Staniland, S. Highest levels of Cu, Mn and Co doped into nanomagnetic magnetosomes through optimized biomineralisation. J. Mater. Chem. 2012, 22, 11919. [Google Scholar] [CrossRef]
- Shimoshige, H.; Nakajima, Y.; Kobayashi, H.; Yanagisawa, K.; Nagaoka, Y.; Shimamura, S.; Mizuki, T.; Inoue, A.; Maekawa, T. Formation of Core-Shell Nanoparticles Composed of Magnetite and Samarium Oxide in Magnetospirillum magneticum strain RSS-1. PLoS ONE 2017, 12, e0170932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, M.; Knowles, W.; Brown, R.; Hondow, N.; Arakaki, A.; Baldwin, S.; Staniland, S.; Matsunaga, T. Biomagnetic Recovery and Bioaccumulation of Selenium Granules in Magnetotactic Bacteria. Appl. Environ. Microbiol. 2016, 82, 3886–3891. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Nakata, Y.; Mori, T.; Okamura, Y.; Miyasaka, H.; Takeyama, H.; Matsunaga, T. Development of a Cell Surface Display System in a Magnetotactic Bacterium, “Magnetospirillum magneticum” AMB-1. Appl. Environ. Microbiol. 2008, 74, 3342–3348. [Google Scholar] [CrossRef] [Green Version]
- Prozorov, T.; Palo, P.; Wang, L.; Nilsen-Hamilton, M.; Jones, D.; Orr, D.; Mallapragada, S.K.; Narasimhan, B.; Canfield, P.C.; Prozorov, R. Cobalt Ferrite Nanocrystals: Out-Performing Magnetotactic Bacteria. ACS Nano 2007, 1, 228–233. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, L.; Yi, Y.; Chen, C.; Wu, L.F.; Song, T. Killing of Staphylococcus aureus via magnetic hyperthermia mediated by magnetotactic bacteria. Appl. Environ. Microbiol. 2016, 82, 2219–2226. [Google Scholar] [CrossRef]
- Felfoul, O.; Mohammadi, M.; Taherkhani, S.; de Lanauze, D.; Xu, Y.Z.; Loghin, D.; Essa, S.; Jancik, S.; Houle, D.; LaFleur, M.; et al. Magneto-aerotactic bacteria deliver drug-containing nanoliposomes to tumour hypoxic regions. Nat. Nanotechnol. 2016, 11, 941–947. [Google Scholar] [CrossRef]
- Smit, B.A.; Van Zyl, E.; Joubert, J.J.; Meyer, W.; Prévéral, S.; Lefèvre, C.T.; Venter, S.N. Magnetotactic bacteria used to generate electricity based on Faraday’s law of electromagnetic induction. Lett. Appl. Microbiol. 2018, 66, 362–367. [Google Scholar] [CrossRef]
- Neter, E. Immunogenicity of Endotoxin. In Beneficial Effects of Endotoxins, 1st ed.; Nowotny, A., Ed.; Springer: New York, NY, USA, 1983; pp. 91–110. ISBN 978-1-4684-4364-6. [Google Scholar]
- Wang, X.; Quinn, P.J. Endotoxins: Lipopolysaccharides of Gram-Negative Bacteria. In Endotoxins: Structure, Function and Recognition, 1st ed.; Wang, X., Quinn, P.J., Eds.; Springer: Dordrecht, The Netherlands, 2010; Volume 53, pp. 3–25. ISBN 978-90-481-9078-2. [Google Scholar]
- Sun, J.B.; Duan, J.H.; Dai, S.L.; Ren, J.; Guo, L.; Jiang, W.; Li, Y. Preparation and anti-tumor efficiency evaluation of doxorubicin-loaded bacterial magnetosomes: Magnetic nanoparticles as drug carriers isolated from Magnetospirillum gryphiswaldense. Biotechnol. Bioeng. 2008, 101, 1313–1320. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Geng, Y.; Zhang, Y.; Wang, X.; Liu, J.; Basit, A.; Miao, T.; Liu, W.; Jiang, W. Bacterial magnetosomes loaded with doxorubicin and transferrin improve targeted therapy of hepatocellular carcinoma. Nanotheranostics 2019, 3, 284–298. [Google Scholar] [CrossRef] [PubMed]
- Hafsi, M.; Preveral, S.; Hoog, C.; Hérault, J.; Perrier, G.A.; Lefèvre, C.T.; Michel, H.; Pignol, D.; Doyen, J.; Pourcher, T.; et al. RGD-functionalized magnetosomes are efficient tumor radioenhancers for X-rays and protons. Nanomed. Nanotechnol. Biol. Med. 2020, 23, 102084. [Google Scholar] [CrossRef]
- Dai, Q.; Long, R.; Wang, S.; Kankala, R.K.; Wang, J.; Jiang, W.; Liu, Y. Bacterial magnetosomes as an efficient gene delivery platform for cancer theranostics. Microb. Cell Factories 2017, 16, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiesen, B.; Jordan, A. Clinical applications of magnetic nanoparticles for hyperthermia. Int. J. Hyperth. 2008, 24, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Alphandéry, E.; Faure, S.; Seksek, O.; Guyo, T.F.; Chebbi, I. Chains of magnetosomes extracted from AMB-1 magnetotactic bacteria for application in alternative magnetic field cancer therapy. ACS Nano 2011, 5, 6279–6296. [Google Scholar] [CrossRef] [PubMed]
- Alphandéry, E.; Guyot, F.; Chebbi, I. Preparation of chains of magnetosomes, isolated from Magnetospirillum magneticum strain AMB-1 magnetotactic bacteria, yielding efficient treatment of tumors using magnetic hyperthermia. Int. J. Pharm. 2012, 434, 444–452. [Google Scholar] [CrossRef] [Green Version]
- Alphandéry, E.; Idbaih, A.; Adam, C.; Delattre, J.Y.; Schmitt, C.; Guyot, F.; Chebbi, I. Development of non-pyrogenic magnetosome minerals coated with poly-l-lysine leading to full disappearance of intracranial U87-Luc glioblastoma in 100% of treated mice using magnetic hyperthermia. Biomaterials 2017, 141, 210–222. [Google Scholar] [CrossRef]
- Eifler, A.C.; Thaxton, C.S. Nanoparticle Therapeutics: FDA Approval, Clinical Trials, Regulatory Pathways, and Case Study. In Biomedical Nanotechnology: Methods in Molecular Biology; Hurst, S.J., Ed.; Humana Press: New York, NY, USA, 2011; ISBN 978-1-61779-052-2. [Google Scholar]
- Bobo, D.; Robinson, K.J.; Islam, J.; Thurecht, K.J.; Corrie, S.R. Nanoparticle-Based Medicines: A Review of FDA-Approved Materials and Clinical Trials to Date. Pharm. Res. 2016, 33, 2373–2387. [Google Scholar] [CrossRef]
- Soares, S.; Sousa, J.; Pais, A.; Vitorino, C. Nanomedicine: Principles, properties, and regulatory issues. Front. Chem. 2018, 6, 360. [Google Scholar] [CrossRef]
- Qi, L.; Lv, X.; Zhang, T.; Jia, P.; Yan, R.; Li, S.; Zou, R.; Xue, Y.; Dai, L. Cytotoxicity and genotoxicity of bacterial magnetosomes against human retinal pigment epithelium cells. Sci. Rep. 2016, 6, 26961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, L.; Wei, J.; Jianbo, S.; Guili, W.; Feng, G.; Ying, L. Purified and sterilized magnetosomes from Magnetospirillum gryphiswaldense MSR-1 were not toxic to mouse fibroblasts in vitro. Lett. Appl. Microbiol. 2007, 45, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Li, S.Y.; Yang, Y.; Zhao, F.M.; Huang, J.; Chang, J. Research on the structure and performance of bacterial magnetic nanoparticles. J. Biomater. Appl. 2008, 22, 433–448. [Google Scholar] [CrossRef] [PubMed]
- Revathy, T.; Jayasri, M.A.; Suthindhiran, K. Toxicity assessment of magnetosomes in different models. 3 Biotech 2017, 7, 126. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Tang, T.; Duan, J.; Xu, P.X.; Wang, Z.; Zhang, Y.; Wu, L.; Li, Y. Biocompatibility of bacterial magnetosomes: Acute toxicity, immunotoxicity and cytotoxicity. Nanotoxicology 2010, 4, 271–283. [Google Scholar] [CrossRef]
- Sun, J.B.; Wang, Z.L.; Duan, J.H.; Ren, J.; Yang, X.-D.; Dai, S.L.; Li, Y. Targeted distribution of bacterial magnetosomes isolated from magnetospirillum gryphiswaldense MSR-1 in healthy sprague-dawley rats. J. Nanosci. Nanotechnol. 2009, 9, 1881–1885. [Google Scholar] [CrossRef]
- Tang, T.; Zhang, L.; Gao, R.; Dai, Y.; Meng, F.; Li, Y. Fluorescence imaging and targeted distribution of bacterial magnetic particles in nude mice. Appl. Microbiol. Biotechnol. 2012, 94, 495–503. [Google Scholar] [CrossRef]
- Liu, R.T.; Liu, J.; Tong, J.Q.; Tang, T.; Kong, W.C.; Wang, X.W.; Li, Y.; Tang, J.T. Heating effect and biocompatibility of bacterial magnetosomes as potential materials used in magnetic fluid hyperthermia. Prog. Nat. Sci. Mater. Int. 2012, 22, 31–39. [Google Scholar] [CrossRef]
- Nan, X.; Lai, W.; Li, D.; Tian, J.; Hu, Z.; Fang, Q. Biocompatibility of Bacterial Magnetosomes as MRI Contrast Agent: A Long-Term In Vivo Follow-Up Study. Nanomaterials 2021, 11, 1235. [Google Scholar] [CrossRef]
- Cedervall, T.; Lynch, I.; Lindman, S.; Berggård, T.; Thulin, E.; Nilsson, H.; Dawson, K.A.; Linse, S. Understanding the nanoparticle-protein corona using methods to quantify exchange rates and affinities of proteins for nanoparticles. Proc. Natl. Acad. Sci. USA 2007, 104, 2050–2055. [Google Scholar] [CrossRef] [Green Version]
- Lynch, I.; Cedervall, T.; Lundqvist, M.; Cabaleiro-Lago, C.; Linse, S.; Dawson, K.A. The nanoparticle-protein complex as a biological entity; a complex fluids and surface science challenge for the 21st century. Adv. Colloid Interface Sci. 2007, 134–135, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Hadjidemetriou, M.; Kostarelos, K. Nanomedicine: Evolution of the nanoparticle corona. Nat. Nanotechnol. 2017, 12, 288–290. [Google Scholar] [CrossRef] [PubMed]
- Lesniak, A.; Fenaroli, F.; Monopoli, M.P.; Åberg, C.; Dawson, K.A.; Salvati, A. Effects of the presence or absence of a protein corona on silica nanoparticle uptake and impact on cells. ACS Nano 2012, 6, 5845–5857. [Google Scholar] [CrossRef] [PubMed]
- Sim, R.B.; Wallis, R. Surface properties: Immune attack on nanoparticles. Nat. Nanotechnol. 2011, 6, 80–81. [Google Scholar] [CrossRef]
- Tenzer, S.; Docter, D.; Kuharev, J.; Musyanovych, A.; Fetz, V.; Hecht, R.; Schlenk, F.; Fischer, D.; Kiouptsi, K.; Reinhard, T.C.; et al. Rapid formation of plasma protein corona critically affects nanoparticle pathophysiology. Nat. Nanotechnol. 2013, 8, 772–781. [Google Scholar] [CrossRef]
- Lai, W.; Li, D.; Wang, Q.; Nan, X.; Xiang, Z.; Ma, Y.; Liu, Y.; Chen, J.; Tian, J.; Fang, Q. A protein corona adsorbed to a bacterial magnetosome affects its cellular uptake. Int. J. Nanomed. 2020, 15, 1481–1498. [Google Scholar] [CrossRef] [Green Version]
- Desai, N. Challenges in development of nanoparticle-based therapeutics. AAPS J. 2012, 14, 282–295. [Google Scholar] [CrossRef] [Green Version]
- Ilinskaya, A.N.; Dobrovolskaia, M.A. Nanoparticles and the blood coagulation system. Part II: Safety concerns. Nanomedicine 2013, 8, 969–981. [Google Scholar] [CrossRef]
- Radomski, A.; Jurasz, P.; Alonso-Escolano, D.; Drews, M.; Morandi, M.; Malinski, T.; Radomski, M.W. Nanoparticle-induced platelet aggregation and vascular thrombosis. Br. J. Pharmacol. 2005, 146, 882–893. [Google Scholar] [CrossRef] [Green Version]
- Kaur, I.P.; Kakkar, V.; Deol, P.K.; Yadav, M.; Singh, M.; Sharma, I. Issues and concerns in nanotech product development and its commercialization. J. Control. Release 2014, 193, 51–62. [Google Scholar] [CrossRef]
- Landesman-Milo, D.; Peer, D. Transforming Nanomedicines from Lab Scale Production to Novel Clinical Modality. Bioconjugate Chem. 2016, 27, 855–862. [Google Scholar] [CrossRef] [PubMed]
- Weiner, S.; Dove, P.M. An Overview of Biomineralization Processes and the Problem of the Vital Effect. Rev. Mineral. Geochem. 2003, 54, 1–29. [Google Scholar] [CrossRef]
- Faivre, D.; Godec, T.U. From bacteria to mollusks: The principles underlying the biomineralization of iron oxide materials. Angew. Chem. Int. Ed. 2015, 54, 4728–4747. [Google Scholar] [CrossRef] [PubMed]
- Agrahari, V.; Hiremath, P. Challenges associated and approaches for successful translation of nanomedicines into commercial products. Nanomedicine 2017, 12, 819–823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muthu, M.S.; Wilson, B. Challenges posed by the scale-up of nanomedicines. Nanomedicine 2012, 7, 307–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.; Hwang, Y.S. Patent keyword network analysis for improving technology development efficiency. Technol. Forecast. Soc. Chang. 2014, 83, 170–182. [Google Scholar] [CrossRef]
- Yanow, S.K.; Good, M.F. Nonessential research in the new normal: The impact of COVID-19. Am. J. Trop. Med. Hyg. 2020, 102, 1164. [Google Scholar] [CrossRef] [Green Version]
- Chen, L. Do patent citations indicate knowledge linkage? The evidence from text similarities between patents and their citations. J. Informetr. 2017, 11, 63–79. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, X.; Xu, S. Patent co-citation networks of Fortune 500 companies. Scientometris 2011, 88, 761–770. [Google Scholar] [CrossRef]
- Bell, C.B.; Bazarov, A. Host Cells with Artificial Endosymbionts. U.S. Patent 2013106814-A1, 14 January 2013. [Google Scholar]
- Abreu, F.; Correa, T.; Bazilinski, D.A.; Torres-Pinto, F. Processo Para Produção Contínua de Magnetossomos Através do Cultivo de Bactérias Magnetotáticas em Biorreator. Brazilian Patent BR1020200158317, 3 August 2020. [Google Scholar]
- Li, W.; Jiang, X.; Yang, H.; Liu, Q.; Sun, Y.; Cao, Y.; Zhang, Y.; Zhang, A. Method of Repairing to Improve Remediation Effect of Heavy Metal Polluted Soil Comprises e.g., Stripping Soil on Surface of Soil Polluted by Heavy Metals, Stacking in Remediation Tank, and Mixing with Magnetotactic Bacteria Liquid Medium. China Patent 111570501-A, 16 August 2020. [Google Scholar]
- Xu, Q. Purifying Domestic Sewage by Crushing Domestic Sewage, Passing Sewage through Grid Filter, Fermenting Sewage, Passing into Biological Water Pond, Irradiating Sewage, Adding Flocculant, Adding Sterilant and Purifying Sewage. China Patent 108409056-A, 4 December 2018. [Google Scholar]
- Dai, L.; Si, C.; Liu, R. Preparing Lignin Nanoparticles Used in Field of e.g. Food by Dissolving Lignin, Adding Non-Solvent, Purifying, Adding Soluble Medicine, Purifying Adding Non-Solvent, Purifying and Drying to Obtain Magnetic Drug-Loaded Lignin Nanoparticles. China Patent 107693506-A, 4 December 2018. [Google Scholar]
- Alphandéry, E. Magnetic Nanoparticles Sequentially Irradiated by Laser Radiation for Medical or Biological or Cosmetic Applications. France Patent 3569247-A1, 15 May 2019. [Google Scholar]
- Abreu, F.; Morillo, V.; Nascimento, F.; Werneck, C.; Cantão, M.E.; Ciapina, L.P.; de Almeida, L.G.P.; Lefèvre, C.T.; Bazylinski, D.A.; de Vasconcelos, A.T.R.; et al. Deciphering unusual uncultured magnetotactic multicellular prokaryotes through genomics. ISME J. 2014, 8, 1055–1068. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species/Strain | Process Conduction | Production (mg/L) | Productivity (mg/L/day) | Fe Source | Fe Concentration (µM) | Fe Feeding | Reference |
|---|---|---|---|---|---|---|---|
| Mv. blakemorei/MV-1T | FB | 26 | 3.2 | Ferrous sulfate | 100 | Yes | [50] |
| Mv. blakemorei/MV-1T | SB | 22.4 | 5.6 | Ferrous sulfate | 100 | No | [50] |
| Mv. blakemorei/MV-1T | FB | 24.5 | 16.8 | Ferrous sulfate | 100 | Yes | [60] |
| Mv. blakemorei/MV-1T | C | 27.1 | 22.7 | Ferrous sulfate | 100 | Yes | [60] |
| Ms. magneticum/AMB-1 | FB | 9 | 3.7 | Ferrous sulfate | 33 | Yes | [61] |
| Ms. gryphiswaldense/MSR-1 | FB | 225.53 | 112.77 | Ferric chloride | 71.3 | Yes | [62] |
| Ms. gryphiswaldense/MSR-1 | SC | 168.3 | 83.5 | Ferric chloride | 101.2 | Yes | [62] |
| Ms. gryphiswaldense/MSR-1 | FB | 139 | 47 | Ferric citrate | 100 | Yes | [63] |
| Ms. gryphiswaldense/MSR-1 | FB | 83.23 | 55.49 | Ferric citrate | 100 | Yes | [64] |
| Ms. gryphiswaldense/MSR-1 | FB | 58.4 | -- | Ferric citrate | 60 | Yes | [65] |
| Ms. gryphiswaldense/MSR-1 | FB | 41.7 | 16.7 | Ferric citrate | 60 | Yes | [66] |
| Ms. gryphiswaldense/MSR-1 | FB | 8–10 | 3.8–4.8 | Ferric chloride | -- | Yes | [67] |
| Ms. gryphiswaldense/MSR-1 | SB | 7.9 | 6.3 | Ferric citrate | 100–150 | No | [59] |
| Ms. sp./ME-1 | FB | 120 | 58.7 | Ferric citrate | 500 | Yes | [68] |
| IPC Codes | Patents | Description |
|---|---|---|
| C12N-001/20 | 22 | Bacteria, Culture media therefor |
| A61P-035/00 | 21 | Antineoplastic agents |
| A61K-041/00 | 15 | Medicinal preparations obtained by treating materials with wave energy or particle radiation |
| A61K-049/00 | 14 | Preparations for testing in vitro |
| A61K-049/18 | 12 | Preparations for testing in vitro, characterized by a special physical form, e.g., emulsions, microcapsules, liposomes |
| A61K-035/74 | 11 | Medicinal preparations containing materials or reaction products thereof with undetermined constitution, Bacteria |
| C12N-001/21 | 10 | Microorganisms, e.g., protozoa; modified by introduction of foreign genetic material |
| C12P-003/00 | 10 | Preparations of elements or inorganic compounds except carbon dioxide |
| B82Y-005/00 | 10 | Nanobiotechnology- or Nanomedicine, e.g., protein engineering or drug delivery |
| C12M-001/42 | 10 | Apparatus for the treatment of microorganisms or enzymes with electrical or wave energy, e.g., magnetism, sonic wave |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Souza Cabral, A.; Verdan, M.; Presciliano, R.; Silveira, F.; Correa, T.; Abreu, F. Large-Scale Cultivation of Magnetotactic Bacteria and the Optimism for Sustainable and Cheap Approaches in Nanotechnology. Mar. Drugs 2023, 21, 60. https://doi.org/10.3390/md21020060
de Souza Cabral A, Verdan M, Presciliano R, Silveira F, Correa T, Abreu F. Large-Scale Cultivation of Magnetotactic Bacteria and the Optimism for Sustainable and Cheap Approaches in Nanotechnology. Marine Drugs. 2023; 21(2):60. https://doi.org/10.3390/md21020060
Chicago/Turabian Stylede Souza Cabral, Anderson, Mariana Verdan, Rogerio Presciliano, Felipe Silveira, Tarcisio Correa, and Fernanda Abreu. 2023. "Large-Scale Cultivation of Magnetotactic Bacteria and the Optimism for Sustainable and Cheap Approaches in Nanotechnology" Marine Drugs 21, no. 2: 60. https://doi.org/10.3390/md21020060
APA Stylede Souza Cabral, A., Verdan, M., Presciliano, R., Silveira, F., Correa, T., & Abreu, F. (2023). Large-Scale Cultivation of Magnetotactic Bacteria and the Optimism for Sustainable and Cheap Approaches in Nanotechnology. Marine Drugs, 21(2), 60. https://doi.org/10.3390/md21020060