

Effect of Iron Concentration on the Co-Production of Fucoxanthin and Fatty Acids in Conticribra weissflogii



Abstract
:1. Introduction
2. Results
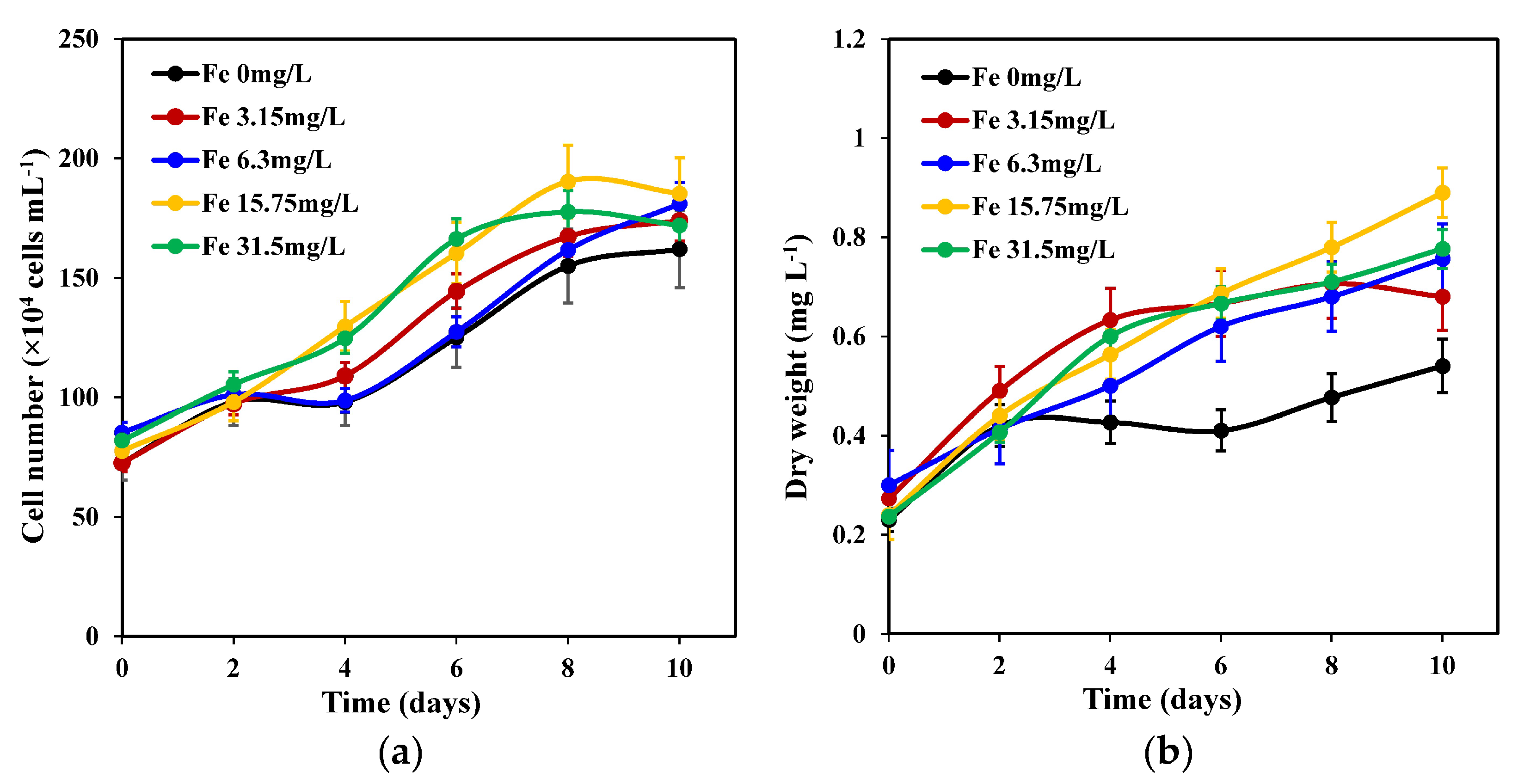
2.1. Growth of C. weissflogii under Different Iron Concentrations
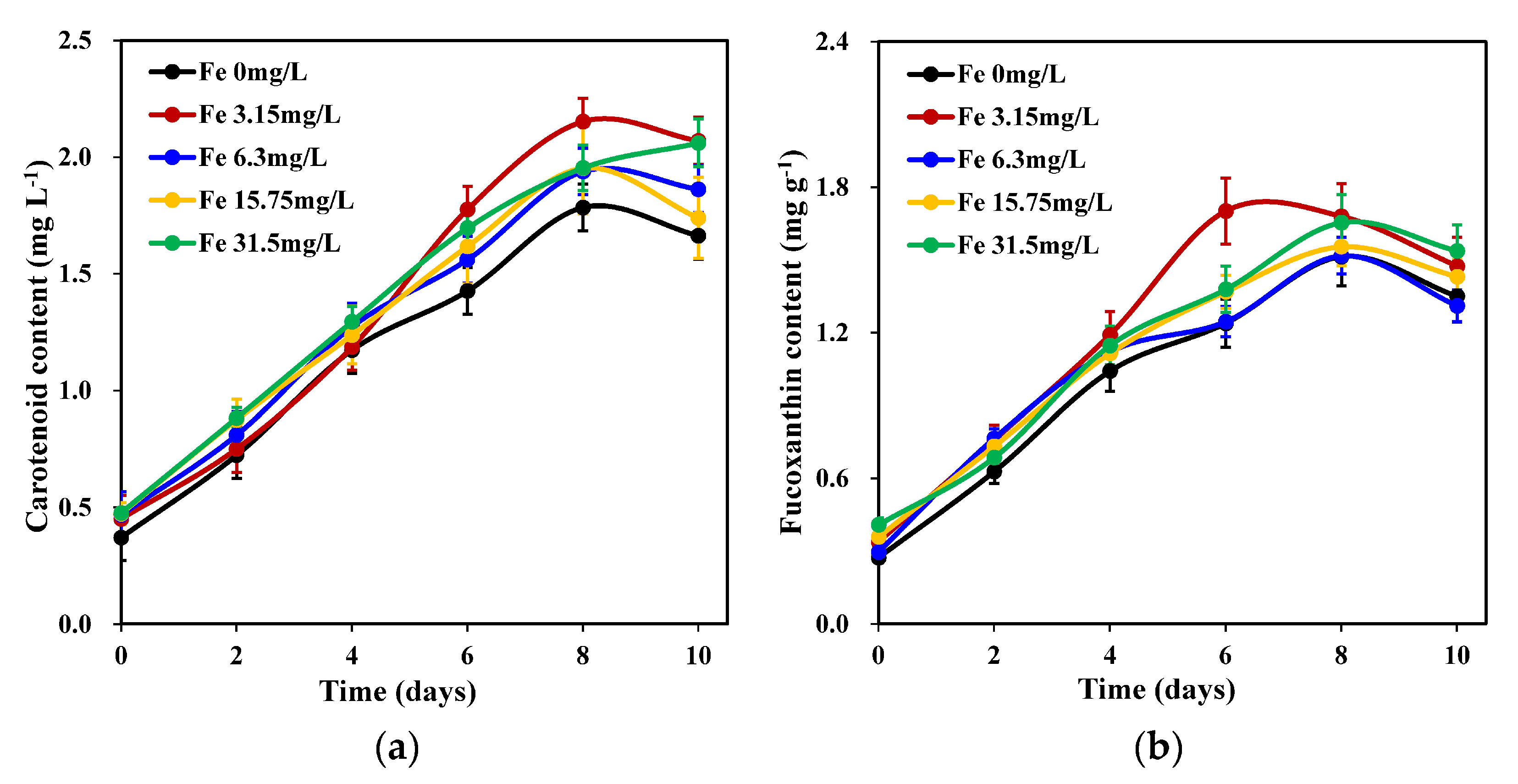
2.2. Changes in the Pigment Content of C. weissflogii under Different Iron Concentrations
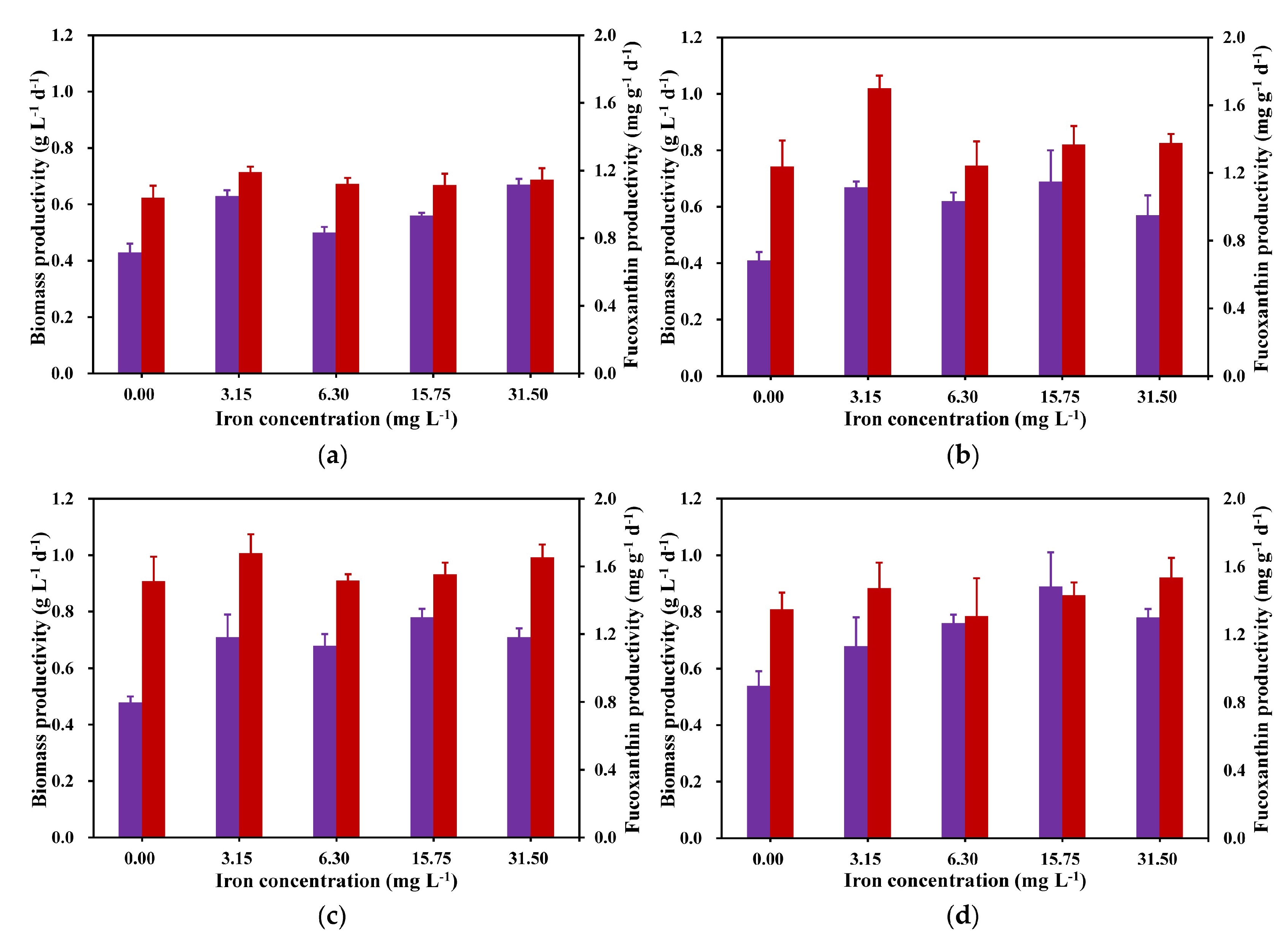
2.3. Effects of Different Iron Concentrations on C. weissflosi Biomass and Fucoxanthin Productivity
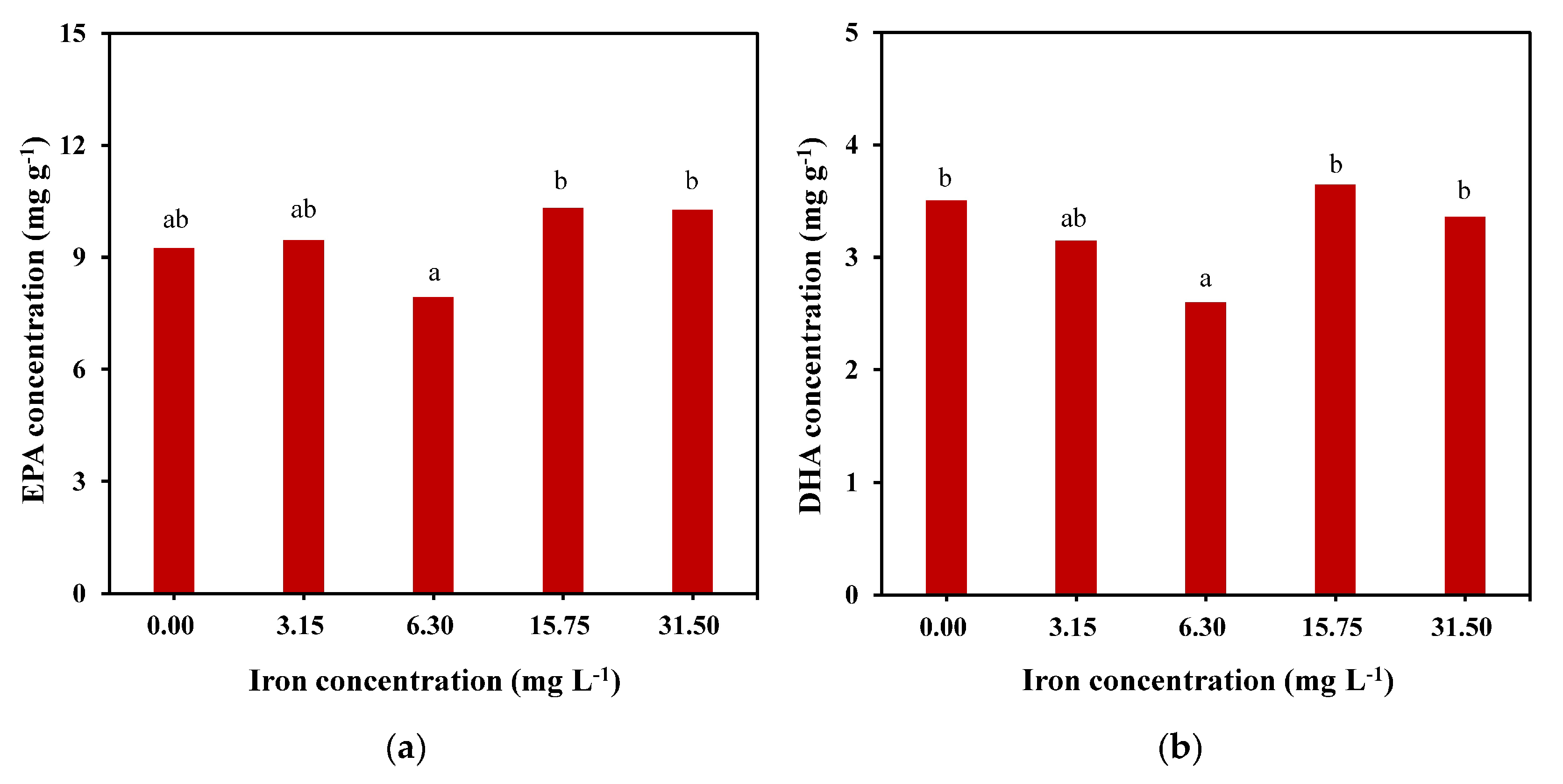
2.4. Fatty Acid Composition and Concentration of C. weissflosi at Different Iron Concentrations
3. Discussion
4. Materials and Methods
4.1. C. weissflogii Strain and Culture Conditions
4.2. Experimental Setup
4.3. Analytical Methods
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jónasdóttir, S.H. Fatty acid profiles and production in marine phytoplankton. Mar. Drugs 2019, 17, 151. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Li, K.; Duan, X.; Hill, D.; Barrow, C.; Dunshea, F.; Marin, G.; Suleria, H. Bioactive compounds in microalgae and their potential health benefits. Food Biosci. 2022, 49, 101932. [Google Scholar] [CrossRef]
- Khan, M.I.; Shin, J.H.; Kim, J.D. The promising future of microalgae: Current status, challenges, and optimization of a sustainable and renewable industry for biofuels, feed, and other products. Microb. Cell Fact. 2018, 17, 36. [Google Scholar] [CrossRef] [PubMed]
- Dolganyuk, V.; Belova, D.; Babich, O.; Prosekov, A.; Ivanova, S.; Katserov, D.; Patyukov, N.; Sukhikh, S. Microalgae: A promising source of valuable bioproducts. Biomolecules 2020, 10, 1153. [Google Scholar] [CrossRef] [PubMed]
- Ampofo, J.; Abbey, L. Microalgae: Bioactive composition, health benefits, safety, and prospects as potential high-value ingredients for the functional food industry. Foods 2022, 11, 1744. [Google Scholar] [CrossRef]
- Fernández, F.G.A.; Reis, A.; Wijffels, R.H.; Barbosa, M.; Verdelho, V.; Llamas, B. The role of microalgae in the bioeconomy. New Biotechnol. 2021, 61, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Yarnold, J.; Karan, H.; Oey, M.; Hankamer, B. Microalgal aquafeeds as part of a circular bioeconomy. Trends Plant Sci. 2019, 24, 959–970. [Google Scholar] [CrossRef]
- Maeda, H.; Hosokawa, M.; Sashima, T.; Miyashita, K. Dietary combination of fucoxanthin and fish oil attenuates the weight gain of white adipose tissue and decreases blood glucose in obese/diabetic KK-Ay mice. J. Agric. Food Chem. 2007, 55, 7701–7706. [Google Scholar] [CrossRef]
- Zhang, H.; Tang, Y.; Zhang, Y.; Zhang, S.; Qu, J.; Wang, X.; Kong, R.; Han, C.; Liu, Z. Fucoxanthin: A promising medicinal and nutritional ingredient. Evid. Based Complement. Altern. Med. 2015, 2015, 723515. [Google Scholar] [CrossRef]
- Yoong, K.L.; Chun-Yen, C.; Sunita, V.; Jo-Shu, C. Producing fucoxanthin from algae—Recent advances in cultivation strategies and downstream processing. Bioresour. Technol. 2022, 344, 126170. [Google Scholar]
- Pajot, A.; Hao Huynh, G.; Picot, L.; Marchal, L.; Nicolau, E. Fucoxanthin from algae to human, an extraordinary bioresource: Insights and advances in up and downstream processes. Mar. Drugs 2022, 20, 222. [Google Scholar] [CrossRef] [PubMed]
- Khaw, Y.S.; Yusoff, F.M.; Tan, H.T.; Noor Mazli, N.A.I.; Nazarudin, M.F.; Shaharuddin, N.A.; Omar, A.R.; Takahashi, K. Fucoxanthin production of microalgae under different culture factors: A systematic review. Mar. Drugs 2022, 20, 592. [Google Scholar] [CrossRef]
- Lu, Q.; Li, H.; Xiao, Y.; Liu, H. A state-of-the-art review on the synthetic mechanisms, production technologies, and practical application of polyunsaturated fatty acids from microalgae. Algal Res. 2021, 55, 102281. [Google Scholar] [CrossRef]
- Tyagi, R.; Rastogi, R.P.; Babich, O.; Awasthi, M.K.; Tiwari, A. New perspectives of omega-3 fatty acids from diatoms. Syst. Microbiol. Biomanuf 2023. [Google Scholar] [CrossRef]
- Rui, X.; Amenorfenyo, D.K.; Peng, K.; Li, H.; Wang, L.; Huang, X.; Li, C.; Li, F. Effects of different nitrogen concentrations on co-production of fucoxanthin and fatty acids in Conticribra weissflogii. Mar. Drugs 2023, 21, 106. [Google Scholar] [CrossRef]
- Marella, T.K.; Tiwari, A. Marine diatom Thalassiosira weissflogii based biorefinery for co-production of eicosapentaenoic acid and fucoxanthin. Bioresour. Technol. 2020, 307, 123245. [Google Scholar] [CrossRef] [PubMed]
- Ece, P.; Ebubekir, Y.; Mahmut, A. Effect of different iron sources on sustainable microalgae-based biodiesel production using Auxenochlorella protothecoides. Renew. Energy 2020, 162, 1970–1978. [Google Scholar]
- Rizwan, M.; Mujtaba, G.; Lee, K. Effects of iron sources on the growth and lipid/carbohydrate production of marine microalga Dunaliella tertiolecta. Biotechnol. Bioproc. Eng. 2017, 22, 68–75. [Google Scholar] [CrossRef]
- Terauchi, A.M.; Peers, G.; Kobayashi, M.C.; Niyogi, K.K.; Merchant, S.S. Trophic status of Chlamydomonas reinhardtii influences the impact of iron deficiency on photosynthesis. Photosynth. Res. 2010, 105, 39–49. [Google Scholar] [CrossRef]
- Yuan, X.; Liang, L.; Liu, K.; Xie, L.; Huang, L.; He, W.; Chen, Y.; Xue, T. Spent yeast as an efficient medium supplement for fucoxanthin and eicosapentaenoic acid (EPA) production by Phaeodactylum tricornutum. J. Appl. Phycol. 2020, 32, 59–69. [Google Scholar] [CrossRef]
- Sunda, W.; Huntsman, S. Interrelated influence of iron, light, and cell size on marine phytoplankton growth. Nature 1997, 390, 389–392. [Google Scholar] [CrossRef]
- Chen, M.; Dei, R.C.H.; Wang, W.X.; Guo, L. Marine diatom uptake of iron bound with natural colloids of different origins. Mar. Chem. 2003, 81, 177–189. [Google Scholar] [CrossRef]
- Naito, K.; Matsui, M.; Imai, I. Ability of marine eukaryotic red tide microalgae to utilize insoluble iron. Harmful Algae 2005, 4, 1021–1032. [Google Scholar] [CrossRef]
- Liu, Z.Y.; Wang, G.C.; Zhou, B.C. Effect of iron on growth and lipid accumulation in Chlorella vulgaris. Bioresour. Technol. 2008, 99, 4717–4722. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Jiang, X.; Li, Y.; Han, Q. Effects of nitrogen, phosphorus and iron on the growth, total lipid content and fatty acid composition of Phaeodactylum tricornutum mutant strain. Chin. J. Ecol. 2016, 35, 189–198. (In Chinese) [Google Scholar]
- Sajjadi, B.; Chen, W.Y.; Aziz, A.; Raman, A.; Ibrahim, S. Microalgae lipid and biomass for biofuel production: A comprehensive review on lipid enhancement strategies and their effects on fatty acid composition. Renew. Sustain. Energy Rev. 2018, 97, 200–232. [Google Scholar] [CrossRef]
- Zhou, H.; Hu, T.; Xuan, Y.; Zhou, J. Study on influence of typical microscale metal elements on growth of Microcystis aeruginosa. J. Water Resour. Water Eng. 2016, 27, 9–13. [Google Scholar]
- Mohsen, G.; Behzad, R.; Berat, Z.H. Effects of macro and micronutrients on neutral lipid accumulation in oleaginous microalgae. Biofuels 2018, 9, 147–156. [Google Scholar]
- Mohit, S.R.; Shashi, B.; Sudhakar, D.R.; Sanjeev, K.P. Effect of iron oxide nanoparticles on growth and biofuel potential of Chlorella spp. Algal Res. 2020, 49, 101942. [Google Scholar]
- Mohit, S.R.; Sanjeev, K.P. Resolving the dilemma of iron bioavailability to microalgae for commercial sustenance. Algal Res. 2021, 59, 102458. [Google Scholar]
- Wu, S.; Zhang, X.; Zheng, W.; Li, J. Optimization of culture media for spaceflight and non-spaceflight Navicula tenera. J. Plant Resour. Environ. 2006, 15, 33–37. (In Chinese) [Google Scholar]
- Sahin, M.S.; Khazi, M.I.; Demirel, Z.; Dalay, D.M. Variation in growth, fucoxanthin, fatty acids profile and lipid content of marine diatoms Nitzschia sp. and Nanofrustulum shiloi in response to nitrogen and iron. Biocatal. Agric. Biotechnol. 2019, 17, 390–398. [Google Scholar] [CrossRef]
- Zhu, M.; Mu, X.; Li, R.; Lü, R. The effects of iron on growth, photosynthesis, and biochemical composition of a marine algae Phaeodactylum tricornutum. Acta Oceanol. Sin. 2000, 22, 110–116. (In Chinese) [Google Scholar]
- Kosakowska, A.; Lewandowska, J.; Stoń, J.; Burkiewicz, K. Qualitative and quantitative composition of pigments in Phaeodactylum tricornutum (Bacillariophyceae) stressed by iron. Biometals 2004, 17, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Wei, D.; Zhang, X.C. New advances on fatty acid desaturase and regulation of these genes’ expression by ecological factors in microalgae cells. Mar. Sci. 2000, 24, 42–46. (In Chinese) [Google Scholar]
- Liang, J.; Jiang, X.; Zhang, Z.; Han, Q. Effects of phosphorus iron and silicon on the growth, total lipid content and fatty acid composition of Tropidoneis maxima. China Oils Fats 2016, 41, 69–74. (In Chinese) [Google Scholar]
- Jiang, H. Studies on Optimizing Production of Polyunsaturated Fatty Acids by Marine Microalgae. Master Dissertation, Shantou University, Guangdong, China, 2003. (In Chinese). [Google Scholar]
- Li, F.; Rui, X.; Amenorfenyo, D.K.; Pan, Y.; Huang, X.; Li, C. Effects of temperature, light, and salt on the production of fucoxanthin from Conticribra weissflogii. Mar. Drugs 2023, 21, 495. [Google Scholar] [CrossRef]
- Parsons, T.R.; Maita, Y.; Lalli, C.M. Determination of chlorophylls and total carotenoids: Spectrophotometric method—Science Direct. In A Manual of Chemical & Biological Methods for Seawater Analysis; Pergamon Press: Oxford, UK, 1984; pp. 101–104. [Google Scholar]
- Xu, R.R.; Gong, Y.F.; Chen, W.T.; Li, S.R.; Chen, R.Y.; Zheng, X.Y.; Chen, X.M.Z.; Wang, H.Y. Effects of LED monochromatic light quality of different colors on fucoxanthin content and expression levels of related genes in Phaeodactylum Tricornutum. Acta Opt. Sin. 2019, 9, 299–307. [Google Scholar]
- Zhang, W.Y.; Gao, B.Y.; Li, A.F.; Zhang, C.W. Effects of different culture conditions on growth and accumulation of bioactive components by Phaeodactylum tricornutum. Mar. Sci. 2016, 40, 57–65. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test Items | 0 mg/L | 3.15 mg/L | 6.3 mg/L | 15.75 mg/L | 31.5 mg/L |
|---|---|---|---|---|---|
| C4:0 | 18.13 | 19.57 | 45.97 | 15.83 | 43.11 |
| C6:0 | 30.33 | 137.49 | 30.65 | 23.48 | 102.16 |
| C8:0 | 263.9 | 112.91 | 119.47 | 130.3 | 269.79 |
| C10:0 | 384.5 | 25.29 | 254.38 | 310.88 | 326.63 |
| C11:0 | Not detected | 25.95 | Not detected | 35.37 | 53 |
| C12:0 | 48.36 | 45.24 | 34.13 | 42.32 | 40.47 |
| C13:0 | Not detected | 47 | Not detected | Not detected | Not detected |
| C14:0 | 8321.98 | 8272.94 | 7789.73 | 8754.77 | 8622.26 |
| C14:1n5 | 161.41 | 154.7 | 134.28 | 179.01 | 159.08 |
| C15:0 | 950.01 | 1120.68 | 1055.68 | 1224.67 | 1180.82 |
| C16:0 | 14,727.31 | 16,135.16 | 14,813.51 | 16,884.4 | 15,317.65 |
| C16:1n7 | 22,365.49 | 25,421.25 | 24,049.79 | 26,828.85 | 24,082.01 |
| C17:0 | 32.67 | 20.12 | 62.36 | 67.49 | 55.43 |
| C18:0 | 501.14 | 425.01 | 338.4 | 470.36 | 367.42 |
| C18:1n9c | 999.17 | 1013.63 | 864.56 | 1102.62 | 988.37 |
| C18:2n6c | 281.33 | 260.08 | 198.92 | 252.58 | 264.04 |
| C18:3n6 | Not detected | 170.72 | 60.06 | 20.83 | 27.57 |
| C18:3n3 | 119.37 | 78.65 | 76.48 | 91.64 | 78.98 |
| C20:0 | 57.75 | 69.59 | 92.43 | 126.45 | 85.36 |
| C20:2 | 64.61 | Not detected | 101.77 | Not detected | Not detected |
| C20:3n6 | 112.49 | Not detected | 66.83 | Not detected | Not detected |
| C21:0 | 134.28 | 121.97 | 98.69 | 112.28 | 138.49 |
| C20:3n3 | 126.25 | Not detected | 59.34 | 164.18 | 145.83 |
| C20:4n6 | 123.17 | 86.67 | 78.95 | 97.39 | 114.95 |
| C20:5n3 | 9254.11 | 9465.2 | 7936.26 | 10,328.37 | 10,280.99 |
| C22:0 | 119.21 | 107.57 | 79.92 | 118.49 | 108.39 |
| C22:6n3 | 3511.2 | 3146.65 | 2600.33 | 3647.48 | 3364.62 |
| C23:0 | 318.16 | Not detected | 447.67 | Not detected | Not detected |
| C24:0 | 1035.7 | 888.02 | 634.48 | 1025.92 | 967.25 |
| C24:1n9 | 167.1 | 37.87 | 93.67 | 146.45 | 28.88 |
| SFA | 38,745.22 | 27,574.51 | 25,897.47 | 40,458.73 | 27,678.23 |
| MUFA | 23,693.17 | 26,683.25 | 25,142.3 | 28,256.93 | 25,258.34 |
| PUFA | 13,592.53 | 13,152.17 | 11,178.94 | 14,602.47 | 14,276.98 |
| UFA | 37,285.7 | 39,835.42 | 36,321.24 | 42,859.4 | 39,535.32 |
| FA | 76,030.92 | 67,409.93 | 62,218.71 | 83,318.13 | 67,213.55 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, K.; Amenorfenyo, D.K.; Rui, X.; Huang, X.; Li, C.; Li, F. Effect of Iron Concentration on the Co-Production of Fucoxanthin and Fatty Acids in Conticribra weissflogii. Mar. Drugs 2024, 22, 106. https://doi.org/10.3390/md22030106
Peng K, Amenorfenyo DK, Rui X, Huang X, Li C, Li F. Effect of Iron Concentration on the Co-Production of Fucoxanthin and Fatty Acids in Conticribra weissflogii. Marine Drugs. 2024; 22(3):106. https://doi.org/10.3390/md22030106
Chicago/Turabian StylePeng, Ke, David Kwame Amenorfenyo, Xiangyu Rui, Xianghu Huang, Changling Li, and Feng Li. 2024. "Effect of Iron Concentration on the Co-Production of Fucoxanthin and Fatty Acids in Conticribra weissflogii" Marine Drugs 22, no. 3: 106. https://doi.org/10.3390/md22030106
APA StylePeng, K., Amenorfenyo, D. K., Rui, X., Huang, X., Li, C., & Li, F. (2024). Effect of Iron Concentration on the Co-Production of Fucoxanthin and Fatty Acids in Conticribra weissflogii. Marine Drugs, 22(3), 106. https://doi.org/10.3390/md22030106