

Anti-Parkinson Effects of Holothuria leucospilota-Derived Palmitic Acid in Caenorhabditis elegans Model of Parkinson’s Disease

 , ,
, ,
Abstract
:1. Introduction
2. Results
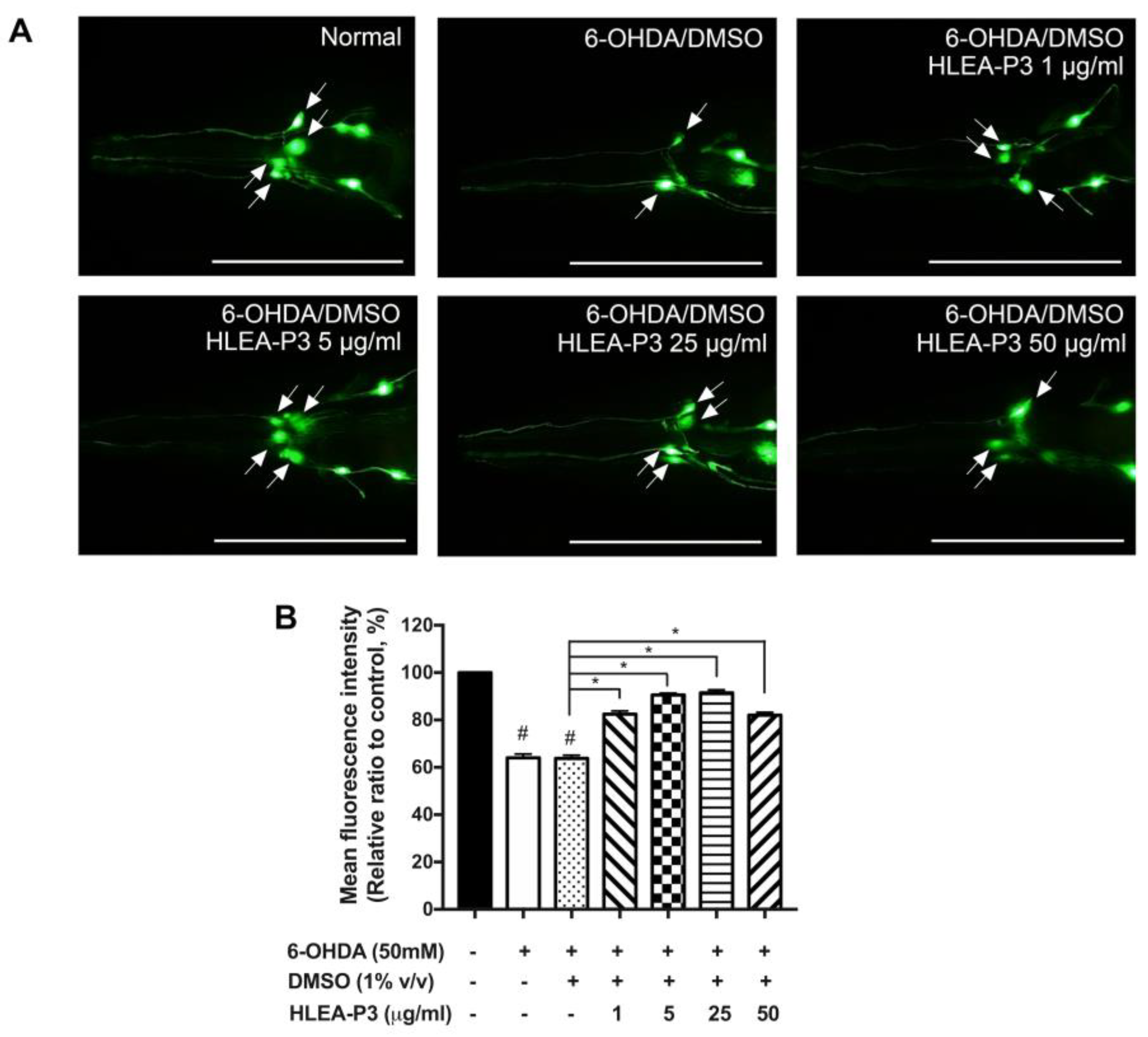
2.1. HLEA-P3 Attenuated DAergic Neurodegeneration Induced by 6-OHDA
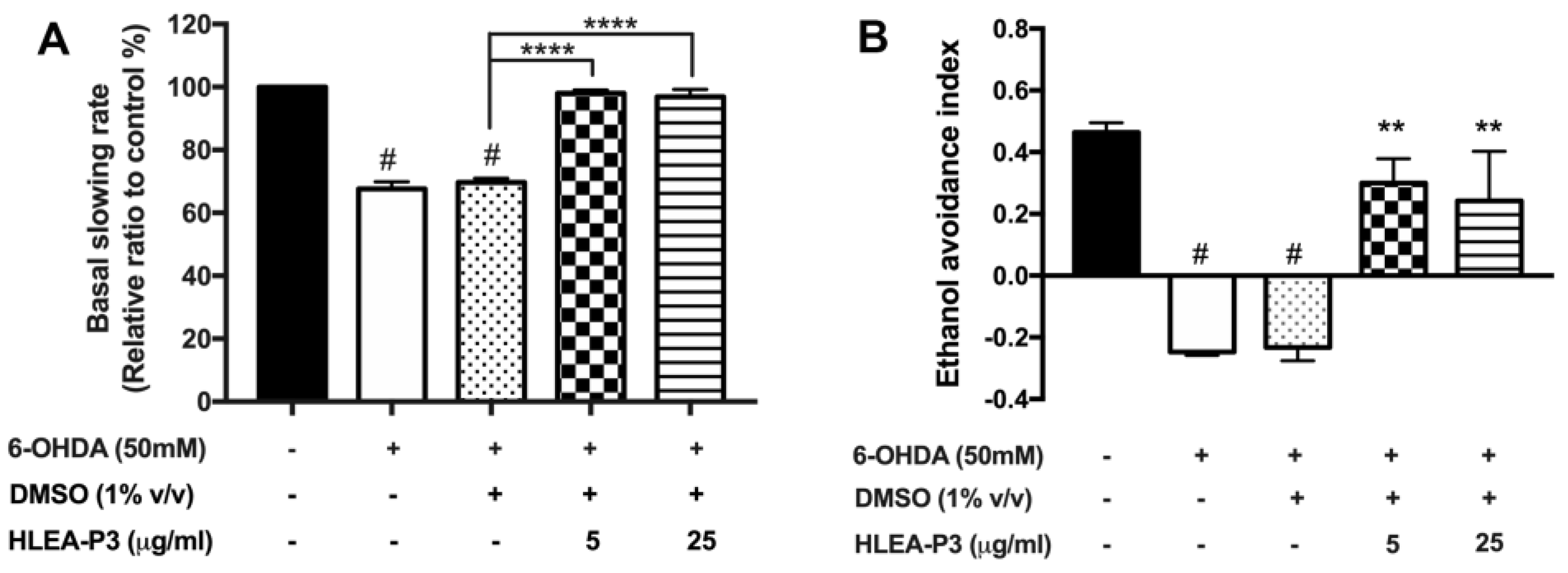
2.2. HLEA-P3 Significantly Improved Dopamine-Dependent Behaviors in 6-OHDA-Treated C. elegans
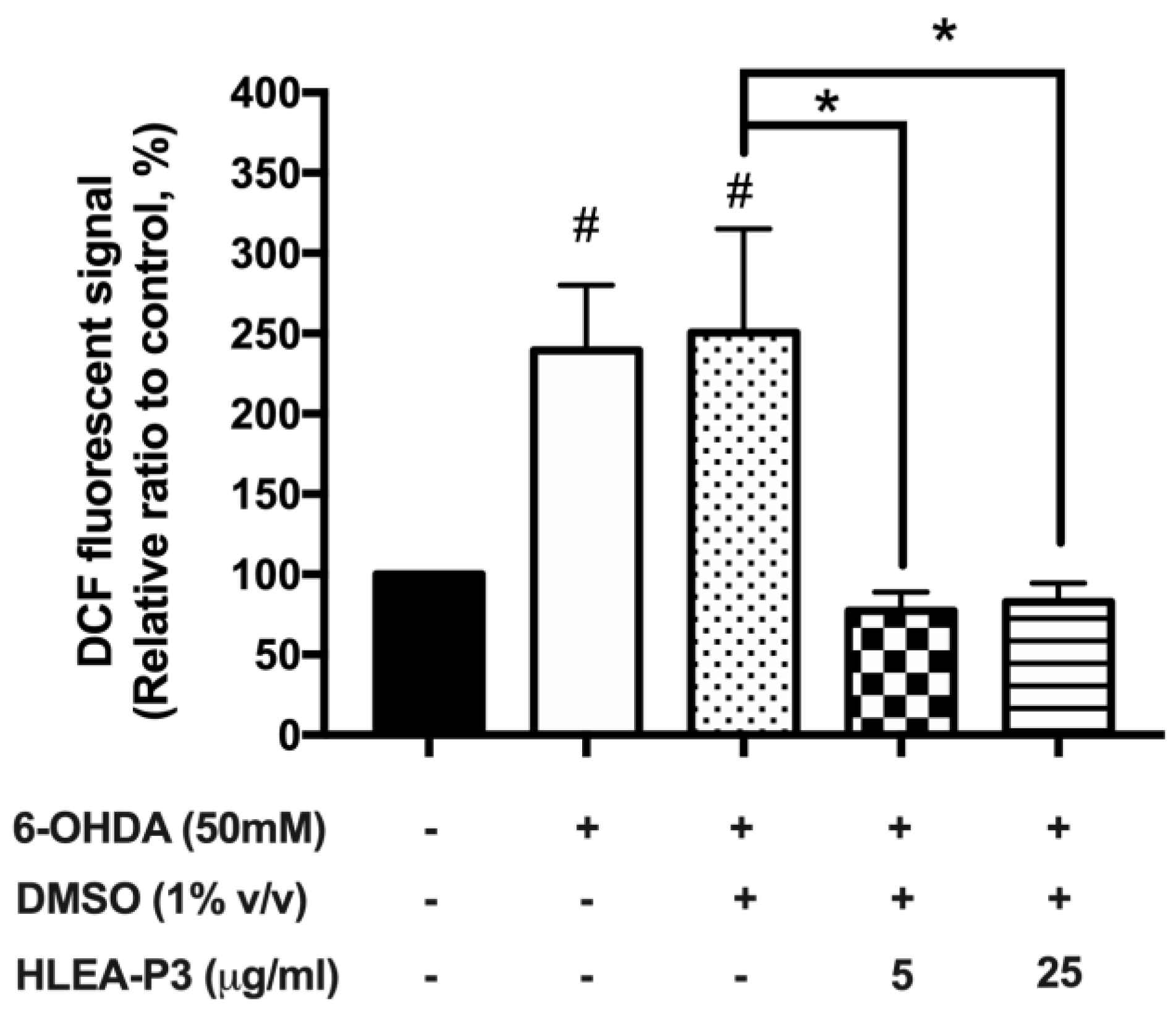
2.3. HLEA-P3 Reduced Oxidative Stress Induced by 6-OHDA in C. elegans
2.4. HLEA-P3 Reduced α-Synuclein Aggregation and Improved Thrashing Behavior in Transgenic C. elegans Expressing α-Synuclein
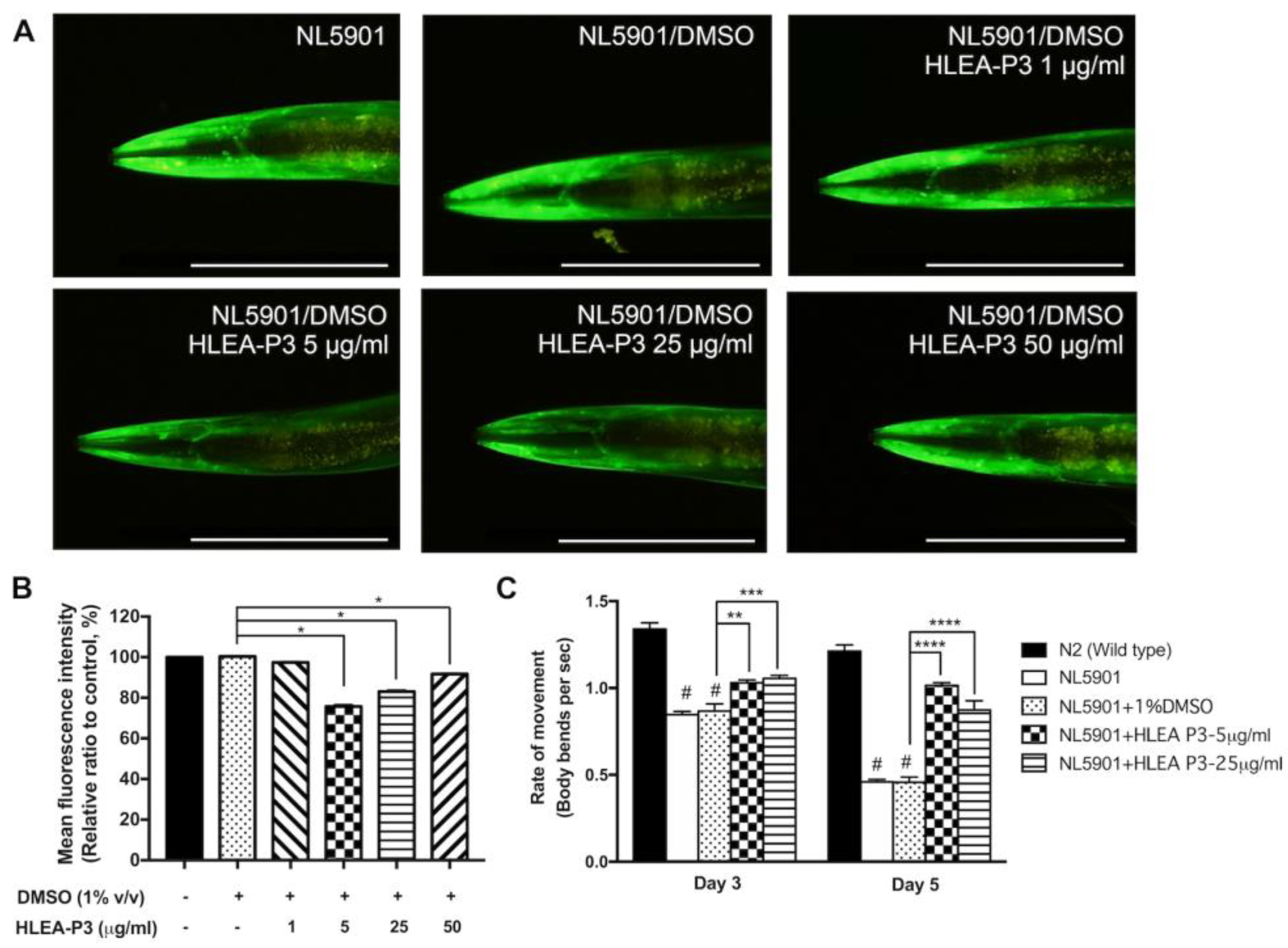
2.5. HLEA-P3 Reduced Lipid Accumulation in Transgenic C. elegans NL5901 Strain
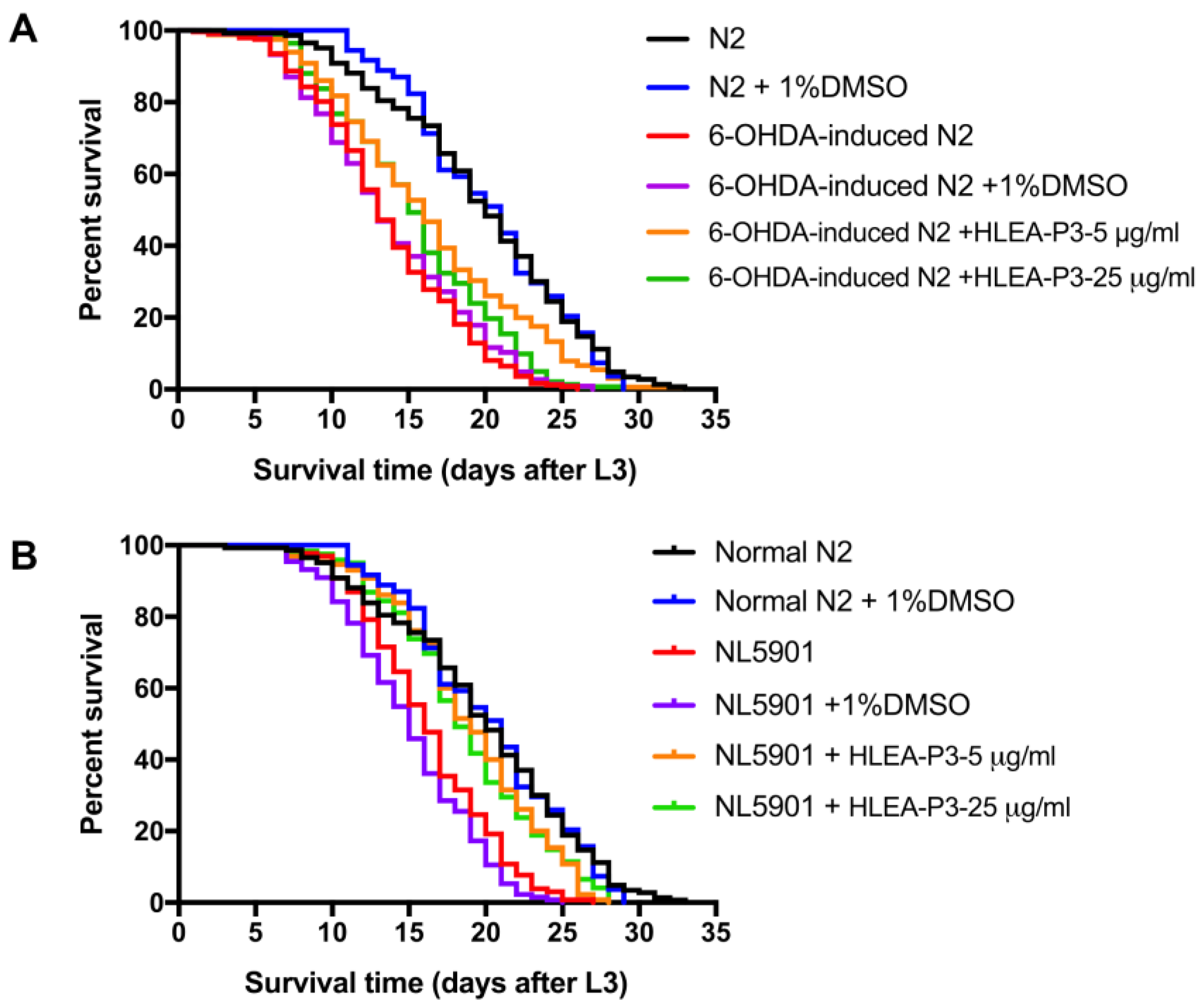
2.6. HLEA-P3 Significantly Prolonged Lifespan of C. elegans PD Models
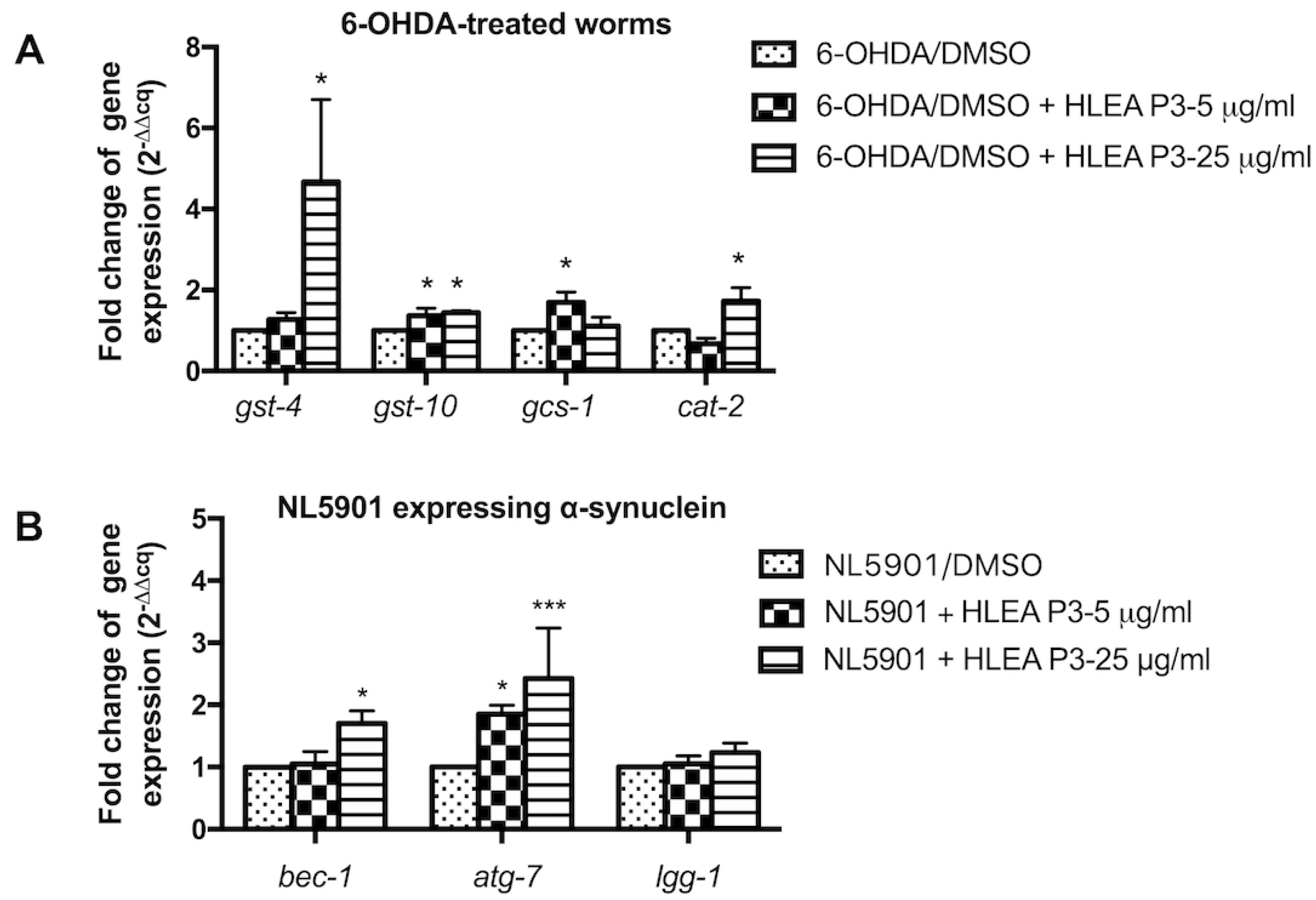
2.7. HLEA-P3 Upregulated Antioxidant Genes and Autophagic Mediators in C. elegans PD Models
2.8. Chemical Structural Analysis Identified HLEA-P3 as Palmitic Acid
3. Discussion
4. Materials and Methods
4.1. Strains, Growth Condition and Synchronization of C. elegans
4.2. Extraction, Isolation and Chemical Characterization of HLEA-P3 from H. leucospilota Ethyl Acetate Fraction
4.3. 6-OHDA-Induced DAergic Neurodegeneration Assay and HLEA-P3 Treatment
4.4. Quantitative Analysis of the Viability of DAergic Neurons
4.5. Assay for Basal Slowing Response Behavior
4.6. Assay for Ethanol Avoidance Behavior
4.7. Quantitative Analysis of α-Synuclein Aggregation
4.8. Assay for Thrashing Behavior
4.9. Quantitative Analysis of Lipid Accumulation
4.10. Quantitative Analysis of Intracellular ROS
4.11. Lifespan Analysis
4.12. Quantitative RT-PCR
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Dorsey, E.; Sherer, T.; Okun, M.S.; Bloem, B.R. The emerging evidence of the Parkinson pandemic. J. Park. Dis. 2018, 8, S3–S8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingelsson, M. Alpha-synuclein oligomers—Neurotoxic molecules in Parkinson’s disease and other Lewy body disorders. Front. Neurosci. 2016, 10, 408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dauer, W.; Przedborski, S. Parkinson’s disease: Mechanisms and models. Neuron 2003, 39, 889–909. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, K.-H.; Chen, C.-M. The role of oxidative stress in Parkinson’s disease. Antioxidants 2020, 9, 597. [Google Scholar] [CrossRef] [PubMed]
- Moore, D.J.; West, A.B.; Dawson, V.L.; Dawson, T.M. Molecular pathophysiology of Parkinson’s disease. Annu. Rev. Neurosci. 2005, 28, 57–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.; Jo, M.G.; Kim, S.Y.; Chung, C.G.; Lee, S.B. Dietary antioxidants and the mitochondrial quality control: Their potential roles in Parkinson’s disease treatment. Antioxidants 2020, 9, 1056. [Google Scholar] [CrossRef]
- Poewe, W.; Seppi, K.; Tanner, C.M.; Halliday, G.M.; Brundin, P.; Volkmann, J.; Schrag, A.-E.; Lang, A.E. Parkinson disease. Nat. Rev. Dis. Prim. 2017, 3, 1–21. [Google Scholar] [CrossRef]
- He, S.; Zhong, S.; Liu, G.; Yang, J. Alpha-synuclein: The interplay of pathology, neuroinflammation, and environmental factors in Parkinson’s disease. Neurodegener. Dis. 2020, 20, 55–64. [Google Scholar] [CrossRef]
- Cai, R.; Zhang, Y.; Simmering, J.E.; Schultz, J.L.; Li, Y.; Fernandez-Carasa, I.; Consiglio, A.; Raya, A.; Polgreen, P.M.; Narayanan, N.S. Enhancing glycolysis attenuates Parkinson’s disease progression in models and clinical databases. J. Clin. Investig. 2019, 129, 4539–4549. [Google Scholar] [CrossRef] [Green Version]
- Mishra, B.B.; Tiwari, V.K. Natural products: An evolving role in future drug discovery. Eur. J. Med. Chem. 2011, 46, 4769–4807. [Google Scholar] [CrossRef]
- Siahaan, E.A.; Pangestuti, R.; Munandar, H.; Kim, S.-K. Cosmeceuticals properties of sea cucumbers: Prospects and trends. Cosmetics 2017, 4, 26. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Zhang, R.; Wen, Z. Bioactive compounds and biological functions of sea cucumbers as potential functional foods. J. Funct. Foods 2018, 49, 73–84. [Google Scholar] [CrossRef]
- Hossain, A.; Dave, D.; Shahidi, F. Antioxidant potential of sea cucumbers and their beneficial effects on human health. Mar. Drugs 2022, 20, 521. [Google Scholar] [CrossRef] [PubMed]
- Chalorak, P.; Jattujan, P.; Nobsathian, S.; Poomtong, T.; Sobhon, P.; Meemon, K. Holothuria scabra extracts exhibit anti-Parkinson potential in C. elegans: A model for anti-Parkinson testing. Nutr. Neurosci. 2018, 21, 427–438. [Google Scholar] [CrossRef]
- Chalorak, P.; Sornkaew, N.; Manohong, P.; Niamnont, N.; Malaiwong, N.; Limboonreung, T.; Sobhon, P.; Aschner, M.; Meemon, K. Diterpene glycosides from Holothuria scabra exert the α-synuclein degradation and neuroprotection against α-synuclein-Mediated neurodegeneration in C. elegans model. J. Ethnopharmacol. 2021, 279, 114347. [Google Scholar] [CrossRef]
- Kitisin, T.; Suphamungmee, W.; Meemon, K. Saponin-rich extracts from Holothuria leucospilota mediate lifespan extension and stress resistance in Caenorhabditis elegans via daf-16. J. Food Biochem. 2019, 43, e13075. [Google Scholar] [CrossRef]
- Malaiwong, N.; Chalorak, P.; Jattujan, P.; Manohong, P.; Niamnont, N.; Suphamungmee, W.; Sobhon, P.; Meemon, K. Anti-Parkinson activity of bioactive substances extracted from Holothuria leucospilota. Biomed. Pharmacother. 2019, 109, 1967–1977. [Google Scholar] [CrossRef]
- Sanguanphun, T.; Sornkaew, N.; Malaiwong, N.; Chalorak, P.; Jattujan, P.; Niamnont, N.; Sobhon, P.; Meemon, K. Neuroprotective effects of a medium chain fatty acid, decanoic acid, isolated from H. leucospilota against Parkinsonism in C. elegans PD model. Front. Pharmacol. 2022, 13, 1004568. [Google Scholar] [CrossRef]
- Caldwell, K.A.; Willicott, C.W.; Caldwell, G.A. Modeling neurodegeneration in Caenorhabditis elegans. Dis. Model. Mech. 2020, 13, dmm046110. [Google Scholar] [CrossRef]
- Brunetti, G.; Di Rosa, G.; Scuto, M.; Leri, M.; Stefani, M.; Schmitz-Linneweber, C.; Calabrese, V.; Saul, N. Healthspan Maintenance and Prevention of Parkinson’s-like Phenotypes with Hydroxytyrosol and Oleuropein Aglycone in C. elegans. Int. J. Mol. Sci. 2020, 21, 2588. [Google Scholar] [CrossRef] [Green Version]
- Hsu, Y.-L.; Hung, H.-S.; Tsai, C.-W.; Liu, S.-P.; Chiang, Y.-T.; Kuo, Y.-H.; Shyu, W.-C.; Lin, S.-Z.; Fu, R.-H. Peiminine reduces ARTS-mediated degradation of XIAP by modulating the PINK1/parkin pathway to ameliorate 6-hydroxydopamine toxicity and α-synuclein accumulation in Parkinson’s disease models in vivo and in vitro. Int. J. Mol. Sci. 2021, 22, 10240. [Google Scholar] [CrossRef]
- Nass, R.; Hall, D.H.; Miller, D.M.; Blakely, R.D. Neurotoxin-induced degeneration of dopamine neurons in Caenorhabditis elegans. Proc. Natl. Acad. Sci. USA 2002, 99, 3264–3269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maulik, M.; Mitra, S.; Bult-Ito, A.; Taylor, B.E.; Vayndorf, E.M. Behavioral phenotyping and pathological indicators of Parkinson’s disease in C. elegans models. Front. Genet. 2017, 8, 77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blesa, J.; Trigo-Damas, I.; Quiroga-Varela, A.; Lopez-Gonzalez del Rey, N. Animal models of Parkinson’s disease. Challenges in Parkinson’s Disease; InTechOpen: London, UK, 2016; pp. 195–216. [Google Scholar]
- Van Ham, T.J.; Thijssen, K.L.; Breitling, R.; Hofstra, R.M.; Plasterk, R.H.; Nollen, E.A.C. elegans model identifies genetic modifiers of α-synuclein inclusion formation during aging. PLoS Genet. 2008, 4, e1000027. [Google Scholar] [CrossRef] [PubMed]
- Cooper, J.F.; Dues, D.J.; Spielbauer, K.K.; Machiela, E.; Senchuk, M.M.; Van Raamsdonk, J.M. Delaying aging is neuroprotective in Parkinson’s disease: A genetic analysis in C. elegans models. NPJ Park. Dis. 2015, 1, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Maulik, M.; Mitra, S.; Basmayor, A.M.; Lu, B.; Taylor, B.E.; Bult-Ito, A. Genetic silencing of fatty acid desaturases modulates α-synuclein toxicity and neuronal loss in Parkinson-like models of C. elegans. Front. Aging Neurosci. 2019, 11, 207. [Google Scholar] [CrossRef] [Green Version]
- Maiti, P.; Manna, J.; Dunbar, G.L. Current understanding of the molecular mechanisms in Parkinson’s disease: Targets for potential treatments. Transl. Neurodegener. 2017, 6, 28. [Google Scholar] [CrossRef] [Green Version]
- Blum, D.; Torch, S.; Lambeng, N.; Nissou, M.-F.; Benabid, A.-L.; Sadoul, R.; Verna, J.-M. Molecular pathways involved in the neurotoxicity of 6-OHDA, dopamine and MPTP: Contribution to the apoptotic theory in Parkinson’s disease. Prog. Neurobiol. 2001, 65, 135–172. [Google Scholar] [CrossRef]
- Simonian, N.; Coyle, J. Oxidative stress in neurodegenerative diseases. Annu. Rev. Pharmacol. Toxicol. 1996, 36, 83–106. [Google Scholar] [CrossRef]
- Ferguson, G.D.; Bridge, W.J. The glutathione system and the related thiol network in Caenorhabditis elegans. Redox Biol. 2019, 24, 101171. [Google Scholar] [CrossRef]
- Schommer, J.; Marwarha, G.; Nagamoto-Combs, K.; Ghribi, O. Palmitic acid-enriched diet increases α-Synuclein and tyrosine hydroxylase expression levels in the mouse brain. Front. Neurosci. 2018, 12, 552. [Google Scholar] [CrossRef] [PubMed]
- Van Raamsdonk, J.M.; Hekimi, S. Superoxide dismutase is dispensable for normal animal lifespan. Proc. Natl. Acad. Sci. USA 2012, 109, 5785–5790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lapointe, J.; Hekimi, S. When a theory of aging ages badly. Cell. Mol. Life Sci. 2010, 67, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Mutlu, A.S.; Duffy, J.; Wang, M.C. Lipid metabolism and lipid signals in aging and longevity. Dev. Cell 2021, 56, 1394–1407. [Google Scholar] [CrossRef]
- Jattujan, P.; Srisirirung, S.; Watcharaporn, W.; Chumphoochai, K.; Kraokaew, P.; Sanguanphun, T.; Prasertsuksri, P.; Thongdechsri, S.; Sobhon, P.; Meemon, K. 2-Butoxytetrahydrofuran and Palmitic Acid from Holothuria scabra Enhance C. elegans Lifespan and Healthspan via DAF-16/FOXO and SKN-1/NRF2 Signaling Pathways. Pharmaceuticals 2022, 15, 1374. [Google Scholar] [CrossRef] [PubMed]
- López-Domínguez, J.A.; Ramsey, J.J.; Tran, D.; Imai, D.M.; Koehne, A.; Laing, S.T.; Griffey, S.M.; Kim, K.; Taylor, S.L.; Hagopian, K. The influence of dietary fat source on life span in calorie restricted mice. J. Gerontol. Ser. A Biomed. Sci. Med. Sci. 2015, 70, 1181–1188. [Google Scholar] [CrossRef] [Green Version]
- Kumar, P.P.; Kumaravel, S.; Lalitha, C. Screening of antioxidant activity, total phenolics and GC-MS study of Vitex negundo. Afr. J. Biochem. Res. 2010, 4, 191–195. [Google Scholar]
- Hidajati, N.; Tukiran, T.; Setiabudi, D.A.; Wardana, A.P. Antioxidant activity of palmitic acid and pinostrobin from methanol extract of Syzygium litoralle (Myrtaceae). In Proceedings of the International Conference on Science and Technology (ICST 2018), Limerick, Ireland, 4–6 December 2018; pp. 183–187. [Google Scholar]
- Lambertucci, R.H.; Hirabara, S.M.; Silveira, L.D.R.; Levada-Pires, A.C.; Curi, R.; Pithon-Curi, T.C. Palmitate increases superoxide production through mitochondrial electron transport chain and NADPH oxidase activity in skeletal muscle cells. J. Cell. Physiol. 2008, 216, 796–804. [Google Scholar] [CrossRef]
- Xu, S.; Nam, S.; Kim, J.; Das, R.; Choi, S.; Nguyen, T.; Quan, X.; Choi, S.; Chung, C.; Lee, E. Palmitate induces ER calcium depletion and apoptosis in mouse podocytes subsequent to mitochondrial oxidative stress. Cell Death Dis. 2015, 6, e1976. [Google Scholar] [CrossRef] [Green Version]
- Yuan, H.; Zhang, X.; Huang, X.; Lu, Y.; Tang, W.; Man, Y.; Wang, S.; Xi, J.; Li, J. NADPH oxidase 2-derived reactive oxygen species mediate FFAs-induced dysfunction and apoptosis of β-cells via JNK, p38 MAPK and p53 pathways. PloS ONE 2010, 5, e15726. [Google Scholar] [CrossRef] [Green Version]
- Ng, Y.-W.; Say, Y.-H. Palmitic acid induces neurotoxicity and gliatoxicity in SH-SY5Y human neuroblastoma and T98G human glioblastoma cells. PeerJ 2018, 6, e4696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidović, M.; Rikalovic, M.G. Alpha-Synuclein Aggregation Pathway in Parkinson’s Disease: Current Status and Novel Therapeutic Approaches. Cells 2022, 11, 1732. [Google Scholar] [CrossRef]
- Friedman, L.G.; Lachenmayer, M.L.; Wang, J.; He, L.; Poulose, S.M.; Komatsu, M.; Holstein, G.R.; Yue, Z. Disrupted autophagy leads to dopaminergic axon and dendrite degeneration and promotes presynaptic accumulation of α-synuclein and LRRK2 in the brain. J. Neurosci. 2012, 32, 7585–7593. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.-S.; Chen, X.-M.; Wan, J.-M.; Gui, H.-B.; Ruan, X.-Z.; Du, X.-G. Autophagy protects against palmitic acid-induced apoptosis in podocytes in vitro. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, S.H.; Shui, G.; Zhou, J.; Li, J.J.E.; Bay, B.-H.; Wenk, M.R.; Shen, H.-M. Induction of autophagy by palmitic acid via protein kinase C-mediated signaling pathway independent of mTOR (mammalian target of rapamycin). J. Biol. Chem. 2012, 287, 14364–14376. [Google Scholar] [CrossRef] [Green Version]
- Galvagnion, C. The role of lipids interacting with α-synuclein in the pathogenesis of Parkinson’s disease. J. Park. Dis. 2017, 7, 433–450. [Google Scholar] [CrossRef] [Green Version]
- Kiechle, M.; Grozdanov, V.; Danzer, K.M. The role of lipids in the initiation of α-synuclein misfolding. Front. Cell Dev. Biol. 2020, 8, 562241. [Google Scholar] [CrossRef]
- Sharon, R.; Bar-Joseph, I.; Frosch, M.P.; Walsh, D.M.; Hamilton, J.A.; Selkoe, D.J. The formation of highly soluble oligomers of α-synuclein is regulated by fatty acids and enhanced in Parkinson’s disease. Neuron 2003, 37, 583–595. [Google Scholar] [CrossRef] [Green Version]
- Sachdeva, M.; Karan, M.; Singh, T.; Dhingra, S. Oxidants and antioxidants in complementary and alternative medicine: A review. Spatula DD 2014, 4, 1–16. [Google Scholar] [CrossRef]
- Näsström, T.; Fagerqvist, T.; Barbu, M.; Karlsson, M.; Nikolajeff, F.; Kasrayan, A.; Ekberg, M.; Lannfelt, L.; Ingelsson, M.; Bergström, J. The lipid peroxidation products 4-oxo-2-nonenal and 4-hydroxy-2-nonenal promote the formation of α-synuclein oligomers with distinct biochemical, morphological, and functional properties. Free. Radic. Biol. Med. 2011, 50, 428–437. [Google Scholar] [CrossRef]
- Almandoz-Gil, L.; Welander, H.; Ihse, E.; Khoonsari, P.E.; Musunuri, S.; Lendel, C.; Sigvardson, J.; Karlsson, M.; Ingelsson, M.; Kultima, K. Low molar excess of 4-oxo-2-nonenal and 4-hydroxy-2-nonenal promote oligomerization of alpha-synuclein through different pathways. Free. Radic. Biol. Med. 2017, 110, 421–431. [Google Scholar] [CrossRef]
- Shanklin, J.; Cahoon, E.B. Desaturation and related modifications of fatty acids. Annu. Rev. Plant Biol. 1998, 49, 611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paton, C.M.; Ntambi, J.M. Biochemical and physiological function of stearoyl-CoA desaturase. Am. J. Physiol. -Endocrinol. Metab. 2009, 297, E28–E37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores-León, M.; Alcaraz, N.; Pérez-Domínguez, M.; Torres-Arciga, K.; Rebollar-Vega, R.; la Rosa-Velázquez, D.; Arriaga-Canon, C.; Herrera, L.; Arias, C.; González-Barrios, R. Transcriptional Profiles Reveal Deregulation of Lipid Metabolism and Inflammatory Pathways in Neurons Exposed to Palmitic Acid. Mol. Neurobiol. 2021, 58, 4639–4651. [Google Scholar] [CrossRef] [PubMed]
- Vincent, B.M.; Tardiff, D.F.; Piotrowski, J.S.; Aron, R.; Lucas, M.C.; Chung, C.Y.; Bacherman, H.; Chen, Y.; Pires, M.; Subramaniam, R. Inhibiting stearoyl-CoA desaturase ameliorates α-synuclein cytotoxicity. Cell Rep. 2018, 25, 2742–2754.e2731. [Google Scholar] [CrossRef] [Green Version]
- Fanning, S.; Haque, A.; Imberdis, T.; Baru, V.; Barrasa, M.I.; Nuber, S.; Termine, D.; Ramalingam, N.; Ho, G.P.; Noble, T. Lipidomic analysis of α-synuclein neurotoxicity identifies stearoyl CoA desaturase as a target for Parkinson treatment. Mol. Cell 2019, 73, 1001–1014.e1008. [Google Scholar] [CrossRef] [Green Version]
- Chumphoochai, K.; Chalorak, P.; Suphamungmee, W.; Sobhon, P.; Meemon, K. Saponin-enriched extracts from body wall and Cuvierian tubule of Holothuria leucospilota reduce fat accumulation and suppress lipogenesis in Caenorhabditis elegans. J. Sci. Food Agric. 2019, 99, 4158–4166. [Google Scholar] [CrossRef]
- Vesga-Jiménez, D.J.; Martin, C.; Barreto, G.E.; Aristizábal-Pachón, A.F.; Pinzón, A.; González, J. Fatty Acids: An Insight into the Pathogenesis of Neurodegenerative Diseases and Therapeutic Potential. Int. J. Mol. Sci. 2022, 23, 2577. [Google Scholar] [CrossRef]
- Pujols, J.; Peña-Díaz, S.; Lázaro, D.F.; Peccati, F.; Pinheiro, F.; González, D.; Carija, A.; Navarro, S.; Conde-Giménez, M.; García, J. Small molecule inhibits α-synuclein aggregation, disrupts amyloid fibrils, and prevents degeneration of dopaminergic neurons. Proc. Natl. Acad. Sci. USA 2018, 115, 10481–10486. [Google Scholar] [CrossRef] [Green Version]
- Yoon, D.S.; Lee, M.-H.; Cha, D.S. Measurement of intracellular ROS in Caenorhabditis elegans using 2’, 7’-dichlorodihydrofluorescein diacetate. Bio-Protocol 2018, 8, e2774. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Mean Lifespan ± SD (Days) | Maximum Lifespan (Days) | % Increase Lifespan | Number of Worms | p Value (Log-Rank Test) |
|---|---|---|---|---|---|
| N2 | 19.76 ± 0.51 | 33 | - | 143 | - |
| N2 + 1% DMSO | 20.20 ± 0.49 | 29 | - | 108 | - |
| N2 + 50 mM 6-OHDA | 13.61± 0.31 | 26 | −31.12 | 248 | ####, p < 0.0001 |
| N2 + 50 mM 6-OHDA + 1%DMSO | 13.75 ± 0.35 | 27 | −30.41 | 224 | ####, p < 0.0001 |
| N2 + 50 mM 6-OHDA + HLEA-P3 5 μg/mL | 16.36 ± 0.49 | 33 | 18.98 | 165 | ****, p < 0.0001 |
| N2 + 50 mM 6-OHDA + HLEA-P3 25 μg/mL | 15.36 ± 0.43 | 29 | 11.71 | 142 | *, p = 0.0105 |
| NL5901 | 16.28 ± 0.37 | 27 | −17.61 | 130 | ####, p < 0.0001 |
| NL5901 + 1% DMSO | 15.02 ± 0.36 | 25 | −23.99 | 133 | ####, p < 0.0001 |
| NL5901 + HLEA-P3 5 μg/mL | 18.92 ± 0.43 | 28 | 25.97 | 130 | ****, p < 0.0001 |
| NL5901 + HLEA-P3 25 μg/mL | 18.71 ± 0.44 | 28 | 24.57 | 122 | ****, p < 0.0001 |
| Gene Name | Forward Primer (5′ to 3′) | Reverse Primer (5′ to 3′) |
|---|---|---|
| Antioxidative system | ||
| gst-4 | CCCATTTTACAAGTCGATGG | CTTCCTCTGCAGTTTTTCCA |
| gst-10 | GTCTACCACGTTTTGGATGC | ACTTTGTCGGCCTTTCTCTT |
| gcs-1 | AATCGATTCCTTTGGAGACC | TGTTTGCCTCGACAATGTT |
| Dopamine synthesis | ||
| cat-2 | AAAGCGTGTGAAACGTCAGT | TCTGTCCGACTCCTTTCTCCT |
| Protein degradation pathway | ||
| bec-1 | AGATCTCAAAGCTGCGTGTG | AAAAGGCAGAATTCCAGCAGA |
| atg-7 | TCTGCAGGATGGATGGTTCG | CTCGGCAAGGTCCATGTGTA |
| lgg-1 | AATGGAAACCCAAAGCCCCT | AGGGGAGAAGAGCAACTTCG |
| Fatty acid desaturation | ||
| fat-5 | GCCCTCTTCCGTTACTGCTT | CTCCGACTGCCGCAATAGAT |
| fat-6 | GCGCTGCTCACTATTTCGGATGG | GTGGGAATGTGTGATGGAAGTTGTG |
| fat-7 | CATGGAGGCAAACTCGACCT | GTGGCGTGAAGTGTGAAACA |
| Housekeeping gene | ||
| act-1 | ATCGTCACCACCAGCTTTCT | CACACCCGCAAATGAGTGAA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanguanphun, T.; Promtang, S.; Sornkaew, N.; Niamnont, N.; Sobhon, P.; Meemon, K. Anti-Parkinson Effects of Holothuria leucospilota-Derived Palmitic Acid in Caenorhabditis elegans Model of Parkinson’s Disease. Mar. Drugs 2023, 21, 141. https://doi.org/10.3390/md21030141
Sanguanphun T, Promtang S, Sornkaew N, Niamnont N, Sobhon P, Meemon K. Anti-Parkinson Effects of Holothuria leucospilota-Derived Palmitic Acid in Caenorhabditis elegans Model of Parkinson’s Disease. Marine Drugs. 2023; 21(3):141. https://doi.org/10.3390/md21030141
Chicago/Turabian StyleSanguanphun, Tanatcha, Sukrit Promtang, Nilubon Sornkaew, Nakorn Niamnont, Prasert Sobhon, and Krai Meemon. 2023. "Anti-Parkinson Effects of Holothuria leucospilota-Derived Palmitic Acid in Caenorhabditis elegans Model of Parkinson’s Disease" Marine Drugs 21, no. 3: 141. https://doi.org/10.3390/md21030141
APA StyleSanguanphun, T., Promtang, S., Sornkaew, N., Niamnont, N., Sobhon, P., & Meemon, K. (2023). Anti-Parkinson Effects of Holothuria leucospilota-Derived Palmitic Acid in Caenorhabditis elegans Model of Parkinson’s Disease. Marine Drugs, 21(3), 141. https://doi.org/10.3390/md21030141