
Growth Behavior, Biomass Composition and Fatty Acid Methyl Esters (FAMEs) Production Potential of Chlamydomonas reinhardtii, and Chlorella vulgaris Cultures
 ,
,  , , ,
, , , 
 and
and 
Abstract
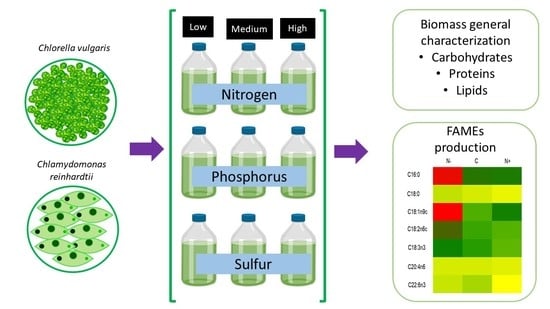
:
1. Introduction
2. Results and Discussion
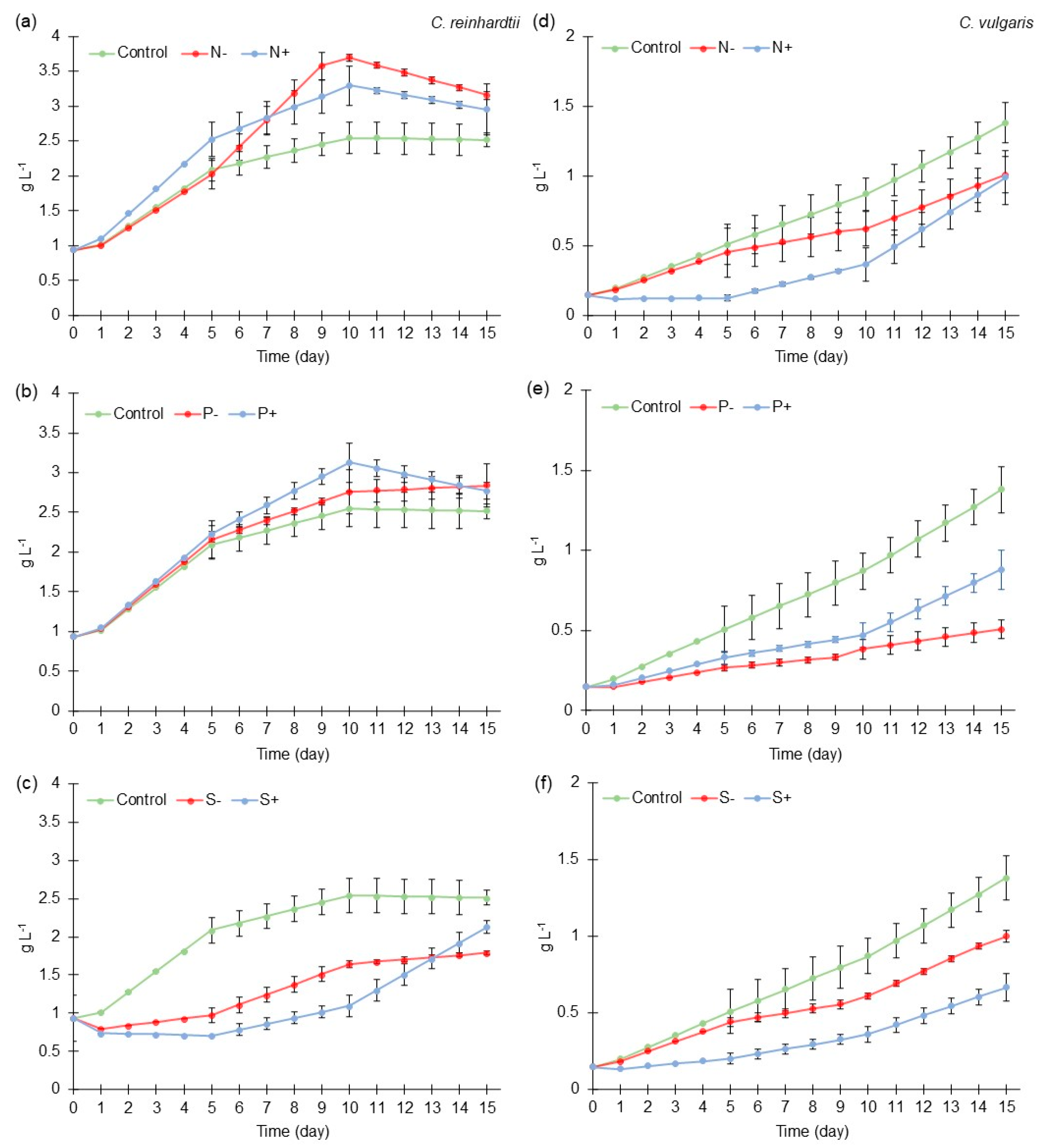
2.1. Growth Behavior of Chlamydomonas reinhardtii and Chlorella vulgaris with Different Nitrogen, Phosphorus, and Sulfur Concentrations
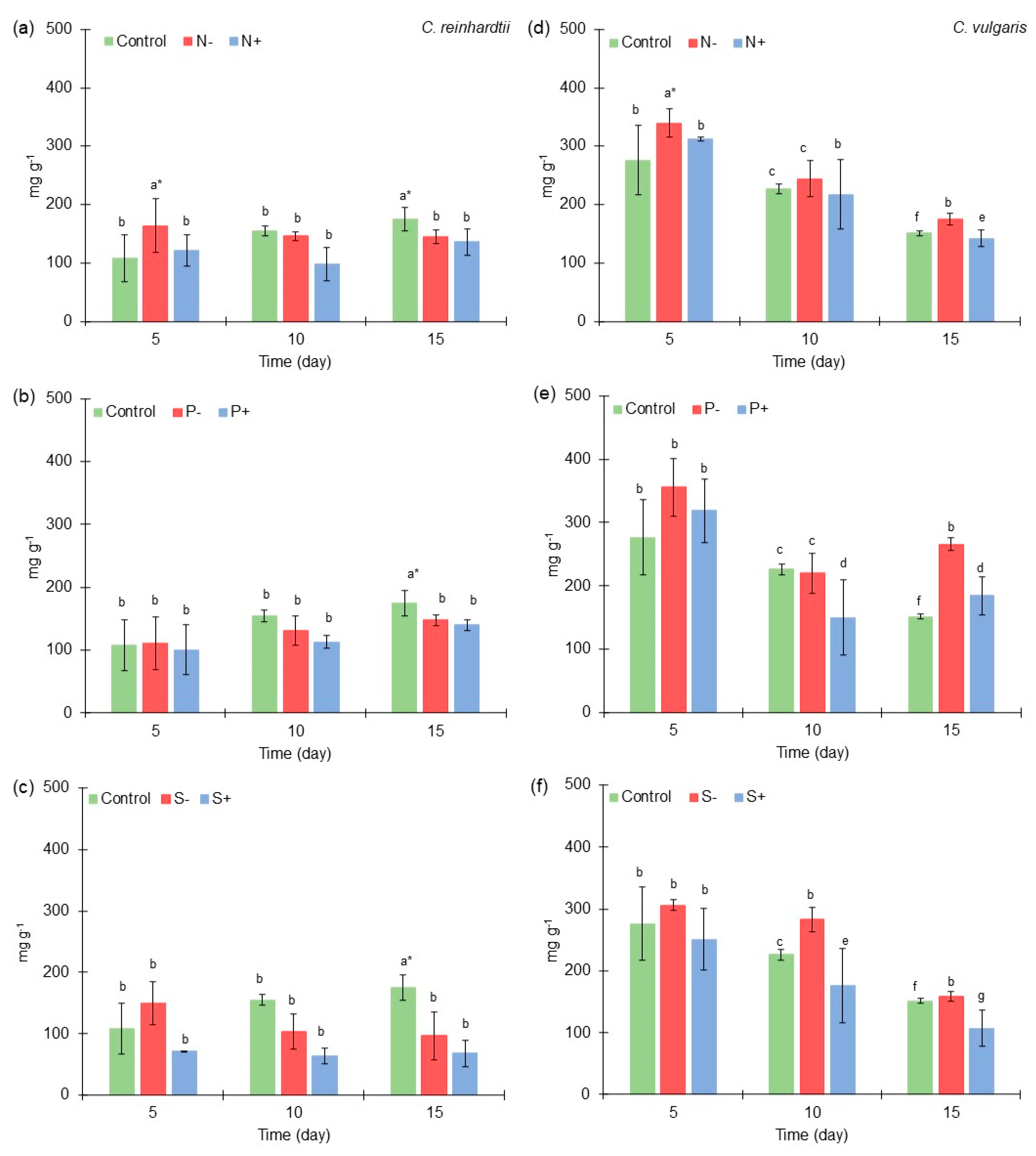
2.2. Carbohydrates Content of Chlamydomonas reinhardtii and Chlorella vulgaris Grown with Different Nitrogen, Phosphorus, and Sulfur Concentrations
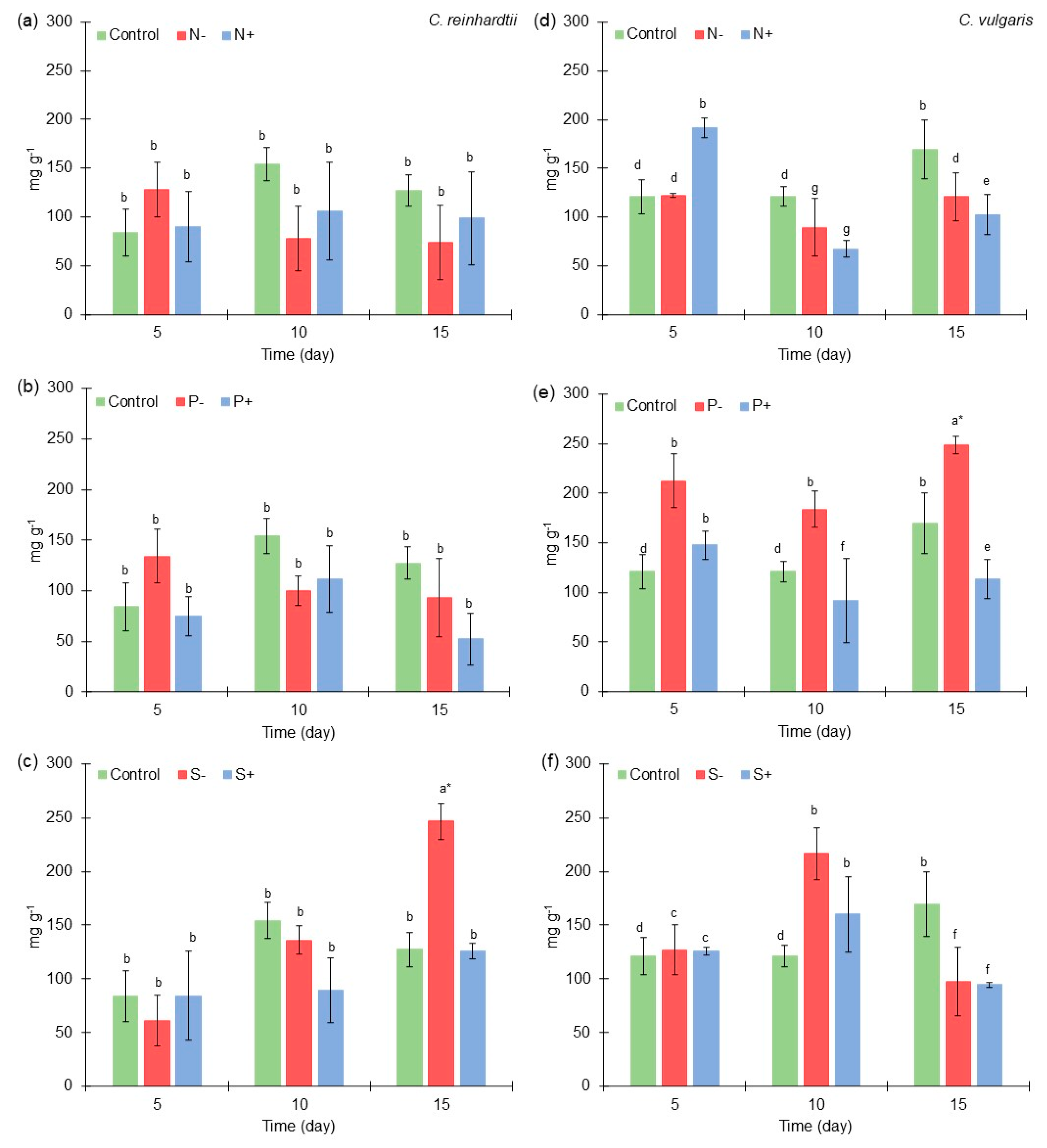
2.3. Protein Content of Chlamydomonas reinhardtii and Chlorella vulgaris Grown with Different Nitrogen, Phosphorus, and Sulfur Concentrations
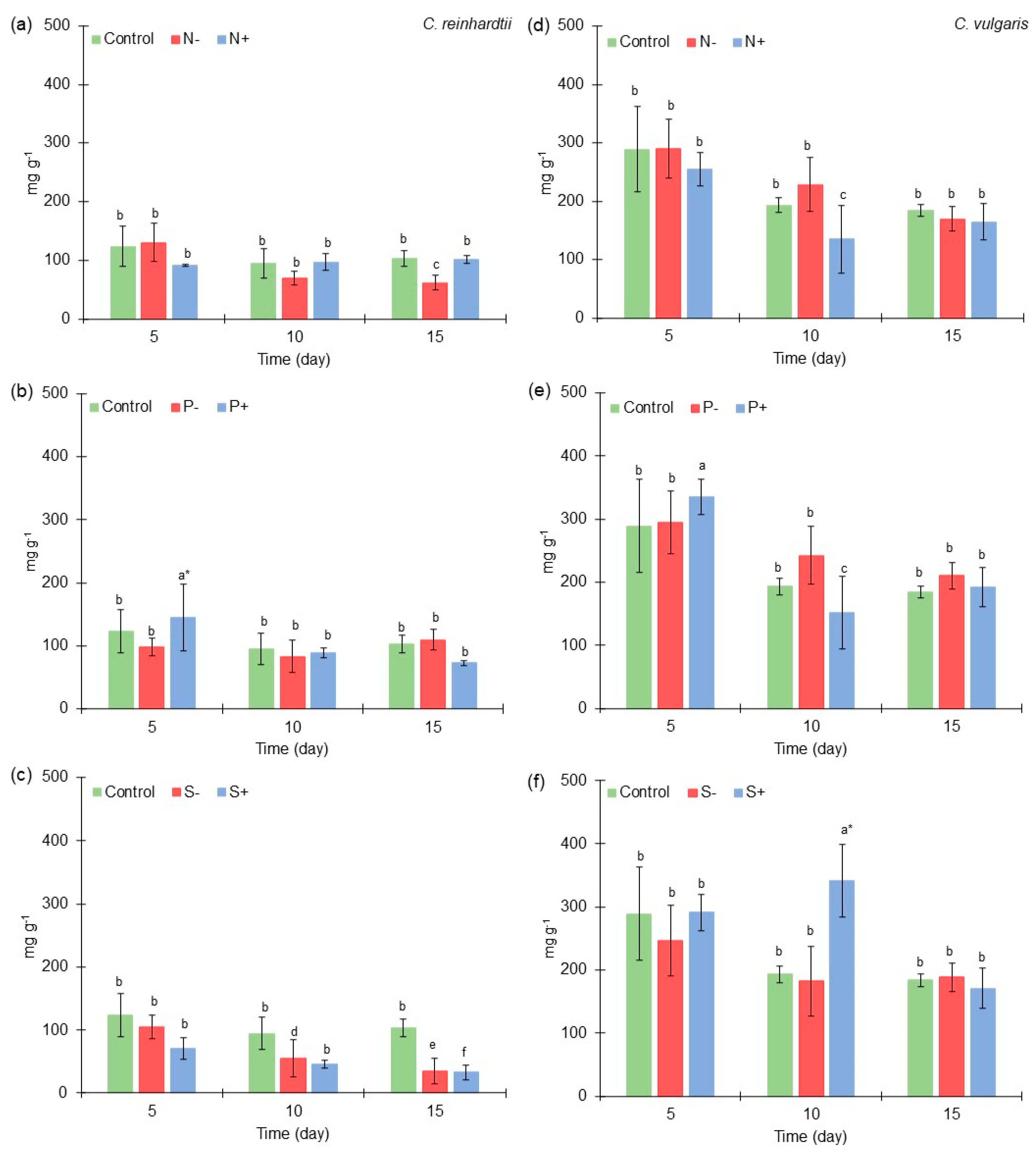
2.4. Lipid Content of Chlamydomonas reinhardtii and Chlorella vulgaris Grown with Different Nitrogen, Phosphorus, and Sulfur Concentrations
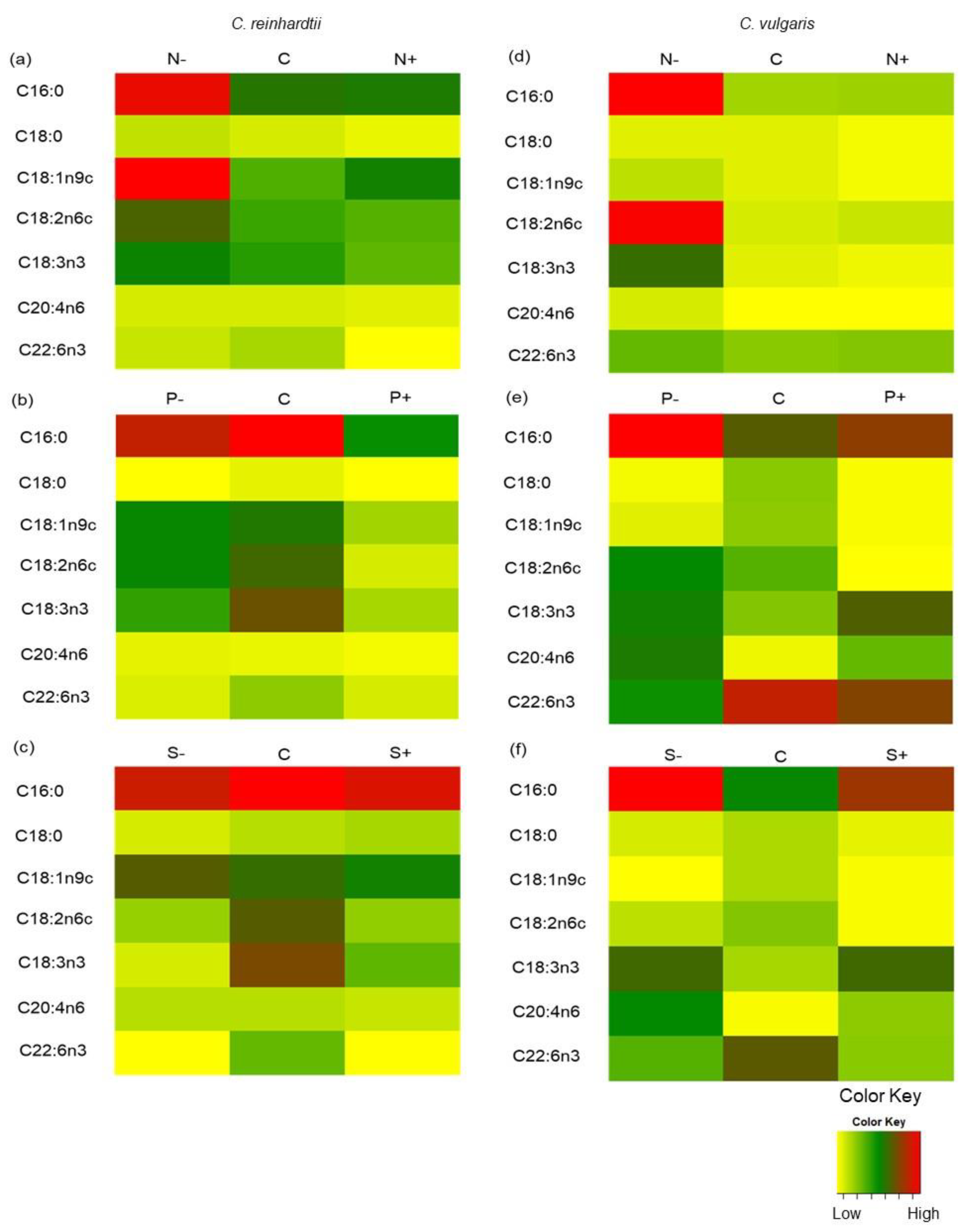
2.5. FAMEs Accumulation of Chlamydomonas reinhardtii and Chlorella vulgaris Grown with Different Nitrogen, Phosphorus, and Sulfur Concentrations
3. Materials and Methods
3.1. Reagents and Equipment
3.2. Inoculum Culture Conditions
3.3. Growth Measure of Chlamydomonas reinhardtii and Chlorella vulgaris with Different Nitrogen, Phosphorus, and Sulfur Concentrations
3.4. Biomass Characterization of Chlamydomonas reinhardtii and Chlorella vulgaris Grown with Different Nitrogen, Phosphorus, and Sulfur Concentrations
3.5. Total Carbohydrates Content of Chlamydomonas reinhardtii and Chlorella vulgaris Grown with Different Nitrogen, Phosphorus, and Sulfur Concentrations
3.6. Total Protein Content of Chlamydomonas reinhardtii and Chlorella vulgaris Grown with Different Nitrogen, Phosphorus, and Sulfur Concentrations
3.7. Total Lipid Content of Chlamydomonas reinhardtii and Chlorella vulgaris Grown with Different Nitrogen, Phosphorus, and Sulfur Concentrations
3.8. FAMEs of Chlamydomonas reinhardtii and Chlorella vulgaris Grown with Different Nitrogen, Phosphorus, and Sulfur Concentrations
3.9. Data Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khan, M.I.; Shin, J.H.; Kim, J.D. The promising future of microalgae: Current status, challenges, and optimization of a sustainable and renewable industry for biofuels, feed, and other products. Microb. Cell Fact. 2018, 17, 36. [Google Scholar] [CrossRef] [PubMed]
- Hossain, S.M.Z. Biochemical Conversion of Microalgae Biomass into Biofuel. Chem. Eng. Technol. 2019, 42, 2594–2607. [Google Scholar] [CrossRef]
- López-Pacheco, I.Y.; Silva-Núñez, A.; García-Perez, J.S.; Carrillo-Nieves, D.; Salinas-Salazar, C.; Castillo-Zacarías, C.; Afewerki, S.; Barceló, D.; Iqbal, H.N.M.; Parra-Saldívar, R. Phyco-remediation of swine wastewater as a sustainable model based on circular economy. J. Environ. Manag. 2021, 278, 111534. [Google Scholar] [CrossRef] [PubMed]
- Adamczyk, M.; Lasek, J.; Skawińska, A. CO2 Biofixation and Growth Kinetics of Chlorella vulgaris and Nannochloropsis gaditana. Appl. Biochem. Biotechnol. 2016, 179, 1248–1261. [Google Scholar] [CrossRef]
- Dineshbabu, G.; Uma, V.S.; Mathimani, T.; Deviram, G.; Arul Ananth, D.; Prabaharan, D.; Uma, L. On-site concurrent carbon dioxide sequestration from flue gas and calcite formation in ossein effluent by a marine cyanobacterium Phormidium valderianum BDU 20041. Energy Convers. Manag. 2017, 141, 315–324. [Google Scholar] [CrossRef]
- Hariz, H.B.; Takriff, M.S.; Mohd Yasin, N.H.; Ba-Abbad, M.M.; Mohd Hakimi, N.I.N. Potential of the microalgae-based integrated wastewater treatment and CO2 fixation system to treat Palm Oil Mill Effluent (POME) by indigenous microalgae; Scenedesmus sp. and Chlorella sp. J. Water Process Eng. 2019, 32, 100907. [Google Scholar] [CrossRef]
- Srimongkol, P.; Sangtanoo, P.; Songserm, P.; Watsuntorn, W.; Karnchanatat, A. Microalgae-based wastewater treatment for developing economic and environmental sustainability: Current status and future prospects. Front. Bioeng. Biotechnol. 2022, 10, 1048819. [Google Scholar] [CrossRef]
- Kuo, C.M.; Chen, T.Y.; Lin, T.H.; Kao, C.Y.; Lai, J.T.; Chang, J.S.; Lin, C.S. Cultivation of Chlorella sp. GD using piggery wastewater for biomass and lipid production. Bioresour. Technol. 2015, 194, 326–333. [Google Scholar] [CrossRef]
- Ahmad, A.; Hassan, S.; Banat, F. An overview of microalgae biomass as a sustainable aquaculture feed ingredient: Food security and circular economy. Bioengineered 2022, 13, 9521–9547. [Google Scholar] [CrossRef]
- Gouda, M.; Tadda, M.A.; Zhao, Y.; Farmanullah, F.; Chu, B.; Li, X.; He, Y. Microalgae Bioactive Carbohydrates as a Novel Sustainable and Eco-Friendly Source of Prebiotics: Emerging Health Functionality and Recent Technologies for Extraction and Detection. Front. Nutr. 2022, 9, 806692. [Google Scholar] [CrossRef]
- Sui, Y.; Vlaeminck, S.E. Effects of salinity, pH and growth phase on the protein productivity by Dunaliella salina. J. Chem. Technol. Biotechnol. 2019, 94, 1032–1040. [Google Scholar] [CrossRef]
- Baldisserotto, C.; Sabia, A.; Giovanardi, M.; Ferroni, L.; Maglie, M.; Pancaldi, S. Chlorophyta microalgae as dietary protein supplement: A comparative analysis of productivity related to photosynthesis. J. Appl. Phycol. 2022, 34, 1323–1340. [Google Scholar] [CrossRef]
- Udayan, A.; Pandey, A.K.; Sirohi, R.; Sreekumar, N.; Sang, B.-I.; Sim, S.J.; Kim, S.H.; Pandey, A. Production of microalgae with high lipid content and their potential as sources of nutraceuticals. Phytochem. Rev. 2022. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, G.F.; Ríos Pinto, L.F.; Maciel Filho, R.; Fregolente, L.V. A review on lipid production from microalgae: Association between cultivation using waste streams and fatty acid profiles. Renew. Sustain. Energy Rev. 2019, 109, 448–466. [Google Scholar] [CrossRef]
- Deshmukh, S.; Kumar, R.; Bala, K. Microalgae biodiesel: A review on oil extraction, fatty acid composition, properties and effect on engine performance and emissions. Fuel Process. Technol. 2019, 191, 232–247. [Google Scholar] [CrossRef]
- Bayly, G.R. Lipids and disorders of lipoprotein metabolism. In Clinical Biochemistry: Metabolic and Clinical Aspects; Elsevier: Amsterdam, The Netherlands, 2014; pp. 702–736. [Google Scholar]
- Maltsev, Y.; Maltseva, K. Fatty acids of microalgae: Diversity and applications. Rev. Environ. Sci. Bio/Technol. 2021, 20, 515–547. [Google Scholar] [CrossRef]
- Babu, S.S.; Gondi, R.; Vincent, G.S.; JohnSamuel, G.C.; Jeyakumar, R.B. Microalgae Biomass and Lipids as Feedstock for Biofuels: Sustainable Biotechnology Strategies. Sustainability 2022, 14, 15070. [Google Scholar] [CrossRef]
- Akubude, V.C.; Nwaigwe, K.N.; Dintwa, E. Production of biodiesel from microalgae via nanocatalyzed transesterification process: A review. Mater. Sci. Energy Technol. 2019, 2, 216–225. [Google Scholar] [CrossRef]
- Farfan-Cabrera, L.I.; Franco-Morgado, M.; González-Sánchez, A.; Pérez-González, J.; Marín-Santibáñez, B.M. Microalgae Biomass as a New Potential Source of Sustainable Green Lubricants. Molecules 2022, 27, 1205. [Google Scholar] [CrossRef]
- Moreira, J.B.; Vaz, B.d.S.; Cardias, B.B.; Cruz, C.G.; Almeida, A.C.A.d.; Costa, J.A.V.; Morais, M.G.d. Microalgae Polysaccharides: An Alternative Source for Food Production and Sustainable Agriculture. Polysaccharides 2022, 3, 441–457. [Google Scholar] [CrossRef]
- Lopez-Pacheco, I.Y.; Rodas-Zuluaga, L.I.; Cuellar-Bermudez, S.P.; Hidalgo-Vázquez, E.; Molina-Vazquez, A.; Araújo, R.G.; Martínez-Ruiz, M.; Varjani, S.; Barceló, D.; Iqbal, H.M.N.; et al. Revalorization of Microalgae Biomass for Synergistic Interaction and Sustainable Applications: Bioplastic Generation. Mar. Drugs 2022, 20, 601. [Google Scholar] [CrossRef]
- De Carvalho Silvello, M.A.; Severo Gonçalves, I.; Patrícia Held Azambuja, S.; Silva Costa, S.; Garcia Pereira Silva, P.; Oliveira Santos, L.; Goldbeck, R. Microalgae-based carbohydrates: A green innovative source of bioenergy. Bioresour. Technol. 2022, 344, 126304. [Google Scholar] [CrossRef] [PubMed]
- Amorim, M.L.; Soares, J.; Coimbra, J.S.D.R.; Leite, M.D.O.; Albino, L.F.T.; Martins, M.A. Microalgae proteins: Production, separation, isolation, quantification, and application in food and feed. Crit. Rev. Food Sci. Nutr. 2021, 61, 1976–2002. [Google Scholar] [CrossRef] [PubMed]
- Bertsch, P.; Böcker, L.; Mathys, A.; Fischer, P. Proteins from microalgae for the stabilization of fluid interfaces, emulsions, and foams. Trends Food Sci. Technol. 2021, 108, 326–342. [Google Scholar] [CrossRef]
- Morales, M.; Aflalo, C.; Bernard, O. Microalgal lipids: A review of lipids potential and quantification for 95 phytoplankton species. Biomass Bioenergy 2021, 150, 106108. [Google Scholar] [CrossRef]
- De Luca, M.; Pappalardo, I.; Limongi, A.R.; Viviano, E.; Radice, R.P.; Todisco, S.; Martelli, G.; Infantino, V.; Vassallo, A. Lipids from Microalgae for Cosmetic Applications. Cosmetics 2021, 8, 52. [Google Scholar] [CrossRef]
- Farfan-Cabrera, L.I.; Rojo-Valerio, A.; Calderon-Najera, J.D.D.; Coronado-Apodaca, K.G.; Iqbal, H.M.N.; Parra-Saldivar, R.; Franco-Morgado, M.; Elias-Zuñiga, A. Microalgae Oil-Based Metal Working Fluids for Sustainable Minimum Quantity Lubrication (MQL) Operations—A Perspective. Lubricants 2023, 11, 215. [Google Scholar] [CrossRef]
- Wang, Y.; Tibbetts, S.; McGinn, P. Microalgae as Sources of High-Quality Protein for Human Food and Protein Supplements. Foods 2021, 10, 3002. [Google Scholar] [CrossRef]
- Cheng, C.-L.; Lo, Y.-C.; Huang, K.-L.; Nagarajan, D.; Chen, C.-Y.; Lee, D.-J.; Chang, J.-S. Effect of pH on biomass production and carbohydrate accumulation of Chlorella vulgaris JSC-6 under autotrophic, mixotrophic, and photoheterotrophic cultivation. Bioresour. Technol. 2022, 351, 127021. [Google Scholar] [CrossRef]
- Cheirsilp, B.; Torpee, S. Enhanced growth and lipid production of microalgae under mixotrophic culture condition: Effect of light intensity, glucose concentration and fed-batch cultivation. Bioresour. Technol. 2012, 110, 510–516. [Google Scholar] [CrossRef]
- Salman, J.M.; Grmasha, R.A.; Stenger-Kovács, C.; Lengyel, E.; Al-sareji, O.J.; AL-Cheban, A.M.A.A.; Meiczinger, M. Influence of magnesium concentrations on the biomass and biochemical variations in the freshwater algae, Chlorella vulgaris. Heliyon 2023, 9, e13072. [Google Scholar] [CrossRef] [PubMed]
- Beigbeder, J.-B.; Lavoie, J.-M. Effect of photoperiods and CO2 concentrations on the cultivation of carbohydrate-rich P. kessleri microalgae for the sustainable production of bioethanol. J. CO2 Util. 2022, 58, 101934. [Google Scholar] [CrossRef]
- Morales-Sánchez, D.; Schulze, P.S.C.; Kiron, V.; Wijffels, R.H. Production of carbohydrates, lipids and polyunsaturated fatty acids (PUFA) by the polar marine microalga Chlamydomonas malina RCC2488. Algal Res. 2020, 50, 102016. [Google Scholar] [CrossRef]
- Josephine, A.; Kumar, T.S.; Surendran, B.; Rajakumar, S.; Kirubagaran, R.; Dharani, G. Evaluating the effect of various environmental factors on the growth of the marine microalgae, Chlorella vulgaris. Front. Mar. Sci. 2022, 9, 954622. [Google Scholar] [CrossRef]
- Zarrinmehr, M.J.; Farhadian, O.; Heyrati, F.P.; Keramat, J.; Koutra, E.; Kornaros, M.; Daneshvar, E. Effect of nitrogen concentration on the growth rate and biochemical composition of the microalga, Isochrysis galbana. Egypt. J. Aquat. Res. 2020, 46, 153–158. [Google Scholar] [CrossRef]
- Chin, G.J.W.L.; Andrew, A.R.; Abdul-Sani, E.R.; Yong, W.T.L.; Misson, M.; Anton, A. The effects of light intensity and nitrogen concentration to enhance lipid production in four tropical microalgae. Biocatal. Agric. Biotechnol. 2023, 48, 102660. [Google Scholar] [CrossRef]
- Lovio Fragoso, J.; Hayano Kanashiro, C.; Lopez Elias, J. Effect of different phosphorus concentrations on growth and biochemical composition of Chaetoceros muelleri. Lat. Am. J. Aquat. Res. 2019, 47, 361–366. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Li, Y.; Zhang, Y.; Xu, Q.; Iqbal, M.S.; Xi, Y.; Xiang, X. The significance of phosphorus in algae growth and the subsequent ecological response of consumers. J. Freshw. Ecol. 2022, 37, 57–69. [Google Scholar] [CrossRef]
- Mao, X.; Lao, Y.; Sun, H.; Li, X.; Yu, J.; Chen, F. Time-resolved transcriptome analysis during transitions of sulfur nutritional status provides insight into triacylglycerol (TAG) and astaxanthin accumulation in the green alga Chromochloris zofingiensis. Biotechnol. Biofuels 2020, 13, 128. [Google Scholar] [CrossRef]
- Cakmak, T.; Angun, P.; Ozkan, A.D.; Cakmak, Z.; Olmez, T.T.; Tekinay, T. Nitrogen and sulfur deprivation differentiate lipid accumulation targets of Chlamydomonas reinhardtii. Bioengineered 2012, 3, 343–346. [Google Scholar] [CrossRef] [Green Version]
- Krzemińska, I.; Szymańska, M.; Ciempiel, W.; Piasecka, A. Auxin supplementation under nitrogen limitation enhanced oleic acid and MUFA content in Eustigmatos calaminaris biomass with potential for biodiesel production. Sci. Rep. 2023, 13, 594. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; Li, D.; Yuan, Y.; Zhou, L.; Li, X.; Wu, T.; Wang, L.; Zhao, Q.; Wei, W.; Sun, Y. Improving carbohydrate and starch accumulation in Chlorella sp. AE10 by a novel two-stage process with cell dilution. Biotechnol. Biofuels 2017, 10, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michelon, W.; Da Silva, M.L.B.; Mezzari, M.P.; Pirolli, M.; Prandini, J.M.; Soares, H.M. Effects of Nitrogen and Phosphorus on Biochemical Composition of Microalgae Polyculture Harvested from Phycoremediation of Piggery Wastewater Digestate. Appl. Biochem. Biotechnol. 2016, 178, 1407–1419. [Google Scholar] [CrossRef]
- Zhuang, L.-L.; Azimi, Y.; Yu, D.; Wu, Y.-H.; Hu, H.-Y. Effects of nitrogen and phosphorus concentrations on the growth of microalgae Scenedesmus. LX1 in suspended-solid phase photobioreactors (ssPBR). Biomass Bioenergy 2018, 109, 47–53. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, H.; Yang, J.; Zhou, Y.; Wang, X.; Dou, S.; Li, L.; Liu, G.; Yang, M. Effect of sulfur limitation strategies on glucose-based carbohydrate production from Chlorella sorokiniana. Renew. Energy 2022, 200, 449–456. [Google Scholar] [CrossRef]
- Narchonai, G.; Arutselvan, C.; LewisOscar, F.; Thajuddin, N. Enhancing starch accumulation/production in Chlorococcum humicola through sulphur limitation and 2,4-D treatment for butanol production. Biotechnol. Rep. 2020, 28, e00528. [Google Scholar] [CrossRef]
- Subramanian, S.; Barry, A.N.; Pieris, S.; Sayre, R.T. Comparative energetics and kinetics of autotrophic lipid and starch metabolism in chlorophytic microalgae: Implications for biomass and biofuel production. Biotechnol. Biofuels 2013, 6, 150. [Google Scholar] [CrossRef] [Green Version]
- Fattore, N.; Bellan, A.; Pedroletti, L.; Vitulo, N.; Morosinotto, T. Acclimation of photosynthesis and lipids biosynthesis to prolonged nitrogen and phosphorus limitation in Nannochloropsis gaditana. Algal Res. 2021, 58, 102368. [Google Scholar] [CrossRef]
- Bišová, K.; Zachleder, V. Cell-cycle regulation in green algae dividing by multiple fission. J. Exp. Bot. 2014, 65, 2585–2602. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.W.M.; Lin, H.; Shen, H.; Lee, Y.K. Nitrogen-induced metabolic changes and molecular determinants of carbon allocation in Dunaliella tertiolecta. Sci. Rep. 2016, 6, 37235. [Google Scholar] [CrossRef] [Green Version]
- Geigenberger, P. Regulation of Starch Biosynthesis in Response to a Fluctuating Environment. Plant Physiol. 2011, 155, 1566–1577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ran, W.; Wang, H.; Liu, Y.; Qi, M.; Xiang, Q.; Yao, C.; Zhang, Y.; Lan, X. Storage of starch and lipids in microalgae: Biosynthesis and manipulation by nutrients. Bioresour. Technol. 2019, 291, 121894. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Yang, J.; Wang, X.; Peng, Q.; Han, Y.; Liu, X.; Liu, K.; Dou, S.; Li, L.; Liu, G.; et al. Starch accumulation dynamics and transcriptome analysis of Chlorella sorokiniana during transition of sulfur nutritional status. Front. Mar. Sci. 2022, 9, 986400. [Google Scholar] [CrossRef]
- Rosa, R.M.; Machado, M.; Vieira Vaz, M.G.M.; dos Santos, R.L.; do Nascimento, A.G.; Araújo, W.L.; Nunes-Nesi, A. Urea as a source of nitrogen and carbon leads to increased photosynthesis rates in Chlamydomonas reinhardtii under mixotrophy. J. Biotechnol. 2023, 367, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Zhang, Z.; Tang, Z.; Ho, S.-H.; Shi, X.; Liu, L.; Xie, Y.; Chen, J. Enhancement of co-production of lutein and protein in Chlorella sorokiniana FZU60 using different bioprocess operation strategies. Bioresour. Bioprocess. 2021, 8, 82. [Google Scholar] [CrossRef]
- Kamalanathan, M.; Pierangelini, M.; Shearman, L.A.; Gleadow, R.; Beardall, J. Impacts of nitrogen and phosphorus starvation on the physiology of Chlamydomonas reinhardtii. J. Appl. Phycol. 2016, 28, 1509–1520. [Google Scholar] [CrossRef]
- Schmollinger, S.; Mühlhaus, T.; Boyle, N.R.; Blaby, I.K.; Casero, D.; Mettler, T.; Moseley, J.L.; Kropat, J.; Sommer, F.; Strenkert, D.; et al. Nitrogen-Sparing Mechanisms in Chlamydomonas Affect the Transcriptome, the Proteome, and Photosynthetic Metabolism. Plant Cell 2014, 26, 1410–1435. [Google Scholar] [CrossRef] [Green Version]
- Plouviez, M.; Abyadeh, M.; Hasan, M.; Mirzaei, M.; Paulo, J.A.; Guieysse, B. The proteome of Chlamydomonas reinhardtii during phosphorus depletion and repletion. Algal Res. 2023, 71, 103037. [Google Scholar] [CrossRef]
- Shang, C.; Zhu, S.; Wang, Z.; Qin, L.; Alam, M.A.; Xie, J.; Yuan, Z. Proteome response of Dunaliella parva induced by nitrogen limitation. Algal Res. 2017, 23, 196–202. [Google Scholar] [CrossRef]
- Estevam, B.R.; Pinto, L.F.R.; Filho, R.M.; Fregolente, L.V. Growth and Metabolite Production in Chlorella sp.: Analysis of Cultivation System and Nutrient Reduction. BioEnergy Res. 2022. [Google Scholar] [CrossRef]
- Salbitani, G.; Perrone, A.; Rosati, L.; Laezza, C.; Carfagna, S. Sulfur Starvation in Extremophilic Microalga Galdieria sulphuraria: Can Glutathione Contribute to Stress Tolerance? Plants 2022, 11, 481. [Google Scholar] [CrossRef] [PubMed]
- Irihimovitch, V.; Yehudai-Resheff, S. Phosphate and sulfur limitation responses in the chloroplast of Chlamydomonas reinhardtii. FEMS Microbiol. Lett. 2008, 283, 1–8. [Google Scholar] [CrossRef]
- Xu, L.; Cheng, X.; Wang, Q. Enhanced Lipid Production in Chlamydomonas reinhardtii by Co-culturing with Azotobacter chroococcum. Front. Plant Sci. 2018, 9, 741. [Google Scholar] [CrossRef]
- Yadavalli, R.; Ratnapuram, H.; Motamarry, S.; Reddy, C.N.; Ashokkumar, V.; Kuppam, C. Simultaneous production of flavonoids and lipids from Chlorella vulgaris and Chlorella pyrenoidosa. Biomass Convers. Biorefinery 2022, 12, 683–691. [Google Scholar] [CrossRef]
- Gao, B.; Wang, F.; Huang, L.; Liu, H.; Zhong, Y.; Zhang, C. Biomass, lipid accumulation kinetics, and the transcriptome of heterotrophic oleaginous microalga Tetradesmus bernardii under different carbon and nitrogen sources. Biotechnol. Biofuels 2021, 14, 4. [Google Scholar] [CrossRef]
- Sirikhachornkit, A.; Suttangkakul, A.; Vuttipongchaikij, S.; Juntawong, P. De novo transcriptome analysis and gene expression profiling of an oleaginous microalga Scenedesmus acutus TISTR8540 during nitrogen deprivation-induced lipid accumulation. Sci. Rep. 2018, 8, 3668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, F.; Xiang, W.; Li, T.; Long, L. Transcriptome analysis for phosphorus starvation-induced lipid accumulation in Scenedesmus sp. Sci. Rep. 2018, 8, 16420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Çakmak, Z.E.; Ölmez, T.T.; Çakmak, T.; Menemen, Y.; Tekinay, T. Induction of triacylglycerol production in Chlamydomonas reinhardtii: Comparative analysis of different element regimes. Bioresour. Technol. 2014, 155, 379–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-De la Torre, A.E.; Ochoa-Alfaro, A.E.; Rocha-Uribe, A.; Soria-Guerra, R.E. Effects of sulfur and phosphorus concentration on the lipid accumulation and fatty acid profile in Chlorella vulgaris (Chlorophyta). Folia Microbiol. 2023, 68, 453–463. [Google Scholar] [CrossRef]
- Boyle, N.R.; Page, M.D.; Liu, B.; Blaby, I.K.; Casero, D.; Kropat, J.; Cokus, S.J.; Hong-Hermesdorf, A.; Shaw, J.; Karpowicz, S.J.; et al. Three Acyltransferases and Nitrogen-responsive Regulator Are Implicated in Nitrogen Starvation-induced Triacylglycerol Accumulation in Chlamydomonas. J. Biol. Chem. 2012, 287, 15811–15825. [Google Scholar] [CrossRef] [Green Version]
- Zheng, S.; Zou, S.; Feng, T.; Sun, S.; Guo, X.; He, M.; Wang, C.; Chen, H.; Wang, Q. Low temperature combined with high inoculum density improves alpha-linolenic acid production and biochemical characteristics of Chlamydomonas reinhardtii. Bioresour. Technol. 2022, 348, 126746. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Fosse, H.K.; Li, K.; Chauton, M.S.; Vadstein, O.; Reitan, K.I. Influence of Nitrogen Limitation on Lipid Accumulation and EPA and DHA Content in Four Marine Microalgae for Possible Use in Aquafeed. Front. Mar. Sci. 2019, 6, 95. [Google Scholar] [CrossRef]
- López-legarda, X.; Taramuel-gallardo, A.; Arboleda-, C. Comparison of methods using sulfuric acid for determination of total sugars. Rev. Cuba. Quím. 2017, 29, 180–198. [Google Scholar]
- Mishra, S.K.; Suh, W.I.; Farooq, W.; Moon, M.; Shrivastav, A.; Park, M.S.; Yang, J. Rapid quantification of microalgal lipids in aqueous medium by a simple colorimetric method. Bioresour. Technol. 2014, 155, 330–333. [Google Scholar] [CrossRef] [PubMed]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RunEst | Run | Blocks | Concentration | Nutrient |
|---|---|---|---|---|
| 11 | 1 | 2 | Low | S |
| 17 | 2 | 2 | High | S |
| 18 | 3 | 2 | High | P |
| 12 | 4 | 2 | Low | P |
| 15 | 5 | 2 | Med | P |
| 16 | 6 | 2 | High | N |
| 13 | 7 | 2 | Med | N |
| 10 | 8 | 2 | Low | N |
| 14 | 9 | 2 | Med | S |
| 24 | 10 | 3 | Med | P |
| 20 | 11 | 3 | Low | S |
| 19 | 12 | 3 | Low | N |
| 22 | 13 | 3 | Med | N |
| 25 | 14 | 3 | High | N |
| 26 | 15 | 3 | High | S |
| 23 | 16 | 3 | Med | S |
| 21 | 17 | 3 | Low | P |
| 27 | 18 | 3 | High | P |
| 8 | 19 | 1 | High | S |
| 6 | 20 | 1 | Med | P |
| 3 | 21 | 1 | Low | P |
| 4 | 22 | 1 | Med | N |
| 1 | 23 | 1 | Low | N |
| 5 | 24 | 1 | Med | S |
| 7 | 25 | 1 | High | N |
| 2 | 26 | 1 | Low | S |
| 9 | 27 | 1 | High | P |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Pacheco, I.Y.; Ayala-Moreno, V.G.; Mejia-Melara, C.A.; Rodríguez-Rodríguez, J.; Cuellar-Bermudez, S.P.; González-González, R.B.; Coronado-Apodaca, K.G.; Farfan-Cabrera, L.I.; González-Meza, G.M.; Iqbal, H.M.N.; et al. Growth Behavior, Biomass Composition and Fatty Acid Methyl Esters (FAMEs) Production Potential of Chlamydomonas reinhardtii, and Chlorella vulgaris Cultures. Mar. Drugs 2023, 21, 450. https://doi.org/10.3390/md21080450
López-Pacheco IY, Ayala-Moreno VG, Mejia-Melara CA, Rodríguez-Rodríguez J, Cuellar-Bermudez SP, González-González RB, Coronado-Apodaca KG, Farfan-Cabrera LI, González-Meza GM, Iqbal HMN, et al. Growth Behavior, Biomass Composition and Fatty Acid Methyl Esters (FAMEs) Production Potential of Chlamydomonas reinhardtii, and Chlorella vulgaris Cultures. Marine Drugs. 2023; 21(8):450. https://doi.org/10.3390/md21080450
Chicago/Turabian StyleLópez-Pacheco, Itzel Y., Victoria Guadalupe Ayala-Moreno, Catherinne Arlette Mejia-Melara, José Rodríguez-Rodríguez, Sara P. Cuellar-Bermudez, Reyna Berenice González-González, Karina G. Coronado-Apodaca, Leonardo I. Farfan-Cabrera, Georgia María González-Meza, Hafiz M. N. Iqbal, and et al. 2023. "Growth Behavior, Biomass Composition and Fatty Acid Methyl Esters (FAMEs) Production Potential of Chlamydomonas reinhardtii, and Chlorella vulgaris Cultures" Marine Drugs 21, no. 8: 450. https://doi.org/10.3390/md21080450
APA StyleLópez-Pacheco, I. Y., Ayala-Moreno, V. G., Mejia-Melara, C. A., Rodríguez-Rodríguez, J., Cuellar-Bermudez, S. P., González-González, R. B., Coronado-Apodaca, K. G., Farfan-Cabrera, L. I., González-Meza, G. M., Iqbal, H. M. N., & Parra-Saldívar, R. (2023). Growth Behavior, Biomass Composition and Fatty Acid Methyl Esters (FAMEs) Production Potential of Chlamydomonas reinhardtii, and Chlorella vulgaris Cultures. Marine Drugs, 21(8), 450. https://doi.org/10.3390/md21080450