
Pro-Apoptotic Activity of Bioactive Compounds from Seaweeds: Promising Sources for Developing Novel Anticancer Drugs

 , ,
, ,
Abstract
:1. Introduction
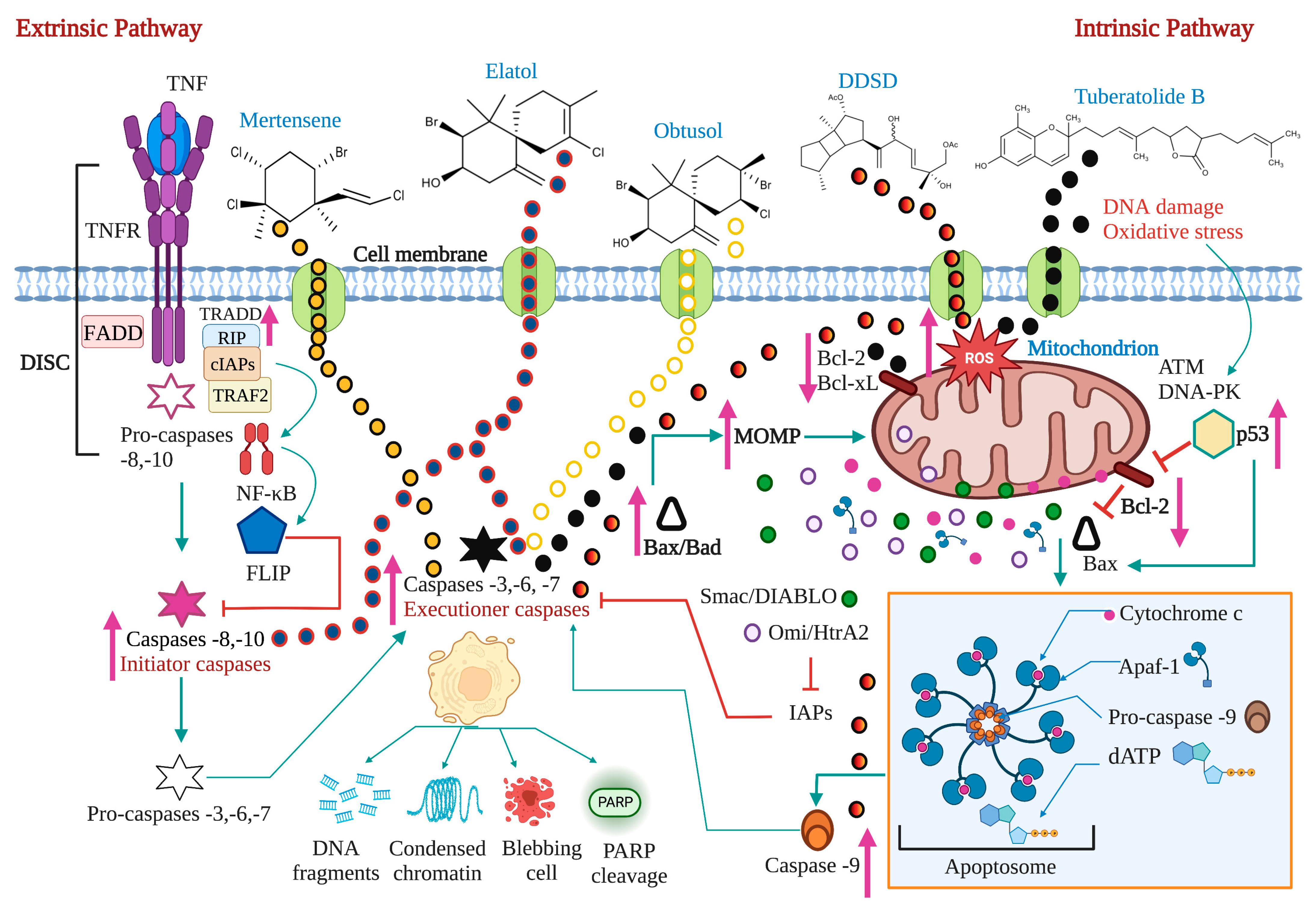
2. Apoptosis: A Target for Anticancer Therapy
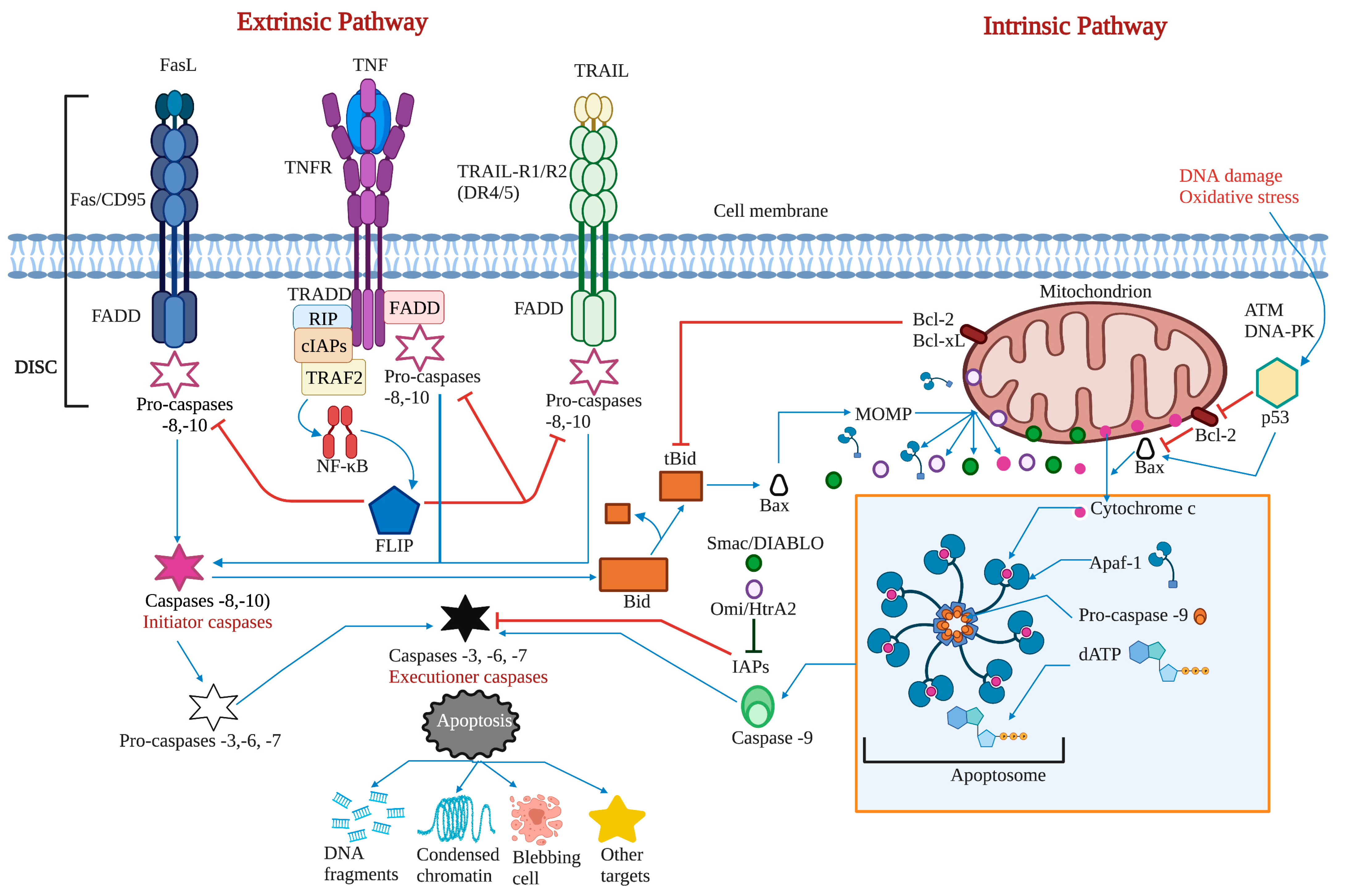
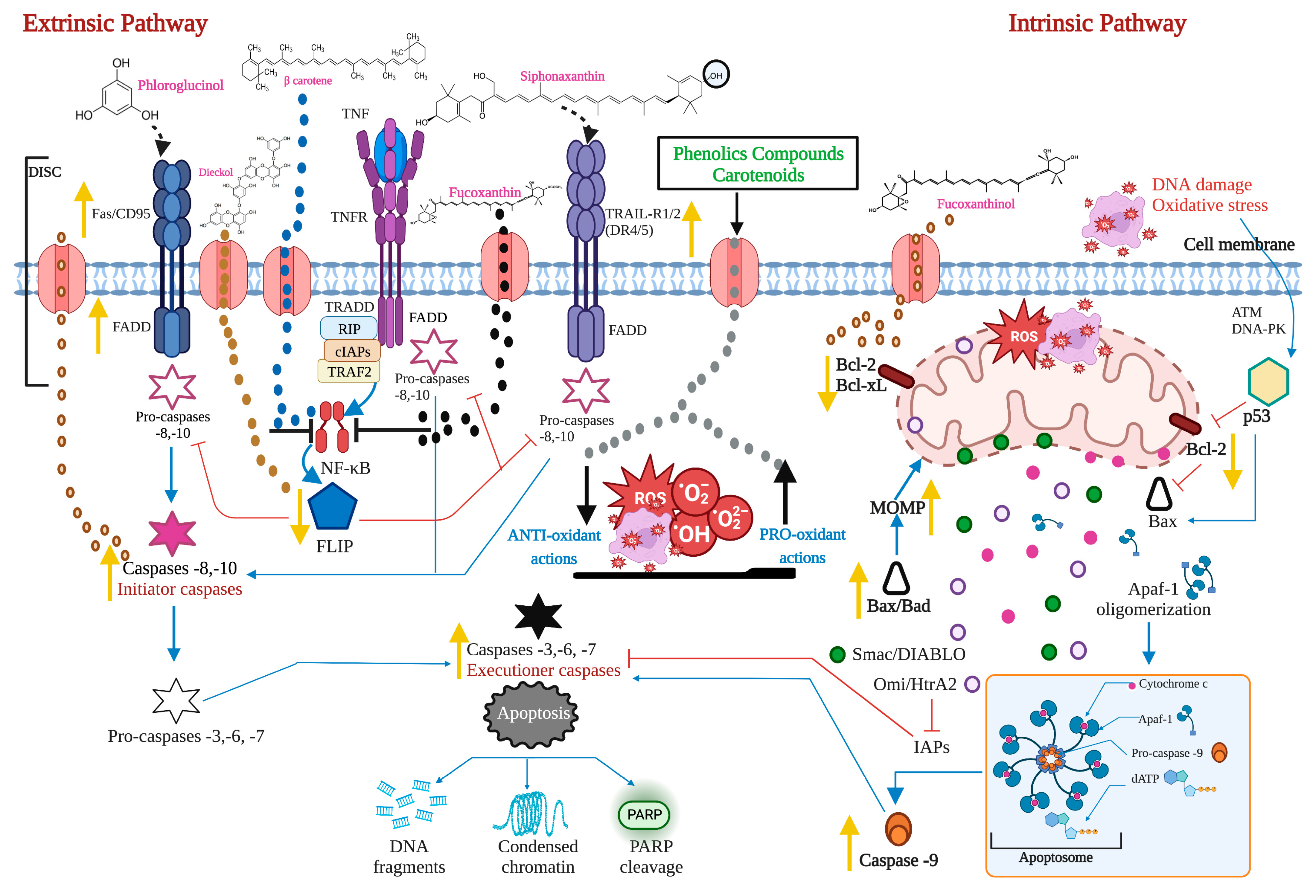
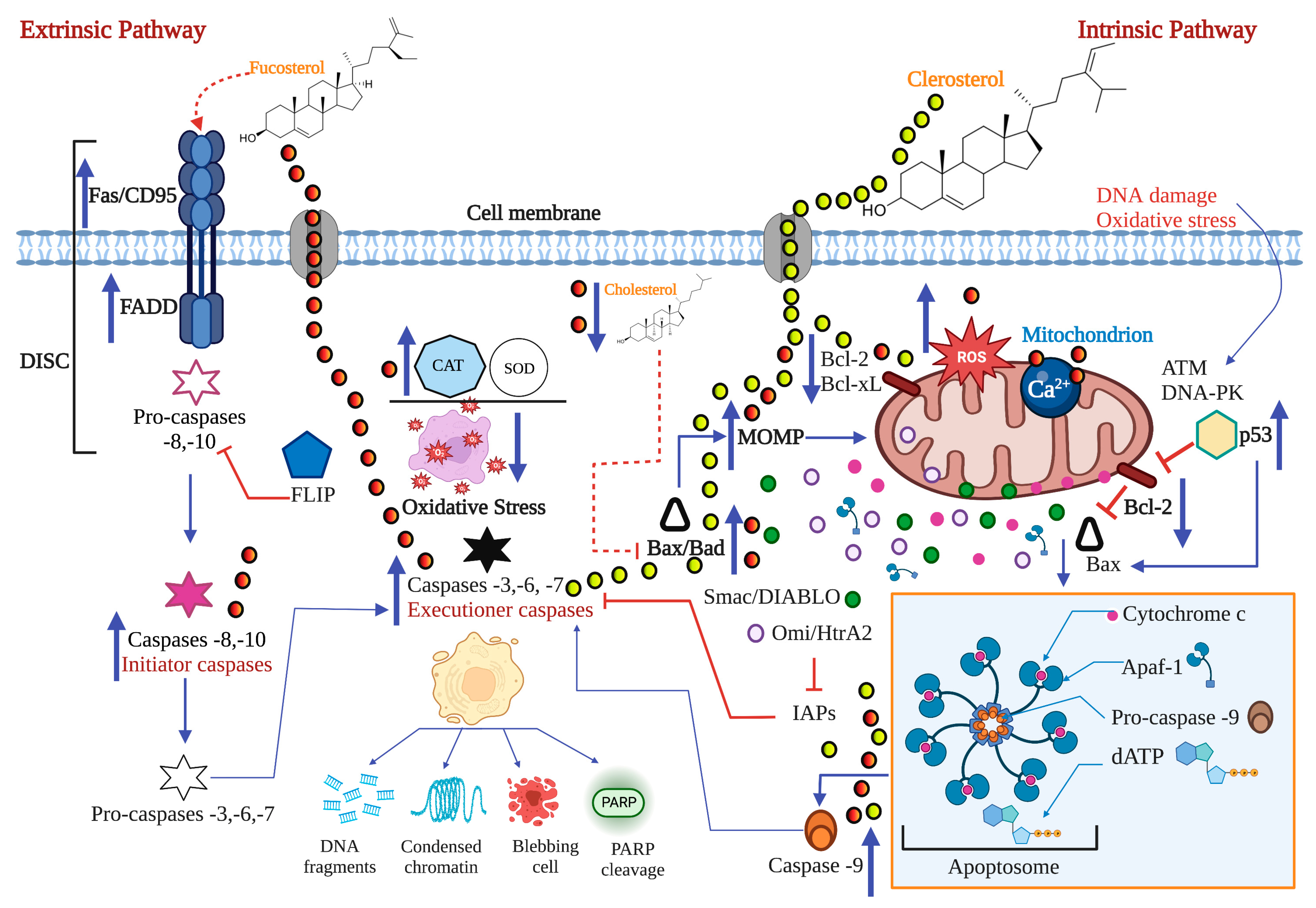
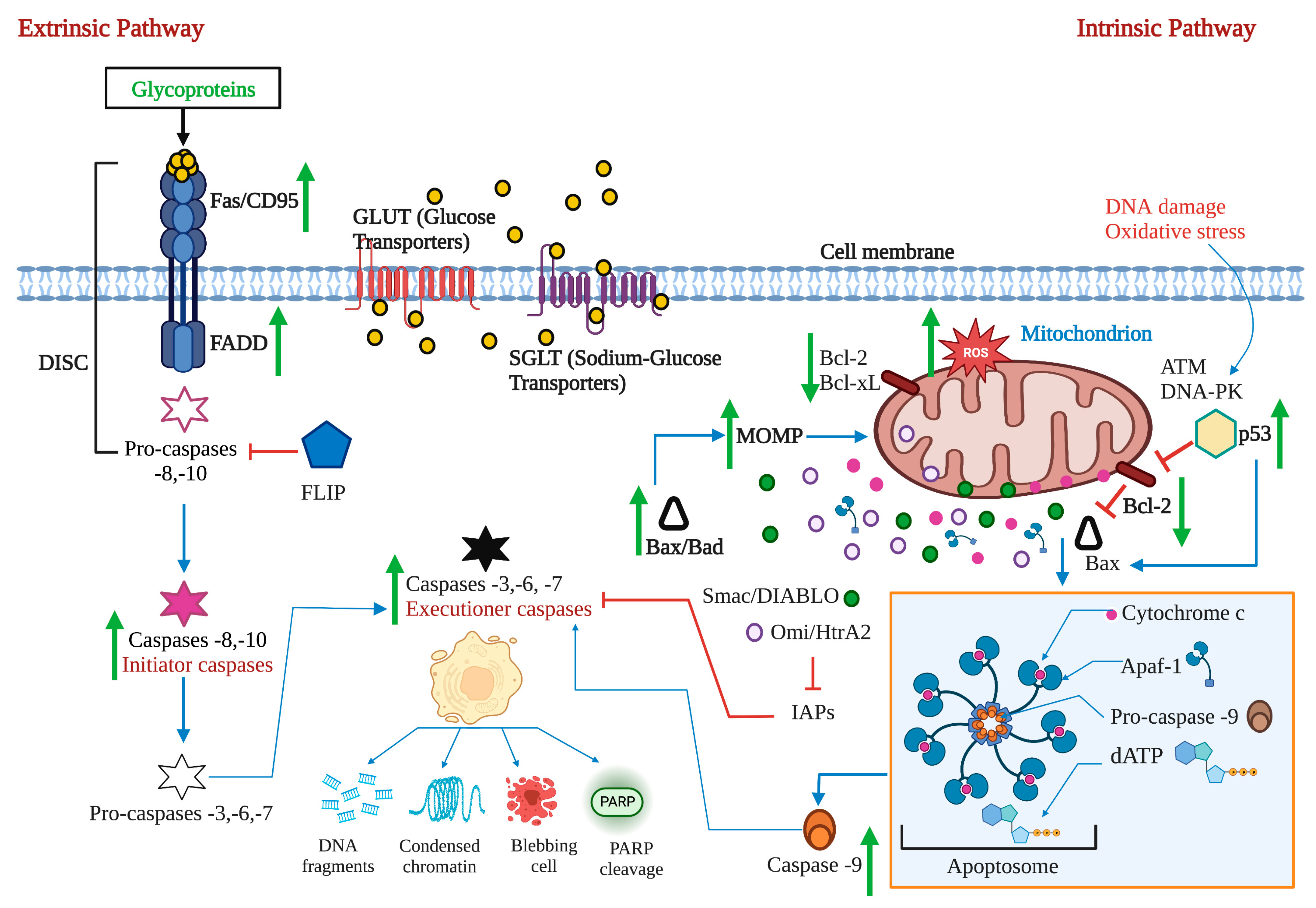
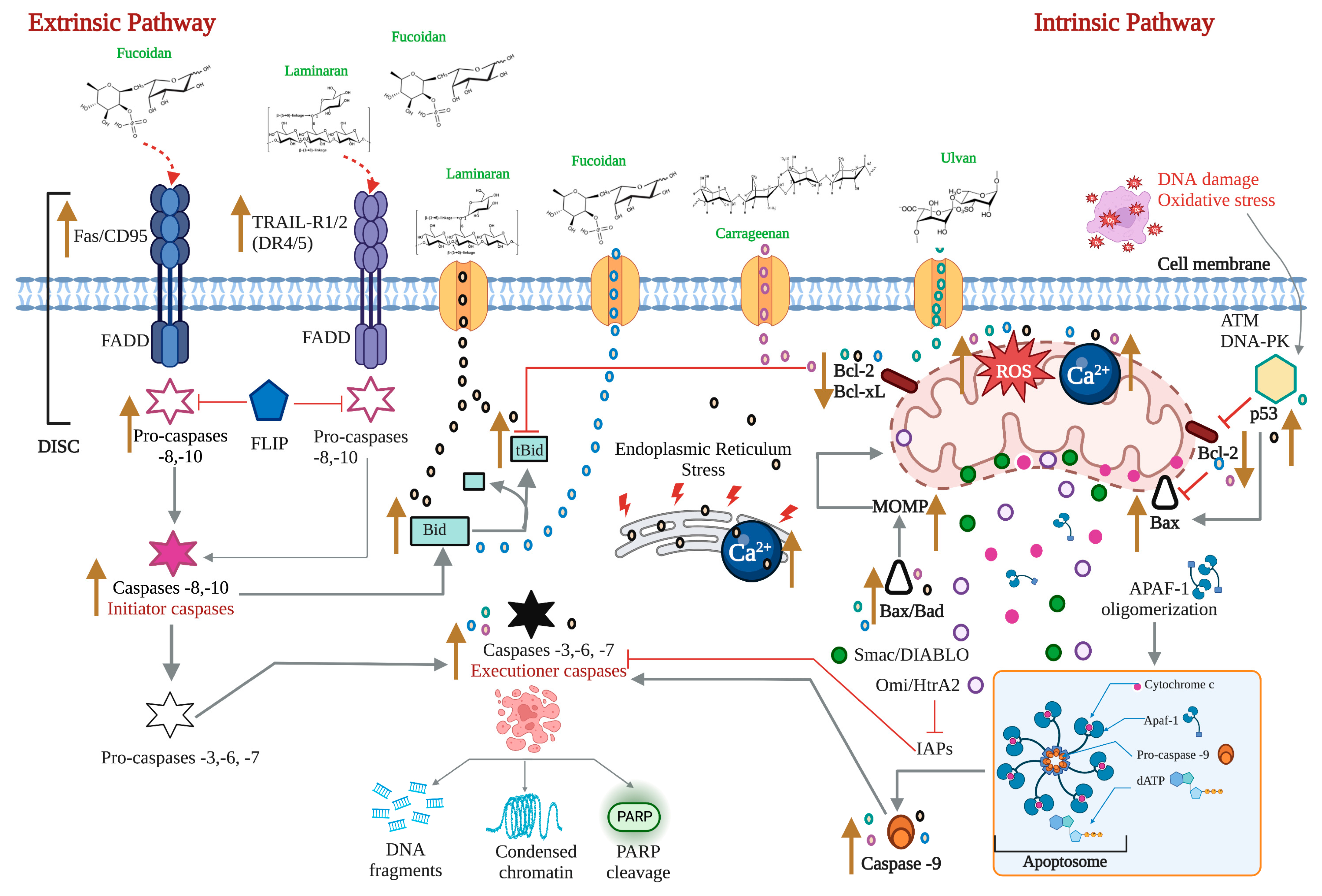
2.1. The Extrinsic or Death Receptor Pathway
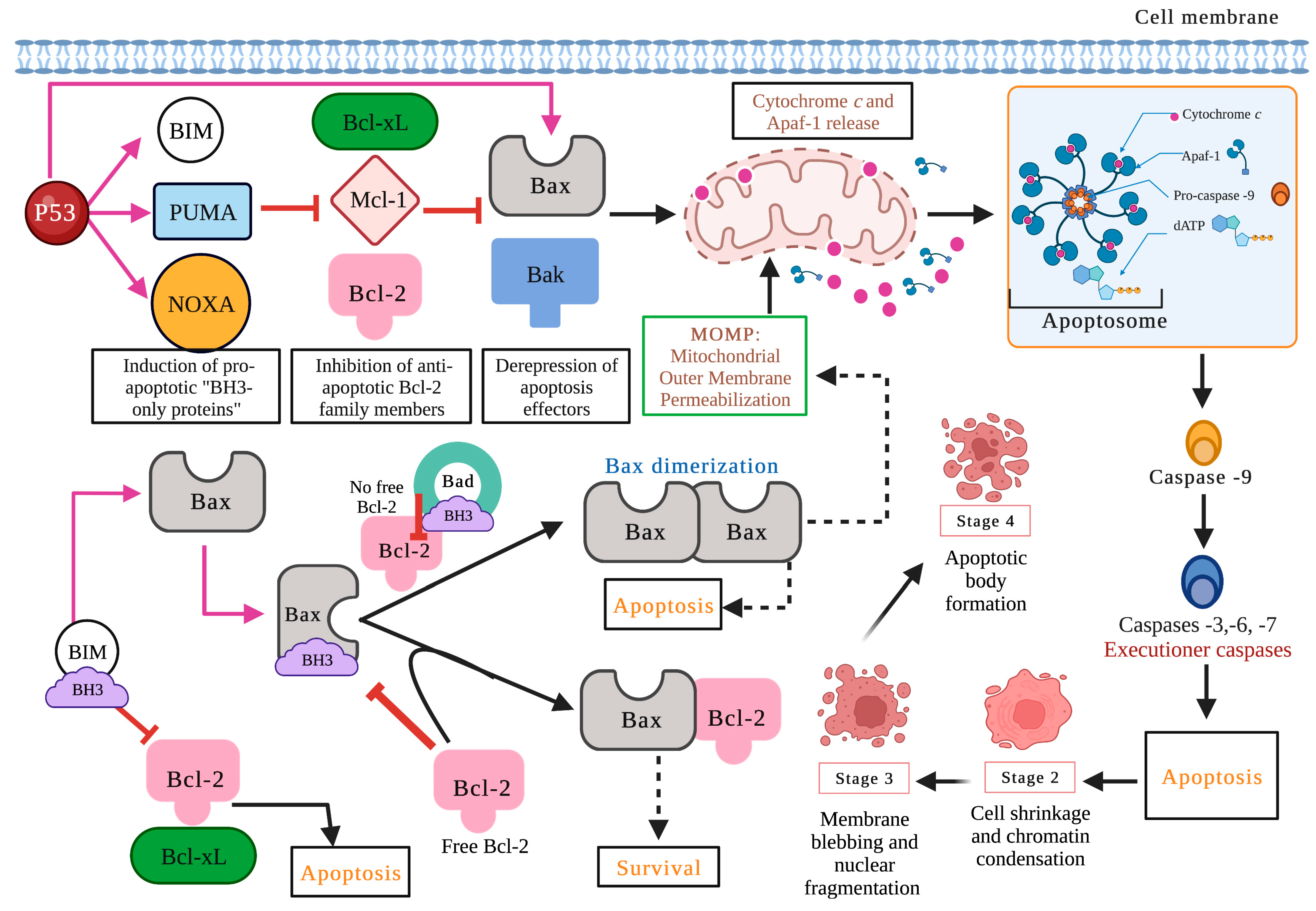
2.2. The Intrinsic or Mitochondrial Pathway
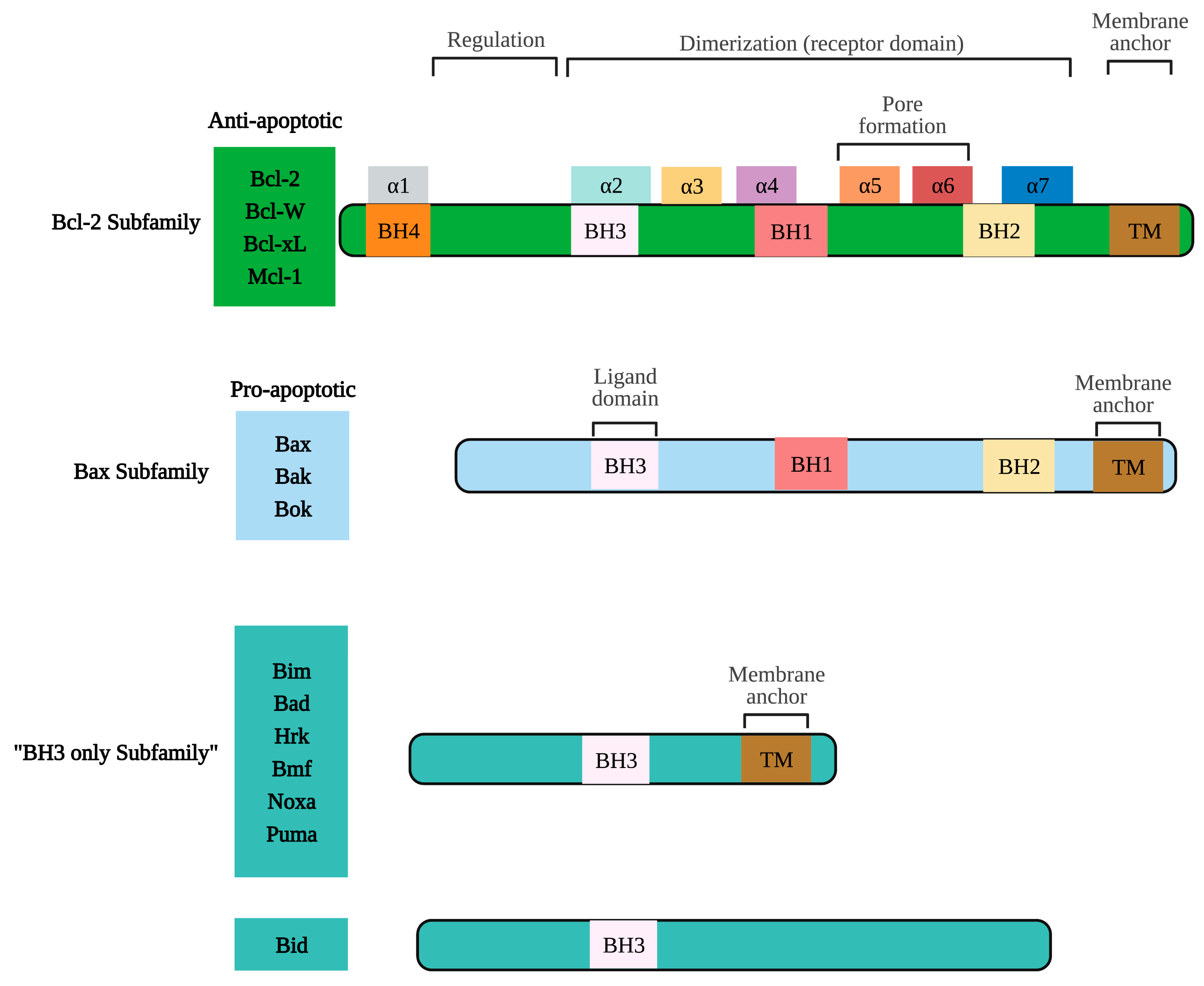
2.3. Proteins of the Bcl-2 Family, P53, and Caspases in Carcinogenesis
2.4. ROS, Ca2+, and Mitochondria in the Apoptotic Process
3. Pro-Apoptotic Activity of Bioactive Compounds from Seaweeds

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Compound | Cell Line | IC50 | Treatment Time | Death Receptor Pathway | Mitochondrial Pathway | ROS/Ca2+ Activity | Bcl-2 Family Regulation | Caspases Activation | Others | Ref. | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Carotenoids | Gracilaria sp. | β-carotene | HepG2 | 5.63 μg/mL | 24 h | ✓ | ✓ | ↑Bax ↓Bcl-2 ↑Bad | 3 | PARP cleavage | [44] | |
| Ishige okamurae | Fucoxanthin | B16F10 | 32.94 μg/mL | ↓Bcl-xL | 3 and 9 | PARP cleavage | [45] | |||||
| Anhui University of Chinese Medicine | HeLa | 0.65 μg/mL | ✓ | ↑Bax ↓Bcl-2 | 3 | [46] | ||||||
| Cladosiphon okamurus | Fucoxanthinol | Saos-2 | 6.16 μg/mL | ↓Bcl-2 ↓Bcl-xL | 3, 8, and 9 | [47] | ||||||
| Codium fragile | Siphonaxanthin | HL-60 | 12.01 μg/mL | ✓ | ↓Bcl-2 | 3 | [48] | |||||
| Phenolic Compounds | Rhodomela confervoides | 1 BDDPM | BEL-7402 | 8.7 μg/mL | 3 and 9 | PARP cleavage | [49] | |||||
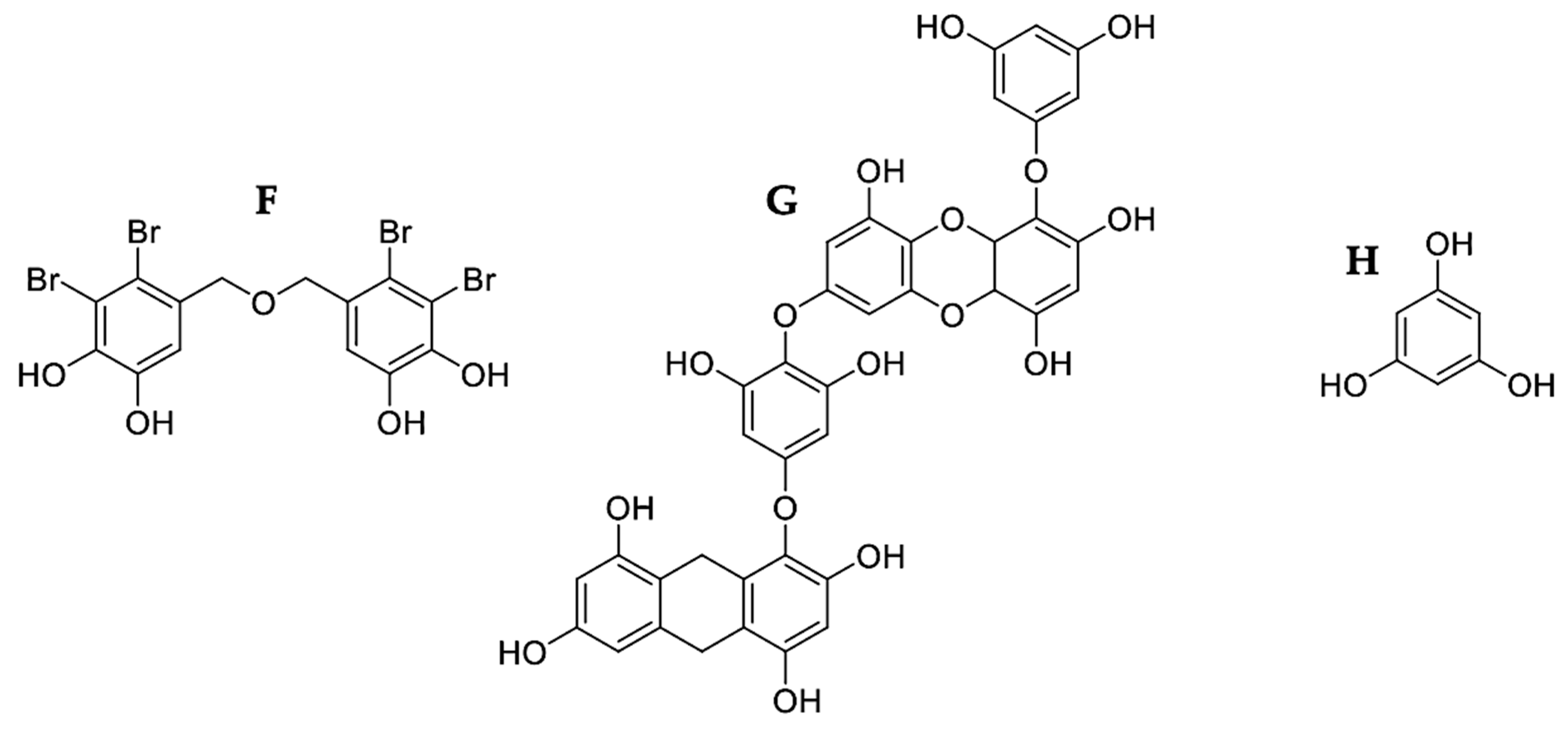
| Ecklonia cava | Dieckol | SKOV3 | 100 μg/mL | ✓ | ✓ | ✓ | ↓Bcl-2 | 3, 8, and 9 | [50] | |||
| AKos Consulting & Solutions | A549 | 25 μg/mL | 3, 8, and 9 | DNA fragmentation Chromatin condensation | [51] | |||||||
| Sigma-Aldrich (St. Louis, MO, USA) | Phloroglucinol | HT-29 | 50 μg/mL | ✓ | ✓ | ↑Bax ↓Bcl-2 ↓Bcl-xL ↑Bad ↑Bid | 3, 8, and 9 | PARP cleavage | [52] | |||
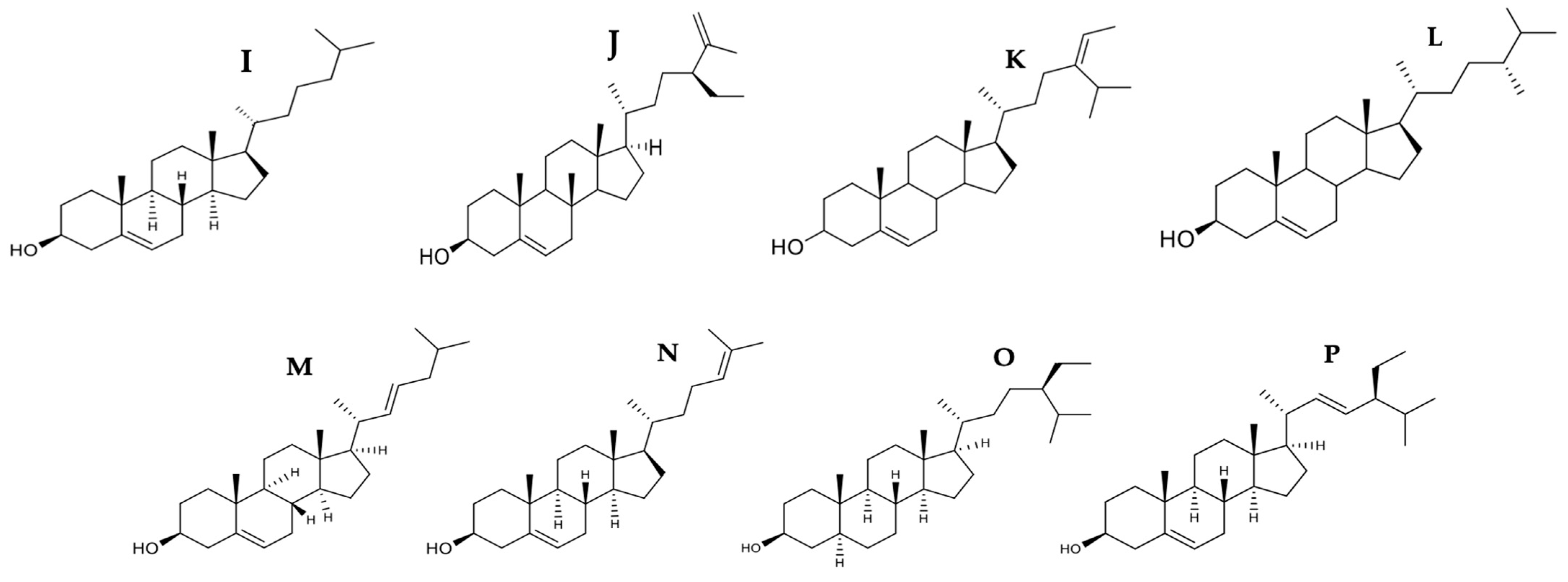
| Phytosterols | Codium fragile | Clerosterol | A2058 | 61.90 μg/mL | ✓ | ↑Bax ↓Bcl-2 | 3 and 9 | DNA fragmentation | [53] | |||
| Marine algae | Fucosterol | HL-60 | 14.19 μg/mL | ✓ | ✓ | 3, 8, and 9 | Presence of apoptotic bodies | [54] | ||||
| Sigma-Aldrich, (St. Louis, MO, USA) | HeLa | 16.50 μg/mL | ✓ | ✓ | PS externalization | [55] | ||||||
| Cayman Chemical Company (Ann Arbor, MI, USA) | ES2 OV90 | 25.75 µg/mL 21.21 µg/mL | ✓ | ✓ | 3 and 9 | [56] | ||||||
| Porphyra dentata | Sterol fraction | 4T1 | 48.3 μg/mL | ✓ | [57] | |||||||
| Glycoproteins | Capsosiphon fulvescens | 2 CF-GP | AGS | 3 μg/mL | ✓ | ✓ | 3, 8, and 9 | [58] | ||||
| Codium decorticatum | 3 GLP | MCF-7 | 60 µg/mL | Nuclear fragmentation | [59] | |||||||
| MDA-MB-231 | 55 μg/mL | ✓ | ✓ | ↑Bax ↓Bcl-2 | 3 and 9 | ↑p53 Chromatin condensation Presence of apoptotic bodies PARP cleavage | [60] | |||||
| Hizikia fusiformis | 4 HFGP | HepG2 | 25 μg/mL | ✓ | ✓ | ↑Bax ↑Bad | [61] | |||||
| Polysaccharides | Enteromorpha intestinalis | 5 EI-SP | HepG2 | 98.4 μg/mL | ✓ | ↑Bax ↓Bcl-2 | 3 and 9 | PARP cleavage Chromatin condensation Nuclear fragmentation | [62] | |||
| Laurencia papillosa | 6 ESC | MDA-MB-231 | 50 µg/mL | ✓ | ↑Bax ↓Bcl-2 | 3, 8, and 9 | ↑p53 | [63] | ||||
| Undaria pinnatifida | Fucoidan | SMMC-7721 | 1000 μg/mL | ✓ | ✓ | ↑Bax ↓Bcl-2 | 3, 8, and 9 | [64] | ||||
| Undaria pinnatifida | PC-3 | 200 μg/mL | ✓ | ↑Bax ↓Bcl-2 | 3, 8, and 9 | PARP cleavage | [65] | |||||
| Sigma-Aldrich (St. Louis, MO, USA) | SKM-1 | 560 μg/mL | ✓ | ✓ | 8 and 9 | [32] | ||||||
| Undaria pinnatifida | DU-145 | 750 µg/mL | ↑Bax ↓Bcl-2 | 9 | PARP cleavage | [66] | ||||||
| Sigma-Aldrich (St. Louis, MO, U.S.A.) | 5637 | 100 μg/mL | ✓ | ✓ | ↑Bax ↓Bcl-2 | ↓c-myc | [67] | |||||
| Sigma-Aldrich (St. Louis, MO, U.S.A.) | MDA-MB-231 | 25 μg/mL | ✓ | ↑Bax ↓Bcl-2 ↓Bcl-xL ↑Bid | 3 and 9 | [68] | ||||||
| Fucus vesiculosus | LM3 | 400 µg/mL | ↑Bax ↓Bcl-2 | 3, 8, and 9 | [69] | |||||||
| Sigma-Aldrich, (St. Louis, MO, USA) | Laminarin | LoVo | 800 μg/mL | ✓ | ✓ | 3 and 9 | [70] | |||||
| Laminaria japonica | LoVo | 800 μg/mL | ✓ | ✓ | ↑Bid ↑tBid | 3, 6, 7, and 9 | [71] | |||||
| Laminaria digitata | ES2 OV90 | 2000 μg/mL | ✓ | ✓ | DNA fragmentation Endoplasmic reticulum stress | [72] | ||||||
| Ulva fasciata | 7 Se-PPC | A549 | 2.5 μg/mL | ✓ | ✓ | ✓ | ↓Bcl-xL ↑Bax ↓Bcl-2 ↑Bid | 3, 8, y 9 | ↑p53 Cell contraction Presence of Apoptotic bodies | [73] | ||
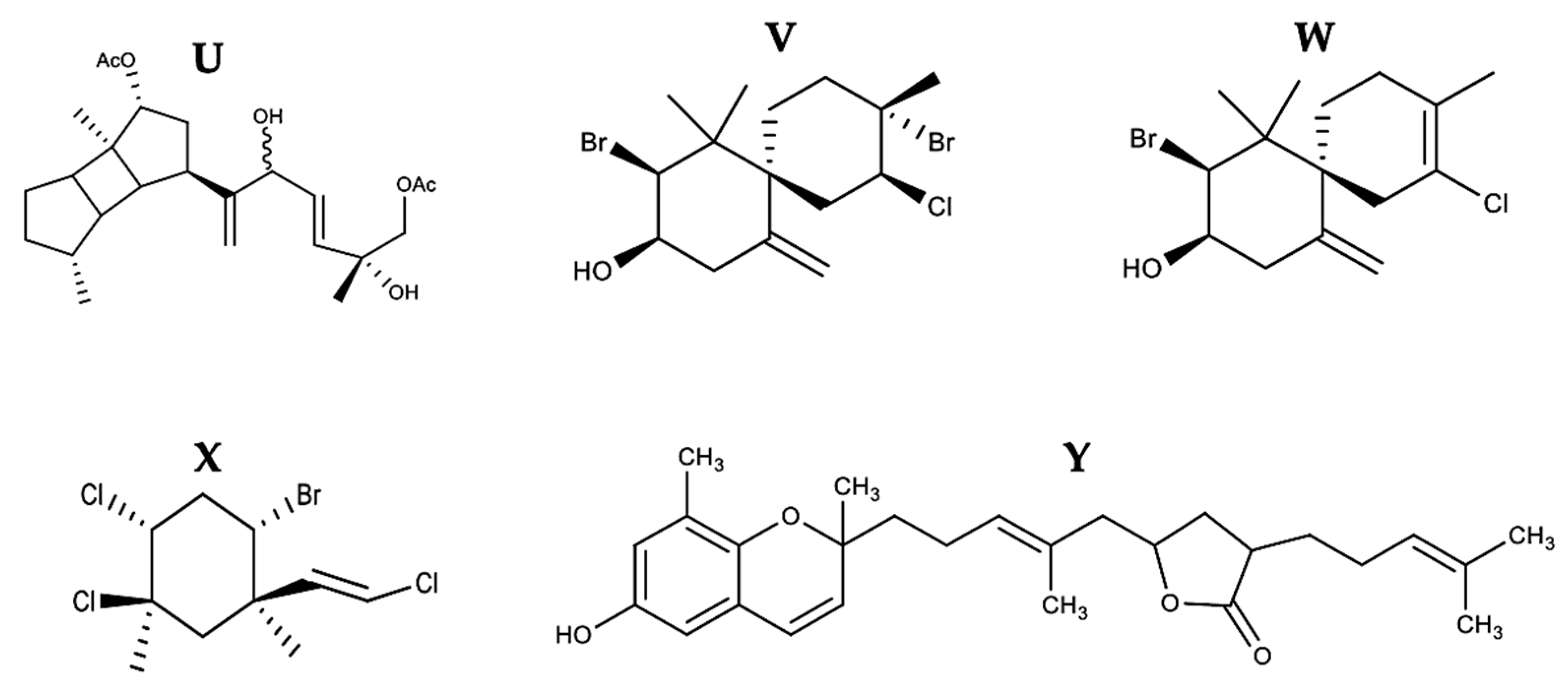
| Terpenes | Stoechospermum marginatum | 8 DDSD | B16F10 | 3.7 μg/mL | ✓ | ✓ | ↑Bax ↓Bcl-2 | 3 and 9 | PS externalization DNA fragmentation Nuclear condensation | [74] | ||
| Laurencia dendroidea | (-)-Elatol | 2.52 µg/mL | 2, 4, 6, and 8 | [75] | ||||||||
| Laurencia dendroidea | + (-) Obtusol | Colo-205 | 1.35 µg/mL | 6 | [75] | |||||||
| Pterocladiella capillacea | Mertensene | HT-29 | 90 μg/mL | ✓ | 3 | PARP cleavage | [76] | |||||
| Sargassum macrocarpum | 9 TTB | MDA-MB-231, A549, and HCT116 | 25.35, 20.28, and 19.86 μg/mL | ✓ | ↓Bcl-2 | 3 | PARP cleavage | [77] |
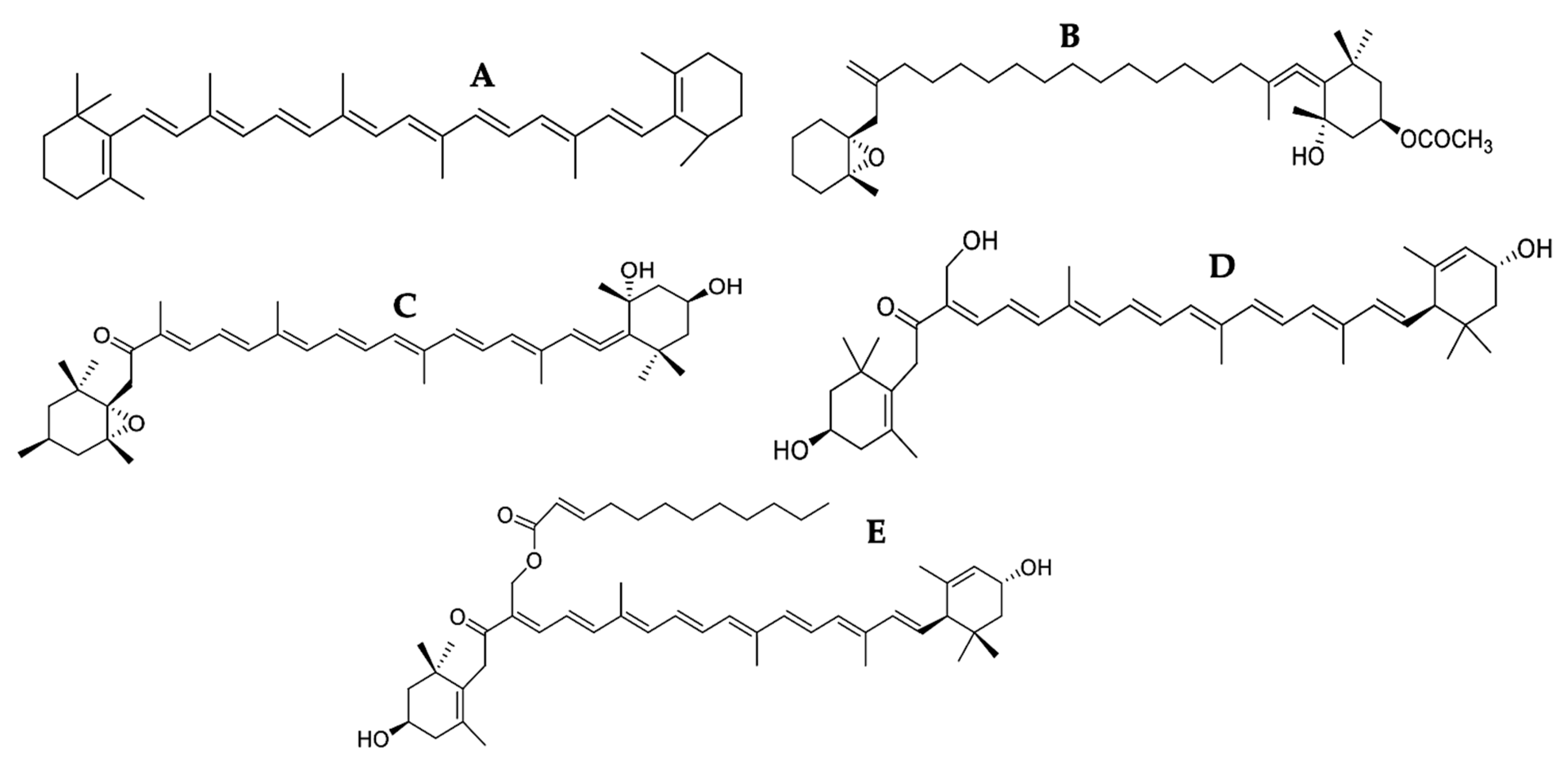
3.1. Carotenoids
3.2. Phenolic Compounds
3.3. Phytosterols
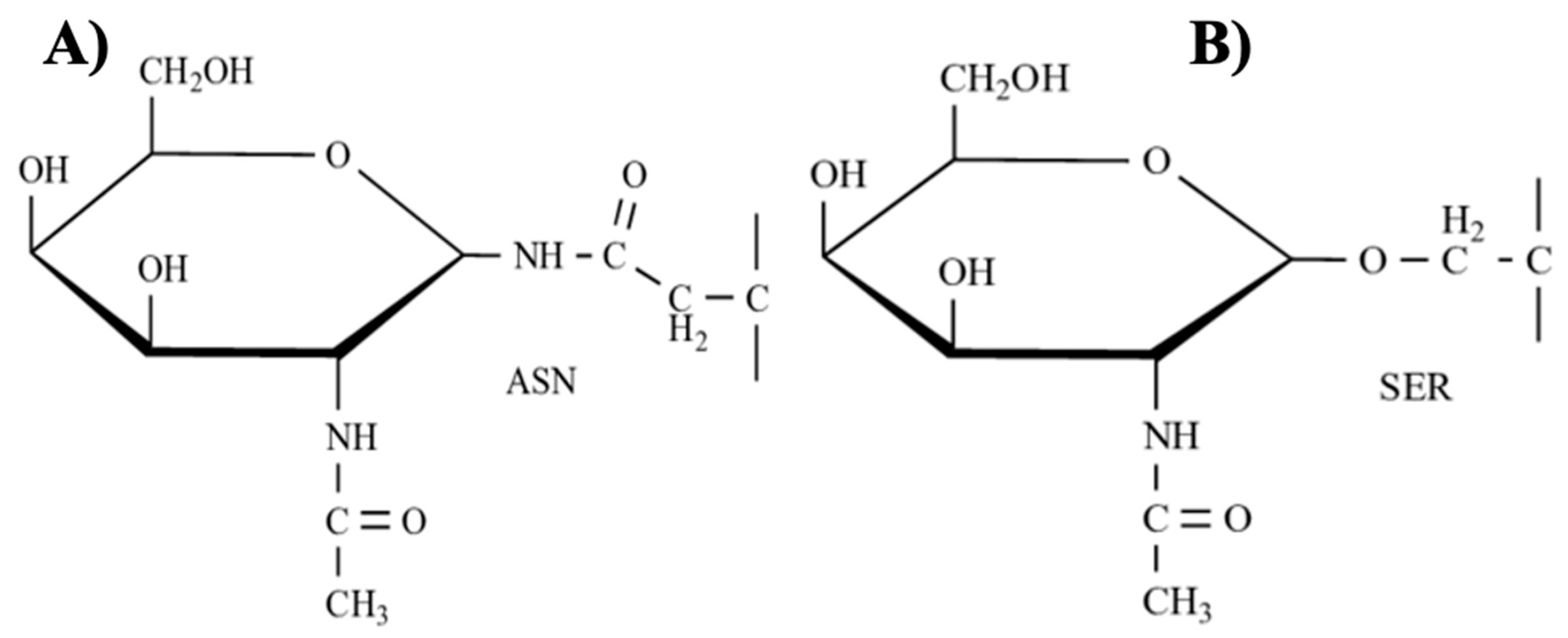
3.4. Glycoproteins
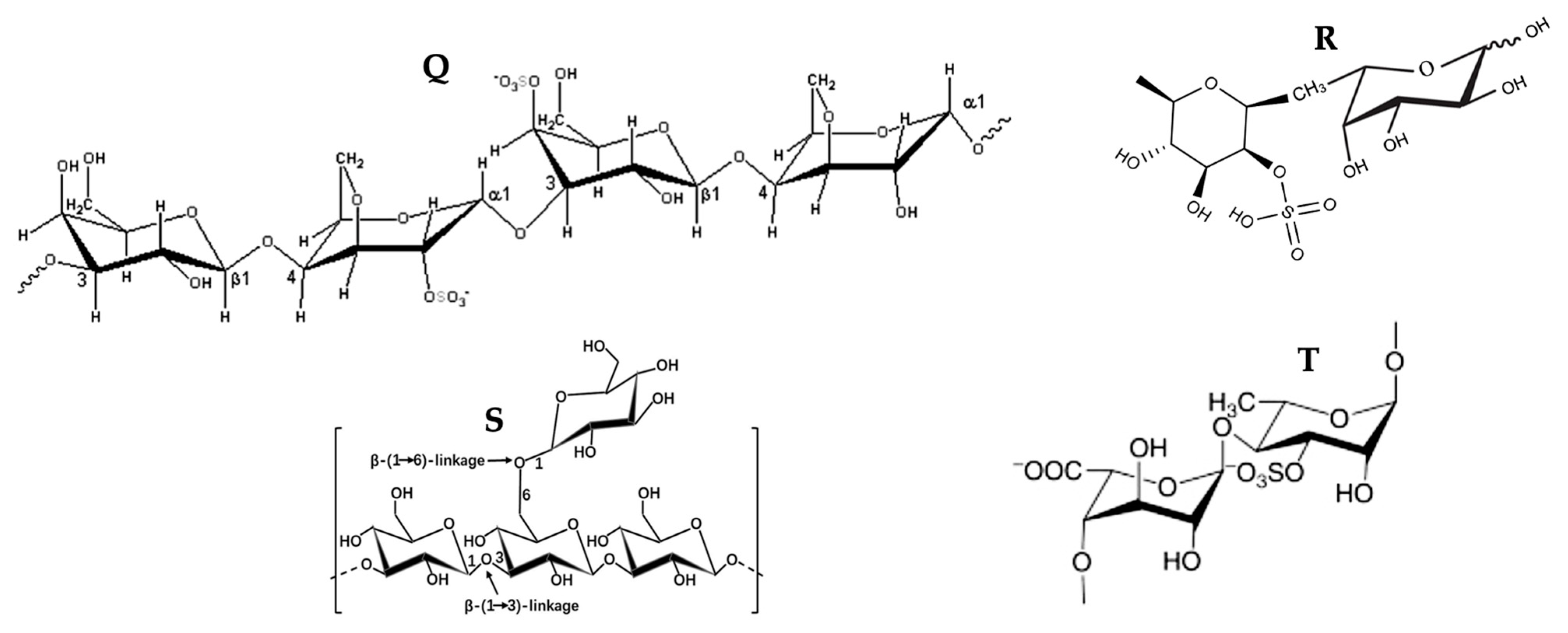
3.5. Polysaccharides
3.6. Terpenes
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- MacCarthy-Morrogh, L.; Martin, P. The hallmarks of cancer are also the hallmarks of wound healing. Sci. Signal 2020, 13, 648. [Google Scholar] [CrossRef] [PubMed]
- Osuna-Ruiz, I.; Hurtado-Oliva, M.Á.; Nieves-Soto, M.; Manzano-Sarabia, M.M.; Burgos-Hernández, A.; Lizardi-Mendoza, J.; Hernández-Garibay, E. Algas marinas: Potencial fuente de compuestos contra el cáncer. Ciencia 2016, 1–8. [Google Scholar]
- Martins, A.; Vieira, H.; Gaspar, H.; Santos, S. Marketed Marine Natural Products in the Pharmaceutical and Cosmeceutical Industries: Tips for Success. Mar. Drugs 2014, 12, 1066–1101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Weelden, G.; Bobiński, M.; Okła, K.; Van Weelden, W.J.; Romano, A.; Pijnenborg, J.M.A. Fucoidan structure, and activity in relation to anti-cancer mechanisms. Mar. Drugs 2019, 17, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, K.G.; Fitzgerald, G.F.; Stanton, C.; Ross, R.P. Looking beyond the terrestrial: The potential of seaweed derived bioactives to treat non-communicable diseases. Mar. Drugs 2016, 14, 60. [Google Scholar] [CrossRef] [Green Version]
- Brown, E.S.; Allsopp, P.J.; Magee, P.J.; Gill, C.I.; Nitecki, S.; Strain, C.R.; McSorley, E.M. Seaweed and human health. Nutr. Rev. 2014, 72, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Ercolano, G.; De Cicco, P.; Ianaro, A. New Drugs from the Sea: Pro-Apoptotic Activity of Sponges and Algae Derived Compounds. Mar. Drugs 2019, 17, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quitral, V.R.; Morales, C.G.; Sepúlveda, M.L.; Schwartz, M.M. Nutritional and health properties of seaweeds and its potential as a functional ingredient. Rev. Child. Nutr. 2012, 39, 196–202. [Google Scholar] [CrossRef]
- Alves, C.; Pinteus, S.; Horta, A.; Pedrosa, R. High citotoxicity and anti-proliferative activity of extracts on an in vitro model of human hepatocellular carcinoma. SpringerPlus 2016, 5, 1339–1351. [Google Scholar] [CrossRef] [Green Version]
- Patra, J.K.; Lee, S.W.; Park, J.G.; Baek, K.H. Antioxidant and Antibacterial Properties of Essential Oil Extracted from an Edible Seaweed Undaria Pinnatifida. J. Food Biochem. 2017, 41, e12278. [Google Scholar] [CrossRef]
- D’Arcy, M.S. Cell death: A review of the major forms of apoptosis, necrosis and autophagy. Cell. Biol. Int. 2019, 43, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Garijo, A. When dying is not the end: Apoptotic caspases as drivers of proliferation. Semin. Cell. Dev. Biol. 2018, 82, 86–95. [Google Scholar] [CrossRef]
- Boice, A.; Bouchier-Hayes, L. Targeting apoptotic caspases in cancer. Biochim. Biophys. Acta Mol. Cell Res. 2020, 1867, 118688. [Google Scholar] [CrossRef]
- Redza-Dutordoir, M.; Averill-Bates, D.A. Activation of apoptosis signalling pathways by reactive oxygen species. Biochim. Biophys. Acta Mol. Cell Res. 2016, 1863, 2977–2992. [Google Scholar] [CrossRef]
- Kumar, S.R.; Hosokawa, M.; Miyashita, K. Fucoxanthin: A marine carotenoid exerting anti-cancer effects by affecting multiple mechanisms. Mar. Drugs 2013, 11, 5130–5147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Figueiredo-Pereira, C.; Oudot, C.; Vieira, H.L.A.; Brenner, C. Mitochondrion: A Common Organelle for Distinct Cell Deaths? Int. Rev. Cell Mol. Biol. 2017, 331, 245–287. [Google Scholar] [CrossRef]
- Jutinico, S.A.P.; Mantilla, G.A.; Sánchez, M.R.M. Regulation of the bcl-2 to family in cells infected with Chlamydia Trachomatis. NOVA 2015, 13, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Warren, C.F.A.; Wong-Brown, M.W.; Bowden, N.A. BCL-2 family isoforms in apoptosis and cancer. Cell Death Dis. 2019, 10, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aubrey, B.J.; Kelly, G.L.; Janic, A.; Herold, M.J.; Strasser, A. How does p53 induce apoptosis and how does this relate to p53-mediated tumour suppression? Cell Death Differ. 2018, 25, 104–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giam, M.; Huang, D.C.S.; Bouillet, P. BH3-only proteins and their roles in programmed cell death. Oncogene 2008, 27, S128–S136. [Google Scholar] [CrossRef] [PubMed]
- Fairlie, W.D.; Tran, S.; Lee, E.F. Crosstalk between apoptosis and autophagy signaling pathways. Int. Rev. Cell Mol. Biol. 2020, 352, 115–158. [Google Scholar] [CrossRef] [PubMed]
- Quarleri, J.; Cevallos, C.; Delpino, M.V. Apoptosis in infectious diseases as a mechanism of immune evasion and survival. Adv. Protein Chem. Struct. Biol. 2021, 125, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Ma, K.; Liu, X.; Chen, S.-H.; Li, P.; Yu, Y.; Leung, A.K.L.; Yu, X. Truncated PARP1 mediates ADP-ribosylation of RNA polymerase III for apoptosis. Cell. Discov. 2022, 8, 3. [Google Scholar] [CrossRef] [PubMed]
- Usmani, H.; Hussain, S.A.; Sheikh, A. PARP inhibitors: Current status and implications for anticancer therapeutics. Infect. Agents Cancer 2013, 8, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.J.; Dawson, T.M.; Dawson, V.L. PARP and the Release of Apoptosis-Inducing Factor from Mitochondria. In Madame Curie Bioscience Database; Landes Bioscience: Austin, TX, USA, 2013; pp. 1–29. [Google Scholar]
- Ramírez-García, M.Á.; Márquez-González, H.; Barranco-Lampón, G.; López-Aguilar, J.E. Bcl-2: Su papel en el ciclo celular, apoptosis y cáncer. Residente 2014, 9, 84–94. [Google Scholar]
- Jan, R.; Chaudhry, G.-e.-S. Understanding Apoptosis and Apoptotic Pathways Targeted Cancer Therapeutics. Adv. Pharm. Bull. 2019, 9, 205–218. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.-F.; Hu, Y.-C.; Kang, B.-H.; Tseng, Y.-K.; Wu, P.-C.; Liang, C.-C.; Hou, Y.-Y.; Fu, T.-Y.; Liou, H.-H.; Hsieh, I.-C.; et al. Expression levels of cleaved caspase-3 and caspase-3 in tumorigenesis and prognosis of oral tongue squamous cell carcinoma. PLoS ONE 2017, 12, e0180620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shalini, S.; Dorstyn, L.; Dawar, S.; Kumar, S. Old, new and emerging functions of caspases. Cell Death Differ. 2015, 22, 526–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uddin, M.S.; Upaganlawar, A.B. Oxidative Stress and Antioxidant Defense: Biomedical Value in Health and Diseases. In Nova Science & Health; Nova Science Pub Inc.: Hauppauge, NY, USA, 2019; ISBN 978-1-53615-687-4. [Google Scholar]
- Wei, C.; Xiao, Q.; Kuang, X.; Zhang, T.; Yang, Z.; Wang, L. Fucoidan inhibits proliferation of the SKM-1 acute myeloid leukaemia cell line via the activation of apoptotic pathways and production of reactive oxygen species. Mol. Med. Rep. 2015, 12, 6649–6655. [Google Scholar] [CrossRef] [Green Version]
- Giorgi, C.; Marchi, S.; Simoes, I.C.M.; Ren, Z.; Morciano, G.; Perrone, M.; Patalas-Krawczyk, P.; Borchard, S.; Jędrak, P.; Pierzynowska, K.; et al. Mitochondria and Reactive Oxygen Species in Aging and Age-Related Diseases. Int. Rev. Cell. Mol. Biol. 2018, 340, 209–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mani, S.; Swargiary, G.; Singh, K.K. Natural agents targeting mitochondria in cancer. Int. J. Mol. Sci. 2020, 21, 96992. [Google Scholar] [CrossRef] [PubMed]
- Ježek, J.; Cooper, K.F.; Strich, R. Reactive Oxygen Species and Mitochondrial Dynamics: The Yin and Yang of Mitochondrial Dysfunction and Cancer Progression. Antioxidants 2018, 7, 13. [Google Scholar] [CrossRef]
- Hernansanz-Agustín, P.; Enríquez, J.A. Generation of Reactive Oxygen Species by Mitochondria. Antioxidants 2021, 10, 415. [Google Scholar] [CrossRef] [PubMed]
- Srinivas, U.S.; Tan, B.W.Q.; Vellayappan, B.A.; Jeyasekharan, A.D. ROS and the DNA damage response in cancer. Redox Biol. 2019, 25, 101084. [Google Scholar] [CrossRef]
- Davalli, P.; Marverti, G.; Lauriola, A.; D’Arca, D. Targeting Oxidatively Induced DNA Damage Response in Cancer: Opportunities for Novel Cancer Therapies. Oxid. Med. Cell Longev. 2018, 2018, 2389523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, P.D.; Huang, B.-W.; Tsuji, G. Reactive oxygen species (ROS) homeostasis and redox regulation in cellular signaling. Cell Signal. 2012, 24, 981–990. [Google Scholar] [CrossRef] [Green Version]
- Begum, R.; Howlader, S.; Mamun-Or-Rashid, A.N.M.; Rafiquzzaman, S.M.; Ashraf, G.M.; Albadrani, G.M.; Sayed, A.A.; Peluso, I.; Abdel-Daim, M.M.; Uddin, M.S. Antioxidant and Signal-Modulating Effects of Brown Seaweed-Derived Compounds against Oxidative Stress-Associated Pathology. Oxid. Med. Cell Longev. 2021, 2021, 9974890. [Google Scholar] [CrossRef] [PubMed]
- Boik, J. Natural Compounds in Cancer Therapy; Oregon Medical Press: Princeton, MN, USA, 2001. [Google Scholar]
- Suffness, M.; Pezzuto, J.M. Assays related to cancer drug discovery. In Methods in Plant Biochemistry: Assays for Bioactivity; Hostettmann, K., Ed.; Academic Press: London, UK, 1990; pp. 71–133. [Google Scholar]
- Ayoub, I.M.; El-Shazly, M.; Lu, M.-C.; Singab, A.N.B. Antimicrobial and cytotoxic activities of the crude extracts of Dietes bicolor leaves, flowers and rhizomes. South Afr. J. Bot. 2014, 95, 97–101. [Google Scholar] [CrossRef] [Green Version]
- Kavalappa, Y.P.; Rudresh, D.U.; Gopal, S.S.; Shivarudrappa, A.H.; Stephen, N.M.; Rangiah, K.; Ponesakki, G. β-carotene isolated from the marine red alga, Gracillaria sp. potently attenuates the growth of human hepatocellular carcinoma (HepG2) cells by modulating multiple molecular pathways. J. Funct. Foods. 2019, 52, 165–176. [Google Scholar] [CrossRef]
- Kim, K.-N.; Ahn, G.; Heo, S.-J.; Kang, S.-M.; Kang, M.-C.; Yang, H.-M.; Kim, D.; Roh, S.-W.; Kim, S.-K.; Jeon, B.-T.; et al. Inhibition of tumor growth in vitro and in vivo by fucoxanthin against melanoma B16F10 cells. Environ. Toxicol. Pharmacol. 2013, 35, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Ye, G.; Lu, Q.; Zhao, W.; Du, D.; Jin, L.; Liu, Y. Fucoxanthin induces apoptosis in human cervical cancer cell line HeLa via PI3K/Akt pathway. Tumour Biol. 2014, 35, 11261–11267. [Google Scholar] [CrossRef] [PubMed]
- Rokkaku, T.; Kimura, R.; Ishikawa, C.; Yasumoto, T.; Senba, M.; Kanaya, F.; Mori, N. Anticancer effects of marine carotenoids, fucoxanthin and its deacetylated product, fucoxanthinol, on osteosarcoma. Int. J. Oncol. 2013, 43, 1176–1186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganesan, P.; Matsubara, K.; Ohkubo, T.; Tanaka, Y.; Noda, K.; Sugawara, T.; Hirata, T. Anti-angiogenic effect of siphonaxanthin from green alga, Codium fragile. Phytomedicine 2010, 17, 1140–1144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, N.; Luo, J.; Jiang, B.; Wang, L.; Wang, S.; Wang, C.; Fu, C.; Li, J.; Shi, D. Marine bromophenol bis (2,3-dibromo-4,5-dihydroxy-phenyl)-methane inhibits the proliferation, migration, and invasion of hepatocellular carcinoma cells via modulating β1-Integrin/FAK signaling. Mar. Drugs 2015, 13, 1010–1025. [Google Scholar] [CrossRef]
- Ahn, J.-H.; Yang, Y.-I.; Lee, K.-T.; Choi, J.-H. Dieckol, isolated from the edible brown algae Ecklonia cava induces apoptosis of ovarian cancer cells and inhibits tumor xenograft growth. J. Cancer Res. Clin. Oncol. 2015, 141, 255–268. [Google Scholar] [CrossRef]
- Wang, C.-H.; Li, X.-F.; Jin, L.-F.; Zhao, Y.; Zhu, G.-J.; Shen, W.-Z. Dieckol inhibits non-small-cell lung cancer cell proliferation and migration by regulating the PI3K/AKT signaling pathway. J. Biochem. Mol. Toxicol. 2019, 33, e22346. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.-H.; Kim, I.-H.; Nam, T.-J. Phloroglucinol induces apoptosis via apoptotic signaling pathways in HT-29 colon cancer cells. Oncol. Rep. 2014, 32, 1341–1346. [Google Scholar] [CrossRef] [Green Version]
- Kim, A.D.; Lee, Y.; Kang, S.-H.; Kim, G.Y.; Kim, H.S.; Hyun, J.W. Cytotoxic Effect of Clerosterol Isolated from Codium fragile on A2058 Human Melanoma Cells. Mar. Drugs 2013, 11, 418–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, Y.-B.; Ji, C.-F.; Yue, L. Study on human promyelocytic leukemia HL-60 cells apoptosis induced by fucosterol. Biomed. Mater. Eng. 2014, 24, 845–851. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Li, J.; Chen, A.; Li, Y.; Xia, M.; Guo, P.; Yao, S.; Chen, S. Fucosterol exhibits selective antitumor anticancer activity against HeLa human cervical cell line by inducing mitochondrial mediated apoptosis, cell cycle migration inhibition and downregulation of m-TOR/PI3K/Akt signalling pathway. Oncol. Lett. 2018, 15, 3458–3463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, H.; Lee, J.-Y.; Song, G.; Lim, W. Fucosterol Suppresses the Progression of Human Ovarian Cancer by Inducing Mitochondrial Dysfunction and Endoplasmic Reticulum Stress. Mar. Drugs 2020, 18, 261. [Google Scholar] [CrossRef] [PubMed]
- Kazłowska, K.; Lin, H.-T.; Chang, S.-H.; Tsai, G.-J. In Vitro and In Vivo Anticancer Effects of Sterol Fraction from Red Algae Porphyra dentata. Evid. Based Complement. Alternat. Med. 2013, 2013, 493869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.-M.; Kim, I.-H.; Nam, T.-J. Induction of apoptosis signaling by glycoprotein of Capsosiphon fulvescens in human gastric cancer (AGS) cells. Nutr. Cancer 2012, 64, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Senthilkumar, D.; Jayanthi, S. Partial characterization and anticancer activities of purified glycoprotein extracted from green seaweed Codium decorticatum. J. Funct. Foods 2016, 25, 323–332. [Google Scholar] [CrossRef]
- Thangam, R.; Senthilkumar, D.; Suresh, V.; Sathuvan, M.; Sivasubramanian, S.; Pazhanichamy, K.; Gorlagunta, P.K.; Kannan, S.; Gunasekaran, P.; Rengasamy, R.; et al. Induction of ROS-dependent mitochondria-mediated intrinsic apoptosis in MDA-MB-231 cells by glycoprotein from Codium decorticatum. J. Agric. Food Chem. 2014, 62, 3410–3421. [Google Scholar] [CrossRef]
- Ryu, J.; Hwang, H.-J.; Kim, I.-H.; Nam, T.-J. Mechanism of Inhibition of HepG2 Cell Proliferation by a Glycoprotein from Hizikia fusiformis. Korean J. Fish Aquat. Sci. 2012, 45, 553–560. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Chen, Y.; Wang, J.; Liu, Z.; Zhao, S. Antitumor activity of a sulfated polysaccharide from Enteromorpha intestinalis targeted against hepatoma through mitochondrial pathway. Tumour Biol. 2014, 35, 1641–1647. [Google Scholar] [CrossRef] [PubMed]
- Murad, H.; Hawat, M.; Ekhtiar, A.; AlJapawe, A.; Abbas, A.; Darwish, H.; Sbenati, O.; Ghannam, A. Induction of G1-phase cell cycle arrest and apoptosis pathway in MDA-MB-231 human breast cancer cells by sulfated polysaccharide extracted from Laurencia papillosa. Cancer Cell Int. 2016, 16, 39. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Wang, P.; Wang, H.; Li, Q.; Teng, H.; Liu, Z.; Yang, W.; Hou, L.; Zou, X. Fucoidan derived from Undaria pinnatifida induces apoptosis in human hepatocellular carcinoma SMMC-7721 cells via the ROS-mediated mitochondrial pathway. Mar. Drugs 2013, 11, 1961–1976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boo, H.-J.; Hong, J.-Y.; Kim, S.-C.; Kang, J.-I.; Kim, M.-K.; Kim, E.-J.; Hyun, J.-W.; Koh, Y.-S.; Yoo, E.-S.; Kwon, J.-M.; et al. The anticancer effect of fucoidan in PC-3 prostate cancer cells. Mar. Drugs 2013, 11, 2982–2999. [Google Scholar] [CrossRef] [Green Version]
- Choo, G.-S.; Lee, H.-N.; Shin, S.-A.; Kim, H.-J.; Jung, J.-Y. Anticancer Effect of Fucoidan on DU-145 Prostate Cancer Cells through Inhibition of PI3K/Akt and MAPK Pathway Expression. Mar. Drugs 2016, 14, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, M.H.; Lee, D.-S.; Jeong, J.-W.; Hong, S.-H.; Choi, I.-W.; Cha, H.-J.; Kim, S.; Kim, H.-S.; Park, C.; Kim, G.-Y.; et al. Fucoidan Induces ROS-Dependent Apoptosis in 5637 Human Bladder Cancer Cells by Downregulating Telomerase Activity via Inactivation of the PI3K/Akt Signaling Pathway. Drug Dev. Res. 2017, 78, 37–48. [Google Scholar] [CrossRef]
- Xue, M.; Ji, X.; Xue, C.; Liang, H.; Ge, Y.; He, X.; Zhang, L.; Bian, K.; Zhang, L. Caspase-dependent and caspase-independent induction of apoptosis in breast cancer by fucoidan via the PI3K/AKT/GSK3β pathway in vivo and in vitro. Biomed. Pharmacother. 2017, 94, 898–908. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y.; Li, J.; Jing, X.; Ding, X.; Yu, Y.; Zhao, Q. Fucoidan Induces Apoptosis and Inhibits Proliferation of Hepatocellular Carcinoma via the p38 MAPK/ERK and PI3K/Akt Signal Pathways. Cancer Manag. Res. 2020, 12, 1713–1723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, Y.B.; Ji, C.F.; Zhang, H. Laminarin Induces Apoptosis of Human Colon Cancer LOVO Cells through a Mitochondrial Pathway. Molecules 2012, 17, 9947–9960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, C.-F.; Ji, Y.-B. Laminarin-induced apoptosis in human colon cancer LoVo cells. Oncol. Lett. 2014, 7, 1728–1732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, H.; Song, G.; Lee, J.-Y.; Hong, T.; Chang, M.-J.; Lim, W. Laminarin-Derived from Brown Algae Suppresses the Growth of Ovarian Cancer Cells via Mitochondrial Dysfunction and ER Stress. Mar. Drugs 2020, 18, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Zhong, Y.; Luo, H.; Yang, Y. Selenium-Containing Polysaccharide-Protein Complex in Se-Enriched Ulva fasciata Induces Mitochondria-Mediated Apoptosis in A549 Human Lung Cancer Cells. Mar. Drugs 2017, 15, 215. [Google Scholar] [CrossRef] [Green Version]
- Velatooru, L.R.; Baggu, C.B.; Janapala, V.R. Spatane diterpinoid from the brown algae, Stoechospermum marginatum induces apoptosis via ROS induced mitochondrial mediated caspase dependent pathway in murine B16F10 melanoma cells. Mol. Carcinog. 2016, 55, 2222–2235. [Google Scholar] [CrossRef]
- Barcellos Marini, M.; Rodrigues de Freitas, W.; Lacerda da Silva Machado, F.; Correa Ramos Leal, I.; Ribeiro Soares, A.; Masahiko Kanashiro, M.; Frazão Muzitano, M. Cytotoxic activity of halogenated sesquiterpenes from Laurencia dendroidea. Phytother. Res. 2018, 32, 1119–1125. [Google Scholar] [CrossRef] [PubMed]
- Tarhouni-Jabberi, S.; Zakraoui, O.; Ioannou, E.; Riahi-Chebbi, I.; Haoues, M.; Roussis, V.; Kharrat, R.; Essafi-Benkhadir, K. Mertensene, a Halogenated Monoterpene, Induces G2/M Cell Cycle Arrest and Caspase Dependent Apoptosis of Human Colon Adenocarcinoma HT29 Cell Line through the Modulation of ERK-1/-2, AKT and NF-κB Signaling. Mar. Drugs 2017, 15, 221. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.K.; Kim, J.; Lee, K.M.; Choi, Y.-J.; Ye, B.-R.; Kim, M.-S.; Ko, S.-G.; Lee, S.-H.; Kang, D.-H.; Heo, S.-J. Tuberatolide B suppresses cancer progression by promoting ROS-mediated inhibition of STAT3 signaling. Mar. Drugs 2017, 15, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Méresse, S.; Fodil, M.; Fleury, F.; Chénais, B. Fucoxanthin, a Marine-Derived Carotenoid from Brown Seaweeds and Microalgae: A Promising Bioactive Compound for Cancer Therapy. Int. J. Mol. Sci. 2020, 21, 39273. [Google Scholar] [CrossRef]
- McNulty, H.P.; Byun, J.; Lockwood, S.F.; Jacob, R.F.; Mason, R.P. Differential effects of carotenoids on lipid peroxidation due to membrane interactions: X-ray diffraction analysis. Biochim. Biophys. Acta 2007, 1768, 167–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, J.; Song, M.-H.; Oh, J.-W.; Keum, Y.-S.; Saini, R.K. Pro-Oxidant Actions of Carotenoids in Triggering Apoptosis of Cancer Cells: A Review of Emerging Evidence. Antioxidants 2020, 9, 532. [Google Scholar] [CrossRef]
- Quitério, E.; Soares, C.; Ferraz, R.; Delerue-Matos, C.; Grosso, C. Marine Health-Promoting Compounds: Recent Trends for Their Characterization and Human Applications. Foods 2021, 10, 3100. [Google Scholar] [CrossRef]
- Jacobsen, C.; Sørensen, A.-D.M.; Holdt, S.L.; Akoh, C.C.; Hermund, D.B. Source, Extraction, Characterization, and Applications of Novel Antioxidants from Seaweed. Annu. Rev. Food Sci. Technol. 2019, 10, 541–568. [Google Scholar] [CrossRef]
- Martin, L.J. Fucoxanthin and Its Metabolite Fucoxanthinol in Cancer Prevention and Treatment. Mar. Drugs 2015, 13, 4784–4798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Linnewiel-Hermoni, K.; Motro, Y.; Miller, Y.; Levy, Y.; Sharoni, Y. Carotenoid derivatives inhibit nuclear factor kappa B activity in bone and cancer cells by targeting key thiol groups. Free Radic. Biol. Med. 2014, 75, 105–120. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, R.; Mishra, D.P. Trailing TRAIL Resistance: Novel Targets for TRAIL Sensitization in Cancer Cells. Front. Oncol. 2015, 5, 69. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, T.; Ganesan, P.; Li, Z.; Manabe, Y.; Hirata, T. Siphonaxanthin, a green algal carotenoid, as a novel functional compound. Mar. Drugs 2014, 12, 3660–3668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Silva Port’s, P.; Chisté, R.C.; Godoy, H.T.; Prado, M.A. The phenolic compounds and the antioxidant potential of infusion of herbs from the Brazilian Amazonian region. Food Res. Int. 2013, 53, 875–881. [Google Scholar] [CrossRef]
- Cotas, J.; Leandro, A.; Monteiro, P.; Pacheco, D.; Figueirinha, A.; Gonçalves, A.M.M.; da Silva, G.J.; Pereira, L. Seaweed Phenolics: From Extraction to Applications. Mar. Drugs 2020, 18, 384. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Lopez, C.; Pereira, A.G.; Lourenço-Lopes, C.; Garcia-Oliveira, P.; Cassani, L.; Fraga-Corral, M.; Prieto, M.A.; Simal-Gandara, J. Main bioactive phenolic compounds in marine algae and their mechanisms of action supporting potential health benefits. Food Chem. 2021, 341, 128262. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Zhao, X.; Xu, J.; Li, C.; Yu, Y.; Wang, W.; Zhu, L. The Protective Effect of Dietary Phytosterols on Cancer Risk: A Systematic Meta-Analysis. J. Oncol. 2019, 2019, 7479518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mignard, V.; Lalier, L.; Paris, F.; Vallette, F.M. Bioactive lipids and the control of Bax pro-apoptotic activity. Cell Death Dis. 2014, 5, e1266. [Google Scholar] [CrossRef] [Green Version]
- Corcos, L.; Le Jossic-Corcos, C. Statins: Perspectives in cancer therapeutics. Dig. Liver Dis. 2013, 45, 795–802. [Google Scholar] [CrossRef] [Green Version]
- Franco, D.; Henao, Y.; Monsalve, M.; Gutiérrez, F.; Hincapie, J.; Amariles, P. Interacciones medicamentosas de agentes hipolipemiantes: Aproximación para establecer y valorar su relevancia clínica: Revisión estructurada. Farm. Hosp. 2013, 37, 539–557. [Google Scholar] [CrossRef] [PubMed]
- Poli, A.; Marangoni, F.; Corsini, A.; Manzato, E.; Marrocco, W.; Martini, D.; Medea, G.; Visioli, F. Phytosterols, Cholesterol Control, and Cardiovascular Disease. Nutrients 2021, 13, 2810. [Google Scholar] [CrossRef]
- Abdul, Q.A.; Choi, R.J.; Jung, H.A.; Choi, J.S. Health benefit of fucosterol from marine algae: A review. J. Sci. Food Agric. 2016, 96, 1856–1866. [Google Scholar] [CrossRef] [PubMed]
- Koutsaviti, A.; Ioannou, E.; Roussis, V. Bioactive Seaweed Substances. In Bioactive Seaweeds for Food Applications: Natural Ingredients for Healthy Diets; Qin, Y., Ed.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 25–52. [Google Scholar] [CrossRef]
- Idler, D.R.; Saito, A.; Wiseman, P. Sterols in red algae (Rhodophyceae). Steroids 1968, 11, 465–473. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Machado, D.I.; López-Hernández, J.; Paseiro-Losada, P.; López-Cervantes, J. An HPLC method for the quantification of sterols in edible seaweeds. Biomed. Chromatogr. 2004, 18, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Awad, A.B.; Fink, C.S. Phytosterols as anticancer dietary components: Evidence and mechanism of action. J. Nutr. 2000, 130, 2127–2130. [Google Scholar] [CrossRef] [Green Version]
- Woyengo, T.A.; Ramprasath, V.R.; Jones, P.J.H. Anticancer effects of phytosterols. Eur. J. Clin. Nutr. 2009, 63, 813–820. [Google Scholar] [CrossRef] [Green Version]
- Vinaiphat, A.; Kwan Sze, S. Advances in extracellular vesicles analysis. In Advances in Clinical Chemistry; Makowski, G.S., Ed.; Elsevier: Amsterdam, The Netherlands, 2020; Volume 97, pp. 73–116. [Google Scholar] [CrossRef]
- Sun, F.; Wu, R. Systematic and site-specific analysis of N-glycoproteins on the cell surface by integrating bioorthogonal chemistry and MS-based proteomics. Methods Enzymol. 2019, 626, 223–247. [Google Scholar] [CrossRef]
- Ahn, Y.H.; Kim, J.Y.; Yoo, J.S. Quantitative mass spectrometric analysis of glycoproteins combined with enrichment methods. Mass Spectrom. Rev. 2015, 34, 148–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swindall, A.F.; Bellis, S.L. Sialylation of the Fas death receptor by ST6Gal-I provides protection against Fas-mediated apoptosis in colon carcinoma cells. J. Biol. Chem. 2011, 286, 22982–22990. [Google Scholar] [CrossRef] [Green Version]
- Peixoto, A.; Relvas-Santos, M.; Azevedo, R.; Santos, L.L.; Ferreira, J.A. Protein Glycosylation and Tumor Microenvironment Alterations Driving Cancer Hallmarks. Front. Oncol. 2019, 9, 380. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Liu, Y.; Jin, K.; Pang, Z. Polysaccharide nanoparticles for cancer drug targeting. In Polysaccharide Carriers for Drug Delivery; Maiti, S., Jana, S., Eds.; Matthew Deans: Oxford, UK, 2019; pp. 365–396. [Google Scholar] [CrossRef]
- Mohammed, A.S.A.; Naveed, M.; Jost, N. Polysaccharides; Classification, Chemical Properties, and Future Perspective Applications in Fields of Pharmacology and Biological Medicine (A Review of Current Applications and Upcoming Potentialities). J. Polym. Environ. 2021, 29, 2359–2371. [Google Scholar] [CrossRef] [PubMed]
- Pei, Y.; Yang, S.; Xiao, Z.; Zhou, C.; Hong, P.; Qian, Z.J. Structural Characterization of Sulfated Polysaccharide Isolated from Red Algae (Gelidium crinale) and Antioxidant and Anti-Inflammatory Effects in Macrophage Cells. Front. Bioeng. Biotechnol. 2021, 9, 1073. [Google Scholar] [CrossRef]
- Shao, P.; Pei, Y.; Fang, Z.; Sun, P. Effects of partial desulfation on antioxidant and inhibition of DLD cancer cell of Ulva fasciata polysaccharide. Int. J. Biol. Macromol. 2014, 65, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.-H.; Wang, Z.-J.; Shen, M.-Y.; Nie, S.-P.; Gong, B.; Li, H.-S.; Zhao, Q.; Li, W.-J.; Xie, M.-Y. Sulfated modification, characterization and antioxidant activities of polysaccharide from Cyclocarya paliurus. In Food Hydrocolloids; Lai, P., Ed.; Elsevier: Amsterdam, The Netherlands, 2016; Volume 53, pp. 7–15. [Google Scholar] [CrossRef]
- Negreanu-Pirjol, B.-S.; Negreanu-Pirjol, T.; Popoviciu, D.R.; Anton, R.-E.; Prelipcean, A.-M. Marine Bioactive Compounds Derived from Macroalgae as New Potential Players in Drug Delivery Systems: A Review. Pharmaceutics 2022, 14, 781. [Google Scholar] [CrossRef] [PubMed]
- Toillon, R.-A.; Chopin, V.; Jouy, N.; Fauquette, W.; Boilly, B.; Le Bourhis, X. Normal breast epithelial cells induce p53-dependent apoptosis and p53-independent cell cycle arrest of breast cancer cells. Breast Cancer Res. Treat. 2002, 71, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.-L.; Tai, C.-J.; Huang, C.-W.; Chang, F.-R.; Wang, J.-Y. Efficacy of low-molecular-weight fucoidan as a supplemental therapy in metastatic colorectal cancer patients: A double-blind randomized controlled trial. Mar. Drugs 2017, 15, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Je, J.G.; Lee, H.G.; Fernando, K.H.N.; Jeon, Y.J.; Ryu, B. Purification and Structural Characterization of Sulfated Polysaccharides Derived from Brown Algae, Sargassum binderi: Inhibitory Mechanism of iNOS and COX-2 Pathway Interaction. Antioxidants 2021, 10, 822. [Google Scholar] [CrossRef] [PubMed]
- Moussavou, G.; Kwak, D.H.; Obiang-Obonou, B.W.; Maranguy, C.A.; Dinzouna-Boutamba, S.-D.; Lee, D.H.; Pissibanganga, O.G.; Ko, K.; Seo, J.I.; Choo, Y.K. Anticancer effects of different seaweeds on human colon and breast cancers. Mar. Drugs 2014, 12, 4898–4911. [Google Scholar] [CrossRef]
- Zhong, Q.; Wei, B.; Wang, S.; Ke, S.; Chen, J.; Zhang, H.; Wang, H. The antioxidant activity of polysaccharides derived from marine organisms: An overview. Mar. Drugs 2019, 17, 674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadam, S.U.; Tiwari, B.K.; O’Donnell, C.P. Extraction, structure and biofunctional activities of laminarin from brown algae. Int. J. Food Sci. Technol. 2015, 50, 24–31. [Google Scholar] [CrossRef]
- Malyarenko, O.S.; Usoltseva, R.V.; Shevchenko, N.M.; Isakov, V.V.; Zvyagintseva, T.N.; Ermakova, S.P. In vitro anticancer activity of the laminarans from Far Eastern brown seaweeds and their sulfated derivatives. J. Appl. Phycol. 2017, 29, 543–553. [Google Scholar] [CrossRef]
- Chattopadhyay, P.; Chaudhury, P.; Wahi, A.K. Ca2+ concentrations are key determinants of ischemia-reperfusion-induced apoptosis: Significance for the molecular mechanism of Bcl-2 action. Appl. Biochem. Biotechnol. 2010, 160, 1968–1977. [Google Scholar] [CrossRef] [PubMed]
- Kondratskyi, A.; Kondratska, K.; Skryma, R.; Prevarskaya, N. Ion channels in the regulation of apoptosis. Biochim. Biophys. Acta 2015, 1848, 2532–2546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Qi, J.; Zhang, X.; Zhao, X.; An, P.; Luo, Y.; Luo, J. The Regulatory Roles of Mitochondrial Calcium and the Mitochondrial Calcium Uniporter in Tumor Cells. Int. J. Mol. Sci. 2022, 23, 26667. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, X.; Vikash, V.; Ye, Q.; Wu, D.; Liu, Y.; Dong, W. ROS and ROS-Mediated Cellular Signaling. Oxid. Med. Cell Longev. 2016, 2016, 4350965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, A.; Hayashi, T.; Su, T.-P.; Betenbaugh, M.J. Bcl-2 family in inter-organelle modulation of calcium signaling; roles in bioenergetics and cell survival. J. Bioenerg. Biomembr. 2014, 46, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montezano, A.C.; Burger, D.; Ceravolo, G.S.; Yusuf, H.; Montero, M.; Touyz, R.M. Novel nox homologues in the vasculature: Focusing on Nox4 and Nox5. Clin. Sci. 2011, 120, 131–141. [Google Scholar] [CrossRef] [Green Version]
- Roe, N.D.; He, E.Y.; Wu, Z.; Ren, J. Folic acid reverses nitric oxide synthase uncoupling and prevents cardiac dysfunction in insulin resistance: Role of Ca2+/calmodulin-activated protein kinase II. Free Radic. Biol. Med. 2013, 65, 234–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raimondi, M.; Fontana, F.; Marzagalli, M.; Audano, M.; Beretta, G.; Procacci, P.; Sartori, P.; Mitro, N.; Limonta, P. Ca2+ overload- and ROS-associated mitochondrial dysfunction contributes to δ-tocotrienol-mediated paraptosis in melanoma cells. Apoptosis 2021, 26, 277–292. [Google Scholar] [CrossRef]
- Ji, C.-F.; Ji, Y.-B.; Meng, D.-Y. Sulfated modification and anti-tumor activity of laminaran. Exp. Ther. Med. 2013, 6, 1259–1264. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.-T.; Sun, X.-Y.; Yu, K.; Gui, B.-S.; Gui, Q.; Ouyang, J.-M. Effect of Content of Sulfate Groups in Seaweed Polysaccharides on Antioxidant Activity and Repair Effect of Subcellular Organelles in Injured HK-2 Cells. Oxid. Med. Cell Longev. 2017, 2017, 2542950. [Google Scholar] [CrossRef] [Green Version]
- Farooqi, A.A.; Butt, G.; Razzaq, Z. Algae extracts and methyl jasmonate anti-cancer activities in prostate cancer: Choreographers of ‘the dance macabre’. Cancer Cell Int. 2012, 12, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, H.; Liu, X.; Ma, J.; Zhang, Q.; Li, Z. In vitro antioxidant activity of acetylated derivatives of polysaccharide extracted from Ulva pertusa (Cholorophta). J. Med. Plant Res. 2010, 4, 2445–2451. [Google Scholar] [CrossRef]
- El Baky, H.H.A.; El-Baz, F.K.; El Baroty, G.S. Potential biological properties of sulfated polysaccharides extracted from the macroalgae Ulva lactuca L. Cancer Res. 2009, 2, 1–11. [Google Scholar] [CrossRef]
- Qi, H.; Sun, Y. Antioxidant activity of high sulfate content derivative of ulvan in hyperlipidemic rats. Int. J. Biol. Macromol. 2015, 76, 326–329. [Google Scholar] [CrossRef] [PubMed]
- Sanmartín, C.; Plano, D.; Sharma, A.K.; Palop, J.A. Selenium compounds, apoptosis and other types of cell death: An overview for cancer therapy. Int. J. Mol. Sci. 2012, 13, 9649–9672. [Google Scholar] [CrossRef]
- Guo, Y.; Pan, D.; Li, H.; Sun, Y.; Zeng, X.; Yan, B. Antioxidant and immunomodulatory activity of selenium exopolysaccharide produced by Lactococcus lactis subsp. lactis. Food Chem. 2013, 138, 84–89. [Google Scholar] [CrossRef]
- Cheng, L.; Wang, Y.; He, X.; Wei, X. Preparation, Structural Characterization and Bioactivities of Se-Containing Polysaccharide: A Review. Int. J. Biol. Macromoles 2018, 120, 82–92. [Google Scholar] [CrossRef]
- Kaleta, B.; Górski, A.; Zagożdżon, R.; Cieślak, M.; Kaźmierczak-Barańska, J.; Nawrot, B.; Klimaszewska, M.; Malinowska, E.; Górska, S.; Turło, J. Selenium-Containing Polysaccharides from Lentinula Edodes—Biological Activity. Carbohydr. Polym. 2019, 223, 115078. [Google Scholar] [CrossRef]
- Malinowska, E.; Klimaszewska, M.; Strączek, T.; Schneider, K.; Kapusta, C.; Podsadni, P.; Łapienis, G.; Dawidowski, M.; Kleps, J.; Górska, S.; et al. Selenized Polysaccharides—Biosynthesis and Structural Analysis. Carbohydrate Polymers 2018, 198, 407–417. [Google Scholar] [CrossRef]
- Górska, S.; Maksymiuk, A.; Turło, J. Selenium-Containing Polysaccharides—Structural Diversity, Biosynthesis, Chemical Modifications and Biological Activity. Appl. Sci. 2021, 11, 3717. [Google Scholar] [CrossRef]
- He, N.; Shi, X.; Zhao, Y.; Tian, L.; Wang, D.; Yang, X. Inhibitory effects and molecular mechanisms of selenium-containing tea polysaccharides on human breast cancer MCF-7 cells. J. Agric. Food Chem. 2013, 61, 579–588. [Google Scholar] [CrossRef]
- Letavayová, L.; Vlckova, V.; Brozmanova, J. Selenium: From cancer prevention to DNA damage. Toxicology 2006, 227, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Nilsonne, G.; Sun, X.; Nyström, C.; Rundlöf, A.; Fernandes, A.P.; Björnstedt, M.; Dobra, K. Selenite induces apoptosis in sarcomatoid malignant mesothelioma cells through oxidative stress. Free Radic. Biol. Med. 2006, 41, 874–885. [Google Scholar] [CrossRef] [PubMed]
- Radomska, D.; Czarnomysy, R.; Radomski, D.; Bielawski, K. Selenium Compounds as Novel Potential Anticancer Agents. Int. J. Mol. Sci. 2021, 22, 31009. [Google Scholar] [CrossRef]
- Huang, M.; Lu, J.-J.; Huang, M.-Q.; Bao, J.-L.; Chen, X.-P.; Wang, Y.-T. Terpenoids: Natural products for cancer therapy. Expert Opin. Investig. Drugs 2012, 21, 1801–1818. [Google Scholar] [CrossRef]
- Rocha, D.H.A.; Seca, A.M.L.; Pinto, D.C.G.A. Seaweed Secondary Metabolites In Vitro and In Vivo Anticancer Activity. Mar. Drugs 2018, 16, 410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, B.-G.; Gloer, J.B.; Ji, N.-Y.; Zhao, J.-C. Halogenated Organic Molecules of Rhodomelaceae Origin: Chemistry and Biology. Chem. Rev. 2013, 113, 3632–3685. [Google Scholar] [CrossRef] [PubMed]
- Qi, R.-Y.; Guo, C.; Peng, X.-N.; Tang, J.-J. Sesquiterpenoids from Inula britannica and Their Potential Effects against Triple-Negative Breast Cancer Cells. Molecules 2022, 27, 5230. [Google Scholar] [CrossRef] [PubMed]














Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agena, R.; de Jesús Cortés-Sánchez, A.; Hernández-Sánchez, H.; Jaramillo-Flores, M.E. Pro-Apoptotic Activity of Bioactive Compounds from Seaweeds: Promising Sources for Developing Novel Anticancer Drugs. Mar. Drugs 2023, 21, 182. https://doi.org/10.3390/md21030182
Agena R, de Jesús Cortés-Sánchez A, Hernández-Sánchez H, Jaramillo-Flores ME. Pro-Apoptotic Activity of Bioactive Compounds from Seaweeds: Promising Sources for Developing Novel Anticancer Drugs. Marine Drugs. 2023; 21(3):182. https://doi.org/10.3390/md21030182
Chicago/Turabian StyleAgena, Rosette, Alejandro de Jesús Cortés-Sánchez, Humberto Hernández-Sánchez, and María Eugenia Jaramillo-Flores. 2023. "Pro-Apoptotic Activity of Bioactive Compounds from Seaweeds: Promising Sources for Developing Novel Anticancer Drugs" Marine Drugs 21, no. 3: 182. https://doi.org/10.3390/md21030182
APA StyleAgena, R., de Jesús Cortés-Sánchez, A., Hernández-Sánchez, H., & Jaramillo-Flores, M. E. (2023). Pro-Apoptotic Activity of Bioactive Compounds from Seaweeds: Promising Sources for Developing Novel Anticancer Drugs. Marine Drugs, 21(3), 182. https://doi.org/10.3390/md21030182