

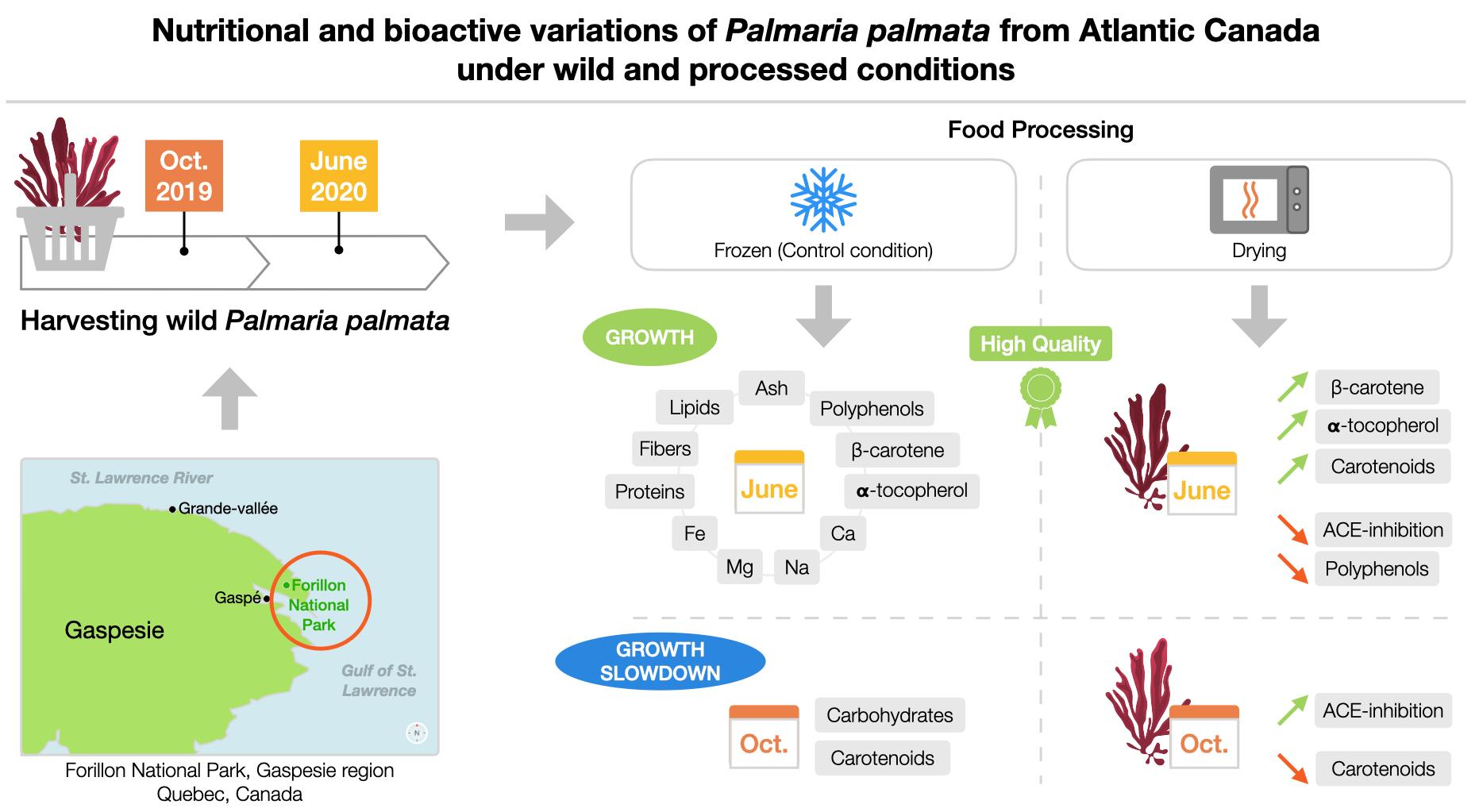
Impact of Harvest Month and Drying Process on the Nutritional and Bioactive Properties of Wild Palmaria palmata from Atlantic Canada

 , and
, and
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Chemical Composition
2.1.1. Crude Palmaria palmata
2.1.2. Dried Palmaria palmata
2.2. Mineral Composition
2.2.1. Crude Palmaria palmata
2.2.2. Dried Palmaria palmata
2.3. Bioactive Compound Content
2.3.1. Crude Palmaria palmata
2.3.2. Dried Palmaria palmata
2.4. In Vitro Bioactive Potential
2.4.1. Protein Contents and Extraction Yields of Soluble Palmaria palmata Extracts
- Crude Palmaria palmata
- Dried Palmaria palmata
2.4.2. Antioxidant Capacity of Water-Soluble Palmaria palmata Extracts
- Crude Palmaria palmata extracts
- Dried Palmaria palmata Extracts
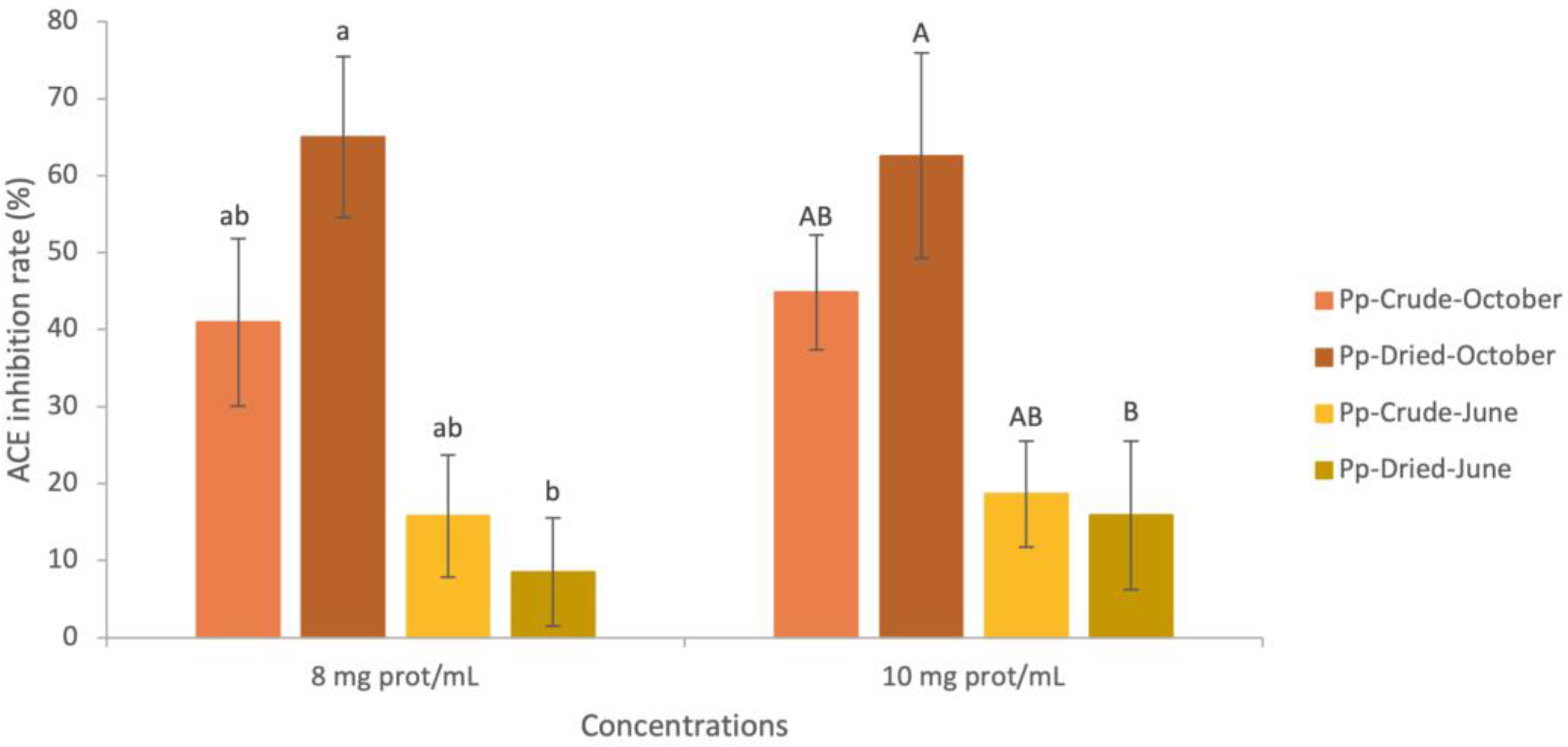
2.4.3. ACE-Inhibition Capacity of Water-Soluble Palmaria palmata Extracts
- Crude Palmaria palmata Extracts
- Dried Palmaria palmata Extracts
3. Materials and Methods
3.1. Macroalgal Biomass and Processing
3.2. Chemical Composition (Lipids, Ash, Proteins, Carbohydrates, Fibers) and Mineral Composition
3.3. Potential Bioactive Compound Determination
3.3.1. Total Phenolic Content (TPC) and Carotenoids
3.3.2. Vitamins (β-Carotene and α-Tocopherol)
3.4. In Vitro Bioactive Potential
3.4.1. Water-Soluble P. palmata Extracts
3.4.2. Oxygen Radical Absorbance Capacity (Orac) Assay
3.4.3. Ferric Ion Reducing Antioxidant Power (FRAP) Assay
3.4.4. Angiotensin-Converting Enzyme (Ace) Inhibition
3.5. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yue, S.; Munir, I.U.; Hyder, S.; Nassani, A.A.; Qazi Abro, M.M.; Zaman, K. Sustainable food production, forest biodiversity and mineral pricing: Interconnected global issues. Resour. Policy 2020, 65, 101583. [Google Scholar] [CrossRef]
- Hsu, S.Y.; Wang, H.C.; Ho, J.L.; Chen, H.C. Exploring Consumers’ Interest in Choosing Sustainable Food. Front. Psychol. 2020, 11, 489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grote, B. Recent developments in aquaculture of Palmaria palmata (Linnaeus) (Weber & Mohr 1805): Cultivation and uses. Rev. Aquac. 2019, 11, 25–41. [Google Scholar] [CrossRef] [Green Version]
- Mouritsen, O.G.; Rhatigan, P.; Pérez-Lloréns, J.L. The rise of seaweed gastronomy: Phycogastronomy. Bot. Mar. 2019, 62, 195–209. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture 2022; FAO: Rome, Italy, 2022; ISBN 978-92-5-136364-5. [Google Scholar]
- Mouritsen, O.G.; Rhatigan, P.; Pérez-Lloréns, J.-L. World cuisine of seaweeds: Science meets gastronomy. Int. J. Gastron. Food Sci. 2018, 14, 55–65. [Google Scholar] [CrossRef]
- Rioux, L.-E.; Beaulieu, L.; Turgeon, S.L. Seaweeds: A traditional ingredients for new gastronomic sensation. Food Hydrocoll. 2017, 68, 255–265. [Google Scholar] [CrossRef]
- Dupré-Gilbert, È.; Berger, K.; Gendron-Lemieux, I.; Sirois, M.; Gaudreault, S. Stratégie Algues; Merinov: Grande-Rivière, QC, Canada, 2018; p. 117. [Google Scholar]
- Bleakley, S.; Hayes, M. Algal Proteins: Extraction, Application, and Challenges Concerning Production. Foods 2017, 6, 33. [Google Scholar] [CrossRef] [Green Version]
- Scieszka, S.; Klewicka, E. Algae in food: A general review. Crit. Rev. Food Sci. Nutr. 2019, 59, 3538–3547. [Google Scholar] [CrossRef]
- Faes, V.A.; Viejo, R.M. Structure and dynamics of a population of Palmaria palmata (rhodophyta) in Northern Spain. J. Phycol. 2003, 39, 1038–1049. [Google Scholar] [CrossRef]
- Stévant, P.; Schmedes, P.S.; Le Gall, L.; Wegeberg, S.; Dumay, J.; Rebours, C. Concise review of the red macroalga dulse, Palmaria palmata (L.) Weber & Mohr. J. Appl. Phycol. 2023, 1, 28. [Google Scholar] [CrossRef]
- Yuan, Y.V.; Carrington, M.F.; Walsh, N.A. Extracts from dulse (Palmaria palmata) are effective antioxidants and inhibitors of cell proliferation in vitro. Food Chem. Toxicol. 2005, 43, 1073–1081. [Google Scholar] [CrossRef] [PubMed]
- Cherry, P.; O’Hara, C.; Magee, P.J.; McSorley, E.M.; Allsopp, P.J. Risks and benefits of consuming edible seaweeds. Nutr. Rev. 2019, 77, 307–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wells, M.L.; Potin, P.; Craigie, J.S.; Raven, J.A.; Merchant, S.S.; Helliwell, K.E.; Smith, A.G.; Camire, M.E.; Brawley, S.H. Algae as nutritional and functional food sources: Revisiting our understanding. J. Appl. Phycol. 2017, 29, 949–982. [Google Scholar] [CrossRef]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Parjikolaei, B.R.; Bruhn, A.; Eybye, K.L.; Larsen, M.M.; Rasmussen, M.B.; Christensen, K.V.; Fretté, X.C. Valuable Biomolecules from Nine North Atlantic Red Macroalgae: Amino Acids, Fatty Acids, Carotenoids, Minerals and Metals. Nat. Resour. 2016, 7, 157–183. [Google Scholar] [CrossRef] [Green Version]
- Beaulieu, L.; Sirois, M.; Tamigneaux, É. Evaluation of the in vitro biological activity of protein hydrolysates of the edible red alga, Palmaria palmata (dulse) harvested from the Gaspe coast and cultivated in tanks. J. Appl. Phycol. 2016, 28, 3101–3115. [Google Scholar] [CrossRef]
- Bondu, S.; Bonnet, C.; Gaubert, J.; Deslandes, É.; Turgeon, S.L.; Beaulieu, L. Bioassay-guided fractionation approach for determination of protein precursors of proteolytic bioactive metabolites from macroalgae. J. Appl. Phycol. 2014, 27, 2059–2074. [Google Scholar] [CrossRef]
- Tanna, B.; Mishra, A. Nutraceutical Potential of Seaweed Polysaccharides: Structure, Bioactivity, Safety, and Toxicity. Compr. Rev. Food Sci. Food Saf. 2019, 18, 817–831. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Jónsdóttir, R.; Kristinsson, H.G.; Hreggvidsson, G.O.; Jónsson, J.Ó.; Thorkelsson, G.; Ólafsdóttir, G. Enzyme-enhanced extraction of antioxidant ingredients from red algae Palmaria palmata. Food Sci. Technol. 2010, 43, 1387–1393. [Google Scholar] [CrossRef]
- Bartsch, I.; Wiencke, C.; Bischof, K.; Buchholz, C.M.; Buck, B.H.; Eggert, A.; Feuerpfeil, P.; Hanelt, D.; Jacobsen, S.; Karez, R.; et al. The genus Laminaria sensu lato: Recent insights and developments. Eur. J. Phycol. 2008, 43, 1–86. [Google Scholar] [CrossRef]
- Brody, S.R.; Lozier, M.S.; Dunne, J.P. A comparison of methods to determine phytoplankton bloom initiation. J. Geophys. Res. Ocean. 2013, 118, 2345–2357. [Google Scholar] [CrossRef]
- Galland-Irmouli, A.; Fleurence, J.; Lamghari, R.; Luçon, L.; Rouxel, C.B.O.; Bronowicki, J.V.C.; Guéant, J. Nutritional value of proteins from edible seaweed Palmaria palmata (dulse). J. Nutr. Biochem. 1999, 10, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Schmid, M.; Guihéneuf, F.; Stengel, D.B. Ecological and commercial implications of temporal and spatial variability in the composition of pigments and fatty acids in five Irish macroalgae. Mar. Biol. 2017, 164, 158. [Google Scholar] [CrossRef]
- Martinez, B.; Rico, J.M. Changes in Nutrient Content of Palmaria palmata in Response to Variable Light and Upwelling in Northern Spain. J. Phycol. 2008, 44, 50–59. [Google Scholar] [CrossRef]
- Harnedy, P.A.; Soler-Vila, A.; Edwards, M.D.; FitzGerald, R.J. The effect of time and origin of harvest on the in vitro biological activity of Palmaria palmata protein hydrolysates. Food Res. Int. 2014, 62, 746–752. [Google Scholar] [CrossRef]
- Parjikolaei, B.R.; Kloster, L.; Bruhn, A.; Rasmussen, M.B.; Fretté, X.C.; Christensen, K.V. Effect of light quality and nitrogen availability on the biomass production and pigment content of Palmaria palmata (Rhodophyta). Chem. Eng. Trans. 2013, 32, 967–972. [Google Scholar]
- Vasconcelos, M.M.M.; Vollet Marson, G.; Turgeon, S.L.; Tamigneaux, É.; Beaulieu, L. Environmental conditions influence on the physicochemical properties of wild and cultivated Palmaria palmata in the Canadian Atlantic shore. J. Appl. Phycol. 2022, 34, 2565–2578. [Google Scholar] [CrossRef]
- Bartle, R.S.; Macleod, A. The effect of natural convection air temperature on the drying kinetics and desorption isotherms of Alaria esculenta and Palmaria palmata. Appl. Phycol. 2022, 3, 27–35. [Google Scholar] [CrossRef]
- Neoh, Y.Y.; Matanjun, P.; Lee, J.S. Comparative study of drying methods on chemical constituents of Malaysian red seaweed. Dry Technol. 2016, 34, 1745–1751. [Google Scholar] [CrossRef]
- Mouritsen, O.G.; Dawczynski, C.; Duelund, L.; Jahreis, G.; Vetter, W.; Schröder, M. On the human consumption of the red seaweed dulse (Palmaria palmata (L.) Weber & Mohr). J. Appl. Phycol. 2013, 25, 1777–1791. [Google Scholar] [CrossRef]
- Abdollahi, M.; Axelsson, J.; Carlsson, N.-G.; Nylund, G.M.; Albers, E.; Undeland, I. Effect of stabilization method and freeze/thaw-aided precipitation on structural and functional properties of proteins recovered from brown seaweed (Saccharina latissima). Food Hydrocoll. 2019, 96, 140–150. [Google Scholar] [CrossRef]
- Sappati, P.K.; Nayak, B.; VanWalsum, G.P.; Mulrey, O.T. Combined effects of seasonal variation and drying methods on the physicochemical properties and antioxidant activity of sugar kelp (Saccharina latissima). J. Appl. Phycol. 2018, 31, 1311–1332. [Google Scholar] [CrossRef]
- Uribe, E.; Vega-Gálvez, A.; García, V.; Pastén, A.; Rodríguez, K.; López, J.; Scala, K.D. Evaluation of physicochemical composition and bioactivity of a red seaweed (Pyropia orbicularis) as affected by different drying technologies. Dry Technol. 2019, 38, 1218–1230. [Google Scholar] [CrossRef]
- Llave, Y.; Sakai, N. Dielectric Defrosting of Frozen Foods. In Food Processing for Increased Quality and Consumption; Academic Press: Cambridge, MA, USA, 2018; pp. 383–422. [Google Scholar]
- Malik Erginkaya, Z.; Ahmad, S.; Erten, H. Food Processing: Strategies for Quality Assessment, a Broad Perspective; Food Engineering Series; Spinger: New York, NY, USA, 2014; pp. 395–426. [Google Scholar]
- Silva, C.L.; Gonçalves, E.M.; Brandao, T.R. Freezing of fruits and vegetables. In Frozen Food Science and Technology; Blackwell: Oxford, UK, 2008; pp. 165–193. [Google Scholar]
- Davison, I.R.; Dudgeon, S.R.; Ruan, H.M. Effect of freezing on seaweed photosynthesis. Mar. Ecol. Prog. Ser. 1989, 58, 123–131. [Google Scholar] [CrossRef]
- Ghaliaoui, N.; Mokrane, H.; Hazzit, M.; Hadjadj, M.; Otmani, F.S.; Touati, S.; Seridi, H. Impact of Freezing and Drying Preprocessing on Pigments Extraction from the Brown Seaweed Phyllaria Reniformis Collected in Algerian Coast. Carpathian J. Food Sci. Technol. 2020, 12, 81–94. [Google Scholar] [CrossRef]
- Lafeuille, B.; Tamigneaux, É.; Berger, K.; Provencher, V.; Beaulieu, L. Variation of the Nutritional Composition and Bioactive Potential in Edible Macroalga Saccharina latissima Cultivated from Atlantic Canada Subjected to Different Growth and Processing Conditions. Foods 2023, 12, 1736. [Google Scholar] [CrossRef] [PubMed]
- Harnedy, P.A.; FitzGerald, R.J. In vitro assessment of the cardioprotective, anti-diabetic and antioxidant potential of Palmaria palmata protein hydrolysates. J. Appl. Phycol. 2013, 25, 1793–1803. [Google Scholar] [CrossRef]
- Hell, A.; Labrie, S.; Beaulieu, L. Effect of seaweed flakes addition on the development of bioactivities in functional Camembert-type cheese. Int. J. Food Sci. Technol. 2018, 53, 1054–1064. [Google Scholar] [CrossRef] [Green Version]
- Beaulieu, L.; Bondu, S.; Doiron, K.; Rioux, L.-E.; Turgeon, S.L. Characterization of antibacterial activity from protein hydrolysates of the macroalga Saccharina longicruris and identification of peptides implied in bioactivity. J. Funct. Foods 2015, 17, 685–697. [Google Scholar] [CrossRef]
- Dudonné, S.; Vitrac, X.; Coutière, P.; Woillez, M.; Mérillon, J. Comparative study of antioxidant properties and total phenolic content of 30 plant extracts of industrial interest using DPPH, ABTS, FRAP, SOD, and ORAC assays. J. Agric. Food Chem. 2009, 57, 1768–1774. [Google Scholar] [CrossRef]
- Müller, L.; Fröhlich, K.; Böhm, V. Comparative antioxidant activities of carotenoids measured by ferric reducing antioxidant power (FRAP), ABTS bleaching assay (αTEAC), DPPH assay and peroxyl radical scavenging assay. Food Chem. 2011, 129, 139–148. [Google Scholar] [CrossRef]
- Boisvert, C.; Beaulieu, L.; Bonnet, C.; Pelletier, É. Assessment of the Antioxidant and Antibacterial Activities of Three Species of Edible Seaweeds. J. Food Biochem. 2015, 39, 377–387. [Google Scholar] [CrossRef]
- Tamigneaux, E.; Vazquez, E.; Mingelbier, M.; Klein, B.; Legendre, L. Environmental control of phytoplankton assemblages in nearshore marine waters, with special emphasis on phototrophic ultraplankton. J. Plankton Res. 1995, 17, 1421–1448. [Google Scholar] [CrossRef]
- Perfeto, P.N.M. Relation between chemical composition of Grateloupia doryphore (Montagne) Howe, Gymnogongrus griffithsiae (Turner) Martius, and abiotic parameters. Acta Bot. Bras. 1998, 12, 77–88. [Google Scholar] [CrossRef] [Green Version]
- Lafeuille, B.; Francezon, N.; Goulet, C.; Perreault, V.; Turgeon, S.L.; Beaulieu, L. Impact of temperature and cooking time on the physicochemical properties and sensory potential of seaweed water extracts of Palmaria palmata and Saccharina longicruris. J. Appl. Phycol. 2022, 34, 1731–1747. [Google Scholar] [CrossRef]
- Mabeau, S.; Fleurence, J. Seaweed in food products: Biochemical and nutritional aspects. Trends Food Sci. Technol. 1993, 4, 103–107. [Google Scholar] [CrossRef]
- Bourgougnon, N.; Gervois, A. Les Algues Marines: Biologie, Écologie et Utilisation; Ellipses: Paris, France, 2021; pp. 13–20. [Google Scholar]
- Kikunaga, S.; Ishii, H.; Takahashi, M. The bioavailability of magnesium in spinach and the effect of oxalic acid on magnesium utilization examined in diets of magnesium-deficient rats. J. Nutr. Sci. Vitaminol. 1995, 41, 671–685. [Google Scholar] [CrossRef]
- Roberts, J.L.; Moreau, R. Functional properties of spinach (Spinacia oleracea L.) phytochemicals and bioactives. Food Funct. 2016, 7, 3337–3353. [Google Scholar] [CrossRef] [PubMed]
- Gouvernement du Canada. Fichier Canadien sur les Éléments Nutritifs, Spinach, Raw, Food Code: 2213. Available online: https://aliments-nutrition.canada.ca/cnf-fce/report-rapport (accessed on 15 May 2023).
- Sager, M. Chocolate and Cocoa Products as A Source of Essential Elements in Nutrition. J. Nutr. Food Sci. 2012, 2, 123. [Google Scholar] [CrossRef] [Green Version]
- Gouvernement du Canada. Fichier Canadien sur les Éléments Nutritifs, Candies, Chocolate, Dark, 70–85% Cacao Solids, Food Code: 6672. Available online: https://aliments-nutrition.canada.ca/cnf-fce/report-rapport (accessed on 15 May 2023).
- Venturi, S. Evolutionary Significance of Iodine. Curr. Chem. Biol. 2011, 5, 155–162. [Google Scholar]
- Ranjha, M.M.A.N.; Irfan, S.; Nadeem, M.; Mahmood, S. A Comprehensive Review on Nutritional Value, Medicinal Uses, and Processing of Banana. Food Rev. Int. 2020, 38, 199–225. [Google Scholar] [CrossRef]
- Gouvernement du Canada. Fichier Canadien sur les Éléments Nutritifs, Banana, Raw, Food Code: 1704. Available online: https://aliments-nutrition.canada.ca/cnf-fce/report-rapport (accessed on 15 May 2023).
- Kapoor, A.; Kanwar, P.; Gupta, R. Traditional recipes of district Kangra of Himachal Pradesh. Indian J. Tradit. Knowl. 2010, 9, 282–288. [Google Scholar]
- Ajmera, P.; Kalani, S.; Sharma, L. Parsley-benefits & side effects on health. Int. J. Physiologt. Nutr. Phys. Educ. 2019, 4, 1236–1242. [Google Scholar]
- Gouvernement du Canada. Fichier Canadien sur les Éléments Nutritifs, Parsley, Fresh, Food Code: 2405. Available online: https://aliments-nutrition.canada.ca/cnf-fce/report-rapport (accessed on 15 May 2023).
- Beer, S.; Björk, M.; Beardall, J.; Bjork, M. Photosynthesis in the Marine Environment; John Wiley & Sons: Hoboken, NJ, USA, 2014. [Google Scholar]
- Luta, X.; Hayoz, S.; Grea Krause, C.; Sommerhalder, K.; Roos, E.; Strazzullo, P.; Beer-Borst, S. The relationship of health/food literacy and salt awareness to daily sodium and potassium intake among a workplace population in Switzerland. Nutr. Metab. Cardiovasc. Dis. 2018, 28, 270–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gouvernement du Canada. Fichier Canadien sur les Éléments Nutritifs, Crustaceans, Shrimp, Mixed Species, Raw, Food Code: 3211. Available online: https://aliments-nutrition.canada.ca/cnf-fce/report-rapport (accessed on 15 May 2023).
- Tamigneaux, É.; Pedneault, E.; Gendron, L. Comparaison des Rendements de L’algue Brune Saccharina Longicruris Cultivée en Milieu Ouvert en Gaspésie et en Lagune Aux Îles-de-la-Madeleine; Merinov: Grande-Rivère, QC, Canada, 2014; p. 35. [Google Scholar]
- Maehre, H.K.; Edvinsen, G.K.; Eilertsen, K.-E.; Elvevoll, E.O. Heat treatment increases the protein bioaccessibility in the red seaweed dulse (Palmaria palmata), but not in the brown seaweed winged kelp (Alaria esculenta). J. Appl. Phycol. 2016, 28, 581–590. [Google Scholar] [CrossRef] [Green Version]
- Gross, G. Geology of Iron Deposits in Canada. Volume II Iron deposits in the Appalachian and Greenville Region in Canada; Geological Survey of Canada, Department of Energy, Mines and Ressources: Ottawa, ON, Canada, 1967. [Google Scholar]
- Watzke, H.J. Impact of processing on bioavailability examples of minerals in foods. Trends Food Sci. Technol. 1998, 9, 320–327. [Google Scholar] [CrossRef]
- Kumar, S.A.; Brown, L. Seaweeds as potential therapeutic interventions for the metabolic syndrome. Rev. Endocr. Metab. Disord. 2013, 14, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-X.; Kim, S.-K. Utilization of seaweed derived ingredients as potential antioxidants and functional ingredients in the food industry: An overview. Food Sci. Biotechnol. 2011, 20, 1461–1466. [Google Scholar] [CrossRef]
- Sampath-Wiley, P.; Neefus, C.D.; Jahnke, L.S. Seasonal effects of sun exposure and emersion on intertidal seaweed physiology: Fluctuations in antioxidant contents, photosynthetic pigments and photosynthetic efficiency in the red alga Porphyra umbilicalis Kützing (Rhodophyta, Bangiales). J. Exp. Mar. Biol. Ecol. 2008, 361, 83–91. [Google Scholar] [CrossRef]
- Yuan, Y.V.; Bone, D.E.; Carrington, M.F. Antioxidant activity of dulse (Palmaria palmata) extract evaluated in vitro. Food Chem. 2005, 91, 485–494. [Google Scholar] [CrossRef]
- Vilcanqui, Y.; Mamani-Apaza, L.O.; Flores, M.; Ortiz-Viedma, J.; Romero, N.; Mariotti-Celis, M.S.; Huamán-Castilla, N.L. Chemical Characterization of Brown and Red Seaweed from Southern Peru, a Sustainable Source of Bioactive and Nutraceutical Compounds. Agronomy 2021, 11, 1669. [Google Scholar] [CrossRef]
- Naseri, A.; Holdt, S.L.; Jacobsen, C. Biochemical and Nutritional Composition of Industrial Red Seaweed Used in Carrageenan Production. J. Aquat. Food Prod. Technol. 2019, 28, 967–973. [Google Scholar] [CrossRef]
- Syamila, M.; Gedi, M.A.; Briars, R.; Ayed, C.; Gray, D.A. Effect of temperature, oxygen and light on the degradation of beta-carotene, lutein and alpha-tocopherol in spray-dried spinach juice powder during storage. Food Chem. 2019, 284, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Afonso, C.; Bandarra, N.M.; Nunes, L.; Cardoso, C. Tocopherols in Seafood and Aquaculture Products. Crit. Rev. Food Sci. Nutr. 2016, 56, 128–140. [Google Scholar] [CrossRef]
- Gouvernement du Canada. Fichier Canadien sur les Éléments Nutritifs, Fish, Salmon, Atlantic, Farmed, Raw, Food Code: 3182. Available online: https://aliments-nutrition.canada.ca/cnf-fce/report-rapport (accessed on 15 May 2023).
- USDA; ARS. Portions and Weight. Available online: https://www.ars.usda.gov/northeast-area/beltsville-md-bhnrc/beltsville-human-nutrition-research-center/food-surveys-research-group/docs/fndds-download-databases/ (accessed on 15 May 2023).
- Machu, L.; Misurcova, L.; Ambrozova, J.V.; Orsavova, J.; Mlcek, J.; Sochor, J.; Jurikova, T. Phenolic content and antioxidant capacity in algal food products. Molecules 2015, 20, 1118–1133. [Google Scholar] [CrossRef] [Green Version]
- Fleurence, J. Seaweed proteins: Biochemical, nutritional aspects and potential uses. Trends Food Sci. Technol. 1999, 10, 25–28. [Google Scholar]
- Jimenez-Escrig, A.; Gomez-Ordonez, E.; Ruperez, P. Seaweed as a source of novel nutraceuticals: Sulfated polysaccharides and peptides. Adv. Food Nutr. Res. 2011, 64, 325–337. [Google Scholar] [CrossRef] [PubMed]
- Manríquez-Hernández, J.A.; Duston, J.; Garbary, D.J. Effect of irradiance on bioremediation capacity of the red algae Chondrus crispus and Palmaria palmata. Aquac. Int. 2015, 24, 39–55. [Google Scholar] [CrossRef]
- Stack, J.; Tobin, P.R.; Gietl, A.; Harnedy, P.A.; Stengel, D.B.; FitzGerald, R.J. Seasonal variation in nitrogenous components and bioactivity of protein hydrolysates from Porphyra dioica. J. Appl. Phycol. 2017, 29, 2439–2450. [Google Scholar] [CrossRef]
- Furuta, T.; Miyabe, Y.; Yasui, H.; Kinoshita, Y.; Kishimura, H. Angiotensin I Converting Enzyme Inhibitory Peptides Derived from Phycobiliproteins of Dulse Palmaria palmata. Mar. Drugs 2016, 14, 32. [Google Scholar] [CrossRef] [Green Version]
- Angell, A.R.; Mata, L.; de Nys, R.; Paul, N.A. The protein content of seaweeds: A universal nitrogen-to-protein conversion factor of five. J. Appl. Phycol. 2015, 28, 511–524. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, K.; Kumar, K.S.; Rao, P.V.S. Comparative assessment of antioxidant activity in three edible species of green seaweed, Enteromorpha from Okha, Northwest coast of India. Innov. Food Sci. Emerg. Technol. 2011, 12, 73–78. [Google Scholar] [CrossRef]
- Singleton, V.L.R.; Joseph, A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Abdelhady, M.I.S.; Motaal, A.A.; Beerhues, L. Total Phenolic Content and Antioxidant Activity of Standardized Extracts from Leaves and Cell Cultures of Three Callistemon Species. Am. J. Plant Sci. 2011, 2, 847–850. [Google Scholar] [CrossRef] [Green Version]
- Quan, C.; Turner, C. Extraction of Astaxanthin from Shrimp Waste Using Pressurized Hot Ethanol. Chromatographia 2009, 70, 247–251. [Google Scholar] [CrossRef]
- Sanchez-Machado, D.I.; López-Hernández, J.; Paseiro-Losada, P. High-performance liquid chromatographic determination of a -tocopherol in macroalgae. J. Chromatogr. 2002, 976, 277–284. [Google Scholar] [CrossRef]
- Simonne, A.H.; Simonne, E.H.; Eitenmiller, R.R.; Mills, H.A.; Cresman, C.P. Could the Dumas Method replace the Kjeldahl digestion for nitrogen and crude protein determinations in foods? J. Sci. Food Agric. 1997, 73, 39–45. [Google Scholar] [CrossRef]
- Hayakari, M.; Kondo, Y.; Izumi, H. A Rapid and Simple Spectrophotometric Angiotensin-Converting Enzyme Assay. Anal. Biochem. 1978, 84, 361–369. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| P. palmata | Lipids (%) | Proteins (%) | Ash (%) | Carbohydrates (%) | Fiber (%) |
|---|---|---|---|---|---|
| Crudes—October | 1.17 ± 0.18 d | 11.23 ± 0.17 c | 12.30 ± 0.04 c | 75.30 ± 0.20 a | 35.81 ± 0.27 c |
| Dried—October | 1.65 ± 0.10 c | 11.19 ± 0.17 c | 12.22 ± 0.09 c | 74.94 ± 0.13 ab | 36.54 ± 0.18 b |
| Crudes—June | 4.64 ± 0.19 a | 16.19 ± 0.09 b | 27.26 ± 0.29 a | 51.91 ± 0.52 bc | 36.88 ± 0.10 b |
| Dried—June | 3.65 ± 0.21 b | 17.32 ± 0.17 a | 26.18 ± 0.05 b | 52.85 ± 0.43 ab | 37.80 ± 0.22 a |
| P. palmata | I (mg/100 g) | K (g/100 g) | Na (g/100 g) | Ca (mg/100 g) | Mg (mg/100 g) | Fe (mg/100 g) | Na/K Ratio |
|---|---|---|---|---|---|---|---|
| Crudes—October | 3.00 ± 0.44 bc | 3.43 ± 0.06 bc | 1.30 ± 0.00 c | 120.00 ± 13.00 c | 169.00 ± 4.00 c | 30.00 ± 1.00 c | 0.38 |
| Dried—October | 4.10 ± 0.10 ab | 3.53 ± 0.12 ab | 1.27 ± 0.06 c | 130.00 ± 13.00 c | 168.00 ± 5.00 c | 28.67 ± 0.56 c | 0.36 |
| Crudes—June | 9.60 ± 1.45 ab | 9.23 ± 0.38 ab | 2.58 ± 0.06 a | 317.00 ± 21.00 a | 313.00 ± 9.00 a | 436.67 ± 35.00 a | 0.28 |
| Dried—June | 10.73 ± 2.63 a | 12.65 ± 1.73 a | 2.20 ± 0.04 b | 256.00 ± 2.00 b | 261.00 ± 4.00 b | 352.33 ± 4.16 b | 0.17 |
| P. palmata | Carotenoids (μg/g) | Polyphenols (mg GAE/g) | α-Tocopherol (μg/g) | β-Carotene (μg/g) |
|---|---|---|---|---|
| Crudes—October | 371.00 ± 11.00 b | 1.10 ± 0.00 c | 9.95 ± 1.11 c | 12.87 ± 3.09 c |
| Dried—October | 308.00 ± 3.00 c | 0.80 ± 0.14 c | 9.05 ± 0.63 c | 7.73 ± 0.77 c |
| Crudes—June | 315.00 ± 22.00 c | 2.06 ± 0.05 b | 22.67 ± 2.08 b | 33.33 ± 1.53 b |
| Dried—June | 535.00 ± 24.00 a | 1.89 ± 0.17 a | 50.00 ± 5.29 a | 47.67 ± 2.08 a |
| P. palmata | Crudes—October | Dried—October | Crudes—June | Dried—June |
|---|---|---|---|---|
| Protein (%) | 15.70 ± 2.31 a | 23.00 ± 0.89 ab | 29.03 ± 4.30 bc | 22.32 ± 2.34 ab |
| Extraction yield (%) | 8.40 ± 0.24 a | 3.44 ± 1.25 bc | 5.60 ± 0.47 ab | 5.38 ± 0.20 ab |
| Protein extraction yield (%) | 12.64 ± 1.57 a | 10.37 ± 1.28 a | 7.45 ± 1.16 a | 6.92 ± 0.62 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lafeuille, B.; Tamigneaux, É.; Berger, K.; Provencher, V.; Beaulieu, L. Impact of Harvest Month and Drying Process on the Nutritional and Bioactive Properties of Wild Palmaria palmata from Atlantic Canada. Mar. Drugs 2023, 21, 392. https://doi.org/10.3390/md21070392
Lafeuille B, Tamigneaux É, Berger K, Provencher V, Beaulieu L. Impact of Harvest Month and Drying Process on the Nutritional and Bioactive Properties of Wild Palmaria palmata from Atlantic Canada. Marine Drugs. 2023; 21(7):392. https://doi.org/10.3390/md21070392
Chicago/Turabian StyleLafeuille, Bétina, Éric Tamigneaux, Karine Berger, Véronique Provencher, and Lucie Beaulieu. 2023. "Impact of Harvest Month and Drying Process on the Nutritional and Bioactive Properties of Wild Palmaria palmata from Atlantic Canada" Marine Drugs 21, no. 7: 392. https://doi.org/10.3390/md21070392
APA StyleLafeuille, B., Tamigneaux, É., Berger, K., Provencher, V., & Beaulieu, L. (2023). Impact of Harvest Month and Drying Process on the Nutritional and Bioactive Properties of Wild Palmaria palmata from Atlantic Canada. Marine Drugs, 21(7), 392. https://doi.org/10.3390/md21070392