


Gambierone and Sodium Channel Specific Bioactivity Are Associated with the Extracellular Metabolite Pool of the Marine Dinoflagellate Coolia palmyrensis
 ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Identification of C. palmyrensis
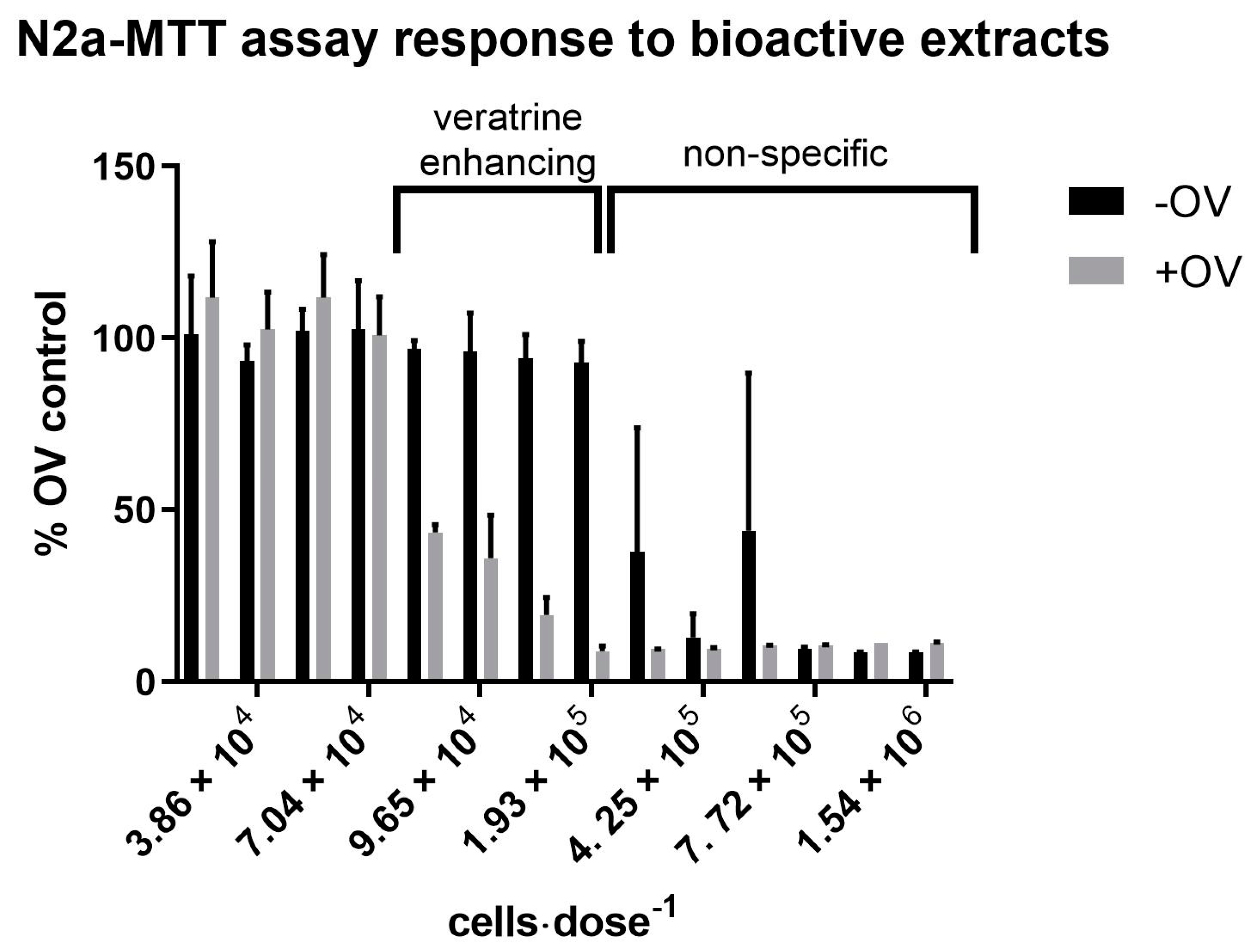
2.2. Bioactivity Associated with C. palmyrensis Media Extracts
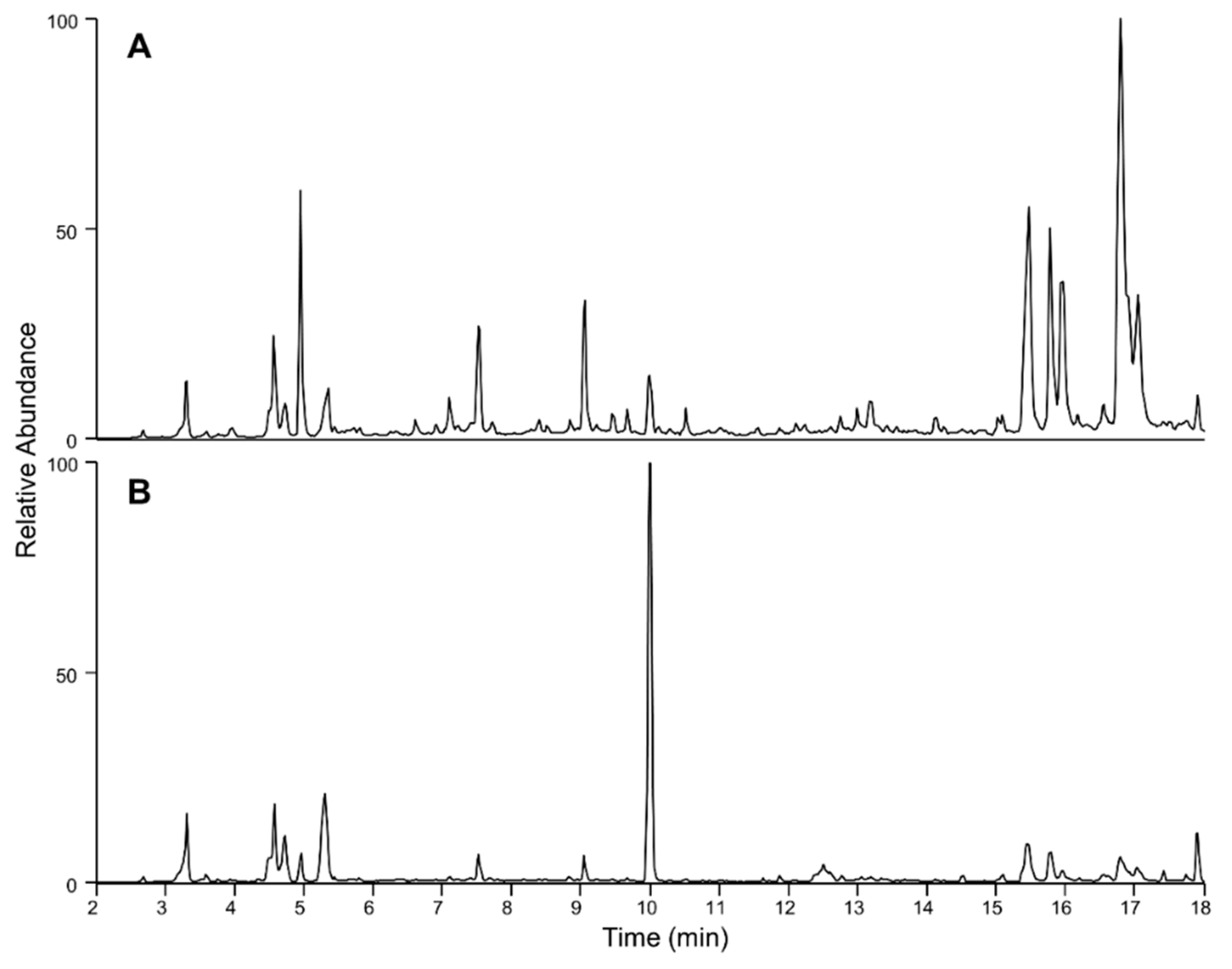
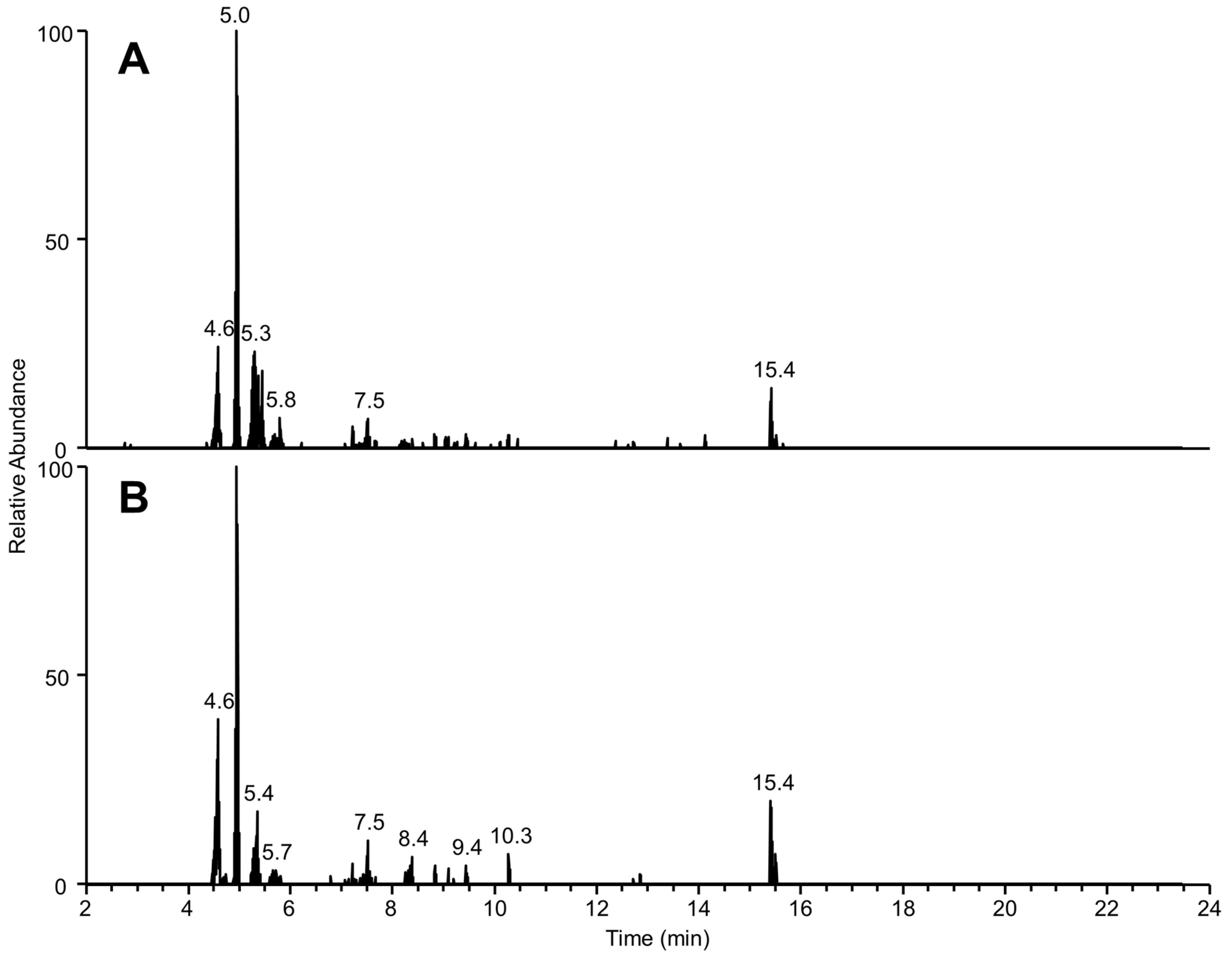
2.3. Targeted LC-MS/MS Analysis of Coolia palmyrensis
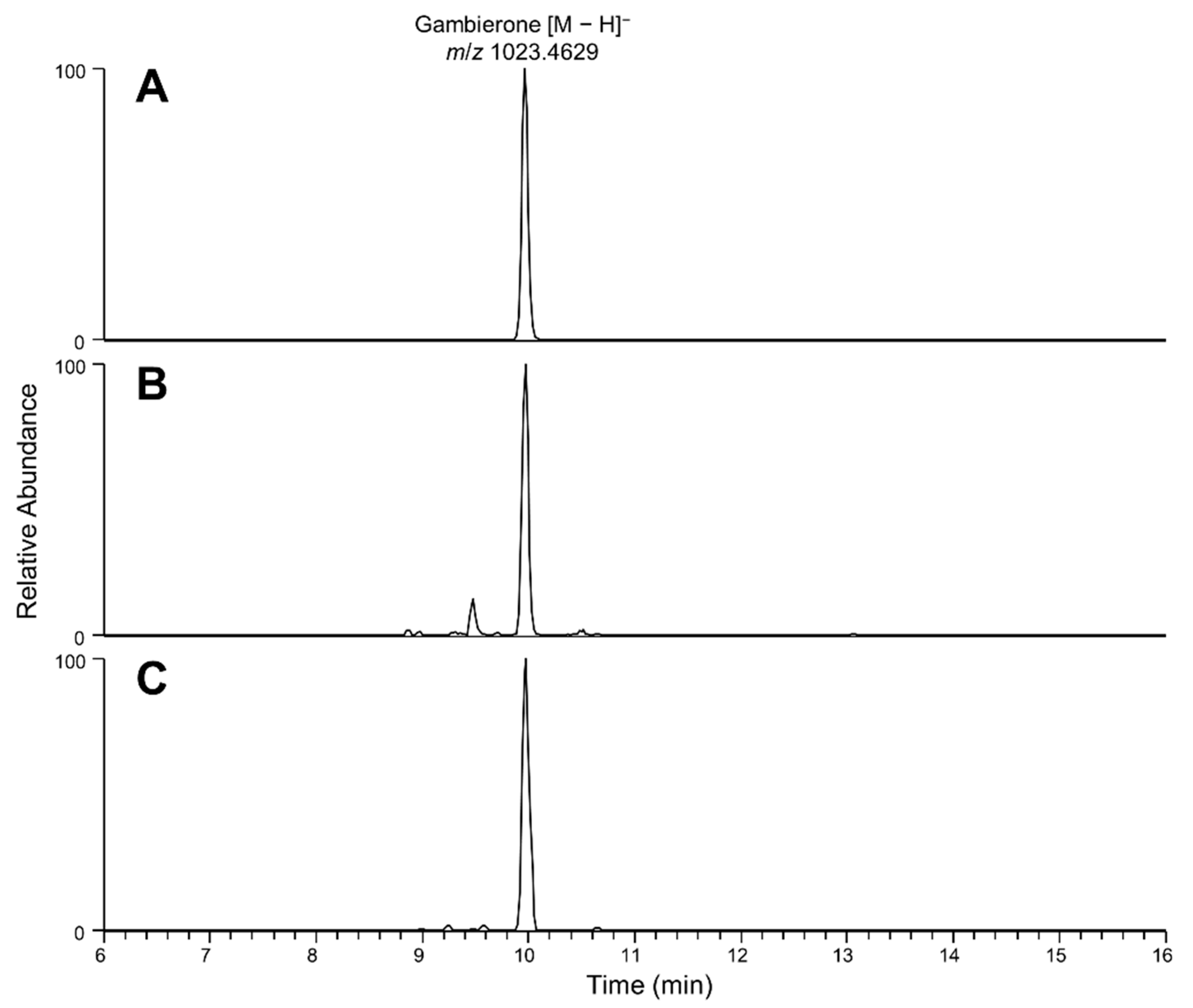
2.4. Liquid Chromatography High Resolution Mass Spectrometry
3. Discussion
3.1. Bioactivity and Gambierones in C. palmyrensis Media Extracts
3.2. Role of Gambierones and C. palmyrensis in CP
3.3. Extracellular Release of Metabolites into C. palymyrensis Media and Potential Ecological Implications
4. Conclusions
5. Materials and Methods
5.1. Materials, Reagents, and Glassware
5.2. Single Cell Isolation and Culture Establishment
5.3. Dinoflagellate Culture Conditions
5.4. Dinoflagellate Species Identification
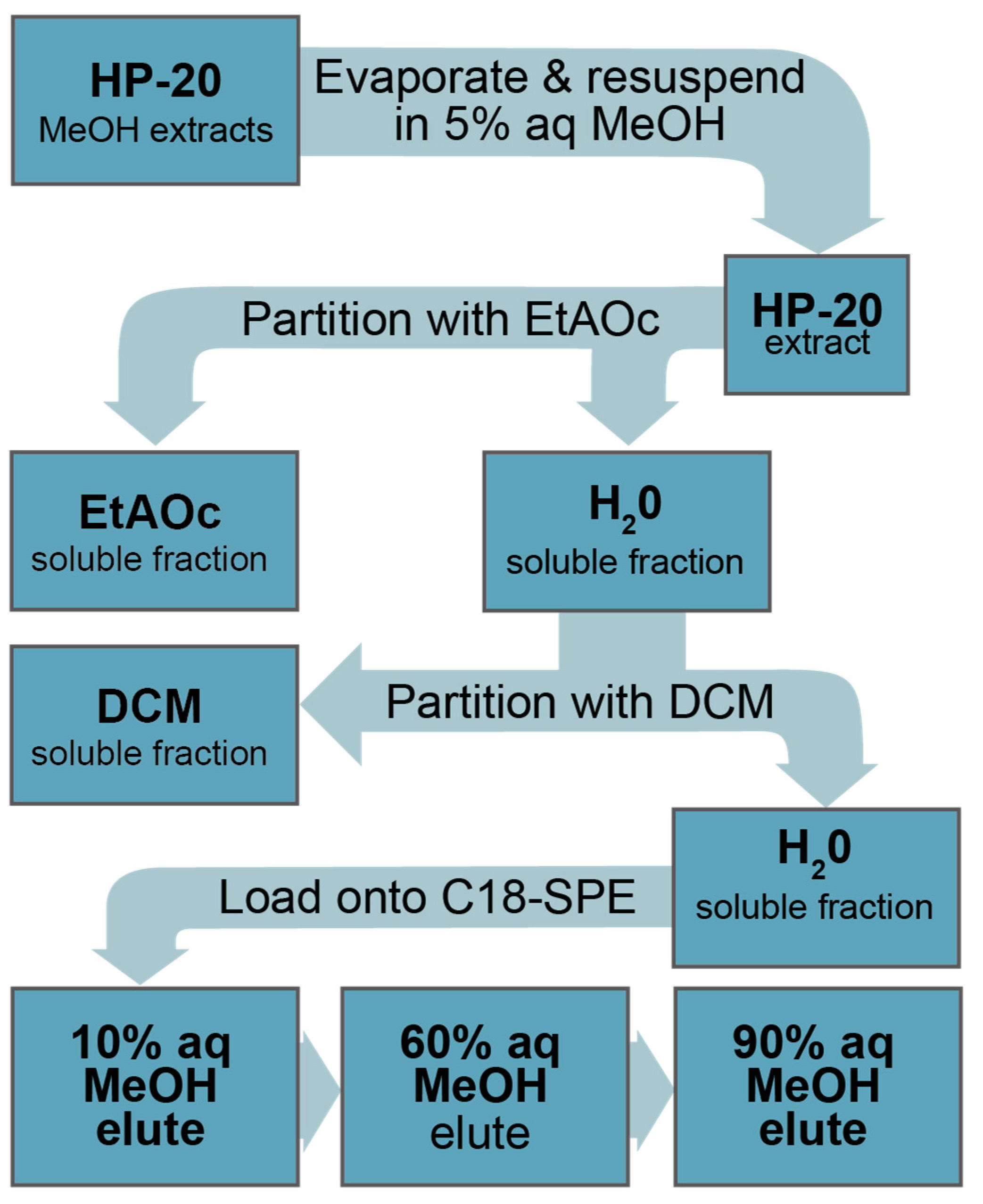
5.5. Isolation, Extraction, and Fractionation of C. palmyrensis from Large Scale Culture
5.6. Algal Cell Counts and Dosing
5.7. N2A-MTT (N2a) Assay
5.8. UHPLC-MS/MS Targeted Phycotoxin Analysis
5.9. LC-HR-MS Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bagnis, R.; Chanteau, S.; Chungue, E.; Hurtel, J.M.; Yasumoto, T.; Inoue, A. Origins of ciguatera fish poisoning: A new dinoflagellate, Gambierdiscus toxicus Adachi and Fukuyo, definitively involved as a causal agent. Toxicon 1980, 18, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, T.; Nakajima, I.; Bagnis, R.; Adachi, R. Finding of a dinoflagellate as a likely culprit of ciguatera. Bull. Jpn. Soc. Sci. Fish. 1977, 43, 1021–1026. [Google Scholar] [CrossRef] [Green Version]
- Litaker, R.W.; Holland, W.C.; Hardison, D.R.; Pisapia, F.; Hess, P.; Kibler, S.R.; Tester, P.A. Ciguatoxicity of Gambierdiscus and Fukuyoa species from the Caribbean and Gulf of Mexico. PLoS ONE 2017, 12, e0185776. [Google Scholar] [CrossRef] [Green Version]
- Chinain, M.; Germain, M.; Deparis, X.; Pauillac, S.; Legrand, A.-M. Seasonal abundance and toxicity of the dinoflagellate Gambierdiscus spp. (Dinophyceae), the causative agent of ciguatera in Tahiti, French Polynesia. Mar. Biol. 1999, 135, 259–267. [Google Scholar] [CrossRef]
- Liefer, J.; Richlen, M.; Smith, T.; DeBose, J.; Xu, Y.; Anderson, D.; Robertson, A. Asynchrony of Gambierdiscus spp. Abundance and Toxicity in the U.S. Virgin Islands: Implications for Monitoring and Management of Ciguatera. Toxins 2021, 13, 413. [Google Scholar] [CrossRef]
- Bennett, C.T.; Robertson, A. Depuration Kinetics and Growth Dilution of Caribbean Ciguatoxin in the Omnivore Lagodon rhomboides: Implications for Trophic Transfer and Ciguatera Risk. Toxins 2021, 13, 774. [Google Scholar] [CrossRef]
- Parsons, M.; Aligizaki, K.; Bottein, M.-Y.D.; Fraga, S.; Morton, S.L.; Penna, A.; Rhodes, L. Gambierdiscus and Ostreopsis: Reassessment of the state of knowledge of their taxonomy, geography, ecophysiology, and toxicology. Harmful Algae 2012, 14, 107–129. [Google Scholar] [CrossRef]
- Chinain, M.; Gatti, C.M.; Roué, M.; Darius, H.T. Ciguatera poisoning in French Polynesia: Insights into the novel trends of an ancient disease. New Microbes New Infect. 2019, 31, 100565. [Google Scholar] [CrossRef]
- de Haro, L.; Schmitt, C.; Sinno-Tellier, S.; Paret, N.; Boels, D.; Le Roux, G.; Langrand, J.; Delcourt, N.; Labadie, M.; Simon, N. Ciguatera fish poisoning in France: Experience of the French Poison Control Centre Network from 2012 to 2019. Clin. Toxicol. 2020, 59, 252–255. [Google Scholar] [CrossRef]
- Bordenstein, S.R.; Theis, K.R. Host Biology in Light of the Microbiome: Ten Principles of Holobionts and Hologenomes. PLoS Biol. 2015, 13, e1002226. [Google Scholar] [CrossRef] [Green Version]
- Agler, M.; Ruhe, J.; Kroll, S.; Morhenn, C.; Kim, S.-T.; Weigel, D.; Kemen, E.M. Microbial Hub Taxa Link Host and Abiotic Factors to Plant Microbiome Variation. PLoS Biol. 2016, 14, e1002352. [Google Scholar] [CrossRef] [Green Version]
- Rosenberg, E.; Zilber-Rosenberg, I. Microbes Drive Evolution of Animals and Plants: The Hologenome Concept. Mbio 2016, 7, e01395-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van der Loos, L.; Eriksson, B.K.; Salles, J.F. The Macroalgal Holobiont in a Changing Sea. Trends Microbiol. 2019, 27, 635–650. [Google Scholar] [CrossRef]
- Barott, K.L.; Rodriguez-Brito, B.; Janouškovec, J.; Marhaver, K.L.; Smith, J.E.; Keeling, P.; Rohwer, F.L. Microbial diversity associated with four functional groups of benthic reef algae and the reef-building coral Montastraea annularis. Environ. Microbiol. 2011, 13, 1192–1204. [Google Scholar] [CrossRef] [PubMed]
- David, H.; Laza-Martínez, A.; Rodríguez, F.; Fraga, S.; Orive, E. Coolia guanchica sp. nov. (Dinophyceae) a new epibenthic dinoflagellate from the Canary Islands (NE Atlantic Ocean). Eur. J. Phycol. 2019, 55, 76–88. [Google Scholar] [CrossRef]
- Leaw, C.P.; Tan, T.H.; Lim, H.C.; Teng, S.T.; Yong, H.L.; Smith, K.F.; Rhodes, L.; Wolf, M.; Holland, W.C.; Vandersea, M.W.; et al. New scenario for speciation in the benthic dinoflagellate genus Coolia (Dinophyceae). Harmful Algae 2016, 55, 137–149. [Google Scholar] [CrossRef]
- Durán-Riveroll, L.M.; Cembella, A.D.; Okolodkov, Y.B. A Review on the Biodiversity and Biogeography of Toxigenic Benthic Marine Dinoflagellates of the Coasts of Latin America. Front. Mar. Sci. 2019, 6, 148. [Google Scholar] [CrossRef] [Green Version]
- Fraga, S.; Penna, A.; Bianconi, I.; Paz, B.; Zapata, M. Coolia canariensis sp. nov. (dinophyceae), a new nontoxic epiphytic benthic dinoflagellate from the Canary Islands. J. Phycol. 2008, 44, 1060–1070. [Google Scholar] [CrossRef] [PubMed]
- David, H.; Laza-Martínez, A.; Miguel, I.; Orive, E. Broad distribution of Coolia monotis and restricted distribution of Coolia cf. canariensis (Dinophyceae) on the Atlantic coast of the Iberian Peninsula. Phycologia 2014, 53, 342–352. [Google Scholar] [CrossRef]
- Karafas, S.; York, R.; Tomas, C. Morphological and genetic analysis of the Coolia monotis species complex with the introduction of two new species, Coolia santacroce sp. nov. and Coolia palmyrensis sp. nov. (Dinophyceae). Harmful Algae 2015, 46, 18–33. [Google Scholar] [CrossRef]
- Larsson, M.E.; Smith, K.F.; Doblin, M.A. First description of the environmental niche of the epibenthic dinoflagellate species Coolia palmyrensis, C. malayensis, and C. tropicalis (Dinophyceae) from Eastern Australia. J. Phycol. 2019, 55, 565–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakeman, K.C.; Yamaguchi, A.; Roy, M.C.; Jenke-Kodama, H. Morphology, phylogeny and novel chemical compounds from Coolia malayensis (Dinophyceae) from Okinawa, Japan. Harmful Algae 2015, 44, 8–19. [Google Scholar] [CrossRef]
- Holmes, M.J.; Lewis, R.J.; Jones, A.; Hoy, A.W.W. Cooliatoxin, the first toxin from Coolia monotis (dinophyceae). Nat. Toxins 1995, 3, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Tibiriçá, C.E.J.D.A.; Sibat, M.; Fernandes, L.F.; Bilien, G.; Chomérat, N.; Hess, P.; Mafra, L.L., Jr. Diversity and Toxicity of the Genus Coolia Meunier in Brazil, and Detection of 44-methyl Gambierone in Coolia tropicalis. Toxins 2020, 12, 327. [Google Scholar] [CrossRef]
- Murray, J.; Finch, S.; Puddick, J.; Rhodes, L.; Harwood, D.; van Ginkel, R.; Prinsep, M. Acute Toxicity of Gambierone and Quantitative Analysis of Gambierones Produced by Cohabitating Benthic Dinoflagellates. Toxins 2021, 13, 333. [Google Scholar] [CrossRef] [PubMed]
- Murray, J.S.; Nishimura, T.; Finch, S.C.; Rhodes, L.L.; Puddick, J.; Harwood, D.T.; Larsson, M.E.; Doblin, M.A.; Leung, P.; Yan, M.; et al. The role of 44-methylgambierone in ciguatera fish poisoning: Acute toxicity, production by marine microalgae and its potential as a biomarker for Gambierdiscus spp. Harmful Algae 2020, 97, 101853. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, L.L.; Thomas, A.E. Coolia monotis (Dinophyceae): A toxic epiphytic microalgal species found in New Zealand (Note). New Zealand J. Mar. Freshw. Res. 1997, 31, 139–141. [Google Scholar] [CrossRef]
- Leung, P.T.; Yan, M.; Yiu, S.K.; Lam, V.T.; Ip, J.C.; Au, M.W.; Chen, C.-Y.; Wai, T.-C.; Lam, P.K. Molecular phylogeny and toxicity of harmful benthic dinoflagellates Coolia (Ostreopsidaceae, Dinophyceae) in a sub-tropical marine ecosystem: The first record from Hong Kong. Mar. Pollut. Bull. 2017, 124, 878–889. [Google Scholar] [CrossRef] [PubMed]
- Laza-Martinez, A.; Orive, E.; Miguel, I. Morphological and genetic characterization of benthic dinoflagellates of the genera Coolia, Ostreopsis and Prorocentrum from the south-eastern Bay of Biscay. Eur. J. Phycol. 2011, 46, 45–65. [Google Scholar] [CrossRef]
- Li, X.; Yan, M.; Gu, J.; Lam, V.T.; Wai, T.-C.; Baker, D.M.; Thompson, P.D.; Yiu, S.K.; Lam, P.K.; Leung, P.T. The effect of temperature on physiology, toxicity and toxin content of the benthic dinoflagellate Coolia malayensis from a seasonal tropical region. Water Res. 2020, 185, 116264. [Google Scholar] [CrossRef]
- Gu, J.; Yan, M.; Leung, P.T.; Tian, L.; Lam, V.T.; Cheng, S.H.; Lam, P.K. Toxicity effects of hydrophilic algal lysates from Coolia tropicalis on marine medaka larvae (Oryzias melastigma). Aquat. Toxicol. 2021, 234, 105787. [Google Scholar] [CrossRef] [PubMed]
- Mendes, M.C.d.Q.; Nunes, J.M.d.C.; Fraga, S.; Rodríguez, F.; Franco, J.M.; Riobó, P.; Branco, S.; Menezes, M. Morphology, molecular phylogeny and toxinology of Coolia and Prorocentrum strains isolated from the tropical South Western Atlantic Ocean. Bot. Mar. 2019, 62, 125–140. [Google Scholar] [CrossRef]
- Giussani, V.; Sbrana, F.; Asnaghi, V.; Vassalli, M.; Faimali, M.; Casabianca, S.; Penna, A.; Ciminiello, P.; Dell’Aversano, C.; Tartaglione, L.; et al. Active role of the mucilage in the toxicity mechanism of the harmful benthic dinoflagellate Ostreopsis cf. ovata. Harmful Algae 2015, 44, 46–53. [Google Scholar] [CrossRef]
- Faimali, M.; Giussani, V.; Piazza, V.; Garaventa, F.; Corrà, C.; Asnaghi, V.; Privitera, D.; Gallus, L.; Cattaneo-Vietti, R.; Mangialajo, L.; et al. Toxic effects of harmful benthic dinoflagellate Ostreopsis ovata on invertebrate and vertebrate marine organisms. Mar. Environ. Res. 2012, 76, 97–107. [Google Scholar] [CrossRef]
- Blossom, H.E.; Daugbjerg, N.; Hansen, P.J. Toxic mucus traps: A novel mechanism that mediates prey uptake in the mixotrophic dinoflagellate Alexandrium pseudogonyaulax. Harmful Algae 2012, 17, 40–53. [Google Scholar] [CrossRef]
- Caillaud, A.; de la Iglesia, P.; Barber, E.; Eixarch, H.; Mohammad-Noor, N.; Yasumoto, T.; Diogène, J. Monitoring of dissolved ciguatoxin and maitotoxin using solid-phase adsorption toxin tracking devices: Application to Gambierdiscus pacificus in culture. Harmful Algae 2011, 10, 433–446. [Google Scholar] [CrossRef]
- Ott, B.M.; Litaker, R.W.; Holland, W.C.; Delwiche, C.F. Using RDNA sequences to define dinoflagellate species. PLoS ONE 2022, 17, e0264143. [Google Scholar] [CrossRef] [PubMed]
- Morquecho, L.; Gárate-Lizárraga, I.; Gu, H. Morphological and molecular characterization of the genus Coolia (Dinophyceae) from Bahía de La Paz, southwest Gulf of California. Algae 2022, 37, 185–204. [Google Scholar] [CrossRef]
- Manger, R.L.; Leja, L.S.; Lee, S.Y.; Hungerford, J.M.; Hokama, Y.; Dickey, R.W.; Granade, H.R.; Lewis, R.; Yasumoto, T.; Wekell, M.M. Detection of Sodium Channel Toxins: Directed Cytotoxicity Assays of Purified Ciguatoxins, Brevetoxins, Saxitoxins, and Seafood Extracts. J. AOAC Int. 1995, 78, 521–527. [Google Scholar] [CrossRef]
- Mudge, E.M.; Robertson, A.; Leynse, A.K.; McCarron, P.; Miles, C.O. Selective extraction of gambierone and related metabolites in Gambierdiscus silvae using m-aminophenylboronic acid–agarose gel and liquid chromatography–high-resolution mass spectrometric detection. J. Chromatogr. B 2021, 1188, 123014. [Google Scholar] [CrossRef]
- Viallon, J.; Chinain, M.; Darius, H.T. Revisiting the Neuroblastoma Cell-Based Assay (CBA-N2a) for the Improved Detection of Marine Toxins Active on Voltage Gated Sodium Channels (VGSCs). Toxins 2020, 12, 281. [Google Scholar] [CrossRef] [PubMed]
- Hogg, R.; Lewis, R.; Adams, D. Ciguatoxin (CTX-1) modulates single tetrodotoxin-sensitive sodium channels in rat parasympathetic neurones. Neurosci. Lett. 1998, 252, 103–106. [Google Scholar] [CrossRef]
- Manger, R.; Woodle, D.; Berger, A.; Dickey, R.W.; Jester, E.; Yasumoto, T.; Lewis, R.; Hawryluk, T.; Hungerford, J. Flow Cytometric-Membrane Potential Detection of Sodium Channel Active Marine Toxins: Application to Ciguatoxins in Fish Muscle and Feasibility of Automating Saxitoxin Detection. J. AOAC Int. 2014, 97, 299–306. [Google Scholar] [CrossRef]
- Lombet, A.; Bidard, J.-N.; Lazdunski, M. Ciguatoxin and Brevetoxins Share a Common Receptor Site on the Neuronal Volt-age-Dependent Na+ Channel. FEBS Lett. 1987, 219, 355–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez, I.; Genta-Jouve, G.; Alfonso, C.; Calabro, K.; Alonso, E.; Sánchez, J.A.; Alfonso, A.; Thomas, O.P.; Botana, L.M. Gambierone, a Ladder-Shaped Polyether from the Dinoflagellate Gambierdiscus belizeanus. Org. Lett. 2015, 17, 2392–2395. [Google Scholar] [CrossRef] [PubMed]
- Boente-Juncal, A.; Álvarez, M.; Antelo, Á.; Rodríguez, I.; Calabro, K.; Vale, C.; Thomas, O.P.; Botana, L.M. Structure Elucidation and Biological Evaluation of Maitotoxin-3, a Homologue of Gambierone, from Gambierdiscus belizeanus. Toxins 2019, 11, 79. [Google Scholar] [CrossRef] [Green Version]
- Yon, T.; Sibat, M.; Robert, E.; Lhaute, K.; Holland, W.C.; Litaker, R.W.; Bertrand, S.; Hess, P.; Réveillon, D. Sulfo-Gambierones, Two New Analogs of Gambierone Produced by Gambierdiscus excentricus. Mar. Drugs 2021, 19, 657. [Google Scholar] [CrossRef]
- MacKenzie, L.A. In situ passive solid-phase adsorption of micro-algal biotoxins as a monitoring tool. Curr. Opin. Biotechnol. 2010, 21, 326–331. [Google Scholar] [CrossRef]
- Roué, M.; Darius, H.T.; Viallon, J.; Ung, A.; Gatti, C.; Harwood, D.T.; Chinain, M. Application of solid phase adsorption toxin tracking (SPATT) devices for the field detection of Gambierdiscus toxins. Harmful Algae 2018, 71, 40–49. [Google Scholar] [CrossRef]
- Smith, J.; Lie, A.A.; Seubert, E.L.; Crowley, N.; Robertson, G.; Caron, D.A. Co-occurring dissolved algal toxins observed at multiple coastal sites in southern California via solid phase adsorption toxin tracking. Toxicon 2019, 171, 62–65. [Google Scholar] [CrossRef]
- Holmes, M.J.; Lewis, R.J. Purification and characterisation of large and small maitotoxins from cultured Gambierdiscus toxicus. Nat. Toxins 1994, 2, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Vale, C.; Antelo, A.; Martin, V. Pharmacology of ciguatoxins. In Phycotoxins: Chemistry and Biochemistry; Wiley Blackwell: Hoboken, NJ, USA, 2015; pp. 23–48. [Google Scholar]
- Hendriks, I.; Sintes, T.; Bouma, T.; Duarte, C. Experimental assessment and modeling evaluation of the effects of the seagrass Posidonia oceanica on flow and particle trapping. Mar. Ecol. Prog. Ser. 2008, 356, 163–173. [Google Scholar] [CrossRef] [Green Version]
- Grosser, K.; Zedler, L.; Schmitt, M.; Dietzek, B.; Popp, J.; Pohnert, G. Disruption-free imaging by Raman spectroscopy reveals a chemical sphere with antifouling metabolites around macroalgae. Biofouling 2012, 28, 687–696. [Google Scholar] [CrossRef]
- Liu, L.; Chen, J.; He, X.; Hao, S.; Lian, Z.; Wang, B. First determination of extracellular paralytic shellfish poisoning toxins in the culture medium of toxigenic dinoflagellates by HILIC–HRMS. Ecotoxicol. Environ. Saf. 2020, 204, 111042. [Google Scholar] [CrossRef] [PubMed]
- Cassell, R.T.; Chen, W.; Thomas, S.; Liu, L.; Rein, K.S. Brevetoxin, the Dinoflagellate Neurotoxin, Localizes to Thylakoid Membranes and Interacts with the Light-Harvesting Complex II (LHCII) of Photosystem II. Chembiochem 2015, 16, 1060–1067. [Google Scholar] [CrossRef] [PubMed]
- Mudge, E.M.; Miles, C.O.; Ivanova, L.; Uhlig, S.; James, K.S.; Erdner, D.L.; Fæste, C.K.; McCarron, P.; Robertson, A. Algal ciguatoxin identified as source of ciguatera poisoning in the Caribbean. Chemosphere 2023, 138659. [Google Scholar] [CrossRef]
- Keller, M.D.; Selvin, R.C.; Claus, W.; Guillard, R.R.L. Media for the culture of oceanic ultraphytoplankton. J. Phycol. 2007, 23, 633–638. [Google Scholar] [CrossRef]
- Scholin, C.A.; Herzog, M.; Sogin, M.; Anderson, D.M. Identification of group- and strain-specific genetic markers for globally distributed Alexandrium (dinophyceae). ii. sequence analysis of a fragment of the lsu rRNA gene. J. Phycol. 1994, 30, 999–1011. [Google Scholar] [CrossRef]
- Nunn, G.B.; Theisen, B.F.; Christensen, B.; Arctander, P. Simplicity-correlated size growth of the nuclear 28S ribosomal RNA D3 expansion segment in the crustacean order isopoda. J. Mol. Evol. 1996, 42, 211–223. [Google Scholar] [CrossRef]
- O’Neill, A.; Morrell, N.; Turner, A.D.; Maskrey, B.H. Method performance verification for the combined detection and quantitation of the marine neurotoxins cyclic imines and brevetoxin shellfish metabolites in mussels (Mytilus edulis) and oysters (Crassostrea gigas) by UHPLC-MS/MS. J. Chromatogr. B 2021, 1179, 122864. [Google Scholar] [CrossRef]
- Botana, L.M. (Ed.) Seafood and Freshwater Toxins: Pharmacology, Physiology, and Detection; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar]
- Turner, A.D.; Dhanji-Rapkova, M.; Fong, S.Y.T.; Hungerford, J.; McNabb, P.S.; Boundy, M.J.; Harwood, D.T.; Aanrud, S.; Alfonso, C.; Alvarez, M.; et al. Ultrahigh-Performance Hydrophilic Interaction Liquid Chromatography with Tandem Mass Spectrometry Method for the Determination of Paralytic Shellfish Toxins and Tetrodotoxin in Mussels, Oysters, Clams, Cockles, and Scallops: Collaborative Study. J. AOAC Int. 2020, 103, 533–562. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract Fraction | EtOAc | DCM | H2O |
|---|---|---|---|
| Partition | Non-specific cytotoxicity 1 | None | Non-specific cytotoxicity |
| 10% aq MeOH | None | None | None |
| 60% aq MeOH | None | None | None |
| 90% aq MeOH | Non-specific cytotoxicity | None | Veratrine enhancing activity 2 |
| RT (min) | [M + H]+ | [M-H]− | Proposed Molecular Formula (Neutral) | RDBE | Loss of 18.0106 (-H2O) | Loss of 79.9573 (-SO3) | m/z 219.1378 Product Ion | m/z 233.1533 Product Ion |
|---|---|---|---|---|---|---|---|---|
| 4.6 | 1383.6826 *† | 1399.6755 | C65H108O32 | 12 | No | No | Yes | Yes |
| 5.0 | 1275.8795 | 1273.8616 | C65H126O23 | 2 | No | No | Yes | Yes |
| 5.3 | 1111.5933 | n/d | C53H90O24 | 9 | Yes | No | Yes | No |
| 6.9 | 1260.6594 † | 1294.6643 | C62H105O25NS | 11 | Yes | Yes | No | No |
| 7.5 | 899.5010 * | 915.4976 | C46H76O18 | 9 | Yes | No | Yes | Yes |
| 15.4 | 1163.8562 | 1161.8408 | C67H118O15 | 9 | No | No | Yes | Yes |
| Species | Strain | Location | No. Isolates | Gambierone | 44-Methyl Gambierone | Citation |
|---|---|---|---|---|---|---|
| C. malayensis | -- | Australia | 3 | na | + | Murray et al., 2020 [26] |
| LM036 | Brazil | 1 | na | − | Tibiricá et al., 2020 [24] | |
| -- | Cook Islands | 3 | na | − | Murray et al., 2020 [26] | |
| -- | Hong Kong | 1 | na | − | Murray et al., 2020 [26] | |
| CAWD175, 154 | New Zealand | 5 | + | +/− (2/5) | Murray et al., 2020 & 2021 [25,26] | |
| C. tropicalis | -- | Australia | 3 | na | +/− (2/3) | Murray et al., 2020 [26] |
| LM141 | Brazil | 1 | na | + | Tibiricá et al., 2020 [24] | |
| -- | Cook Islands | 5 | na | + | Murray et al., 2020 [26] | |
| -- | Hong Kong | 2 | na | + | Murray et al., 2020 [26] | |
| UTS2 | ? | 1 | − | + | Murray et al., 2020 [26] | |
| UTS3 | ? | 1 | − | + | Murray et al., 2020 [26] | |
| C. palmyrensis | -- | Australia | 2 | na | − | Murray et al., 2020 [26] |
| LM112 | Brazil | 1 | na | − | Tibiricá et al., 2020 [24] | |
| -- | Australia | 2 | na | − | Murray et al., 2020 [26] | |
| DISL57 | USVI | 1 | + | trace | This study | |
| C. canariensis | -- | Hong Kong | 2 | na | − | Murray et al., 2020 [26] |
| C. monotis | -- | Spain | 1 | na | − | Murray et al., 2020 [26] |
| C. santacroce | LM113 | Brazil | 1 | na | − | Tibiricá et al., 2020 [24] |
| Method | Toxin | Positive Ions (m/z) | CE | CV | Negative Ions (m/z) | CE | CV |
|---|---|---|---|---|---|---|---|
| A | OA, DTX2 | - | - | - | 803.5 > 255.1; 803.5 > 113 | 48; 55 | 70 |
| A | DTX1 | - | - | - | 817.5 > 255.1; 817.5 > 113 | 45; 60 | 70 |
| A | YTX | - | - | - | 570.5 > 467.4; 570.5 > 396.2 | 30 | 38 |
| A | homo YTX | - | - | - | 577.5 > 474.2; 577.5 > 403.2 | 30 | 38 |
| A | 45 OH YTX | - | - | - | 578.5 > 467.4; 578.5 > 396.2 | 30 | 38 |
| A | 45 OH homo YTX | - | - | - | 585.5 > 474.2; 585.5 > 403.2 | 30 | 38 |
| A | AZA1 | 842.5 > 654.4; 842.5 > 362.3 | 50 | 42 | - | - | - |
| A | AZA2 | 856.6 > 654.4; 856.6 > 362.3 | 50 | 42 | - | - | - |
| A | AZA3 | 828.5 > 658.4; 828.5 > 362.3 | 50 | 42 | - | - | - |
| A | PTX1/PTX11 | 892.5 > 821.5; 892.5 > 213.1 | 25; 37 | 32 | - | - | - |
| A | PTX2 | 876.6 > 823.5; 876.6 > 213.1 | 25; 38 | 32 | - | - | - |
| A | SPX1 | 692.5 > 164.1; 692.5 > 444.3 | 45; 35 | 42 | - | - | - |
| A | GYM | 508.4 > 136.1; 508.4 > 162.1 | 38 | 36 | - | - | - |
| B | PnTx E | 784.2 > 164.0; 784.2 > 446.2 | 55; 45 | 40 | - | - | - |
| B | PnTx F | 766.4 > 164.0; 766.4 > 488.2 | 55; 45 | 40 | - | - | - |
| B | PnTx G | 694.5 > 164.0; 694.5 > 458.1 | 55; 50 | 40 | - | - | - |
| B | BTX B2 | 1034.5 > 929.0; 1034.5 > 947.0 | 40 | 40 | - | - | - |
| B | BTX B4 | 1272.7 > 929.4; 1272.7 > 326.2 | 35 | 80 | - | - | - |
| B | BTX B5 | 911.5 > 875.5; 911.5 > 839.0 | 20 | 60 | - | - | - |
| B | PbTx 2 | 895.5 > 319.2; 895.5 > 877.5 | 30 | 60 | - | - | - |
| B | PbTx 3 | 897.5 > 725.5; 897.5 > 129.0 | 30 | 60 | - | - | - |
| B | S desoxy BTX B2 | 1018.6 > 248.2; 1018.6 > 204.1 | 40 | 40 | - | - | - |
| B | 13, 19 didesmethyl spirolide C | 678.5 > 164.1; 678.5 > 430.1 | 50; 35 | 40 | - | - | - |
| B | 20-methyl spirolide C | 706.5 > 163.6; 706.5 > 346.2 | 50; 35 | 40 | - | - | - |
| B | 12methyl GYM | 522.7 > 135.0; 522.7 > 120.5 | 40 | 40 | - | - | - |
| B | PbTX 1 | 867.2 > 221.0; 867.2 > 611.0 | 30 | 60 | - | - | - |
| B | PnTX A | 712.5 > 458.3; 712.5 > 164.1 | 40 | 40 | - | - | - |
| B | PnTX D | 782.0 > 164.0 | 55 | 40 | - | - | - |
| B | PnTX H | 708.0 > 164.0 | 55 | 40 | - | - | - |
| C | STX | 300.1 > 204.1, 138.0 | 31; 24 | - | - | - | - |
| C | NEO | 316.1 > 126.1, 298.1, 220.1 | 30; 20 | - | - | - | - |
| C | dcSTX | 257.1 > 126.1, 222.0 | 20; 20 | - | - | - | - |
| C | dcNEO | 273.1 > 126.1, 225.1 | 24; 18 | - | - | - | - |
| C | doSTX | 241.1 > 60.0, 206.1 | 25; 20 | - | - | - | - |
| C | TTX | 320.1 > 302.1, 162.1 | 28; 44 | - | - | - | - |
| C | GTX2 | - | - | - | 394.1 > 351.1; 394.1 > 333.1 | 20; 18 | - |
| C | GTX3 | 396.1 > 298.1 | 18 | - | 394.1 > 333.1 | 22 | - |
| C | GTX1 | - | - | - | 410.1 > 367.1; 410.1 > 349.1 | 15; 20 | - |
| C | GTX4 | 412.1 > 314.1 | 18 | - | 410.1 > 367.1 | 15 | - |
| C | GTX5 | 380.1 > 300.1 | 15 | - | 378.1 > 122.0 | 22 | - |
| C | GTX6 | 396.1 > 316.1 | 12 | - | 394.1 > 122.0 | 24 | - |
| C | dcGTX2 | - | - | - | 351.1 > 164.0; 351.1 > 333.1 | 22; 12 | - |
| C | dcGTX3 | 353.1 > 255.1 | 15 | - | 351.1 > 333.1 | 18 | - |
| C | dcGTX1 | - | - | - | 367.1 > 274.1; 367.1 > 349.1 | 20; 17 | - |
| C | dcGTX4 | 369.1 > 271.1 | 20 | - | 367.1 > 349.1 | 16 | - |
| C | C1 | - | - | - | 474.1 > 122.0 | 38; 30 | - |
| C | C2 | 396.1 > 298.1 | 15 | - | 474.1 > 122.0; 474.1 > 351.1 | 38 | - |
| C | C3 | 412.1 > 332.1 | 12 | - | 490.1 > 410.1 | 16 | - |
| C | C4 | 412.1 > 314.1 | 12 | - | 490.1 > 392.1 | 20 | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leynse, A.K.; Mudge, E.M.; Turner, A.D.; Maskrey, B.H.; Robertson, A. Gambierone and Sodium Channel Specific Bioactivity Are Associated with the Extracellular Metabolite Pool of the Marine Dinoflagellate Coolia palmyrensis. Mar. Drugs 2023, 21, 244. https://doi.org/10.3390/md21040244
Leynse AK, Mudge EM, Turner AD, Maskrey BH, Robertson A. Gambierone and Sodium Channel Specific Bioactivity Are Associated with the Extracellular Metabolite Pool of the Marine Dinoflagellate Coolia palmyrensis. Marine Drugs. 2023; 21(4):244. https://doi.org/10.3390/md21040244
Chicago/Turabian StyleLeynse, Alexander K., Elizabeth M. Mudge, Andrew D. Turner, Benjamin H. Maskrey, and Alison Robertson. 2023. "Gambierone and Sodium Channel Specific Bioactivity Are Associated with the Extracellular Metabolite Pool of the Marine Dinoflagellate Coolia palmyrensis" Marine Drugs 21, no. 4: 244. https://doi.org/10.3390/md21040244
APA StyleLeynse, A. K., Mudge, E. M., Turner, A. D., Maskrey, B. H., & Robertson, A. (2023). Gambierone and Sodium Channel Specific Bioactivity Are Associated with the Extracellular Metabolite Pool of the Marine Dinoflagellate Coolia palmyrensis. Marine Drugs, 21(4), 244. https://doi.org/10.3390/md21040244