
Study of the Phytochemical Composition, Antioxidant Properties, and In Vitro Anti-Diabetic Efficacy of Gracilaria bursa-pastoris Extracts

 , , , ,
, , , ,  ,
,  , , and
, , and
Abstract
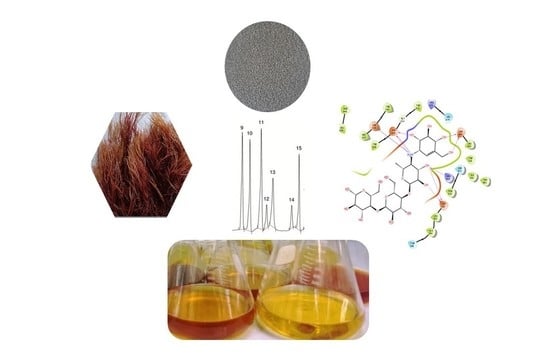
:
1. Introduction
2. Results
2.1. Yields, Phenols and Flavonoids Contents
2.2. Fatty Acid Analysis
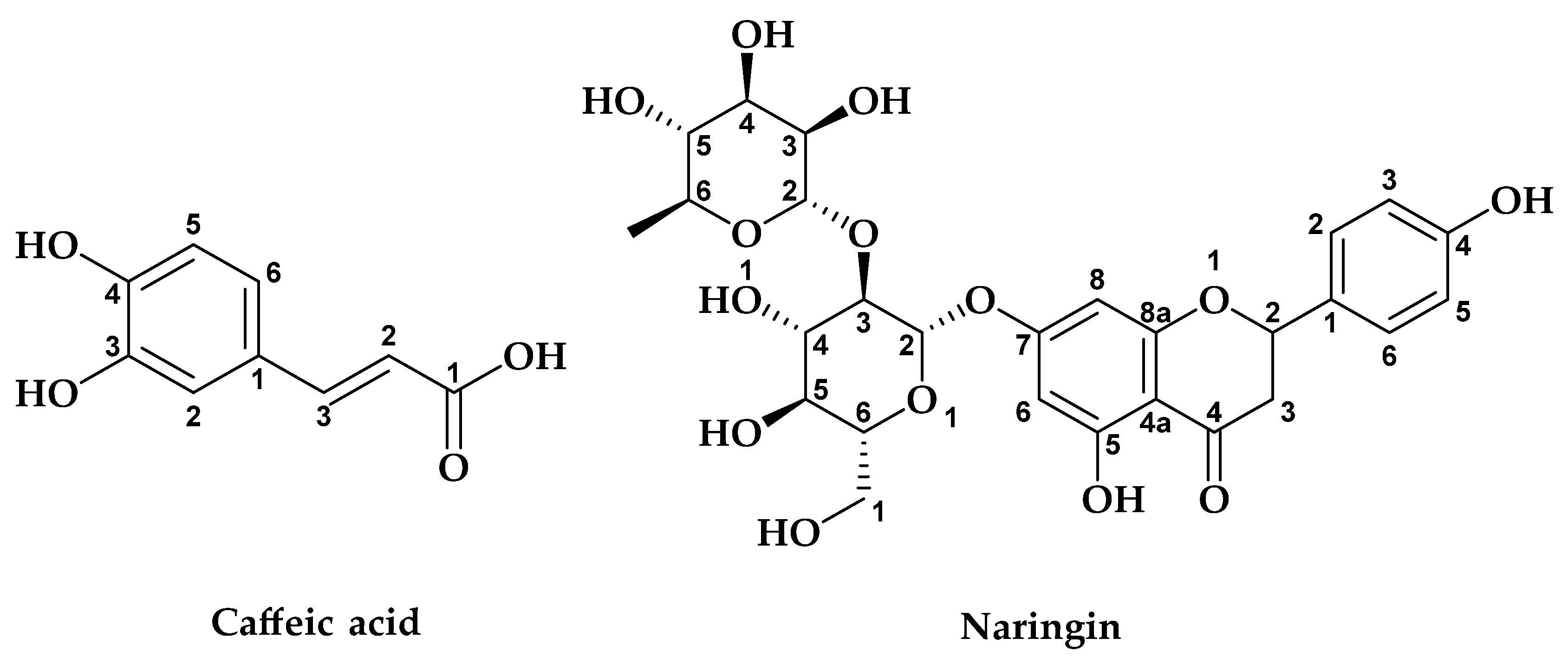
2.3. HPLC Analysis of G. bursa-pastoris Extracts
2.4. Antioxidant Activity
2.5. In Vitro α-Amylase Inhibition
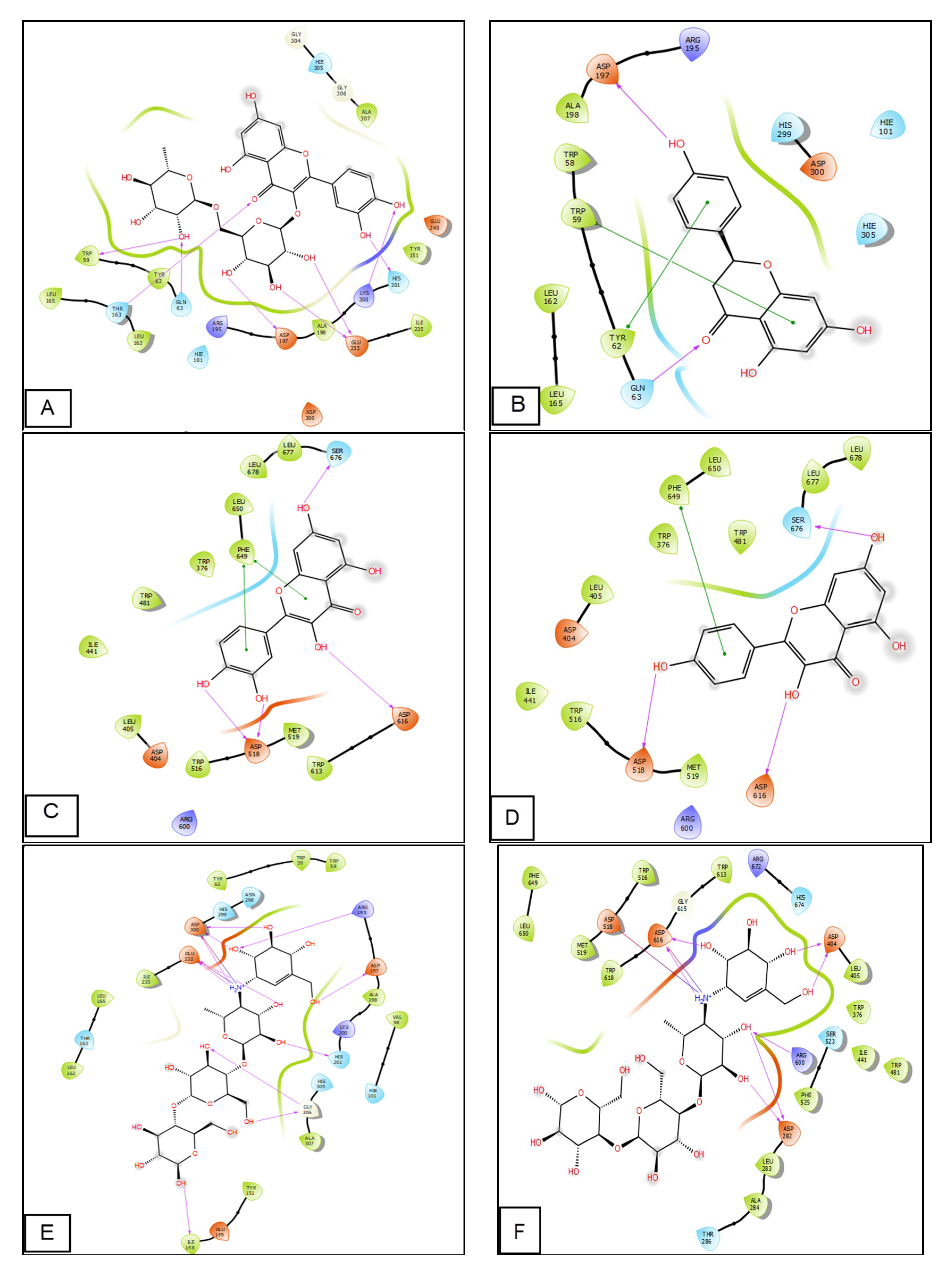
2.6. Molecular Modeling Studies
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
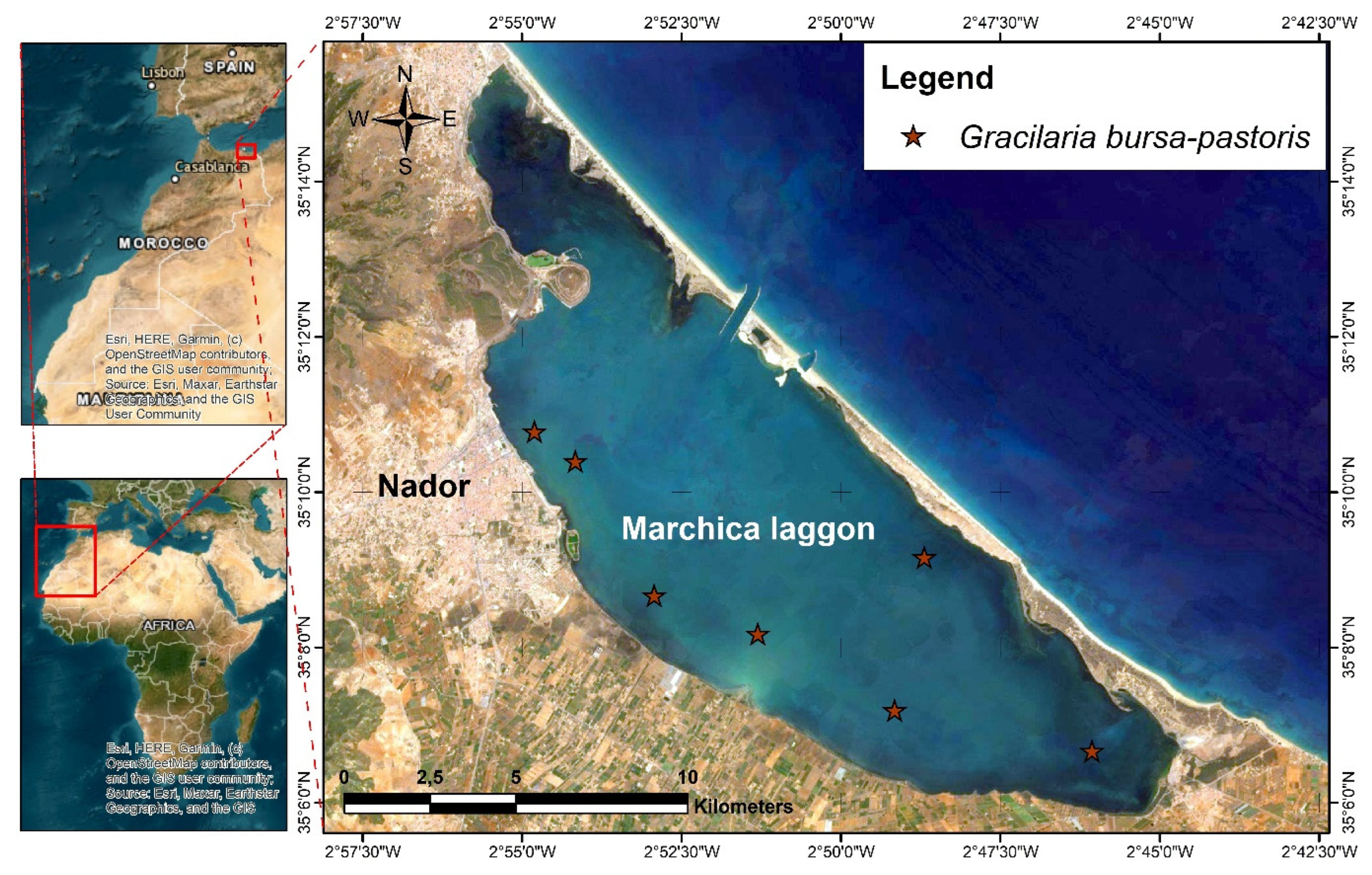
4.2. Plant Material and Extraction
4.2.1. Maceration Extraction
4.2.2. Soxhlet Extraction
4.3. Phytochemicals Compounds
4.3.1. Quantification of Total Phenolic Constituents
4.3.2. Measurement of Total Flavonoids Contents
4.4. Fatty Acid GC-MS Analysis of G. bursa-pastoris Extracts
4.5. HPLC Analyses of G. bursa-pastoris Extracts
4.6. Antioxidant Activity
4.6.1. Scavenging 2, 2-Diphenyl-1-Picrylhydrazyl Radical Test
4.6.2. β-Carotene Bleaching Assay
4.7. In Vitro α-Amylase Inhibition
4.8. In Vitro α-Glucosidase Inhibition Assay
4.9. Molecular Docking Study
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chan, P.T.; Matanjun, P. Chemical composition and physicochemical properties of tropical red seaweed, Gracilaria changii. Food Chem. 2017, 221, 302–310. [Google Scholar] [CrossRef]
- Martirosyan, D.M.; Singh, J. A new definition of functional food by FFC: What makes a new definition unique? Funct. Foods Health Dis. 2015, 5, 209–223. [Google Scholar] [CrossRef]
- Qi, H.; Zhao, T.; Zhang, Q.; Li, Z.; Zhao, Z.; Xing, R. Antioxidant activity of different molecular weight sulfated polysaccharides from Ulva pertusa Kjellm (Chlorophyta). J. Appl. Phycol. 2005, 17, 527–534. [Google Scholar] [CrossRef]
- Van Oosten, M.J.; Pepe, O.; De Pascale, S.; Silletti, S.; Maggio, A. The role of biostimulants and bioeffectors as alleviators of abiotic stress in crop plants. Chem. Biol. Technol. Agric. 2017, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Dawczynski, C.; Schubert, R.; Jahreis, G. Amino acids, fatty acids, and dietary fibre in edible seaweed products. Food Chem. 2007, 103, 891–899. [Google Scholar] [CrossRef]
- Balboa, E.M.; Conde, E.; Moure, A.; Falqué, E.; Domínguez, H. In vitro antioxidant properties of crude extracts and compounds from brown algae. Food Chem. 2013, 138, 1764–1785. [Google Scholar] [CrossRef]
- Ortiz, J.; Romero, N.; Robert, P.; Araya, J.; Lopez-Hernández, J.; Bozzo, C.; Navarrete, E.; Osorio, A.; Rios, A. Dietary fiber, amino acid, fatty acid and tocopherol contents of the edible seaweeds Ulva lactuca and Durvillaea antarctica. Food Chem. 2006, 99, 98–104. [Google Scholar] [CrossRef]
- Fleurence, J. Seaweed proteins: Biochemical, nutritional aspects and potential uses. Trends Food Sci. Technol. 1999, 10, 25–28. [Google Scholar] [CrossRef]
- Lordan, S.; Ross, R.P.; Stanton, C. Marine bioactives as functional food ingredients: Potential to reduce the incidence of chronic diseases. Mar. Drugs 2011, 9, 1056–1100. [Google Scholar] [CrossRef] [Green Version]
- Marinho-Soriano, E.; Fonseca, P.; Carneiro, M.; Moreira, W. Seasonal variation in the chemical composition of two tropical seaweeds. Bioresour. Technol. 2006, 97, 2402–2406. [Google Scholar] [CrossRef]
- Pohnert, G. Chemical defense strategies of marine organisms. In The Chemistry of Pheromones and Other Semiochemicals I; Springer: Berlin/Heidelberg, Germany, 2004; pp. 179–219. [Google Scholar]
- Paul, N.A.; de Nys, R.; Steinberg, P. Chemical defence against bacteria in the red alga Asparagopsis armata: Linking structure with function. Mar. Ecol. Prog. Ser. 2006, 306, 87–101. [Google Scholar] [CrossRef] [Green Version]
- Blunt, J.; Copp, B.; Munro, M.; Northcote, P.; Prinsep, M. Marine natural products. Natural Prod. Rep. 2006, 23, 26–78. [Google Scholar] [CrossRef] [PubMed]
- Etahiri, S.; Bultel-Poncé, V.; Caux, C.; Guyot, M. New bromoditerpenes from the red alga Sphaerococcus coronopifolius. J. Nat. Prod. 2001, 64, 1024–1027. [Google Scholar] [CrossRef] [PubMed]
- Etahiri, S.; Bultel-Ponce, V.; Assobhei, O.; Zaoui, D. Antibacterial activities of marine algae from the Atlantic coast of Morocco. Mar. Life 2003, 13, 3–9. [Google Scholar]
- Smit, A.J. Medicinal and pharmaceutical uses of seaweed natural products: A review. J. Appl. Phycol. 2004, 16, 245–262. [Google Scholar] [CrossRef]
- Bhakuni, D.S.; Rawat, D.S. Bioactive Marine Natural Products; Springer Science & Business Media: Berlin, Germany, 2006. [Google Scholar]
- Etahiri, S.; El Kouria, A.K.; Bultel-Poncé, V.; Guyot, M.; Assobhei, O. Antibacterial bromophenol from the marine red alga Pterosiphonia complanata. Nat. Prod. Commun. 2007, 2, 1934578X0700200708. [Google Scholar] [CrossRef] [Green Version]
- Oumaskour, K.; Boujaber, N.; Etahiri, S.; Assobhei, O. Screening of antibacterial and antifungal activities in green and brown algae from the coast of Sidi Bouzid (El Jadida, Morocco). Afr. J. Biotechnol. 2012, 11, 16831–16837. [Google Scholar]
- Boujaber, N.; Oumaskour, K.; Hassou, N.; Lakhdar, F.; Assobhei, O.; Etahiri, S. Antimicrobial effect of two marine algae Gelidium sesquipedale and Laminaria ochroleuca collected from the coast of El Jadida-Morocco. J. Bio Innov. 2016, 5, 16–23. [Google Scholar]
- Kladi, M.; Xenaki, H.; Vagias, C.; Papazafiri, P.; Roussis, V. New cytotoxic sesquiterpenes from the red algae Laurencia obtusa and Laurencia microcladia. Tetrahedron 2006, 62, 182–189. [Google Scholar] [CrossRef]
- Machado, F.L.d.S.; Kaiser, C.R.; Costa, S.S.; Gestinari, L.M.; Soares, A.R. Atividade biológica de metabólitos secundários de algas marinhas do gênero Laurencia. Rev. Bras. Farmacogn. 2010, 20, 441–452. [Google Scholar] [CrossRef]
- Chan, P.T.; Matanjun, P.; Yasir, S.M.; Tan, T.S. Antioxidant and hypolipidaemic properties of red seaweed, Gracilaria changii. J. Appl. Phycol. 2014, 26, 987–997. [Google Scholar] [CrossRef]
- Garon-Lardière, S. Etude Structurale Des Polysaccharides Pariétaux de L’Algue Rouge Asparagopsis armata (Bonnemaisoniales). Ph.D. Thesis, Université de Bretagne Occidentale, Brest, France, 2004. [Google Scholar]
- Collen, J.; Cornish, M.L.; Craigie, J.; Ficko-Blean, E.; Hervé, C.; Krueger-Hadfield, S.A.; Leblanc, C.; Michel, G.; Potin, P.; Tonon, T. Chondrus crispus–A present and historical model organism for red seaweeds. In Advances in Botanical Research; Elsevier: Amsterdam, The Netherlands, 2014; Volume 71, pp. 53–89. [Google Scholar]
- Rebello, J.; Ohno, M.; Critchley, A.; Sawamura, M. Growth rates and agar quality of Gracilaria gracilis (Stackhouse) Steentoft from Namibia, Southern Africa. Bot. Mar 1996, 39, 273–279. [Google Scholar] [CrossRef]
- Bixler, H.J.; Porse, H. A decade of change in the seaweed hydrocolloids industry. J. Appl. Phycol. 2011, 23, 321–335. [Google Scholar] [CrossRef]
- Bhatti, J.S.; Sehrawat, A.; Mishra, J.; Sidhu, I.S.; Navik, U.; Khullar, N.; Kumar, S.; Bhatti, G.K.; Reddy, P.H. Oxidative stress in the pathophysiology of type 2 diabetes and related complications: Current therapeutics strategies and future perspectives. Free Radic. Biol. Med. 2022, 184, 114–134. [Google Scholar] [CrossRef]
- Deshpande, A.D.; Harris-Hayes, M.; Schootman, M. Schootman: Epidemiology of diabetes and diabetes-related complications. Phys. Ther. 2008, 88, 1254–1264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daoudi, N.E.; Bouziane, O.; Bouhrim, M.; Bnouham, M. Natural aldose reductase inhibitors for treatment and prevention of diabetic cataract: A review. Herba Pol. 2022, 68, 35–58. [Google Scholar] [CrossRef]
- Scheen, A.-J. Antidiabétiques oraux dans le traitement du diabète de type 2: Perspectives historique et médico-économique. Médecine Des Mal. Métaboliques 2015, 9, 186–197. [Google Scholar] [CrossRef]
- Silverthorn, D.U. Physiologie Humaine: Une Approche Intégrée; Pearson Education France: Paris, France, 2007. [Google Scholar]
- Halimi, S. Une place pour des traitements antidiabétiques en plus de l’insuline chez les jeunes diabétiques de type 1? Médecine Des Mal. Métaboliques 2015, 9, 668–676. [Google Scholar] [CrossRef]
- Gautier, J.-F.; Choukem, S.P. Les incrétines. Nutr. Clin. Et Métabolisme 2008, 22, 59–65. [Google Scholar] [CrossRef]
- Gunathilaka, T.L.; Samarakoon, K.; Ranasinghe, P.; Peiris, L.D.C. Antidiabetic potential of marine brown algae—A mini review. J. Diabetes Res. 2020, 2020, 1230218. [Google Scholar] [CrossRef] [Green Version]
- Wong, K.H.; Cheung, P.C. Nutritional evaluation of some subtropical red and green seaweeds: Part I—Proximate composition, amino acid profiles and some physico-chemical properties. Food Chem. 2000, 71, 475–482. [Google Scholar] [CrossRef]
- Sun-Waterhouse, D.; Thakorlal, J.; Zhou, J. Effects of added phenolics on the storage stability of avocado and coconut oils. Int. J. Food Sci. Technol. 2011, 46, 1575–1585. [Google Scholar] [CrossRef]
- Damasceno, S.S.; Santos, N.A.; Santos, I.M.; Souza, A.L.; Souza, A.G.; Queiroz, N. Caffeic and ferulic acids: An investigation of the effect of antioxidants on the stability of soybean biodiesel during storage. Fuel 2013, 107, 641–646. [Google Scholar] [CrossRef]
- Kaneria, M.; Chanda, S. Evaluation of antioxidant and antimicrobial properties of Manilkara zapota L.(chiku) leaves by sequential soxhlet extraction method. Asian Pac. J. Trop. Biomed. 2012, 2, S1526–S1533. [Google Scholar] [CrossRef]
- Me, D.Y.; Me, Q.W.; Be, L.K.; Be, J.J. Antioxidant activities of various extracts of lotus (Nelumbo nuficera Gaertn) rhizome. Asia Pac. J. Clin. Nutr. 2007, 16, 158. [Google Scholar]
- Cho, S.-h.; Kang, S.-e.; Cho, J.-y.; Kim, A.-r.; Park, S.-m.; Hong, Y.-K.; Ahn, D.-H. The antioxidant properties of brown seaweed (Sargassum siliquastrum) extracts. J. Med. Food 2007, 10, 479–485. [Google Scholar] [CrossRef]
- Bandar, H.; Hijazi, A.; Rammal, H.; Hachem, A.; Saad, Z.; Badran, B. Techniques for the extraction of bioactive compounds from Lebanese Urtica Dioica. Am. J. Phytomed. Clin. Ther. 2013, 1, 507–513. [Google Scholar]
- Ramdani, M.; Elasri, O.; Saidi, N.; Elkhiati, N.; Taybi, F.A.; Mostareh, M.; Zaraali, O.; Haloui, B.; Ramdani, M. Evaluation of antioxidant activity and total phenol content of Gracilaria bursa-pastoris harvested in Nador lagoon for an enhanced economic valorization. Chem. Biol. Technol. Agric. 2017, 4, 28. [Google Scholar] [CrossRef] [Green Version]
- Gamze Yildiz, Ö.V. Serap Çelikler & Şükran Dere, Determination of the Phenolic Compounds and Antioxidative Capacity in Red Algae Gracilaria bursa-pastoris. Int. J. Food Prop. 2011, 14, 496–502. [Google Scholar]
- Terouzi, W.; Yacine, Z.A.; Hanine, H.; Boulli, A.; Oussama, A. Comparative study of physical and chemical propriety of the oil of some varieties of olive trees. Int. J. Innov. Appl. Stud. 2014, 6, 1096. [Google Scholar]
- Brahmi, F.; Haddad, S.; Bouamara, K.; Yalaoui-Guellal, D.; Prost-Camus, E.; De Barros, J.-P.P.; Prost, M.; Atanasov, A.G.; Madani, K.; Boulekbache-Makhlouf, L. Comparison of chemical composition and biological activities of Algerian seed oils of Pistacia lentiscus L. Opuntia ficus indica (L.) mill. and Argania spinosa L. Skeels. Ind. Crops Prod. 2020, 151, 112456. [Google Scholar] [CrossRef]
- Attia, Y.A.; Al-Harthi, M.A.; Korish, M.A.; Shiboob, M.M. Fatty acid and cholesterol profiles and hypocholesterolemic, atherogenic, and thrombogenic indices of table eggs in the retail market. Lipids Health Dis. 2015, 14, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, S.; Jiao, W.; Xu, Y.; Hou, L.; Li, H.; Shao, J.; Zhang, X.; Wang, R.; Kong, D. Palmitic acid inhibits prostate cancer cell proliferation and metastasis by suppressing the PI3K/Akt pathway. Life Sci. 2021, 286, 120046. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Oguzhan, B.; Louchami, K.; Chardigny, J.-M.; Portois, L.; Carpentier, Y.; Malaisse, W.-J.; Herchuelz, A.; Sener, A. Pancreatic islet function in ω-3 fatty acid-depleted rats: Alteration of calcium fluxes and calcium-dependent insulin release. Am. J. Physiol. -Endocrinol. Metab. 2006, 291, E441–E448. [Google Scholar] [CrossRef] [Green Version]
- Sawada, K.; Kawabata, K.; Yamashita, T.; Kawasaki, K.; Yamamoto, N.; Ashida, H. Ameliorative effects of polyunsaturated fatty acids against palmitic acid-induced insulin resistance in L6 skeletal muscle cells. Lipids Health Dis. 2012, 11, 36. [Google Scholar] [CrossRef] [Green Version]
- Vassiliou, E.K.; Gonzalez, A.; Garcia, C.; Tadros, J.H.; Chakraborty, G.; Toney, J.H. Oleic acid and peanut oil high in oleic acid reverse the inhibitory effect of insulin production of the inflammatory cytokine TNF-α both in vitro and in vivo systems. Lipids Health Dis. 2009, 8, 25. [Google Scholar] [CrossRef] [Green Version]
- Tsuchiya, A.; Kanno, T.; Nishizaki, T. Stearic acid serves as a potent inhibitor of protein tyrosine phosphatase 1B. Cell. Physiol. Biochem. 2013, 32, 1451–1459. [Google Scholar] [CrossRef] [Green Version]
- Nugent, C.; Prins, J.B.; Whitehead, J.P.; Wentworth, J.M.; Chatterjee, V.K.K.; O’Rahilly, S. Arachidonic acid stimulates glucose uptake in 3T3-L1 adipocytes by increasing GLUT1 and GLUT4 levels at the plasma membrane: Evidence for involvement of lipoxygenase metabolites and peroxisome proliferator-activated receptor γ. J. Biol. Chem. 2001, 276, 9149–9157. [Google Scholar] [CrossRef] [Green Version]
- Manco, M.; Calvani, M.; Mingrone, G. Effects of dietary fatty acids on insulin sensitivity and secretion. Diabetes Obes. Metab. 2004, 6, 402–413. [Google Scholar] [CrossRef] [PubMed]
- Mensink, R.P.; Zock, P.L.; Kester, A.D.; Katan, M.B. Effects of dietary fatty acids and carbohydrates on the ratio of serum total to HDL cholesterol and on serum lipids and apolipoproteins: A meta-analysis of 60 controlled trials. Am. J. Clin. Nutr. 2003, 77, 1146–1155. [Google Scholar] [CrossRef] [Green Version]
- BH, H. Omega-3 fatty acids and inflammatory processes. Pharmacol. Ther. 2002, 96, 67–202. [Google Scholar]
- Calder, P.C. Omega-3 fatty acids and inflammatory processes. Nutrients 2010, 2, 355–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.; Ebersole, J. A novel bioactivity of omega-3 polyunsaturated fatty acids and their ester derivatives. Mol. Oral Microbiol. 2010, 25, 75–80. [Google Scholar] [CrossRef]
- Guedes, A.C.; Amaro, H.M.; Malcata, F.X. Microalgae as sources of high added-value compounds—A brief review of recent work. Biotechnol. Prog. 2011, 27, 597–613. [Google Scholar] [CrossRef]
- Pulz, O.; Gross, W. Valuable products from biotechnology of microalgae. Appl. Microbiol. Biotechnol. 2004, 65, 635–648. [Google Scholar] [CrossRef]
- Schmitz, G.; Ecker, J. The opposing effects of n− 3 and n− 6 fatty acids. Prog. Lipid Res. 2008, 47, 147–155. [Google Scholar] [CrossRef]
- Plaza, M.; Herrero, M.; Cifuentes, A.; Ibanez, E. Innovative natural functional ingredients from microalgae. J. Agric. Food Chem. 2009, 57, 7159–7170. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Wu, J.H. Omega-3 fatty acids and cardiovascular disease: Effects on risk factors, molecular pathways, and clinical events. J. Am. Coll. Cardiol. 2011, 58, 2047–2067. [Google Scholar] [CrossRef] [Green Version]
- Field, C.J.; Schley, P.D. Evidence for potential mechanisms for the effect of conjugated linoleic acid on tumor metabolism and immune function: Lessons from n− 3 fatty acids. Am. J. Clin. Nutr. 2004, 79, 1190S–1198S. [Google Scholar] [CrossRef] [Green Version]
- Das, M.; Zuniga, E.; Ojima, I. Novel taxoid-based tumor-targeting drug conjugates. Chim. Oggi 2009, 27, 54. [Google Scholar]
- Radwan, S.S. Sources of C20-polyunsaturated fatty acids for biotechnological use. Appl. Microbiol. Biotechnol. 1991, 35, 421–430. [Google Scholar] [CrossRef]
- Razzaghi-Asl, N.; Garrido, J.; Khazraei, H.; Borges, F.; Firuzi, O. Antioxidant properties of hydroxycinnamic acids: A review of structure-activity relationships. Curr. Med. Chem. 2013, 20, 4436–4450. [Google Scholar] [CrossRef] [Green Version]
- Silva, F.A.; Borges, F.; Guimarães, C.; Lima, J.L.; Matos, C.; Reis, S. Phenolic acids and derivatives: Studies on the relationship among structure, radical scavenging activity, and physicochemical parameters. J. Agric. Food Chem. 2000, 48, 2122–2126. [Google Scholar] [CrossRef]
- Sroka, Z.; Cisowski, W. Hydrogen peroxide scavenging, antioxidant and anti-radical activity of some phenolic acids. Food Chem. Toxicol. 2003, 41, 753–758. [Google Scholar] [CrossRef]
- Mirzaei, S.; Gholami, M.H.; Zabolian, A.; Saleki, H.; Farahani, M.V.; Hamzehlou, S.; Far, F.B.; Sharifzadeh, S.O.; Samarghandian, S.; Khan, H. Caffeic acid and its derivatives as potential modulators of oncogenic molecular pathways: New hope in the fight against cancer. Pharmacol. Res. 2021, 171, 105759. [Google Scholar] [CrossRef] [PubMed]
- Sul, D.; Kim, H.-S.; Lee, D.; Joo, S.S.; Hwang, K.W.; Park, S.-Y. Protective effect of caffeic acid against beta-amyloid-induced neurotoxicity by the inhibition of calcium influx and tau phosphorylation. Life Sci. 2009, 84, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.-D.; Lee, S.E.; Park, Y.S.; Shin, D.-H.; Park, G.G.; Park, C.-S. Immunosuppressive effects of fisetin against dinitrofluorobenzene-induced atopic dermatitis-like symptoms in NC/Nga mice. Food Chem. Toxicol. 2014, 66, 341–349. [Google Scholar] [CrossRef]
- Khan, N.; Afaq, F.; Khusro, F.H.; Mustafa Adhami, V.; Suh, Y.; Mukhtar, H. Dual inhibition of phosphatidylinositol 3-kinase/Akt and mammalian target of rapamycin signaling in human nonsmall cell lung cancer cells by a dietary flavonoid fisetin. Int. J. Cancer 2012, 130, 1695–1705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pal, H.C.; Sharma, S.; Elmets, C.A.; Athar, M.; Afaq, F. Fisetin inhibits growth, induces G 2/M arrest and apoptosis of human epidermoid carcinoma A 431 cells: Role of mitochondrial membrane potential disruption and consequent caspases activation. Exp. Dermatol. 2013, 22, 470–475. [Google Scholar] [CrossRef] [Green Version]
- Prasath, G.S.; Subramanian, S.P. Modulatory effects of fisetin, a bioflavonoid, on hyperglycemia by attenuating the key enzymes of carbohydrate metabolism in hepatic and renal tissues in streptozotocin-induced diabetic rats. Eur. J. Pharmacol. 2011, 668, 492–496. [Google Scholar] [CrossRef]
- Pal, H.C.; Pearlman, R.L.; Afaq, F. Fisetin and Its Role in Chronic Diseases. In Anti-Inflammatory Nutraceuticals and Chronic Diseases; Springer: Cham, Switzerland, 2016; pp. 213–244. [Google Scholar]
- Benchikh, Y.; Louaileche, H.; George, B.; Merlin, A. Changes in bioactive phytochemical content and in vitro antioxidant activity of carob (Ceratonia siliqua L.) as influenced by fruit ripening. Ind. Crops Prod. 2014, 60, 298–303. [Google Scholar] [CrossRef]
- Singh, M.; Lee, K.E.; Vinayagam, R.; Kang, S.G. Antioxidant and antibacterial profiling of pomegranate-pericarp extract functionalized-zinc oxide nanocomposite. Biotechnol. Bioprocess Eng. 2021, 26, 728–737. [Google Scholar] [CrossRef] [PubMed]
- Kwon, N.; Vinayagam, R.; Do, G.S.; Lee, K.E.; Kang, S.G. Protective Effects of Fermented Houttuynia cordata Against UVA and H2O2-Induced Oxidative Stress in Human Skin Keratinocytes. Appl. Biochem. Biotechnol. 2023, 195, 3027–3046. [Google Scholar] [CrossRef] [PubMed]
- Prieto, M.; Rodríguez-Amado, I.; Vázquez, J.A.; Murado, M. β-Carotene assay revisited. Application to characterize and quantify antioxidant and prooxidant activities in a microplate. J. Agric. Food Chem. 2012, 60, 8983–8993. [Google Scholar] [CrossRef] [Green Version]
- Ganesan, K.; Rana, M.B.M.; Sultan, S. Oral Hypoglycemic Medications; StatPearls: Tampa, FL, USA, 2018. [Google Scholar]
- Marghich, M.; Daoudi, N.E.; Amrani, O.; Addi, M.; Hano, C.; Chen, J.-T.; Mekhfi, H.; Ziyyat, A.; Bnouham, M.; Aziz, M. Antioxidant Activity and Inhibition of Carbohydrate Digestive Enzymes Activities of Artemisia campestris L. Front. Biosci. Sch. 2022, 14, 25. [Google Scholar] [CrossRef]
- Date, K.; Satoh, A.; Iida, K.; Ogawa, H. Pancreatic α-amylase controls glucose assimilation by duodenal retrieval through N-glycan-specific binding, endocytosis, and degradation. J. Biol. Chem. 2015, 290, 17439–17450. [Google Scholar] [CrossRef] [Green Version]
- Daoudi, N.E.; Bouhrim, M.; Ouassou, H.; Legssyer, A.; Mekhfi, H.; Ziyyat, A.; Aziz, M.; Bnouham, M. Inhibitory effect of roasted/unroasted Argania spinosa seeds oil on α-glucosidase, α-amylase and intestinal glucose absorption activities. S. Afr. J. Bot. 2020, 135, 413–420. [Google Scholar] [CrossRef]
- Lordan, S.; Smyth, T.J.; Soler-Vila, A.; Stanton, C.; Ross, R.P. The α-amylase and α-glucosidase inhibitory effects of Irish seaweed extracts. Food Chem. 2013, 141, 2170–2176. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, Y.; Suzuki, M.; Satsu, H.; Arai, S.; Hara, Y.; Suzuki, K.; Miyamoto, Y.; Shimizu, M. Green tea polyphenols inhibit the sodium-dependent glucose transporter of intestinal epithelial cells by a competitive mechanism. J. Agric. Food Chem. 2000, 48, 5618–5623. [Google Scholar] [CrossRef]
- Aleixandre, A.; Gil, J.V.; Sineiro, J.; Rosell, C.M. Understanding phenolic acids inhibition of α-amylase and α-glucosidase and influence of reaction conditions. Food Chem. 2022, 372, 131231. [Google Scholar] [CrossRef]
- Li, K.; Yao, F.; Xue, Q.; Fan, H.; Yang, L.; Li, X.; Sun, L.; Liu, Y. Inhibitory effects against α-glucosidase and α-amylase of the flavonoids-rich extract from Scutellaria baicalensis shoots and interpretation of structure–activity relationship of its eight flavonoids by a refined assign-score method. Chem. Cent. J. 2018, 12, 82. [Google Scholar] [CrossRef] [PubMed]
- Lou, W.; Chen, Y.; Ma, H.; Liang, G.; Liu, B. Antioxidant and α-amylase inhibitory activities of tannic acid. J. Food Sci. Technol. 2018, 55, 3640–3646. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Moneim, A.; Abd El-Twab, S.M.; Yousef, A.I.; Reheim, E.S.A.; Ashour, M.B. Modulation of hyperglycemia and dyslipidemia in experimental type 2 diabetes by gallic acid and p-coumaric acid: The role of adipocytokines and PPARγ. Biomed. Pharmacother. 2018, 105, 1091–1097. [Google Scholar] [CrossRef]
- Panda, S.; Kar, A. Apigenin (4′, 5, 7-trihydroxyflavone) regulates hyperglycaemia, thyroid dysfunction and lipid peroxidation in alloxan-induced diabetic mice. J. Pharm. Pharmacol. 2007, 59, 1543–1548. [Google Scholar] [CrossRef] [PubMed]
- Alkhalidy, H.; Moore, W.; Wang, A.; Luo, J.; McMillan, R.P.; Wang, Y.; Zhen, W.; Hulver, M.W.; Liu, D. Kaempferol ameliorates hyperglycemia through suppressing hepatic gluconeogenesis and enhancing hepatic insulin sensitivity in diet-induced obese mice. J. Nutr. Biochem. 2018, 58, 90–101. [Google Scholar] [CrossRef]
- Srinivasan, P.; Vijayakumar, S.; Kothandaraman, S.; Palani, M. Anti-diabetic activity of quercetin extracted from Phyllanthus emblica L. fruit: In silico and in vivo approaches. J. Pharm. Anal. 2018, 8, 109–118. [Google Scholar] [CrossRef]
- Meng, S.; Cao, J.; Feng, Q.; Peng, J.; Hu, Y. Roles of chlorogenic acid on regulating glucose and lipids metabolism: A review. Evid. Based Complement. Altern. Med. eCAM 2013, 2013, 801457. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, S.; Muthukumaran, J.; Muruganathan, U.; Venkatesan, R.S.; Jalaludeen, A.M. Antihyperglycemic effect of syringic acid on attenuating the key enzymes of carbohydrate metabolism in experimental diabetic rats. Biomed. Prev. Nutr. 2014, 4, 595–602. [Google Scholar] [CrossRef]
- Annadurai, T.; Muralidharan, A.R.; Joseph, T.; Hsu, M.; Thomas, P.; Geraldine, P. Antihyperglycemic and antioxidant effects of a flavanone, naringenin, in streptozotocin–nicotinamide-induced experimental diabetic rats. J. Physiol. Biochem. 2012, 68, 307–318. [Google Scholar] [CrossRef]
- Den Hartogh, D.J.; Tsiani, E. Antidiabetic properties of naringenin: A citrus fruit polyphenol. Biomolecules 2019, 9, 99. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Du, X.; Guo, P.; Huang, L.; Qi, P.; Gong, Q. Ascorbic acid supplementation in type 2 diabetes mellitus: A protocol for systematic review and meta-analysis. Medicine 2020, 99, e23125. [Google Scholar] [CrossRef] [PubMed]
- Manning, P.J.; Sutherland, W.H.; Walker, R.J.; Williams, S.M.; De Jong, S.A.; Ryalls, A.R.; Berry, E.A. Effect of high-dose vitamin E on insulin resistance and associated parameters in overweight subjects. Diabetes Care 2004, 27, 2166–2171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, H.-h.; Shao, Z.-m.; Tang, D.-q.; Lu, Q.; Chen, X.; Yin, X.-x.; Wu, J.; Chen, H. Preventive effects of rutin on the development of experimental diabetic nephropathy in rats. Life Sci. 2012, 91, 959–967. [Google Scholar] [CrossRef] [PubMed]
- Mechchate, H.; Es-Safi, I.; Haddad, H.; Bekkari, H.; Grafov, A.; Bousta, D. Combination of Catechin, Epicatechin, and Rutin: Optimization of a novel complete antidiabetic formulation using a mixture design approach. J. Nutr. Biochem. 2021, 88, 108520. [Google Scholar] [CrossRef]
- Ouffai, K.; Azzi, R.; Abbou, F.; Mahdi, S.; El Haci, I.A.; Belyagoubi-Benhammou, N.; Bekkara, F.A.; Lahfa, F.B. Phenolics compounds, evaluation of Alpha-amylase, alpha-glucosidase inhibitory capacity and antioxidant effect from Globularia alypum L. Vegetos 2021, 34, 477–484. [Google Scholar] [CrossRef]
- Luliana, S.; Desnita, R.; Martien, R.; Nurrochmad, A. Total flavonoid contents and in silico study of flavonoid compounds from Meniran (Phyllanthus niruri L.) towards alpha-amylase and alpha-glucosidase enzyme. Pharmaciana 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Martinez-Gonzalez, A.I.; Díaz-Sánchez, Á.G.; de la Rosa, L.A.; Vargas-Requena, C.L.; Bustos-Jaimes, I.; Alvarez-Parrilla, E. Polyphenolic compounds and digestive enzymes: In vitro non-covalent interactions. Molecules 2017, 22, 669. [Google Scholar] [CrossRef] [Green Version]
- Teng, H.; Chen, L. α-Glucosidase and α-amylase inhibitors from seed oil: A review of liposoluble substance to treat diabetes. Crit. Rev. Food Sci. Nutr. 2017, 57, 3438–3448. [Google Scholar] [CrossRef]
- Chen, L.; Kang, Y.-H. In vitro inhibitory effect of oriental melon (Cucumis melo L. var. makuwa Makino) seed on key enzyme linked to type 2 diabetes: Assessment of anti-diabetic potential of functional food. J. Funct. Foods 2013, 5, 981–986. [Google Scholar] [CrossRef]
- Huie, C.W. A review of modern sample-preparation techniques for the extraction and analysis of medicinal plants. Anal. Bioanal. Chem. 2002, 373, 23–30. [Google Scholar] [CrossRef]
- Kim, J.-H.; Park, S.-M.; Ha, H.-J.; Moon, C.-J.; Shin, T.-K.; Kim, J.-M.; Lee, N.-H.; Kim, H.-C.; Jang, K.-J.; Wie, M.-B. Opuntia ficus-indica attenuates neuronal injury in in vitro and in vivo models of cerebral ischemia. J. Ethnopharmacol. 2006, 104, 257–262. [Google Scholar] [CrossRef]
- Loukili, E.; Abrigach, F.; Bouhrim, M.; Bnouham, M.; Fauconnier, M.-l.; Ramdani, M. Chemical composition and physicochemical analysis of Opuntia dillenii extracts grown in Morocco. J. Chem. 2021, 2021, 8858929. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.-E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Bekkouch, O.; Harnafi, M.; Touiss, I.; Khatib, S.; Harnafi, H.; Alem, C.; Amrani, S. In vitro antioxidant and in vivo lipid-lowering properties of Zingiber officinale crude aqueous extract and methanolic fraction: A follow-up study. Evid. Based Complement. Altern. Med. 2019, 2019, 9734390. [Google Scholar] [CrossRef] [Green Version]
- Hbika, A.; Daoudi, N.E.; Bouyanzer, A.; Bouhrim, M.; Mohti, H.; Loukili, E.H.; Mechchate, H.; Al-Salahi, R.; Nasr, F.A.; Bnouham, M. Artemisia absinthium L. Aqueous and Ethyl Acetate Extracts: Antioxidant Effect and Potential Activity In Vitro and In Vivo against Pancreatic α-Amylase and Intestinal α-Glucosidase. Pharmaceutics 2022, 14, 481. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Rana, H.K.; Singh, V.; Yadav, T.C.; Varadwaj, P.; Pandey, A.K. Evaluation of antidiabetic activity of dietary phenolic compound chlorogenic acid in streptozotocin induced diabetic rats: Molecular docking, molecular dynamics, in silico toxicity, in vitro and in vivo studies. Comput. Biol. Med. 2021, 134, 104462. [Google Scholar] [CrossRef] [PubMed]
- Aboul-Soud, M.A.; Ennaji, H.; Kumar, A.; Alfhili, M.A.; Bari, A.; Ahamed, M.; Chebaibi, M.; Bourhia, M.; Khallouki, F.; Alghamdi, K.M. Antioxidant, Anti-Proliferative Activity and Chemical Fingerprinting of Centaurea calcitrapa against Breast Cancer Cells and Molecular Docking of Caspase-3. Antioxidants 2022, 11, 1514. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Solvent | Extraction Methods | Yield (%) | Polyphenols (mg GAE/g) | Flavonoids (mg QE/g) |
|---|---|---|---|---|
| Hexane | M | 0.11 ± 0.09 | - | - |
| S | 1.16 ± 0.34 | - | - | |
| Ethyl acetate | M | 0.21 ± 0.04 | 165.42 ± 0.21 | 84.53 ± 0.07 |
| S | 1.80 ± 0.08 | 150.24 ± 0.11 | 75.47 ± 0.02 | |
| Methanol | M | 0.81 ± 0.26 | 37.69 ± 1.02 | 15.29 ± 0.18 |
| S | 3.42 ± 0.07 | 28.68 ± 0.07 | 13.70 ± 0.03 | |
| Water | M | 1.25 ± 0.06 | 381.31 ± 0.33 | 201.80 ± 0.21 |
| Fatty Acids | RT (min) | HE (%) | EAcE (%) | ||
|---|---|---|---|---|---|
| M | S | M | S | ||
| Eicosenoic acid (C20:1) | 20.08 | 53.06 ± 0.05 | 33.25 ± 0.41 | 5.21 ± 0.01 | nd |
| 7,10-Hexadecadienoic acid (C16:2) | 21.17 | 2.88 ± 0.23 | 5.40 ± 0.01 | 5.67 ± 0.03 | 3.84 ± 0.06 |
| Palmitic acid (C16:0) | 23.30 | 22.88 ± 0.01 | 40.72 ± 0.31 | 50.55 ± 0.05 | 38.24 ± 0.02 |
| Margaric acid (C17:0) | 23.87 | nd | nd | nd | 16.19 ± 0.07 |
| Oleic acid (C18:1) | 24.54 | 3.32 ± 0.07 | nd | 10.48 ± 0.02 | 13.01 ± 0.09 |
| Linoleic acid (C18:2) | 25.03 | 9.51 ± 0.31 | 10.08 ± 0.11 | 15.57 ± 0.23 | 23.85 ± 0.11 |
| Linolenic acid (C18:3) | 25.09 | 3.58 ± 0.04 | 4.49 ± 0.17 | 6.09 ± 0.06 | nd |
| Stearic acid (C18:0) | 25.26 | 4.77 ± 0.43 | 6.06 ± 0.08 | 6.43 ± 0.07 | 4.87 ± 0.04 |
| SFA a | 27.65 | 46.78 | 56.98 | 59.3 | |
| UFA b | 72.35 | 53.22 | 43.02 | 40.7 | |
| UFA/SFA c | 2.61 | 1.13 | 0.75 | 0.68 | |
| N° | Compounds | RT (min) | EAcE (%) | ME (%) | ||
|---|---|---|---|---|---|---|
| M | S | M | S | |||
| 1 | Gallic acid | 15.47 | nd | nd | nd | 0.637 |
| 2 | Catechin | 18.68 | 1.516 | 8.188 | nd | nd |
| 3 | 4-hydroxy benzoïc acid | 18.91 | 4.105 | 18.778 | nd | 3.561 |
| 4 | Chlorogenic acid | 19.15 | 1.096 | 4.202 | 11.666 | 7.714 |
| 5 | Caffeic acid | 19.45 | nd | nd | 35.642 | 24.239 |
| 6 | Syringic acid | 19.74 | nd | 3.092 | 3.673 | nd |
| 7 | Vanilline | 23.1 | 3.457 | nd | nd | nd |
| 8 | p-Coumaric acid | 23.63 | nd | nd | nd | 5.269 |
| 9 | Sinapic acid | 24.09 | 9.534 | nd | nd | nd |
| 11 | Quercetin-O-3-glucoside | 24.52 | 8.074 | nd | nd | nd |
| 12 | 7,3′,4′-flavon-3-ol | 24.92 | 8.316 | 20.682 | nd | nd |
| 13 | Naringin | 25.01 | 8.344 | 17.754 | 21.069 | 12.626 |
| 14 | Rutin | 25.16 | 8.291 | nd | nd | nd |
| 15 | Salicylic acid | 25.32 | nd | 15.613 | 14.901 | 11.854 |
| 16 | Quercetin | 25.46 | 8.669 | nd | nd | nd |
| 17 | Cinnamic acid | 25.48 | 8.941 | nd | nd | 9.662 |
| 18 | Luteolin | 25.64 | 8.912 | nd | nd | 9.387 |
| 19 | Apigenin | 25.87 | 7.839 | nd | nd | nd |
| 20 | Kaempferol | 26.1 | 9.108 | 11.690 | 8.100 | 9.109 |
| 21 | Flavone | 26.92 | nd | nd | 4.948 | 5.940 |
| 22 | Flavanone | 27.412 | nd | 3.795 | nd | nd |
| Extracts | IC50 (mg/mL) | ||
|---|---|---|---|
| DPPH | β-Carotene | ||
| EAcE | M | 0.40 ± 0.51 | 0.83 ± 0.32 |
| S | 0.30 ± 0.31 | 0.65 ± 0.04 | |
| ME | M | 0.55 ± 0.25 | 0.07 ± 0.06 |
| S | 0.46 ± 0.36 | 0.06 ± 0.64 | |
| AQE | M | 0.17 ± 0.07 | 0.21 ± 0.14 |
| Ascorbic Acid | 0.06 | - | |
| BHA | - | 0.02 | |
| Inhibitors | IC50 (mg/mL) | ||
|---|---|---|---|
| α-Amylase | α-Glucosidase | ||
| Acarbose | 0.35 ± 0.08 | 0.39 ± 0.04 | |
| EAcE | M | 1.86 ± 0.06 *** | 0.44 ± 0.02 ns |
| S | 1.11 ± 0.02 *** | 0.57 ± 0.03 ** | |
| ME | M | 0.72 ± 0.04 ** | 0.25 ± 0.09 * |
| S | 0.76 ± 0.05 ** | 0.37 ± 0.06 ns | |
| AQE | M | 0.85 ± 0.01 *** | 0.32 ± 0.07 ns |
| α-Amylase (PDB: 1B2Y) | α-Gluosidase (PDB: 5NN8) | |||||
|---|---|---|---|---|---|---|
| Glide Gscore | Glide Emodel | Glide Energy | Glide Gscore | Glide Emodel | Glide Energy | |
| 4-Hydroxybenzoic acid | −6.079 | −27.887 | −18.583 | −4.366 | −20.091 | −13.882 |
| 7,3′,4′-flavon-3-ol | −4.801 | −48.996 | −38.542 | −3.695 | −36.338 | −30.331 |
| Apigenin | −7.105 | −60.626 | −41.637 | −5.2 | −44.196 | −33.441 |
| Caffeic Acid | −5.893 | −39.405 | −29.08 | −4.237 | −26.17 | −18.858 |
| Chlorogenic acid | −6.801 | −63.522 | −45.48 | −3.738 | −43.188 | −37.071 |
| Cinnamic acid | −3.632 | −22.468 | −18.689 | −3.353 | −19.589 | −14.942 |
| Flavanone | −5.798 | −41.242 | −30.237 | −4.287 | −35.284 | −27.649 |
| Flavone | −6.121 | −43.458 | −31.445 | −4.326 | −35.845 | −28.673 |
| Kaempferol | −7.2 | −63.076 | −43.51 | −5.698 | −47.8 | −35.946 |
| Luteolin | −6.366 | −52.124 | −38.738 | −5.425 | −51.621 | −37.184 |
| Naringenin | −7.198 | −60.107 | −40.882 | −5.246 | −38.138 | −29.883 |
| p-coumaric acid | −5.395 | −35.602 | −26.441 | −3.558 | −21.069 | −15.352 |
| Quercetin | −7.146 | −67.907 | −45.267 | −7.035 | −60.46 | −42.363 |
| Quercetin 3-glucoside | −6.402 | −67.452 | −49.322 | −5.242 | −60.877 | −49.104 |
| Rutin | −8.109 | −108.772 | −71.453 | −5.237 | −73.009 | −57.522 |
| Salicylic Acid | −4.06 | −23.996 | −18.556 | −3.911 | −15.12 | −12.243 |
| Sinapic acid | −3.894 | −30.39 | −24.001 | −3.543 | −27.834 | −22.072 |
| Syringic acid | −5.104 | −32.584 | −24.351 | −3.472 | −23.213 | −19.151 |
| Vanillin | −5.498 | −24.397 | −17.121 | −4.651 | −30.76 | −23.024 |
| Acarbose | −7.067 | −108.733 | −73.113 | −6.989 | −111.624 | −70.347 |
| Metformin | −3.989 | −35.344 | −22.492 | −4.494 | −36.414 | −21.184 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ouahabi, S.; Loukili, E.H.; Daoudi, N.E.; Chebaibi, M.; Ramdani, M.; Rahhou, I.; Bnouham, M.; Fauconnier, M.-L.; Hammouti, B.; Rhazi, L.; et al. Study of the Phytochemical Composition, Antioxidant Properties, and In Vitro Anti-Diabetic Efficacy of Gracilaria bursa-pastoris Extracts. Mar. Drugs 2023, 21, 372. https://doi.org/10.3390/md21070372
Ouahabi S, Loukili EH, Daoudi NE, Chebaibi M, Ramdani M, Rahhou I, Bnouham M, Fauconnier M-L, Hammouti B, Rhazi L, et al. Study of the Phytochemical Composition, Antioxidant Properties, and In Vitro Anti-Diabetic Efficacy of Gracilaria bursa-pastoris Extracts. Marine Drugs. 2023; 21(7):372. https://doi.org/10.3390/md21070372
Chicago/Turabian StyleOuahabi, Safae, El Hassania Loukili, Nour Elhouda Daoudi, Mohamed Chebaibi, Mohamed Ramdani, Ilyesse Rahhou, Mohamed Bnouham, Marie-Laure Fauconnier, Belkheir Hammouti, Larbi Rhazi, and et al. 2023. "Study of the Phytochemical Composition, Antioxidant Properties, and In Vitro Anti-Diabetic Efficacy of Gracilaria bursa-pastoris Extracts" Marine Drugs 21, no. 7: 372. https://doi.org/10.3390/md21070372
APA StyleOuahabi, S., Loukili, E. H., Daoudi, N. E., Chebaibi, M., Ramdani, M., Rahhou, I., Bnouham, M., Fauconnier, M. -L., Hammouti, B., Rhazi, L., Ayerdi Gotor, A., Dépeint, F., & Ramdani, M. (2023). Study of the Phytochemical Composition, Antioxidant Properties, and In Vitro Anti-Diabetic Efficacy of Gracilaria bursa-pastoris Extracts. Marine Drugs, 21(7), 372. https://doi.org/10.3390/md21070372