

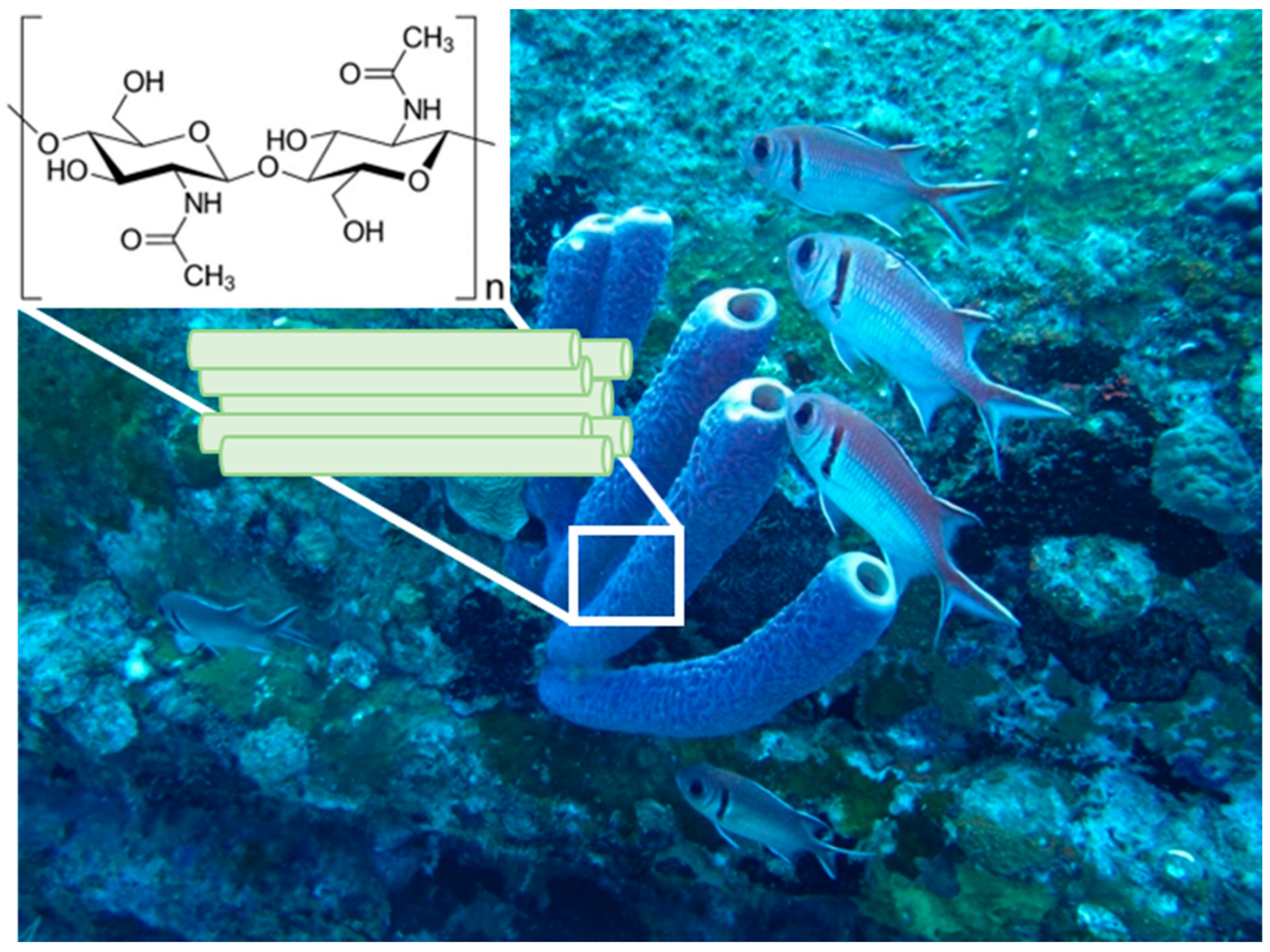
On the Mechanical Properties of Microfibre-Based 3D Chitinous Scaffolds from Selected Verongiida Sponges

 ,
, 

 , , , ,
, , , ,  ,
, 
Abstract
:1. Introduction
2. Results
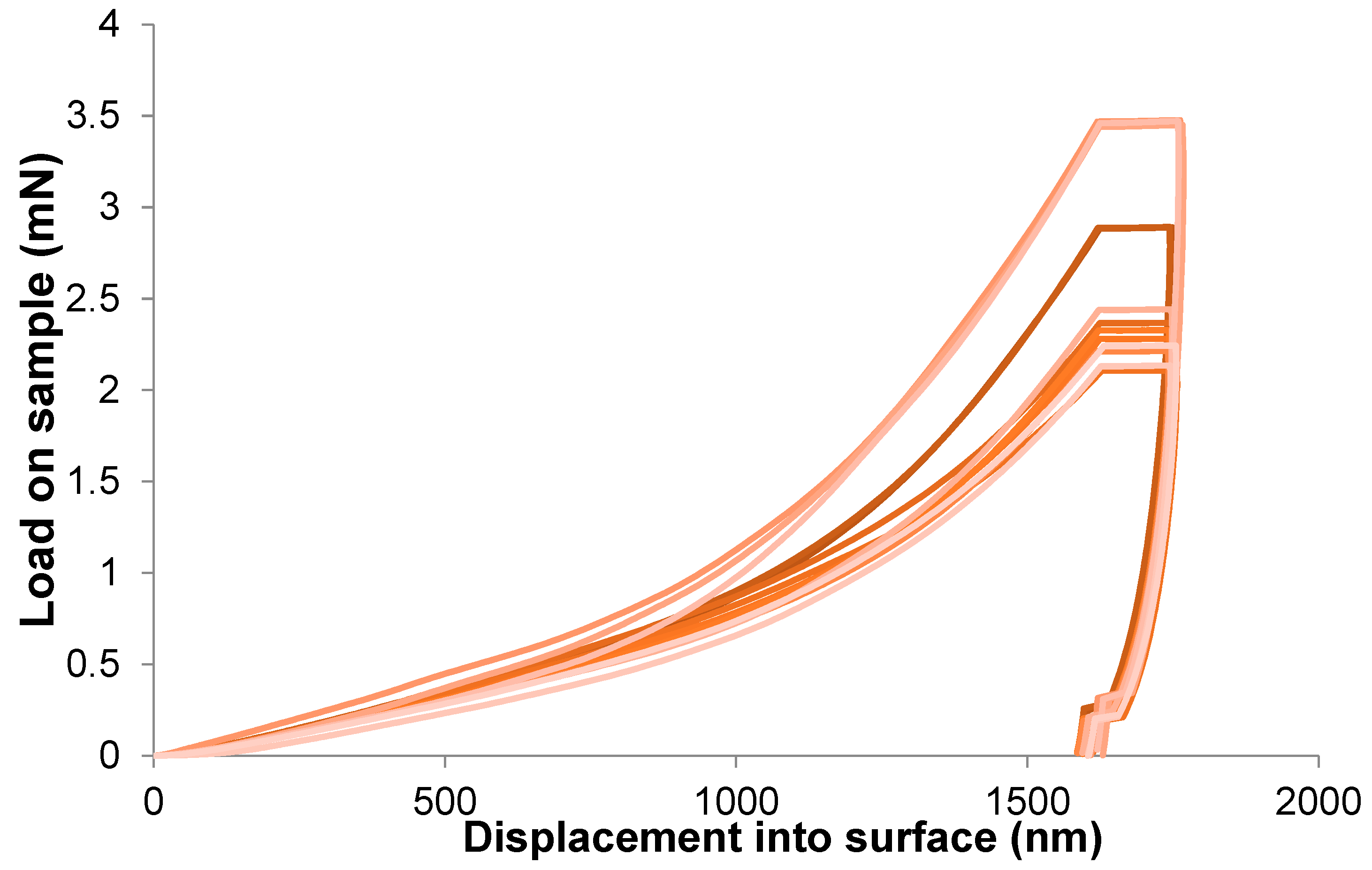
2.1. Monotonic Compression Test
2.2. Nanoindentation
2.3. Scanning Electron Microscopy
2.4. Digital Optical Microscopy
3. Discussion
- At a low-stress level, an elastic stage can be observed. The spongious structure is deformed, but it is still structurally stable. The stresses transferred between the spongious struts are insufficient to induce permanent structural modifications. In this region, a modulus peak is usually observed, indicating the transition from the stable phase toward the subsequent collapse stage;
- During the collapse stage, the characteristic plateau in the case of both of the considered curves is present. The plateau is associated with the collapse of the pores. A series of local collapses percolate through the structure at some critical stress. In particular, for an elastic foam, the plateau is due to elastic buckling, whereas, in the case of elastoplastic foams, it is due to the formation of plastic hinges [44]. Depending on the compressive mechanical behaviour of the cellular material, the plateau region can be characterised by a flat or slightly increasing slope stress plateau;
- When the pores’ closure is almost completed, spongious struts begin to interact together whereby a rapid increase of stress takes place. At the same time, an abrupt increase in the modulus is observed. This last region of the curve is called the densification stage. When analysing the tangent modulus–strain curve, one can observe that its densification region is smoother than the collapse region, which facilitates the accurate discrimination between them [41].
4. Materials and Methods
4.1. Sample Preparation
4.2. Monotonic Compression Test
4.3. Nanoindentation
4.4. Scanning Electron Microscopy
4.5. Digital Optical Microscopy
4.6. Measurment of Porosity, Pore Size, and Fibre Diameter
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tsurkan, D.; Wysokowski, M.; Petrenko, I.; Voronkina, A.; Khrunyk, Y.; Fursov, A.; Ehrlich, H. Modern scaffolding strategies based on naturally pre-fabricated 3D biomaterials of poriferan origin. Appl. Phys. A 2020, 126, 382. [Google Scholar] [CrossRef]
- Binnewerg, B.; Schubert, M.; Voronkina, A.; Muzychka, L.; Wysokowski, M.; Petrenko, I.; Djurović, M.; Kovalchuk, V.; Tsurkan, M.; Martinovic, R.; et al. Marine biomaterials: Biomimetic and pharmacological potential of cultivated Aplysina aerophoba marine demosponge. Mater. Sci. Eng. C Mater. Biol. Appl. 2020, 109, 110566. (In English) [Google Scholar] [CrossRef]
- Khrunyk, Y.; Lach, S.; Petrenko, I.; Ehrlich, H. Progress in Modern Marine Biomaterials Research. Mar. Drugs 2020, 18, 589. [Google Scholar] [CrossRef]
- Ehrlich, H.; Wysokowski, M.; Jesionowski, T. The philosophy of extreme biomimetics. Sustain. Mater. Technol. 2022, 32, e00447. [Google Scholar] [CrossRef]
- Costa, G.; Violi, B.; Bavestrello, G.; Pansini, M.; Bertolino, M. Aplysina aerophoba (Nardo, 1833) (Porifera, Demospongiae): An unexpected miniaturised growth form from the tidal zone of Mediterranean caves: Morphology and DNA barcoding. Eur. Zool. J. 2020, 87, 73–81. [Google Scholar] [CrossRef]
- Mutsenko, V.V.; Gryshkov, O.; Lauterboeck, L.; Rogulska, O.; Tarusin, D.N.; Bazhenov, V.V.; Schütz, K.; Brüggemeier, S.; Gossla, E.; Akkineni, A.R.; et al. Novel chitin scaffolds derived from marine sponge Ianthella basta for tissue engineering approaches based on human mesenchymal stromal cells: Biocompatibility and cryopreservation. Int. J. Biol. Macromol. 2017, 104, 1955–1965. (In English) [Google Scholar] [CrossRef]
- Mutsenko, V.V.; Bazhenov, V.V.; Rogulska, O.; Tarusin, D.N.; Schütz, K.; Brüggemeier, S.; Gossla, E.; Akkineni, A.R.; Meißner, H.; Lode, A.; et al. 3D chitinous scaffolds derived from cultivated marine demosponge Aplysina aerophoba for tissue engineering approaches based on human mesenchymal stromal cells. Int. J. Biol. Macromol. 2017, 104, 1966–1974. (In English) [Google Scholar] [CrossRef]
- Schubert, M.; Binnewerg, B.; Voronkina, A.; Muzychka, L.; Wysokowski, M.; Petrenko, I.; Kovalchuk, V.; Tsurkan, M.; Martinovic, R.; Bechmann, N.; et al. Naturally Prefabricated Marine Biomaterials: Isolation and Applications of Flat Chitinous 3D Scaffolds from Ianthella labyrinthus (Demospongiae: Verongiida). Int. J. Mol. Sci. 2019, 20, 5105. [Google Scholar] [CrossRef]
- Kovalchuk, V.; Voronkina, A.; Binnewerg, B.; Schubert, M.; Muzychka, L.; Wysokowski, M.; Tsurkan, M.V.; Bechmann, N.; Petrenko, I.; Fursov, A.; et al. Naturally Drug-Loaded Chitin: Isolation and Applications. Mar. Drugs 2019, 17, 574. (In English) [Google Scholar] [CrossRef]
- Muzychka, L.; Voronkina, A.; Kovalchuk, V.; Smolii, O.B.; Wysokowski, M.; Petrenko, I.; Youssef, D.T.A.; Ehrlich, I.; Ehrlich, H. Marine biomimetics: Bromotyrosines loaded chitinous skeleton as source of antibacterial agents. Appl. Phys. A Mater. Sci. Process. 2021, 127, 15. (In English) [Google Scholar] [CrossRef]
- Machałowski, T.; Rusak, A.; Wiatrak, B.; Haczkiewicz-Leśniak, K.; Popiel, A.; Jaroszewicz, J.; Żak, A.; Podhorska-Okołów, M.; Jesionowski, T. Naturally Formed Chitinous Skeleton Isolated from the Marine Demosponge Aplysina fistularis as a 3D Scaffold for Tissue Engineering. Materials 2021, 14, 2992. [Google Scholar] [CrossRef]
- Nowacki, K.; Galiński, M.; Fursov, A.; Voronkina, A.; Meissner, H.; Petrenko, I.; Stelling, A.L.; Ehrlich, H. Electrolysis as a Universal Approach for Isolation of Diverse Chitin Scaffolds from Selected Marine Demosponges. Mar. Drugs 2022, 20, 665. [Google Scholar] [CrossRef]
- Klinger, C.; Żółtowska-Aksamitowska, S.; Wysokowski, M.; Tsurkan, M.V.; Galli, R.; Petrenko, I.; Machałowski, T.; Ereskovsky, A.; Martinović, R.; Muzychka, L.; et al. Express Method for Isolation of Ready-to-Use 3D Chitin Scaffolds from Aplysina archeri (Aplysineidae: Verongiida) Demosponge. Mar. Drugs 2019, 17, 131. [Google Scholar] [CrossRef] [PubMed]
- Klinger, C.; Żółtowska, S.; Jesionowski, T. Isolation of Chitin from Aplysina aerophoba Using a Microwave Approach. Prog. Chem. Appl. Chitin Deriv. 2019, XXIV, 61–74. [Google Scholar] [CrossRef]
- Żółtowska, S.; Klinger, C.; Petrenko, I.; Wysokowski, M.; Joseph, Y.; Jesionowski, T.; Ehrlich, H. Methods of Isolating Chitin from Sponges (Porifera). In Chitin and Chitosan: Properties and Applications; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2019; pp. 35–59. [Google Scholar]
- Duminis, T. Natural Polymers with Bioactive Glass Additives for Bone Regeneration: Chemistry and Trends. Nat. Prod. J. 2023, 14, e280423216342. [Google Scholar] [CrossRef]
- Duminis, T.; Shahid, S.; Hill, R.G. Apatite glass-ceramics: A review. Front. Mater. 2017, 3, 59. [Google Scholar] [CrossRef]
- Kaya, M.; Mujtaba, M.; Ehrlich, H.; Salaberria, A.M.; Baran, T.; Amemiya, C.T.; Galli, R.; Akyuz, L.; Sargin, I.; Labidi, J. On chemistry of γ-chitin. Carbohydr. Polym. 2017, 176, 177–186. [Google Scholar] [CrossRef]
- Tsurkan, M.V.; Voronkina, A.; Khrunyk, Y.; Wysokowski, M.; Petrenko, I.; Ehrlich, H. Progress in chitin analytics. Carbohydr. Polym. 2021, 252, 117204. [Google Scholar] [CrossRef]
- Sawada, D.; Nishiyama, Y.; Langan, P.; Forsyth, V.T.; Kimura, S.; Wada, M. Direct Determination of the Hydrogen Bonding Arrangement in Anhydrous β-Chitin by Neutron Fiber Diffraction. Biomacromolecules 2012, 13, 288–291. [Google Scholar] [CrossRef]
- Wei, A.; Fu, J.; Guo, F. Mechanical properties of chitin polymorphs: A computational study. J. Mater. Sci. 2021, 56, 12048–12058. [Google Scholar] [CrossRef]
- Deringer, V.L.; Englert, U.; Dronskowski, R. Nature, Strength, and Cooperativity of the Hydrogen-Bonding Network in α-Chitin. Biomacromolecules 2016, 17, 996–1003. [Google Scholar] [CrossRef]
- Cui, J.; Yu, Z.; Lau, D. Effect of Acetyl Group on Mechanical Properties of Chitin/Chitosan Nanocrystal: A Molecular Dynamics Study. Int. J. Mol. Sci. 2016, 17, 61. [Google Scholar] [CrossRef]
- Montroni, D.; Fermani, S.; Morellato, K.; Torri, G.; Naggi, A.; Cristofolini, L.; Falini, G. β-Chitin samples with similar microfibril arrangement change mechanical properties varying the degree of acetylation. Carbohydr. Polym. 2019, 207, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Dziedzic, I.; Voronkina, A.; Pajewska-Szmyt, M.; Kotula, M.; Kubiak, A.; Meissner, H.; Duminis, T.; Ehrlich, H. The Loss of Structural Integrity of 3D Chitin Scaffolds from Aplysina aerophoba Marine Demosponge after Treatment with LiOH. Mar. Drugs 2023, 21, 334. [Google Scholar] [CrossRef] [PubMed]
- Brunner, E.; Ehrlich, H.; Schupp, P.; Hedrich, R.; Hunoldt, S.; Kammer, M.; Machill, S.; Paasch, S.; Bazhenov, V.V.; Kurek, D.V.; et al. Chitin-based scaffolds are an integral part of the skeleton of the marine demosponge Ianthella basta. J. Struct. Biol. 2009, 168, 539–547. (In English) [Google Scholar] [CrossRef]
- Geahchan, S.; Ehrlich, H.; Rahman, M.A. The Anti-Viral Applications of Marine Resources for COVID-19 Treatment: An Overview. Mar. Drugs 2021, 19, 409. (In English) [Google Scholar] [CrossRef]
- Yusof, N.L.B.M.; Lim, L.Y.; Khor, E. Flexible chitin films: Structural studies. Carbohydr. Res. 2004, 339, 2701–2711. [Google Scholar] [CrossRef] [PubMed]
- Siamantouras, E.; Woodrow, C.; Celiker, E.; Cullen, D.A.; Hills, C.E.; Squires, P.E.; Montealegre-Z, F. Quantification of bush-cricket acoustic trachea mechanics using Atomic Force Microscopy nanoindentation. Acta Biomater. 2022, 153, 399–410. [Google Scholar] [CrossRef]
- Chen, P.-Y.; Lin, A.Y.-M.; McKittrick, J.; Meyers, M.A. Structure and mechanical properties of crab exoskeletons. Acta Biomater. 2008, 4, 587–596. [Google Scholar] [CrossRef]
- Gadgey, K.K.; Bahekar, A. Studies on extraction methods of chitin from crab shell and investigation of its mechanical properties. Int. J. Mech. Eng. Technol. 2017, 8, 220–231. [Google Scholar]
- Gallagher, A.J.; Ní Annaidh, A.; Bruyère-Garnier, K. Dynamic tensile properties of human skin. In Proceedings of the 2012 IRCOBI Conference Proceedings, Dublin, Ireland, 12–14 September 2012; pp. 494–502. [Google Scholar]
- Rohlmann, A.; Zilch, H.; Bergmann, G.; Kolbel, R. Material properties of femoral cancellous bone in axial loading—Part I: Time independent properties. Arch. Orthop. Trauma. Surg. 1980, 97, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Duan, B.; Chang, C.; Zhang, L. Structure and properties of films fabricated from chitin solution by coagulating with heating. J. Appl. Polym. Sci. 2014, 131, 1–7. [Google Scholar] [CrossRef]
- Kim, J.; Ha, C.S.; Jo, N.J. Synthesis and properties of biodegradable chitin-graft-poly(L-lactide) copolymers. Polym. Int. 2002, 51, 1123–1128. [Google Scholar] [CrossRef]
- Chen, B.; Sun, K.; Ren, T. Mechanical and viscoelastic properties of chitin fiber reinforced poly(ε-caprolactone). Eur. Polym. J. 2005, 41, 453–457. [Google Scholar] [CrossRef]
- Jayakumar, R.; Tamura, H. Synthesis, characterization and thermal properties of chitin-g-poly(epsilon-caprolactone) copolymers by using chitin gel. Int. J. Biol. Macromol. 2008, 43, 32–36. (In English) [Google Scholar] [CrossRef] [PubMed]
- Wysokowski, M.; Petrenko, I.; Stelling, A.L.; Stawski, D.; Jesionowski, T.; Ehrlich, H. Poriferan Chitin as a Versatile Template for Extreme Biomimetics. Polymers 2015, 7, 235–265. [Google Scholar] [CrossRef]
- Ifuku, S.; Nogi, M.; Abe, K.; Yoshioka, M.; Morimoto, M.; Saimoto, H.; Yano, H. Preparation of Chitin Nanofibers with a Uniform Width as α-Chitin from Crab Shells. Biomacromolecules 2009, 10, 1584–1588. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.; Choy, S.; Park, Y.; Jung, Y.M.; Koo, J.M.; Hwang, D.S. Different Molecular Interaction between Collagen and α- or β-Chitin in Mechanically Improved Electrospun Composite. Mar. Drugs 2019, 17, 318. [Google Scholar] [CrossRef]
- Yu, Z.; Lau, D. Molecular dynamics study on stiffness and ductility in chitin–protein composite. J. Mater. Sci. 2015, 50, 7149–7157. [Google Scholar] [CrossRef]
- Nowacki, K.; Stępniak, I.; Machałowski, T.; Wysokowski, M.; Petrenko, I.; Schimpf, C.; Rafaja, D.; Langer, E.; Richter, A.; Ziętek, J.; et al. Electrochemical method for isolation of chitinous 3D scaffolds from cultivated Aplysina aerophoba marine demosponge and its biomimetic application. Appl. Phys. A 2020, 126, 368. [Google Scholar] [CrossRef]
- Fu, Y.B.; Ogden, R.W. Nonlinear Elasticity: Theory and Applications; Cambridge University Press: Cambridge, UK, 2001. [Google Scholar]
- Calabrese, L.; Bonaccorsi, L.; Bruzzaniti, P.; Gullì, G.; Freni, A.; Proverbio, E. Zeolite filled siloxane composite foams: Compression property. J. Appl. Polym. Sci. 2018, 135, 46145. [Google Scholar] [CrossRef]
- Granito, R.N.; Custódio, M.R.; Rennó, A.C.M. Natural marine sponges for bone tissue engineering: The state of art and future perspectives. J. Biomed. Mater. Res. B Appl. Biomater. 2017, 105, 1717–1727. (In English) [Google Scholar] [CrossRef] [PubMed]
- Chatzimitakos, T.G.; Stalikas, C.D. Sponges and Sponge-Like Materials in Sample Preparation: A Journey from Past to Present and into the Future. Molecules 2020, 25, 3673. (In English) [Google Scholar] [CrossRef] [PubMed]
- Woesz, A.; Weaver, J.C.; Kazanci, M.; Dauphin, Y.; Aizenberg, J.; Morse, D.E.; Fratzl, P. Micromechanical properties of biological silica in skeletons of deep-sea sponges. J. Mater. Res. 2006, 21, 2068–2078. [Google Scholar] [CrossRef]
- Louden, D.; Inderbitzin, S.; Peng, Z.; de Nys, R. Development of a new protocol for testing bath sponge quality. Aquaculture 2007, 271, 275–285. [Google Scholar] [CrossRef]
- Shen, J.; Min Xie, Y.; Huang, X.; Zhou, S.; Ruan, D. Mechanical properties of luffa sponge. J. Mech. Behav. Biomed. Mater. 2012, 15, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Ghodbane, S.A.; Dunn, M.G. Physical and mechanical properties of cross-linked type I collagen scaffolds derived from bovine, porcine, and ovine tendons. J. Biomed. Mater. Res. A 2016, 104, 2685–2692. (In English) [Google Scholar] [CrossRef]
- Wang, Z.; Lin, S.; Li, X.; Zou, H.; Zhuo, B.; Ti, P.; Yuan, Q. Optimization and absorption performance of wood sponge. J. Mater. Sci. 2021, 56, 8479–8496. [Google Scholar] [CrossRef]
- Ofem, M.I.; Muhammed, M.; Umar, M. Mechanical properties of Dungeness crab based chitin. Int. J. Sci. Eng. Res. 2015, 6, 1737–1743. [Google Scholar]
- Ifuku, S.; Saimoto, H. Chitin nanofibers: Preparations, modifications, and applications. Nanoscale 2012, 4, 3308–3318. [Google Scholar] [CrossRef] [PubMed]
- Mushi, N.E.; Butchosa, N.; Salajkova, M.; Zhou, Q.; Berglund, L.A. Nanostructured membranes based on native chitin nanofibers prepared by mild process. Carbohydr. Polym. 2014, 112, 255–263. [Google Scholar] [CrossRef]
- Wang, J.; Brasch, M.E.; Baker, R.M.; Tseng, L.F.; Peña, A.N.; Henderson, J.H. Shape memory activation can affect cell seeding of shape memory polymer scaffolds designed for tissue engineering and regenerative medicine. J. Mater. Sci. Mater. Med. 2017, 28, 151. (In English) [Google Scholar] [CrossRef]
- Lv, S.; Cai, M.; Leng, F.; Jiang, X. Biodegradable carboxymethyl chitin-based hemostatic sponges with high strength and shape memory for non-compressible hemorrhage. Carbohydr. Polym. 2022, 288, 119369. (In English) [Google Scholar] [CrossRef]
- Ross, C.T.F.; Case, T.J.; Chilver, A. Strength of Materials and Structures; Elsevier Science: Amsterdam, The Netherlands, 1999. [Google Scholar]
- Welinder, B.S. Halogenated tyrosines from the cuticle of Limulus polyphemus (L.). Biochim. Biophys. Acta 1972, 279, 491–497. (In English) [Google Scholar] [CrossRef] [PubMed]
- Hunt, S.; Breuer, S.W. Chlorinated and Brominated Tyrosine Residues in Molluscan Scleroprotein. Biochem. Soc. Trans. 1973, 1, 215–216. [Google Scholar] [CrossRef]
- Montroni, D.; Sparla, F.; Fermani, S.; Falini, G. Influence of proteins on mechanical properties of a natural chitin-protein composite. Acta Biomater. 2021, 120, 81–90. [Google Scholar] [CrossRef]
- Torres-Rendon, J.G.; Schacher, F.H.; Ifuku, S.; Walther, A. Mechanical Performance of Macrofibers of Cellulose and Chitin Nanofibrils Aligned by Wet-Stretching: A Critical Comparison. Biomacromolecules 2014, 15, 2709–2717. [Google Scholar] [CrossRef] [PubMed]
- Mushi, N.E. A review on native well-preserved chitin nanofibrils for materials of high mechanical performance. Int. J. Biol. Macromol. 2021, 178, 591–606. (In English) [Google Scholar] [CrossRef]
- Ehrlich, H. Chitin of poriferan origin as a unique biological material. In Blue Biotechnology: Production and Use of Marine Molecules; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2018; Volume 2, pp. 821–854. [Google Scholar]
- Mutsenko, V.; Gryshkov, O.; Rogulska, O.; Lode, A.; Petrenko, A.Y.; Gelinsky, M.; Glasmacher, B.; Ehrlich, H. Chitinous scaffolds from marine sponges for tissue engineering. In Marine-Derived Biomaterials for Tissue Engineering Applications; Springer: Singapore, 2019; pp. 285–307. [Google Scholar]
- Ehrlich, H.; Steck, E.; Ilan, M.; Maldonado, M.; Muricy, G.; Bavestrello, G.; Kljajic, Z.; Carballo, J.; Schiaparelli, S.; Ereskovsky, A. Three-dimensional chitin-based scaffolds from Verongida sponges (Demospongiae: Porifera). Part II: Biomimetic potential and applications. Int. J. Biol. Macromol. 2010, 47, 141–145. [Google Scholar] [CrossRef]
- Machałowski, T.; Czajka, M.; Petrenko, I.; Meissner, H.; Schimpf, C.; Rafaja, D.; Ziętek, J.; Dzięgiel, B.; Adaszek, Ł.; Voronkina, A.; et al. Functionalization of 3D Chitinous Skeletal Scaffolds of Sponge Origin Using Silver Nanoparticles and Their Antibacterial Properties. Mar. Drugs 2020, 18, 304. [Google Scholar] [CrossRef]
- Ehrlich, H.; Simon, P.; Carrillo-Cabrera, W.; Bazhenov, V.V.; Botting, J.P.; Ilan, M.; Ereskovsky, A.V.; Muricy, G.; Worch, H.; Mensch, A.; et al. Insights into Chemistry of Biological Materials: Newly Discovered Silica-Aragonite-Chitin Biocomposites in Demosponges. Chem. Mater. 2010, 22, 1462–1471. [Google Scholar] [CrossRef]
- Bo, M.; Bavestrello, G.; Kurek, D.; Paasch, S.; Brunner, E.; Born, R.; Galli, R.; Stelling, A.L.; Sivkov, V.N.; Petrova, O.V.; et al. Isolation and identification of chitin in the black coral Parantipathes larix (Anthozoa: Cnidaria). Int. J. Biol. Macromol. 2012, 51, 129–137. (In English) [Google Scholar] [CrossRef]
- Nowacki, K.; Stępniak, I.; Langer, E.; Tsurkan, M.; Wysokowski, M.; Petrenko, I.; Khrunyk, Y.; Fursov, A.; Bo, M.; Bavestrello, G.; et al. Electrochemical Approach for Isolation of Chitin from the Skeleton of the Black Coral Cirrhipathes sp. (Antipatharia). Mar. Drugs 2020, 18, 297. (In English) [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.A.; Halfar, J. First evidence of chitin in calcified coralline algae: New insights into the calcification process of Clathromorphum compactum. Sci. Rep. 2014, 4, 6162. [Google Scholar] [CrossRef]
- Baharlouei, P.; Rahman, A. Chitin and Chitosan: Prospective Biomedical Applications in Drug Delivery, Cancer Treatment, and Wound Healing. Mar. Drugs 2022, 20, 460. (In English) [Google Scholar] [CrossRef]
- Machałowski, T.; Wysokowski, M.; Żółtowska-Aksamitowska, S.; Bechmann, N.; Binnewerg, B.; Schubert, M.; Guan, K.; Bornstein, S.R.; Czaczyk, K.; Pokrovsky, O.; et al. Spider Chitin. The biomimetic potential and applications of Caribena versicolor tubular chitin. Carbohydr. Polym. 2019, 226, 115301. [Google Scholar] [CrossRef] [PubMed]
- Machałowski, T.; Wysokowski, M.; Tsurkan, M.V.; Galli, R.; Schimpf, C.; Rafaja, D.; Brendler, E.; Viehweger, C.; Żółtowska-Aksamitowska, S.; Petrenko, I.; et al. Spider Chitin: An Ultrafast Microwave-Assisted Method for Chitin Isolation from Caribena versicolor Spider Molt Cuticle. Molecules 2019, 24, 3736. [Google Scholar] [CrossRef]
- Ehrlich, H.; Ilan, M.; Maldonado, M.; Muricy, G.; Bavestrello, G.; Kljajic, Z.; Carballo, J.L.; Schiaparelli, S.; Ereskovsky, A.; Schupp, P.; et al. Three-dimensional chitin-based scaffolds from Verongida sponges (Demospongiae: Porifera). Part I. Isolation and identification of chitin. Int. J. Biol. Macromol. 2010, 47, 132–140. [Google Scholar] [CrossRef]
- Oliver, W.C.; Pharr, G.M. An improved technique for determining hardness and elastic modulus using load and displacement sensing indentation experiments. J. Mater. Res. 1992, 7, 1564–1583. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biomaterial | Elastic Modulus (MPa) | Ultimate Tensile Strength (MPa) | Source | |
|---|---|---|---|---|
| Chitin | Bush crickets’ acoustic tracheae | 5200 | - | [29] |
| Sheep crab exoskeleton (wet) | 518 ± 72 | 31.5 ± 5.4 | [30] | |
| Sheep crab exoskeleton (dry) | 764 ± 83 | 12.9 ± 1.7 | ||
| Commercial flake chitin films (solvent-casting) | 1240–3650 | 38–60 | [28] | |
| Philippine blue swimming crab chitin (solvent-casting) | - | 44.22 | [31] | |
| Human Tissues | Human skin (back) | 98.97 ± 97 | 27.2 ± 9.3 | [32] |
| Human femoral cancellous bone | 441 | 6.8 | [33] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duminis, T.; Heljak, M.; Święszkowski, W.; Ereskovsky, A.; Dziedzic, I.; Nowicki, M.; Pajewska-Szmyt, M.; Voronkina, A.; Bornstein, S.R.; Ehrlich, H. On the Mechanical Properties of Microfibre-Based 3D Chitinous Scaffolds from Selected Verongiida Sponges. Mar. Drugs 2023, 21, 463. https://doi.org/10.3390/md21090463
Duminis T, Heljak M, Święszkowski W, Ereskovsky A, Dziedzic I, Nowicki M, Pajewska-Szmyt M, Voronkina A, Bornstein SR, Ehrlich H. On the Mechanical Properties of Microfibre-Based 3D Chitinous Scaffolds from Selected Verongiida Sponges. Marine Drugs. 2023; 21(9):463. https://doi.org/10.3390/md21090463
Chicago/Turabian StyleDuminis, Tomas, Marcin Heljak, Wojciech Święszkowski, Alexander Ereskovsky, Izabela Dziedzic, Marek Nowicki, Martyna Pajewska-Szmyt, Alona Voronkina, Stefan R. Bornstein, and Hermann Ehrlich. 2023. "On the Mechanical Properties of Microfibre-Based 3D Chitinous Scaffolds from Selected Verongiida Sponges" Marine Drugs 21, no. 9: 463. https://doi.org/10.3390/md21090463
APA StyleDuminis, T., Heljak, M., Święszkowski, W., Ereskovsky, A., Dziedzic, I., Nowicki, M., Pajewska-Szmyt, M., Voronkina, A., Bornstein, S. R., & Ehrlich, H. (2023). On the Mechanical Properties of Microfibre-Based 3D Chitinous Scaffolds from Selected Verongiida Sponges. Marine Drugs, 21(9), 463. https://doi.org/10.3390/md21090463