
Structure and Composition of the Cuticle of the Goose Barnacle Pollicipes pollicipes: A Flexible Composite Biomaterial


 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Characterization of Tergum Plate
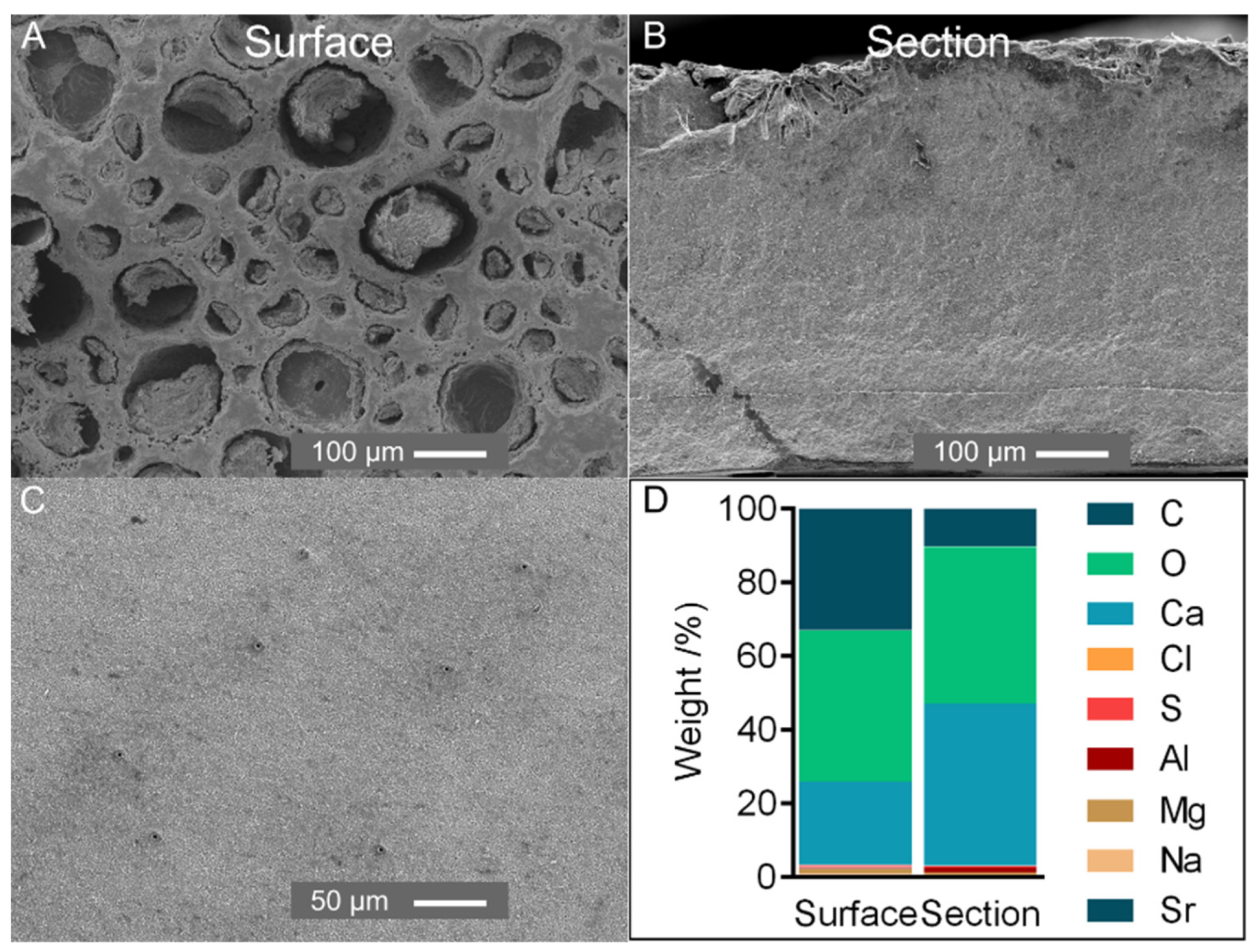
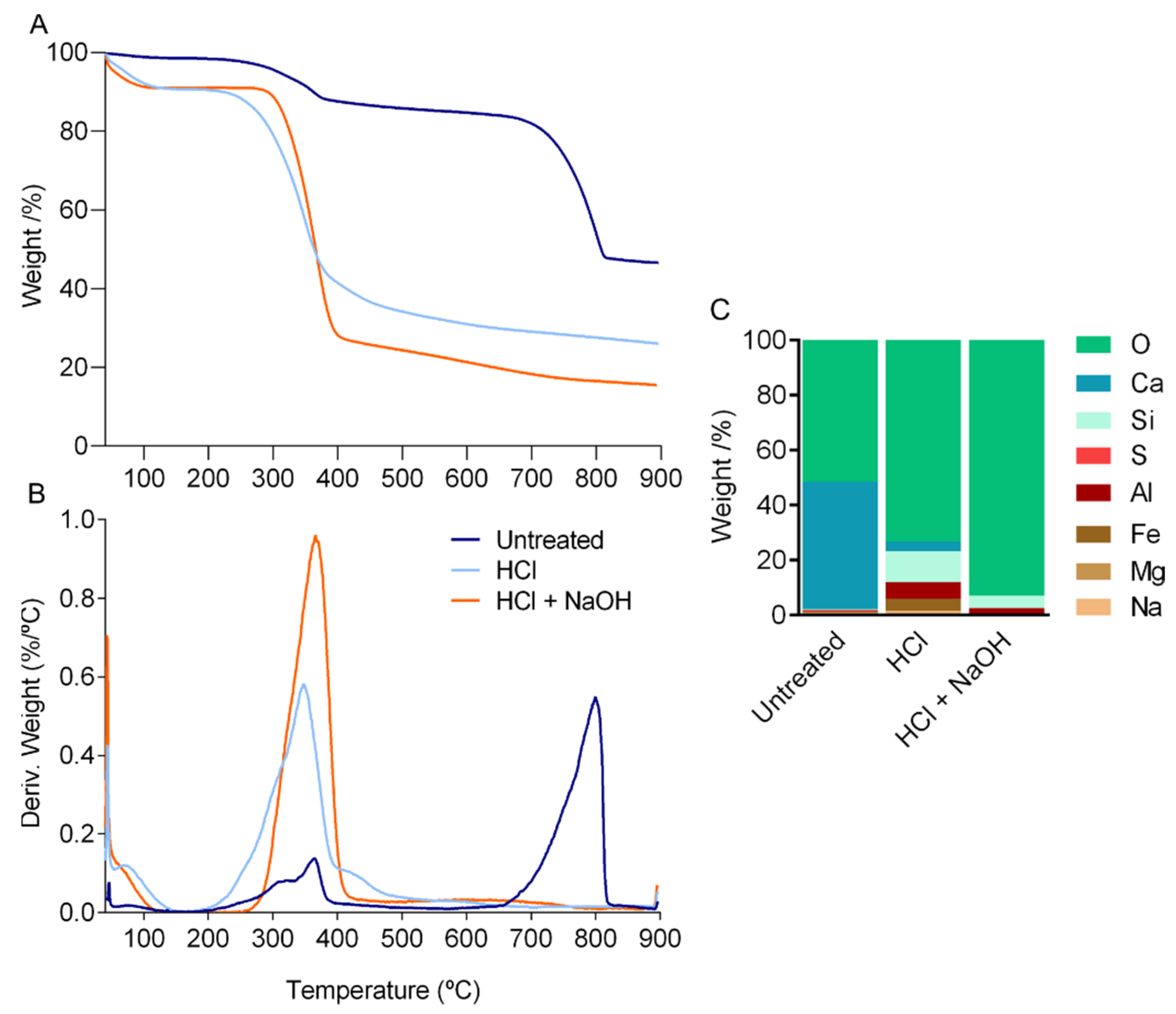
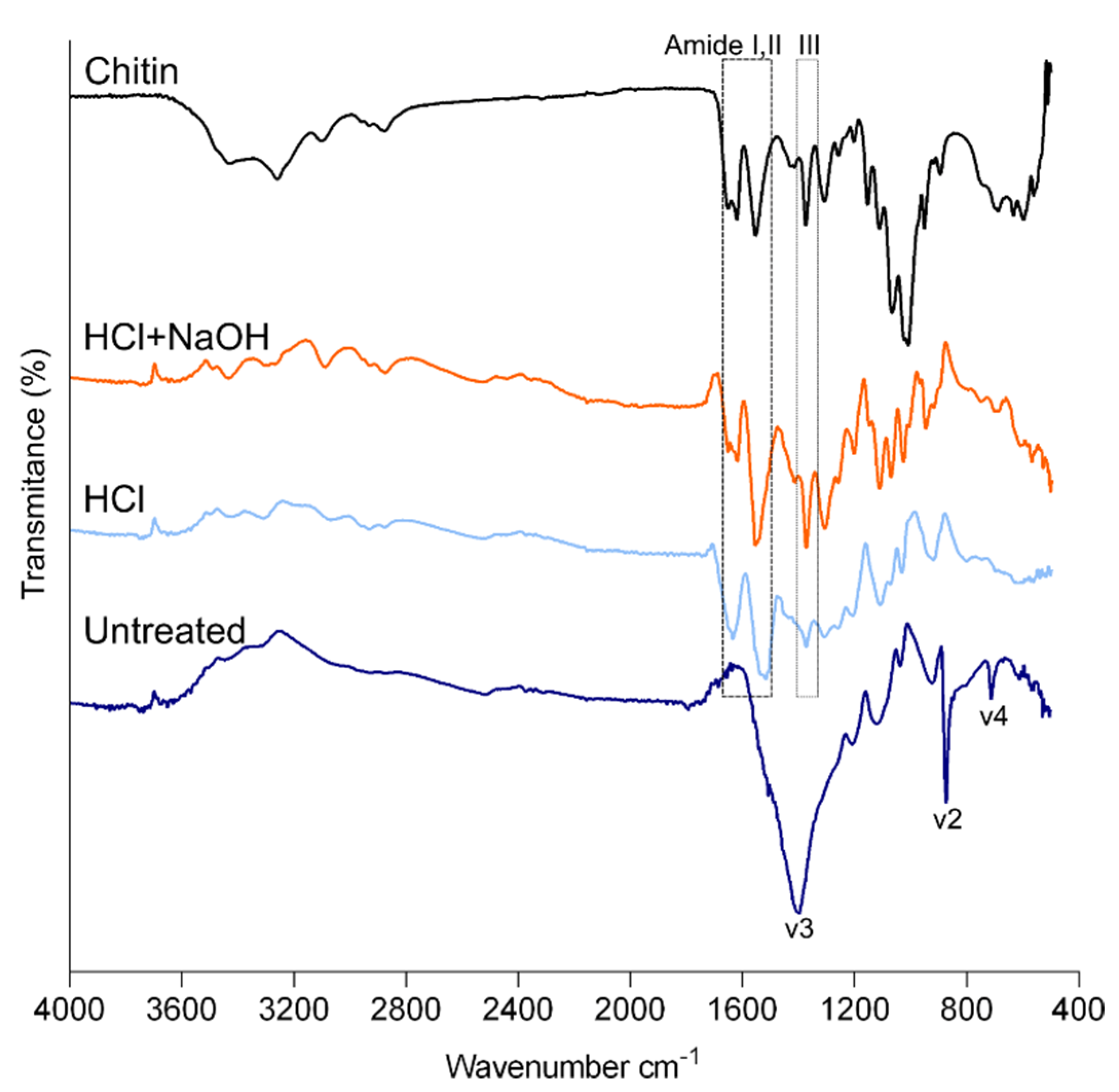
2.2. Composition of the Cuticle
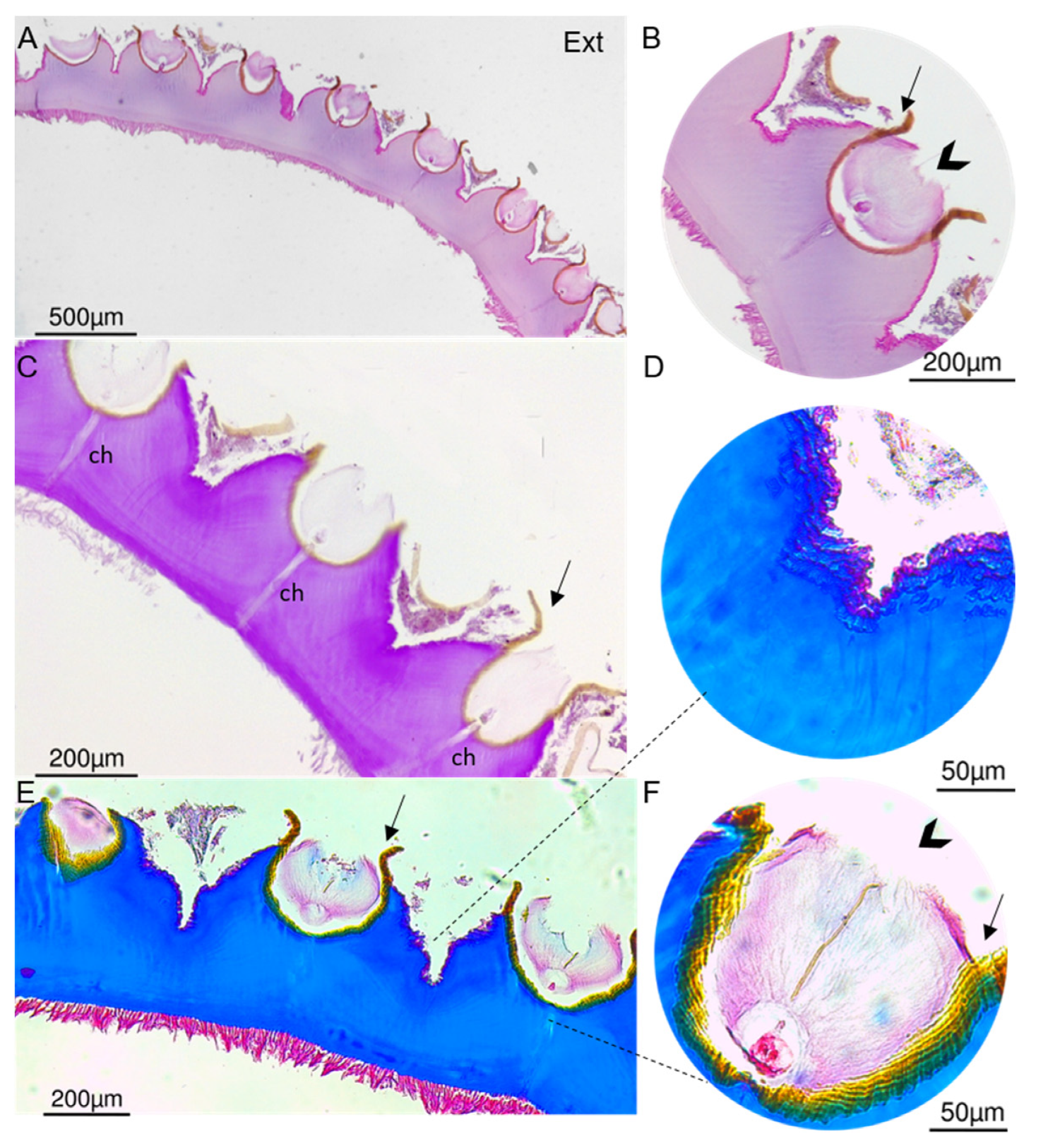
2.3. Organization and General Structure of the Cuticle
2.4. Fine Structure of the Cuticle
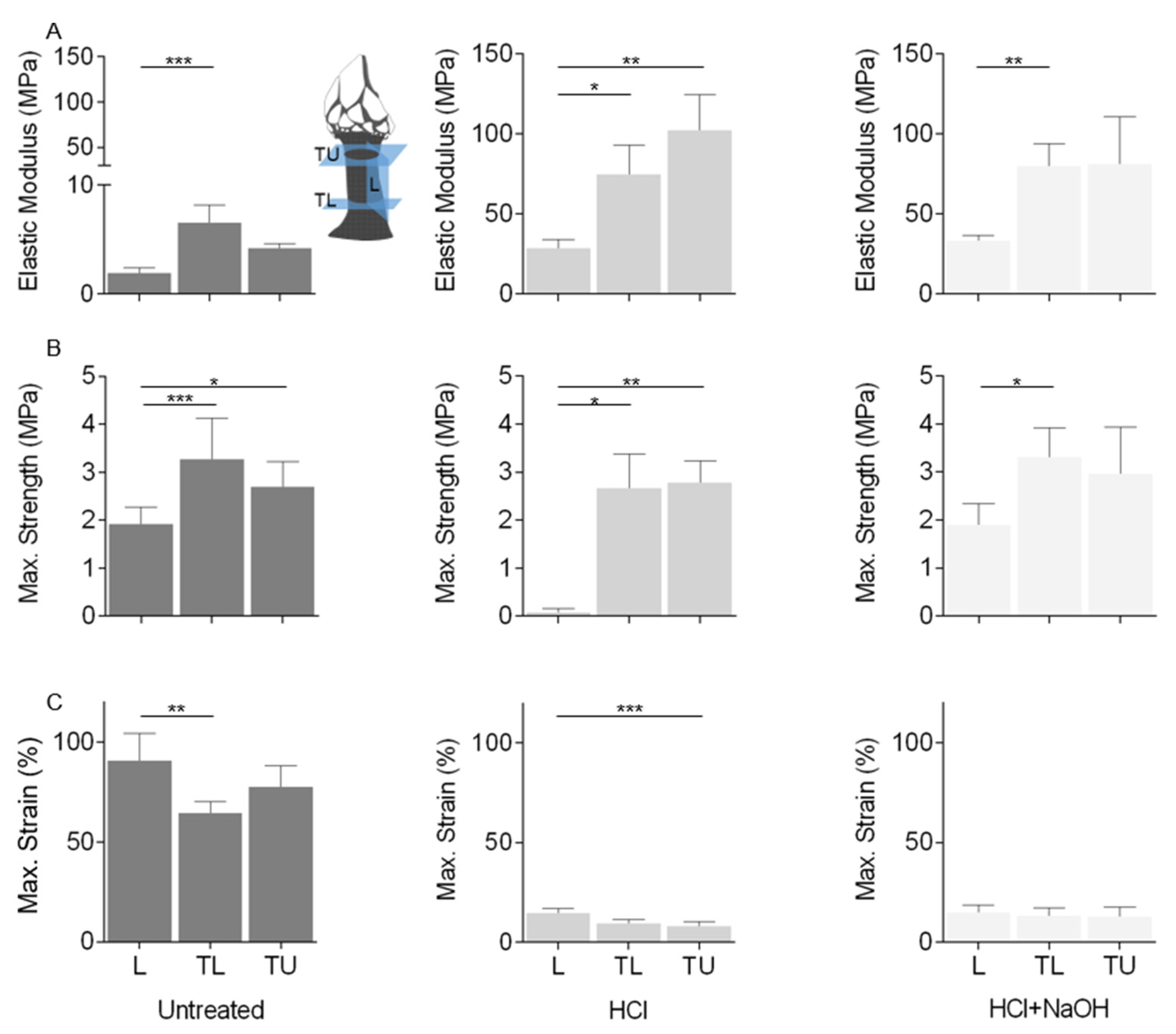
2.5. Mechanical Properties
2.6. Flexible Cuticles as Biomimetic Models for Tissue Engineering and Regenerative Medicine Applications
3. Material and Methods
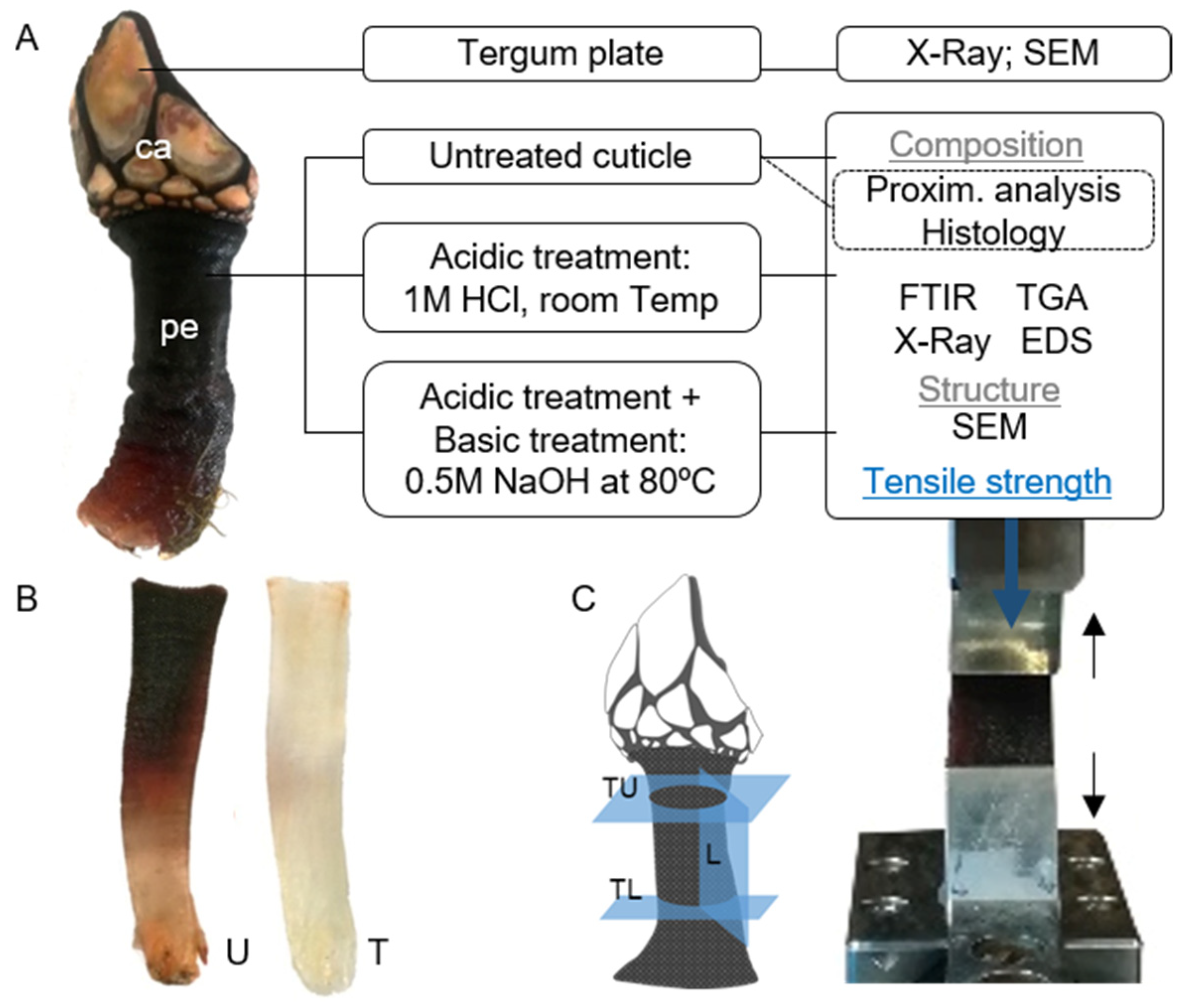
3.1. Sample Collection and Preparation
3.2. Plates Characterization
3.3. Cuticle Characterization
3.3.1. Proximate Composition and Estimation of Chitin Content
3.3.2. Thermal Gravimetric Analysis (TGA) and EDS
3.3.3. Attenuated Total Reflection–Fourier Transform Infrared (ATR-FTIR) Spectroscopy
3.3.4. X-Ray Diffraction (XRD)
3.3.5. Histology
3.3.6. Environmental Scanning Electron Microscopy (ESEM) and Scanning Electron Microscopy (SEM)
3.3.7. Mechanical Testing
3.3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Ehrlich, H.; Krautter, M.; Hanke, T.; Simon, P.; Knieb, C.; Heinemann, S.; Worch, H. First Evidence of the Presence of Chitin in Skeletons of Marine Sponges. Part II. Glass Sponges (Hexactinellida: Porifera). J. Exp. Zool. Part B Mol. Dev. Evol. 2007, 308B, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Nowacki, K.; Galiński, M.; Fursov, A.; Voronkina, A.; Meissner, H.; Petrenko, I.; Stelling, A.L.; Ehrlich, H. Electrolysis as a Universal Approach for Isolation of Diverse Chitin Scaffolds from Selected Marine Demosponges. Mar. Drugs 2022, 20, 665. [Google Scholar] [CrossRef] [PubMed]
- Juárez-de la Rosa, B.A.; Quintana, P.; Ardisson, P.-L.; Yáñez-Limón, J.M.; Alvarado-Gil, J.J. Effects of Thermal Treatments on the Structure of Two Black Coral Species Chitinous Exoskeleton. J. Mater. Sci. 2012, 47, 990–998. [Google Scholar] [CrossRef]
- Nowacki, K.; Stępniak, I.; Langer, E.; Tsurkan, M.; Wysokowski, M.; Petrenko, I.; Khrunyk, Y.; Fursov, A.; Bo, M.; Bavestrello, G.; et al. Electrochemical Approach for Isolation of Chitin from the Skeleton of the Black Coral Cirrhipathes Sp. (Antipatharia). Mar. Drugs 2020, 18, 297. [Google Scholar] [CrossRef]
- Matias, R.S.; Gregory, S.; Ceia, F.R.; Baeta, A.; Seco, J.; Rocha, M.S.; Fernandes, E.M.; Reis, R.L.; Silva, T.H.; Pereira, E.; et al. Show Your Beaks and We Tell You What You Eat: Different Ecology in Sympatric Antarctic Benthic Octopods under a Climate Change Context. Mar. Environ. Res. 2019, 150, 104757. [Google Scholar] [CrossRef]
- Bentov, S.; Abehsera, S. The Mineralized Exoskeletons of Crustaceans. In Extracellular Composite Matrices in Arthropods; Cohen, E., Moussian, B., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 137–263. ISBN 9783319407401. [Google Scholar]
- Ehrlich, H. Chitin and Collagen as Universal and Alternative Templates in Biomineralization. Int. Geol. Rev. 2010, 52, 661–699. [Google Scholar] [CrossRef]
- Boßelmann, F.; Romano, P.; Fabritius, H.; Raabe, D.; Epple, M. The Composition of the Exoskeleton of Two Crustacea: The American Lobster Homarus Americanus and the Edible Crab Cancer Pagurus. Thermochim. Acta 2007, 463, 65–68. [Google Scholar] [CrossRef]
- Watling, L.; Thiel, M. Functional Morphology and Diversity; The Natural History of Crustacea; Oxford University Press: Oxford, UK, 2012; ISBN 0199875456. [Google Scholar]
- Fabritius, H.O.; Ziegler, A.; Friák, M.; Nikolov, S.; Huber, J.; Seidl, B.H.M.; Ruangchai, S.; Alagboso, F.I.; Karsten, S.; Lu, J.; et al. Functional Adaptation of Crustacean Exoskeletal Elements through Structural and Compositional Diversity: A Combined Experimental and Theoretical Study. Bioinspiration Biomim. 2016, 11, 055006. [Google Scholar] [CrossRef]
- Mohan, K.; Muralisankar, T.; Jayakumar, R.; Rajeevgandhi, C. A Study on Structural Comparisons of α-Chitin Extracted from Marine Crustacean Shell Waste. Carbohydr. Polym. Technol. Appl. 2021, 2, 100037. [Google Scholar] [CrossRef]
- Wu, J.; Qin, Z.; Qu, L.; Zhang, H.; Deng, F.; Guo, M. Natural Hydrogel in American Lobster: A Soft Armor with High Toughness and Strength. Acta Biomater. 2019, 88, 102–110. [Google Scholar] [CrossRef]
- Weaver, J.C.; Milliron, G.W.; Miserez, A.; Evans-Lutterodt, K.; Herrera, S.; Gallana, I.; Mershon, W.J.; Swanson, B.; Zavattieri, P.; DiMasi, E.; et al. The Stomatopod Dactyl Club: A Formidable Damage-Tolerant Biological Hammer. Science 2012, 336, 1275–1280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tadayon, M.; Amini, S.; Masic, A.; Miserez, A. The Mantis Shrimp Saddle: A Biological Spring Combining Stiffness and Flexibility. Adv. Funct. Mater. 2015, 25, 6437–6447. [Google Scholar] [CrossRef]
- Yang, T.L. Chitin-Based Materials in Tissue Engineering: Applications in Soft Tissue and Epithelial Organ. Int. J. Mol. Sci. 2011, 12, 1936–1963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kertmen, A.; Ehrlich, H. Patentology of Chitinous Biomaterials. Part I: Chitin. Carbohydr. Polym. 2022, 282, 119102. [Google Scholar] [CrossRef]
- Reys, L.L.; Silva, S.S.; Oliveira, C.; Lopez-Cebral, R.; Neves, N.M.; Martins, A.; Oliveira, J.M.; Silva, T.H.; Reis, R.L. Marine-origin Polysaccharides for Tissue Engineering and Regenerative Medicine. Encycl. Mar. Biotechnol. 2020, 4, 2619–2650. [Google Scholar] [CrossRef]
- Mutsenko, V.; Gryshkov, O.; Rogulska, O.; Lode, A.; Petrenko, A.Y.; Gelinsky, M.; Glasmacher, B.; Ehrlich, H. Chitinous Scaffolds from Marine Sponges for Tissue Engineering. In Marine-Derived Biomaterials for Tissue Engineering Applications; Springer: Singapore, 2019. [Google Scholar]
- Ngoc-Hai, C.V.; Hong-My, D.D.; Bach Phan, T.; Ngoc, Q.T.; Van, T.V.; Nguyen, T.H. Simple Fabrication of a Chitin Wound Healing Membrane from Soft-Shell Crab Carapace. Mater. Lett. 2021, 297, 129995. [Google Scholar] [CrossRef]
- Liang, C.; Li, Y.; Liu, Z.; Wu, W.; Hu, B. Protein Aggregation Formed by Recombinant Cp19k Homologue of Balanus Albicostatus Combined with an 18 KDa N-Terminus Encoded by PET-32a(+) Plasmid Having Adhesion Strength Comparable to Several Commercial Glues. PLoS ONE 2015, 10, e0136493. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, R.L.; Coleman, M.; Davies, P.; North, L.; Pope, E.C.; Pleydell-Pearce, C.; Harris, W.; Johnston, R. Macro-to-Nano Scale Investigation of Wall-Plate Joints in the Acorn Barnacle Semibalanus Balanoides: Correlative Imaging, Biological Form and Function, and Bioinspiration. J. R. Soc. Interface 2019, 16, 20190218. [Google Scholar] [CrossRef] [Green Version]
- Rocha, M.; Antas, P.; Castro, L.F.C.; Campos, A.; Vasconcelos, V.; Pereira, F.; Cunha, I. Comparative Analysis of the Adhesive Proteins of the Adult Stalked Goose Barnacle Pollicipes Pollicipes (Cirripedia: Pedunculata). Mar. Biotechnol. 2019, 21, 38–51. [Google Scholar] [CrossRef]
- Jacinto, D.; Penteado, N.; Pereira, D.; Sousa, A.; Cruz, T. Variación de La Tasa de Crecimiento Del Percebe Pollicipes Pollicipes (Crustacea: Cirripedia) Utilizando Calceína Como Marcador Químico. Sci. Mar. 2015, 79, 117–123. [Google Scholar] [CrossRef]
- Buckeridge, J.S.; Reeves, J.M. Some Insights into How Barnacles Survive as Sessile Organisms. Integr. Zool. 2009, 4, 395–401. [Google Scholar] [CrossRef]
- Thomas, H.J. Tegumental Glands in the Cirripedia Thoracica. J. Cell Sci. 1944, s2–s84, 257–282. [Google Scholar] [CrossRef]
- Mahmoud, M.F. The Structure of the Outer Integument of Some Pedunculate Cirripedes (Ba Rnacles). Proc. Egypt. Acad. Sci. 1959, 14, 61–69. [Google Scholar]
- Wyckoff, R.W.G. The Crystal Structures of Some Carbonates of the Calcite Group. Am. J. Sci. 1920, s4–s50, 317–360. [Google Scholar] [CrossRef]
- Brusca, R.C.; Brusca, G.J. Invertebrates, 2nd ed.; Sinauer Associates: Sunderland, MA, USA, 2003. [Google Scholar]
- Checa, A.G.; González-Segura, A.; Rodríguez-Navarro, A.B.; Lagos, N.A. Microstructure and Crystallography of the Wall Plates of the Giant Barnacle Austromegabalanus Psittacus: A Material Organized by Crystal Growth. J. R. Soc. Interface 2020, 17, 20190743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalifa, G.M.; Weiner, S.; Addadi, L. Mineral and Matrix Components of the Operculum and Shell of the Barnacle Balanus Amphitrite: Calcite Crystal Growth in a Hydrogel. Cryst. Growth Des. 2011, 11, 5122–5130. [Google Scholar] [CrossRef]
- Barnes, M. Pedunculate Cirripedes of the Genus Pollicipes. Oceanogr. Mar. Biol. Annu. Rev. 1996, 34, 303–394. [Google Scholar]
- Younes, I.; Rinaudo, M. Chitin and Chitosan Preparation from Marine Sources. Structure, Properties and Applications. Mar. Drugs 2015, 13, 1133–1174. [Google Scholar] [CrossRef] [Green Version]
- Pires, C.; Marques, A.; Carvalho, M.; Batista, I. Chemical Characterization of Cancer Pagurus, Maja Squinado, NecoraPuber and Carcinus Maenas Shells. Poultry, Fish. Wildl. Sci. 2017, 5, 181. [Google Scholar]
- Kaya, M.; Lelešius, E.; Nagrockaite, R.; Sargin, I.; Arslan, G.; Mol, A.; Baran, T.; Can, E.; Bitim, B. Differentiations of Chitin Content and Surface Morphologies of Chitins Extracted from Male and Female Grasshopper Species. PLoS ONE 2015, 10, e0115531. [Google Scholar] [CrossRef] [Green Version]
- Kaya, M.; Tozak, K.O.; Baran, T.; Sezen, G.; Sargin, I. Natural Porous and Nano Fiber Chitin Structure from Gammarus Argeus (Gammaridea Crustacea). Excli J. 2013, 12, 503–510. [Google Scholar] [PubMed]
- Romano, P.; Fabritius, H.; Raabe, D. The Exoskeleton of the Lobster Homarus Americanus as an Example of a Smart Anisotropic Biological Material. Acta Biomater. 2007, 3, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Neues, F.; Ziegler, A.; Epple, M. The Composition of the Mineralized Cuticle in Marine and Terrestrial Isopods: A Comparative Study. CrystEngComm 2007, 9, 1245–1251. [Google Scholar] [CrossRef]
- Gbenebor, O.P.; Adeosun, S.O.; Lawal, G.I.; Jun, S.; Olaleye, S.A. Acetylation, Crystalline and Morphological Properties of Structural Polysaccharide from Shrimp Exoskeleton. Eng. Sci. Technol. Int. J. 2017, 20, 1155–1165. [Google Scholar] [CrossRef]
- Sajomsang, W.; Gonil, P. Preparation and Characterization of α-Chitin from Cicada Sloughs. Mater. Sci. Eng. C 2010, 30, 357–363. [Google Scholar] [CrossRef]
- Kaya, M.; Baublys, V.; Can, E.; Šatkauskienė, I.; Bitim, B.; Tubelytė, V.; Baran, T. Comparison of Physicochemical Properties of Chitins Isolated from an Insect (Melolontha Melolontha) and a Crustacean Species (Oniscus Asellus). Zoomorphology 2014, 133, 285–293. [Google Scholar] [CrossRef]
- Machałowski, T.; Wysokowski, M.; Tsurkan, M.V.; Galli, R.; Schimpf, C.; Rafaja, D.; Brendler, E.; Viehweger, C.; Żółtowska-Aksamitowska, S.; Petrenko, I.; et al. Spider Chitin: An Ultrafast Microwave-Assisted Method for Chitin Isolation from Caribena Versicolor Spider Molt Cuticle. Molecules 2019, 24, 3736. [Google Scholar] [CrossRef] [Green Version]
- Nardone, J.A.; Patel, S.; Siegel, K.R.; Tedesco, D.; McNicholl, C.G.; O’Malley, J.; Herrick, J.; Metzler, R.A.; Orihuela, B.; Rittschof, D.; et al. Assessing the Impacts of Ocean Acidification on Adhesion and Shell Formation in the Barnacle Amphibalanus Amphitrite. Front. Mar. Sci. 2018, 5, 369. [Google Scholar] [CrossRef] [Green Version]
- Kaya, M.; Mujtaba, M.; Ehrlich, H.; Salaberria, A.M.; Baran, T.; Amemiya, C.T.; Galli, R.; Akyuz, L.; Sargin, I.; Labidi, J. On Chemistry of γ-Chitin. Carbohydr. Polym. 2017, 176, 177–186. [Google Scholar] [CrossRef]
- Tsurkan, M.V.; Voronkina, A.; Khrunyk, Y.; Wysokowski, M.; Petrenko, I.; Ehrlich, H. Progress in Chitin Analytics. Carbo-Hydr. Polym. 2021, 252, 117204. [Google Scholar] [CrossRef]
- Franco, S.C. Aquaculture of Stalked Barnacles (Pollicipes Pollicipes). Ph.D. Thesis, Newcastle University, Newcastle, UK, 2014. [Google Scholar]
- Znidarsic, N.; Mrak, P.; Rajh, E.; Soderznik, K.; Ceh, M.; Strus, J. Cuticle Matrix Imaging by Histochemistry, Fluorescence, and Electron Microscopy. Resolut. Discov. 2018, 3, 5–12. [Google Scholar] [CrossRef]
- Crenshaw, D.G. Hydrostatic Support of the Pedunculate Barnacle Pollicipes Polymerus. Comp. Biochem. Physiol. 1979, 62, 423–425. [Google Scholar] [CrossRef]
- Temereva, E.; Shcherbakova, T.; Tzetlin, A. First Data on the Structure of Tubes Formed by Phoronids. Zoology 2020, 143, 125849. [Google Scholar] [CrossRef]
- Merz, R.A. Textures and Traction: How Tube-Dwelling Polychaetes Get a Leg Up. Invertebr. Biol. 2015, 134, 61–77. [Google Scholar] [CrossRef]
- De Andrade, S.M.B.; Ladchumananandasivam, R.; da Rocha, B.G.; Belarmino, D.D.; Galvão, A.O. The Use of Exoskeletons of Shrimp (Litopenaeus Vanammei) and Crab (Ucides Cordatus) for the Extraction of Chitosan and Production of Nanomembrane. Mater. Sci. Appl. 2012, 3, 21160. [Google Scholar]
- De Queiroz Antonino, R.S.C.M.; Lia Fook, B.R.P.; de Oliveira Lima, V.A.; de Farias Rached, R.Í.; Lima, E.P.N.; da Silva Lima, R.J.; Peniche Covas, C.A.; Lia Fook, M.V. Preparation and Characterization of Chitosan Obtained from Shells of Shrimp (Litopenaeus Vannamei Boone). Mar. Drugs 2017, 15, 141. [Google Scholar] [CrossRef] [Green Version]
- Taylor, J.R.A.; Hebrank, J.; Kier, W.M. Mechanical Properties of the Rigid and Hydrostatic Skeletons of Molting Blue Crabs, Callinectes Sapidus Rathbun. J. Exp. Biol. 2007, 210, 4272–4278. [Google Scholar] [CrossRef] [Green Version]
- Michels, J.; Appel, E.; Gorb, S.N. Functional Diversity of Resilin in Arthropoda. Beilstein J. Nanotechnol. 2016, 7, 1241–1259. [Google Scholar] [CrossRef] [Green Version]
- Torres, F.; Troncoso Heros, O.; Torres, C. Mussel Byssus Fibres: A Tough Biopolymer. RSC Green Chem. 2012, 1, 305–329. [Google Scholar]
- Waite, J.H.; Harrington, M.J. Following the Thread: Mytilus Mussel Byssus as an Inspired Multi-Functional Biomaterial. Can. J. Chem. 2021, 100, 197–211. [Google Scholar] [CrossRef]
- Chaffee, J.; Lewis, C.A. Pedunculate Barnacle Stalk Growth. J. Exp. Mar. Bio. Ecol. 1988, 124, 145–162. [Google Scholar] [CrossRef]
- Lobo-da-Cunha, A.; Alves, Â.; Oliveira, E.; Cunha, I. The Cement Apparatus of the Stalked Barnacle Pollicipes Pollicipes. Mar. Biol. 2017, 164, 11. [Google Scholar] [CrossRef]
- Machałowski, T.; Wysokowski, M.; Żółtowska-Aksamitowska, S.; Bechmann, N.; Binnewerg, B.; Schubert, M.; Guan, K.; Bornstein, S.R.; Czaczyk, K.; Pokrovsky, O.; et al. Spider Chitin. The Biomimetic Potential and Applications of Caribena Versicolor Tubular Chitin. Carbohydr. Polym. 2019, 226, 115301. [Google Scholar] [CrossRef]
- Cruz, T.; Jacinto, D.; Sousa, A.; Penteado, N.; Pereira, D.; Fernandes, J.N.; Silva, T.; Castro, J.J. The State of the Fishery, Conservation and Management of the Stalked Barnacle Pollicipes Pollicipes in Portugal. Mar. Environ. Res. 2015, 112, 73–80. [Google Scholar] [CrossRef]
- Franco, S.C.; Aldred, N.; Cruz, T.; Clare, A.S. Effects of Culture Conditions on Larval Growth and Survival of Stalked Barnacles (Pollicipes Pollicipes). Aquac. Res. 2016, 48, 2920–2933. [Google Scholar] [CrossRef]
- Song, G.; Delroisse, J.; Schoenaers, D.; Kim, H.; Nguyen, T.C.; Horbelt, N.; Leclère, P.; Hwang, D.S.; Harrington, M.J.; Flammang, P. Structure and Composition of the Tunic in the Sea Pineapple Halocynthia Roretzi: A Complex Cellulosic Composite Biomaterial. Acta Biomater. 2020, 111, 290–301. [Google Scholar] [CrossRef]
- Ehrlich, H. Marine Biological Materials of Invertebrate Origin; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis of the AOAC, 15th ed.; Association of Official Analytical Chemists: Washington, DC, USA, 1990. [Google Scholar]
- Clark, A.J.; Triblehorn, J.D. Mechanical Properties of the Cuticles of Three Cockroach Species That Differ in Their Wind-Evoked Escape Behavior. PeerJ 2014, 2, e501. [Google Scholar] [CrossRef]
- Promwikorn, W.; Boonyoung, P.; Pornpimol, K. Histological Characterization of Cuticular Depositions throughout the Molting Cycle of the Black Tiger Shrimp (Penaeus Monodon). Songklanakarin J. Sci. Technol. 2005, 27, 765–772. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| % Dry Weight | |||
|---|---|---|---|
| Ash | Protein | Lipids | Carbohydrates |
| 75.16 ± 0.86 | 19.91 ± 0.13 | 0.33 ± 0.01 | 3.51 ± 0.86 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Almeida, M.; Fernandes, E.M.; Marques, C.F.; Lobo, F.C.M.; Sousa, R.O.; Reis, R.L.; Silva, T.H. Structure and Composition of the Cuticle of the Goose Barnacle Pollicipes pollicipes: A Flexible Composite Biomaterial. Mar. Drugs 2023, 21, 96. https://doi.org/10.3390/md21020096
Almeida M, Fernandes EM, Marques CF, Lobo FCM, Sousa RO, Reis RL, Silva TH. Structure and Composition of the Cuticle of the Goose Barnacle Pollicipes pollicipes: A Flexible Composite Biomaterial. Marine Drugs. 2023; 21(2):96. https://doi.org/10.3390/md21020096
Chicago/Turabian StyleAlmeida, Mariana, Emanuel M. Fernandes, Catarina F. Marques, Flávia C. M. Lobo, Rita O. Sousa, Rui L. Reis, and Tiago H. Silva. 2023. "Structure and Composition of the Cuticle of the Goose Barnacle Pollicipes pollicipes: A Flexible Composite Biomaterial" Marine Drugs 21, no. 2: 96. https://doi.org/10.3390/md21020096
APA StyleAlmeida, M., Fernandes, E. M., Marques, C. F., Lobo, F. C. M., Sousa, R. O., Reis, R. L., & Silva, T. H. (2023). Structure and Composition of the Cuticle of the Goose Barnacle Pollicipes pollicipes: A Flexible Composite Biomaterial. Marine Drugs, 21(2), 96. https://doi.org/10.3390/md21020096