
Self-Assembly Nanostructure of Myristoylated ω-Conotoxin MVIIA Increases the Duration of Efficacy and Reduces Side Effects
 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Myristoylated MVIIA Self-Assembled into Supramolecular Nanostructures
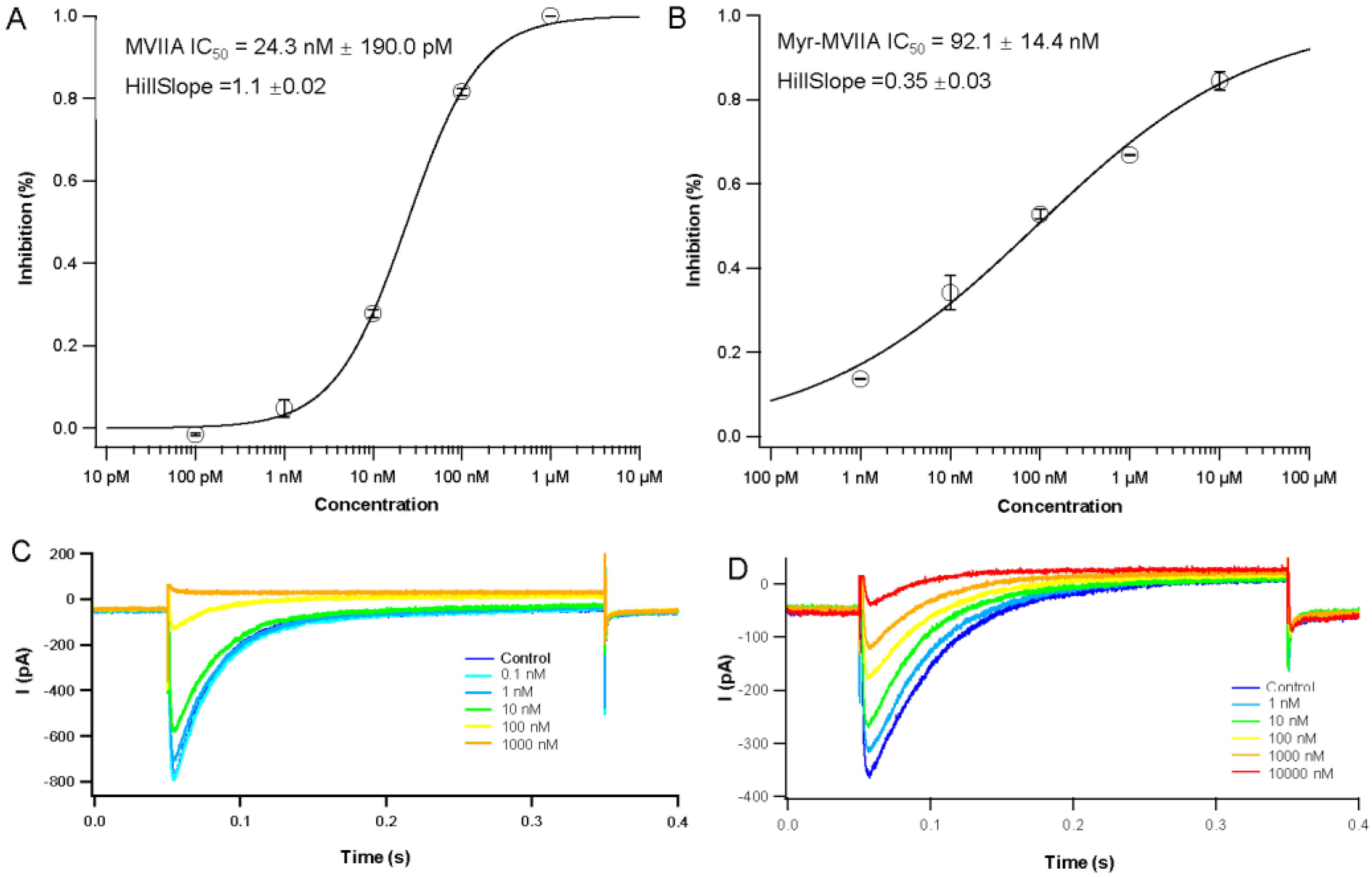
2.2. Inhibition of Cav2.2 Channel Currents Induced by MVIIA and Myr-MVIIA
2.3. Analgesic Effects of Peptides on Acetic-Acid-Induced Visceral Pain in Mice
2.4. Coordinated Locomotion Effects of Peptides
2.5. Tremor Symptoms in Mice Induced by Peptides
2.6. Serum Stability of Peptides
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. MD Simulation
4.3. Peptide Synthesis and Peptide Sample Preparation
4.4. TEM Measurements
4.5. Dynamic Light Scattering
4.6. ANS Binding Assay
4.7. Determination of CMC
4.8. CD Spectroscopy
4.9. Electrophysiology
4.10. Animals
4.11. ICV Injection Procedures
4.12. Acetic-Acid-Induced Writhing Test
4.13. Coordinated Locomotion Test
4.14. Tremor Test
4.15. Peptide Stability in Human Sera
4.16. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Treede, R.-D.; Rief, W.; Barke, A.; Aziz, Q.; Bennett, M.I.; Benoliel, R.; Cohen, M.; Evers, S.; Finnerup, N.B.; First, M.B.; et al. Chronic pain as a symptom or a disease: The IASP Classification of Chronic Pain for the International Classification of Diseases (ICD-11). Pain 2019, 160, 19–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cohen, S.P.; Vase, L.; Hooten, W.M. Chronic pain: An update on burden, best practices, and new advances. Lancet 2021, 397, 2082–2097. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Sun, J.; Liu, H.; Sun, J.; Yu, Y.; Su, Y.; Cui, Y.; Zhao, M.; Zhang, J. Scorpion Toxins Targeting Voltage-gated Sodium Channels Associated with Pain. Curr. Pharm. Biotechnol. 2018, 19, 848–855. [Google Scholar] [CrossRef] [PubMed]
- Vink, S.; Alewood, P.F. Targeting voltage-gated calcium channels: Developments in peptide and small-molecule inhibitors for the treatment of neuropathic pain. Br. J. Pharmacol. 2012, 167, 970–989. [Google Scholar] [CrossRef] [Green Version]
- De Logu, F.; Geppetti, P. Ion Channel Pharmacology for Pain Modulation. Handb. Exp. Pharmacol. 2019, 260, 161–186. [Google Scholar] [CrossRef]
- Hannon, H.E.; Atchison, W.D. Omega-Conotoxins as Experimental Tools and Therapeutics in Pain Management. Mar. Drugs 2013, 11, 680–699. [Google Scholar] [CrossRef] [Green Version]
- Sekiguchi, F.; Tsubota, M.; Kawabata, A. Involvement of Voltage-Gated Calcium Channels in Inflammation and Inflammatory Pain. Biol. Pharm. Bull. 2018, 41, 1127–1134. [Google Scholar] [CrossRef] [Green Version]
- Lee, S. Pharmacological Inhibition of Voltage-gated Ca(2+) Channels for Chronic Pain Relief. Curr. Neuropharmacol. 2013, 11, 606–620. [Google Scholar]
- Snutch, T.P. Targeting chronic and neuropathic pain: The N-type calcium channel comes of age. Neurorx 2005, 2, 662–670. [Google Scholar] [CrossRef]
- Schmidtko, A.; Lötsch, J.; Freynhagen, R.; Geisslinger, G. Ziconotide for treatment of severe chronic pain. Lancet 2010, 375, 1569–1577. [Google Scholar] [CrossRef]
- Garber, K. Peptide leads new class of chronic pain drugs. Nat. Biotechnol. 2005, 23, 399. [Google Scholar] [CrossRef] [PubMed]
- Newcomb, R.; Abbruscato, T.J.; Singh, T.; Nadasdi, L.; Davis, T.P.; Miljanich, G. Bioavailability of Ziconotide in brain: Influx from blood, stability, and diffusion. Peptides 2000, 21, 491–501. [Google Scholar] [CrossRef]
- Scott, D.A.; Wright, C.E.; Angus, J.A. Actions of intrathecal ω-conotoxins CVID, GVIA, MVIIA, and morphine in acute and neuropathic pain in the rat. Eur. J. Pharmacol. 2002, 451, 279–286. [Google Scholar] [CrossRef]
- Penn, R.D.; Paice, J.A. Adverse effects associated with the intrathecal administration of ziconotide. Pain 2000, 85, 291–296. [Google Scholar] [CrossRef]
- Smith, H.S.; Deer, T.R. Safety and efficacy of intrathecal ziconotide in the management of severe chronic pain. Ther. Clin. Risk Manag. 2009, 5, 521–534. [Google Scholar] [CrossRef] [Green Version]
- Maletínská, L.; Nagelová, V.; Tichá, A.; Zemenová, J.; Pirník, Z.; Holubová, M.; Špolcová, A.; Mikulášková, B.; Blechová, M.; Sýkora, D.; et al. Novel lipidized analogs of prolactin-releasing peptide have prolonged half-lives and exert anti-obesity effects after peripheral administration. Int. J. Obes. (Lond.) 2015, 39, 986–993. [Google Scholar] [CrossRef]
- Dishon, S.; Schumacher-Klinger, A.; Gilon, C.; Hoffman, A.; Nussbaum, G. Myristoylation Confers Oral Bioavailability and Improves the Bioactivity of c(MyD 4-4), a Cyclic Peptide Inhibitor of MyD88. Mol. Pharm. 2019, 16, 1516–1522. [Google Scholar] [CrossRef] [PubMed]
- Mráziková, L.; Neprašová, B.; Mengr, A.; Popelová, A.; Strnadová, V.; Holá, L.; Železná, B.; Kuneš, J.; Maletínská, L. Lipidized Prolactin-Releasing Peptide as a New Potential Tool to Treat Obesity and Type 2 Diabetes Mellitus: Preclinical Studies in Rodent Models. Front. Pharmacol. 2021, 12, 779962. [Google Scholar] [CrossRef]
- Weroński, K.J.; Diez-Pérez, I.; Busquets, M.A.; López-Iglesias, C.; Girona, V.; Prat, J. Interaction of lipidated GBV-C/HGV NS3 (513-522) and (505-514) peptides with phospholipids monolayer. An AFM study. Colloids Surf. B Biointerfaces 2010, 75, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Italiya, K.S.; Basak, M.; Mazumdar, S.; Sahel, D.K.; Shrivastava, R.; Chitkara, D.; Mittal, A. Scalable Self-Assembling Micellar System for Enhanced Oral Bioavailability and Efficacy of Lisofylline for Treatment of Type-I Diabetes. Mol. Pharm. 2019, 16, 4954–4967. [Google Scholar] [CrossRef]
- La Manna, S.; Di Natale, C.; Onesto, V.; Marasco, D. Self-Assembling Peptides: From Design to Biomedical Applications. Int. J. Mol. Sci. 2021, 22, 12662. [Google Scholar] [CrossRef] [PubMed]
- Kurtzhals, P. Pharmacology of insulin detemir. Endocrinol. Metab. Clin. 2007, 36 (Suppl. 1), 14–20. [Google Scholar] [CrossRef] [PubMed]
- Hordern, S.V. Insulin detemir: A review. Drugs Today 2006, 42, 505–517. [Google Scholar] [CrossRef]
- Home, P.; Kurtzhals, P. Insulin detemir: From concept to clinical experience. Expert Opin. Pharmacother. 2006, 7, 325–343. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Zhang, S.; Yang, Z.; Wang, J.; Shan, A. Application of supramolecular peptide self-assembly in biomedicine. Sheng Wu Gong Cheng Xue Bao Chin. J. Biotechnol. 2021, 37, 2240–2255. [Google Scholar] [CrossRef]
- Wermeling, D.P. Ziconotide, an Intrathecally Administered N-Type Calcium Channel Antagonist for the Treatment of Chronic Pain. Pharmacother. J. Hum. Pharmacol. Drug Ther. 2005, 25, 1084–1094. [Google Scholar] [CrossRef]
- Kristipati, R.; Nádasdi, L.; Tarczy-Hornoch, K.; Lau, K.; Miljanich, G.P.; Ramachandran, J.; Bell, J.R. Characterization of the Binding of Omega-Conopeptides to Different Classes of Non-L-Type Neuronal Calcium Channels. Mol. Cell. Neurosci. 1994, 5, 219–228. [Google Scholar] [CrossRef]
- Yu, S.; Li, Y.; Chen, J.; Zhang, Y.; Tao, X.; Dai, Q.; Wang, Y.; Li, S.; Dong, M. TAT-Modified ω-Conotoxin MVIIA for Crossing the Blood-Brain Barrier. Mar. Drugs 2019, 17, 286. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Yan, Z.; Liu, Z.; Wang, S.; Wu, Q.; Yu, S.; Ding, J.; Dai, Q. Molecular basis of toxicity of N-type calcium channel inhibitor MVIIA. Neuropharmacology 2016, 101, 137–145. [Google Scholar] [CrossRef]
- Kunisawa, N.; Iha, H.A.; Shimizu, S.; Tokudome, K.; Mukai, T.; Kinboshi, M.; Serikawa, T.; Ohno, Y. Nicotine evokes kinetic tremor by activating the inferior olive via α7 nicotinic acetylcholine receptors. Behav. Brain Res. 2016, 314, 173–180. [Google Scholar] [CrossRef]
- Green, B.D.; Gault, V.A.; Mooney, M.H.; Irwin, N.; Harriott, P.; Greer, B.; Bailey, C.J.; O’Harte, F.P.; Flatt, P.R. Degradation, receptor binding, insulin secreting and antihyperglycaemic actions of palmitate-derivatised native and Ala8-substituted GLP-1 analogues. Biol. Chem. 2004, 385, 169–177. [Google Scholar] [CrossRef]
- Heck, T.; Limbach, M.; Geueke, B.; Zacharias, M.; Gardiner, J.; Kohler, H.P.; Seebach, D. Enzymatic degradation of beta- and mixed alpha,beta-oligopeptides. Chem. Biodivers. 2006, 3, 1325–1348. [Google Scholar] [CrossRef]
- Zhou, X.-R.; Cao, Y.; Zhang, Q.; Tian, X.-B.; Dong, H.; Chen, L.; Luo, S.-Z. Self-assembly nanostructure controlled sustained release, activity and stability of peptide drugs. Int. J. Pharm. 2017, 528, 723–731. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Sun, F.; Zhou, X.-R.; Luo, S.-Z.; Chen, L. Role of peptide self-assembly in antimicrobial peptides. J. Pept. Sci. 2015, 21, 530–539. [Google Scholar] [CrossRef] [PubMed]
- Van Der Spoel, D.; Lindahl, E.; Hess, B.; Groenhof, G.; Mark, A.E.; Berendsen, H.J. GROMACS: Fast, flexible, and free. J. Comput. Chem. 2005, 26, 1701–1718. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.; Cheng, X.; Islam, M.S.; Huang, L.; Rui, H.; Zhu, A.; Lee, H.S.; Qi, Y.; Han, W.; Vanommeslaeghe, K.; et al. CHARMM-GUI PDB manipulator for advanced modeling and simulations of proteins containing nonstandard residues. Adv. Protein Chem. Struct. Biol. 2014, 96, 235–265. [Google Scholar]
- Liu, H.; Zhao, X.; Liang, S.; Fan, L.; Li, Z.; Zhang, Y.; Ni, J. Amphiphilic Endomorphin-1 Derivative Functions as Self-assembling Nanomedicine for Effective Brain Delivery. Chem. Pharm. Bull. 2019, 67, 977–984. [Google Scholar] [CrossRef] [Green Version]
- Lei, R.; Hou, J.; Chen, Q.; Yuan, W.; Cheng, B.; Sun, Y.; Jin, Y.; Ge, L.; Ben-Sasson, S.A.; Chen, J.; et al. Self-Assembling Myristoylated Human α-Defensin 5 as a Next-Generation Nanobiotics Potentiates Therapeutic Efficacy in Bacterial Infection. ACS Nano 2018, 12, 5284–5296. [Google Scholar] [CrossRef]
- François-Moutal, L.; Wang, Y.; Moutal, A.; Cottier, K.E.; Melemedjian, O.K.; Yang, X.; Wang, Y.; Ju, W.; Largent-Milnes, T.M.; Khanna, M.; et al. A membrane-delimited N-myristoylated CRMP2 peptide aptamer inhibits CaV2.2 trafficking and reverses inflammatory and postoperative pain behaviors. Pain 2015, 156, 1247–1264. [Google Scholar] [CrossRef] [Green Version]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, X.; Wang, Y.; Zhang, S.; Zhang, R.; Chen, D.; Chen, L.; Zhang, Y.; Luo, S.-Z.; Xu, J.; Pei, C. Self-Assembly Nanostructure of Myristoylated ω-Conotoxin MVIIA Increases the Duration of Efficacy and Reduces Side Effects. Mar. Drugs 2023, 21, 229. https://doi.org/10.3390/md21040229
Ding X, Wang Y, Zhang S, Zhang R, Chen D, Chen L, Zhang Y, Luo S-Z, Xu J, Pei C. Self-Assembly Nanostructure of Myristoylated ω-Conotoxin MVIIA Increases the Duration of Efficacy and Reduces Side Effects. Marine Drugs. 2023; 21(4):229. https://doi.org/10.3390/md21040229
Chicago/Turabian StyleDing, Xiufang, Yue Wang, Sida Zhang, Ruihua Zhang, Dong Chen, Long Chen, Yu Zhang, Shi-Zhong Luo, Jianfu Xu, and Chengxin Pei. 2023. "Self-Assembly Nanostructure of Myristoylated ω-Conotoxin MVIIA Increases the Duration of Efficacy and Reduces Side Effects" Marine Drugs 21, no. 4: 229. https://doi.org/10.3390/md21040229
APA StyleDing, X., Wang, Y., Zhang, S., Zhang, R., Chen, D., Chen, L., Zhang, Y., Luo, S. -Z., Xu, J., & Pei, C. (2023). Self-Assembly Nanostructure of Myristoylated ω-Conotoxin MVIIA Increases the Duration of Efficacy and Reduces Side Effects. Marine Drugs, 21(4), 229. https://doi.org/10.3390/md21040229