
Exploring the Hypocholesterolemic Potential of a Fucus vesiculosus Extract: Omic Insights into Molecular Mechanisms at the Intestinal Level



Abstract
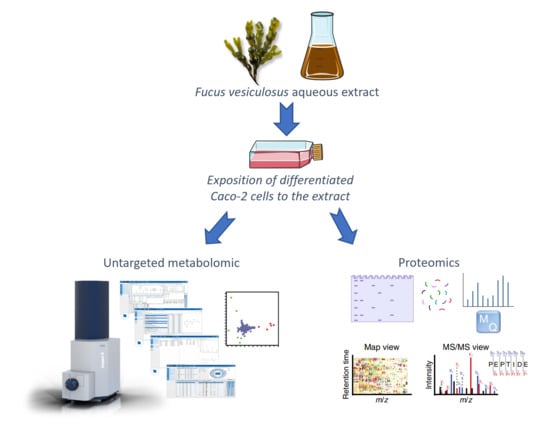
:
1. Introduction
2. Results and Discussion
2.1. Effect of the Purified F. vesiculosus Aqueous Extract in Enterocyte-Like Caco-2 Cells
2.1.1. Metabolomic Analysis
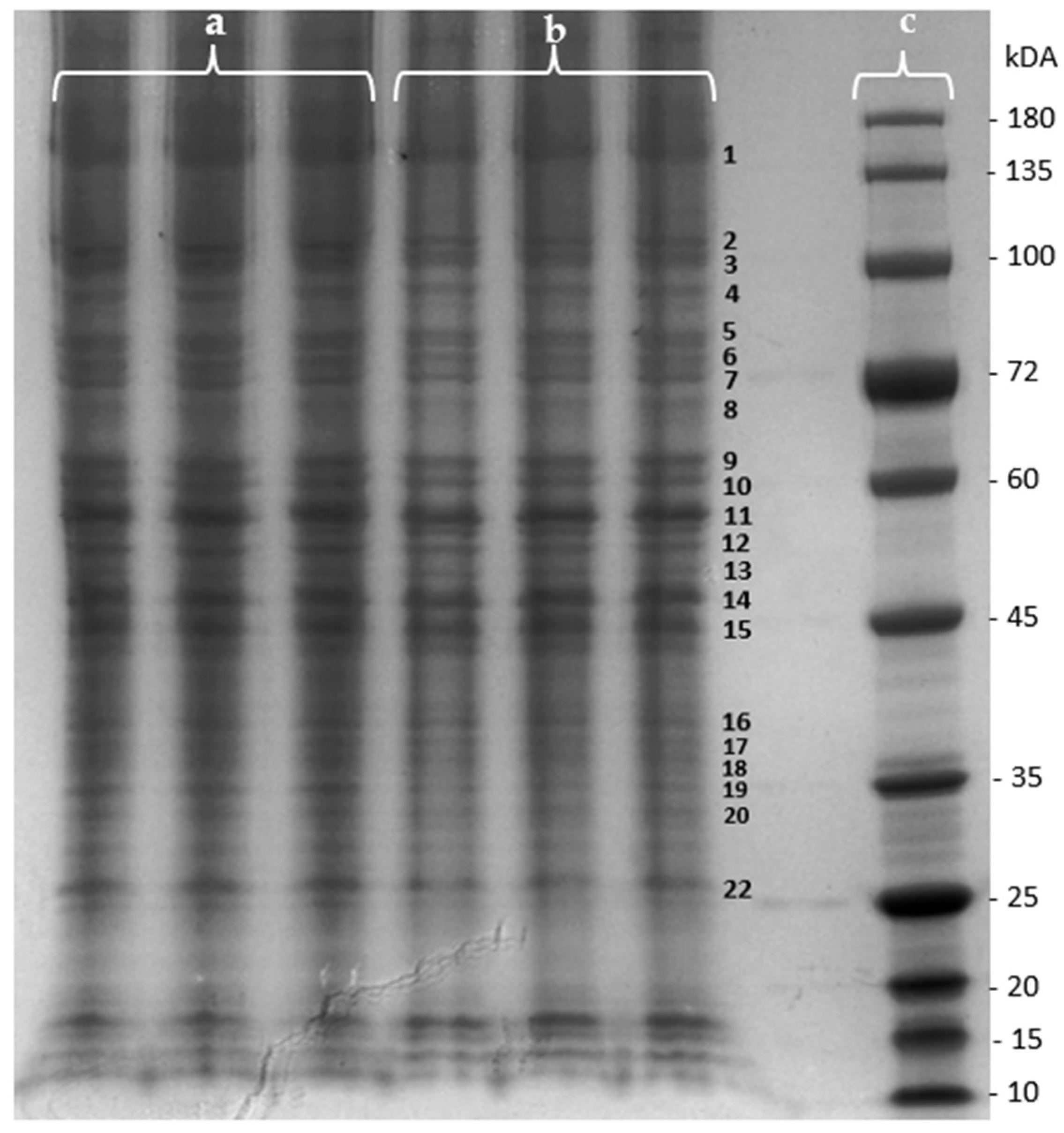
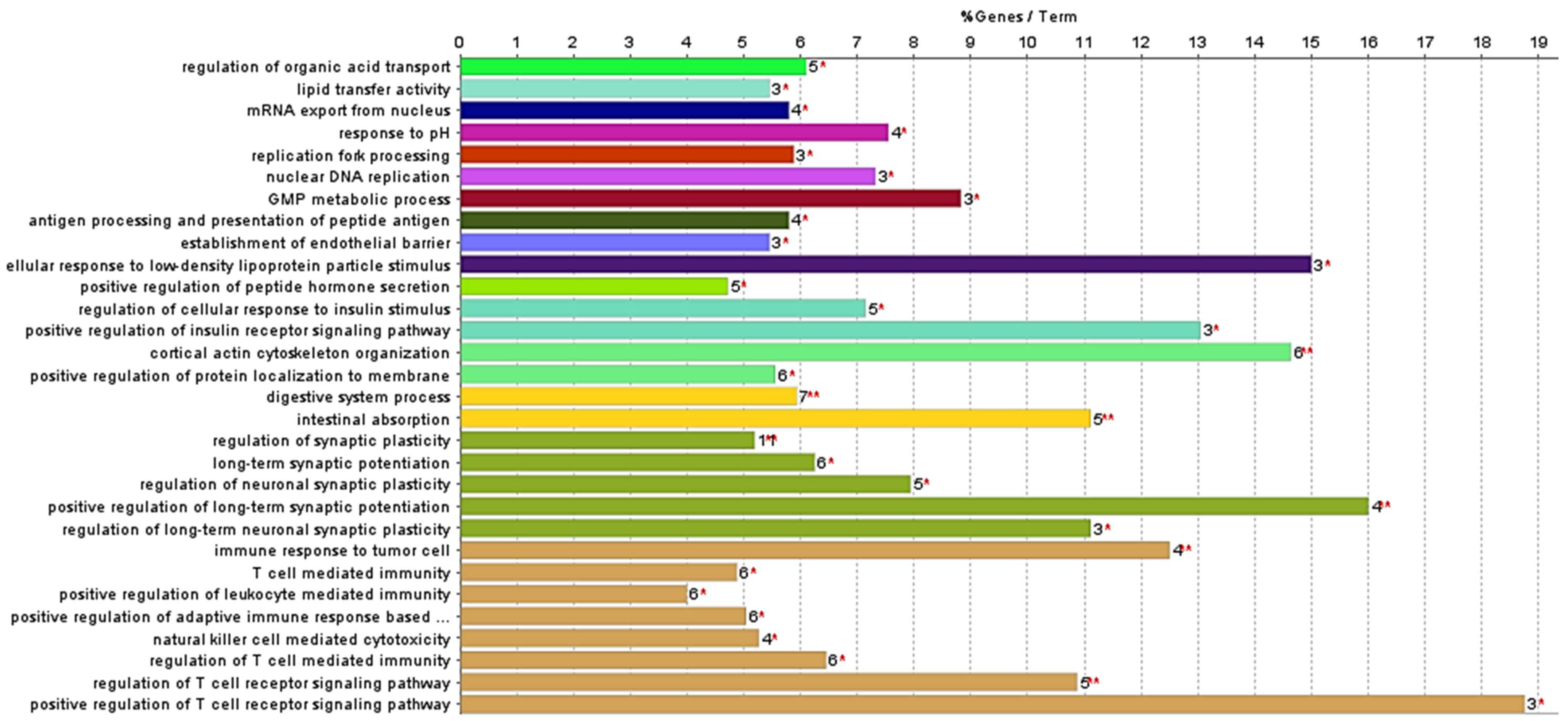
2.1.2. Proteomic Analysis
3. Materials and Methods
3.1. Chemicals
3.2. Preparation and Characterization of Algae Extract
3.3. Cell Culturing and Differentiation
3.4. Cytotoxicity Studies in Caco-2 Cells
3.5. Metabolomic Analysis through Liquid Chromatography Combined with High-Resolution Tandem Mass Spectrometry (LC/HRMS)
3.6. Membrane Protein Extract and One-Dimension Polyacrylamide Gel Electrophoresis (SDS-PAGE)
3.7. In-Gel Protein Digestion, Nano-LC−ESI−MS/MS, and Data Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Hansson, G.K.; Hermansson, A. The immune system in atherosclerosis. Nat. Immunol. 2011, 12, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Townsend, N.; Kazakiewicz, D.; Lucy Wright, F.; Timmis, A.; Huculeci, R.; Torbica, A.; Gale, C.P.; Achenbach, S.; Weidinger, F.; Vardas, P. Epidemiology of cardiovascular disease in Europe. Nat. Rev. Cardiol. 2022, 19, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, E.; Wilson, L.; Wickramasinghe, K.; Bhatnagar, P. European Cardiovascular Disease Statistics 2017; European Heart Network: Brussels, Belgium, 2017; pp. 94–100. [Google Scholar]
- Carson, J.A.S.; Lichtenstein, A.H.; Anderson, C.A.M.; Appel, L.J.; Kris-Etherton, P.M.; Meyer, K.A.; Petersen, K.; Polonsky, T.; Van Horn, L. Dietary Cholesterol and Cardiovascular Risk: A Science Advisory from the American Heart Association. Circulation 2020, 141, e39–e53. [Google Scholar] [CrossRef] [PubMed]
- André, R.; Pacheco, R.; Bourbon, M.; Serralheiro, M.L. Brown Algae Potential as a Functional Food against Hypercholesterolemia: Review. Foods 2021, 10, 234. [Google Scholar] [CrossRef] [PubMed]
- Aniket, K.; Roshan, D. Algae Products Market—Global Opportunity Analysis and Industry Forecast, 2022–2031; Allied Market Research: Pune, India, 2022. [Google Scholar]
- Ferruzza, S.; Rossi, C.; Scarino, M.L.; Sambuy, Y. A protocol for differentiation of human intestinal Caco-2 cells in asymmetric serum-containing medium. Toxicol. In Vitro 2012, 26, 1252–1255. [Google Scholar] [CrossRef] [PubMed]
- Lea, T. Caco-2 Cell Line BT—The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Verhoeckx, K., Cotter, P., López-Expósito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 103–111. ISBN 978-3-319-16104-4. [Google Scholar]
- Tung, Y.-T.; Wu, C.-H.; Chen, W.-C.; Pan, C.-H.; Chen, Y.-W.; Tsao, S.-P.; Chen, C.-J.; Huang, H.-Y. Ascophyllum nodosum and Fucus vesiculosus Extracts Improved Lipid Metabolism and Inflammation in High-Energy Diet-Induced Hyperlipidemia Rats. Nutrients 2022, 14, 4665. [Google Scholar] [CrossRef] [PubMed]
- André, R.; Guedes, L.; Melo, R.; Ascensão, L.; Pacheco, R.; Vaz, P.D.; Serralheiro, M.L. Effect of food preparations on in vitro bioactivities and chemical components of fucus vesiculosus. Foods 2020, 9, 955. [Google Scholar] [CrossRef] [PubMed]
- André, R.; Guedes, R.; López, J.; Serralheiro, M.L. Untargeted metabolomic study of HepG2 cells under the effect of Fucus vesiculosus aqueous extract. Rapid Commun. Mass Spectrom. 2021, 35, e9197. [Google Scholar] [CrossRef] [PubMed]
- Yoon, N.Y.; Kim, H.R.; Chung, H.Y.; Choi, J.S. Anti-hyperlipidemic effect of an edible brown algae, Ecklonia stolonifera, and its constituents on poloxamer 407-induced hyperlipidemic and cholesterol-fed rats. Arch. Pharm. Res. 2008, 31, 1564–1571. [Google Scholar] [CrossRef]
- André, R.; Pacheco, R.; Alves, A.C.; Santos, H.M.; Bourbon, M.; Serralheiro, M.L. The Hypocholesterolemic Potential of the Edible Algae Fucus vesiculosus: Proteomic and Quantitative PCR Analysis. Foods 2023, 12, 2758. [Google Scholar] [CrossRef]
- Laekeman, G. Assessment Report on Fucus vesiculosus L., Thallus; European Medicines Agency: Amsterdam, The Netherlands, 2014; Volume 44. [Google Scholar]
- Ososki, A.L.; Lohr, P.; Reiff, M.; Balick, M.J.; Kronenberg, F.; Fugh-Berman, A.; O’Connor, B. Ethnobotanical literature survey of medicinal plants in the Dominican Republic used for women’s health conditions. J. Ethnopharmacol. 2002, 79, 285–298. [Google Scholar] [CrossRef] [PubMed]
- Romm, A.; Hardy, M.L.; Mills, S. Endocrine Disorders and Adrenal Support. In Botanical Medicine for Women’s Health; Elsevier: St. Louis, MO, USA, 2017; pp. 186–210. [Google Scholar]
- Skibola, C.F. The effect of Fucus vesiculosus, an edible brown seaweed, upon menstrual cycle length and hormonal status in three pre-menopausal women: A case report. BMC Complement. Altern. Med. 2004, 4, 10. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, S.M.; Pereira, O.R.; Seca, A.M.L.; Pinto, D.C.G.A.; Silva, A.M.S. Seaweeds as Preventive Agents for Cardiovascular Diseases: From nutrients to functional foods. Mar. Drugs 2015, 13, 6838–6865. [Google Scholar] [CrossRef] [PubMed]
- Neto, R.; Marçal, C.; Queirós, A.; Abreu, H.; Silva, A.; Cardoso, S. Screening of Ulva rigida, Gracilaria sp., Fucus vesiculosus and Saccharina latissima as Functional Ingredients. Int. J. Mol. Sci. 2018, 19, 2987. [Google Scholar] [CrossRef] [PubMed]
- Catarino, M.; Silva, A.; Cardoso, S. Phycochemical Constituents and Biological Activities of Fucus spp. Mar. Drugs 2018, 16, 249. [Google Scholar] [CrossRef] [PubMed]
- Catarino, M.D.; Fernandes, I.; Oliveira, H.; Carrascal, M.; Ferreira, R.; Silva, A.M.S.; Cruz, M.T.; Mateus, N.; Cardoso, S.M. Antitumor Activity of Fucus vesiculosus-Derived Phlorotannins through Activation of Apoptotic Signals in Gastric and Colorectal Tumor Cell Lines. Int. J. Mol. Sci. 2021, 22, 7604. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Rubio, M.E.; Pérez-Jiménez, J.; Saura-Calixto, F. Dietary fiber and antioxidant capacity in Fucus vesiculosus products. Int. J. Food Sci. Nutr. 2009, 60, 23–34. [Google Scholar] [CrossRef]
- Jun, J.-Y.; Jung, M.-J.; Jeong, I.-H.; Yamazaki, K.; Kawai, Y.; Kim, B.-M. Antimicrobial and Antibiofilm Activities of Sulfated Polysaccharides from Marine Algae against Dental Plaque Bacteria. Mar. Drugs 2018, 16, 301. [Google Scholar] [CrossRef]
- van der Wulp, M.Y.M.; Verkade, H.J.; Groen, A.K. Regulation of cholesterol homeostasis. Mol. Cell. Endocrinol. 2013, 368, 1–16. [Google Scholar] [CrossRef]
- Shannon, E.; Conlon, M.; Hayes, M. Seaweed Components as Potential Modulators of the Gut Microbiota. Mar. Drugs 2021, 19, 358. [Google Scholar] [CrossRef]
- Schumacher-Schuh, A.; Bieger, A.; Borelli, W.V.; Portley, M.K.; Awad, P.S.; Bandres-Ciga, S. Advances in Proteomic and Metabolomic Profiling of Neurodegenerative Diseases. Front. Neurol. 2021, 12, 792227. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-Z.; Gerszten, R.E. Metabolomics and Proteomics in Type 2 Diabetes. Circ. Res. 2020, 126, 1613–1627. [Google Scholar] [CrossRef] [PubMed]
- Panner Selvam, M.K.; Finelli, R.; Agarwal, A.; Henkel, R. Proteomics and metabolomics—Current and future perspectives in clinical andrology. Andrologia 2021, 53, e13711. [Google Scholar] [CrossRef] [PubMed]
- Duarte, M.M.M.F.; Moresco, R.N.; Duarte, T.; Santi, A.; Bagatini, M.D.; Da Cruz, I.B.M.; Schetinger, M.R.C.; Loro, V.L. Oxidative stress in hypercholesterolemia and its association with Ala16Val superoxide dismutase gene polymorphism. Clin. Biochem. 2010, 43, 1118–1123. [Google Scholar] [CrossRef] [PubMed]
- Niu, B.; Liao, K.; Zhou, Y.; Wen, T.; Quan, G.; Pan, X.; Wu, C. Application of glutathione depletion in cancer therapy: Enhanced ROS-based therapy, ferroptosis, and chemotherapy. Biomaterials 2021, 277, 121110. [Google Scholar] [CrossRef] [PubMed]
- Yoo, D.; Jung, E.; Noh, J.; Hyun, H.; Seon, S.; Hong, S.; Kim, D.; Lee, D. Glutathione-Depleting Pro-Oxidant as a Selective Anticancer Therapeutic Agent. ACS Omega 2019, 4, 10070–10077. [Google Scholar] [CrossRef] [PubMed]
- Dang, H.T.; Kang, G.J.; Yoo, E.S.; Hong, J.; Choi, J.S.; Kim, H.S.; Chung, H.Y.; Jung, J.H. Evaluation of endogenous fatty acid amides and their synthetic analogues as potential anti-inflammatory leads. Bioorg. Med. Chem. 2011, 19, 1520–1527. [Google Scholar] [CrossRef] [PubMed]
- Bagheri, S.; Zolghadri, S.; Stanek, A. Beneficial Effects of Anti-Inflammatory Diet in Modulating Gut Microbiota and Controlling Obesity. Nutrients 2022, 14, 3985. [Google Scholar] [CrossRef] [PubMed]
- Tsoupras, A.; Lordan, R.; O’Keefe, E.; Shiels, K.; Saha, S.K.; Zabetakis, I. Structural Elucidation of Irish Ale Bioactive Polar Lipids with Antithrombotic Properties. Biomolecules 2020, 10, 1075. [Google Scholar] [CrossRef]
- Jia, L.; Betters, J.L.; Yu, L. Niemann-Pick C1-Like 1 (NPC1L1) Protein in Intestinal and Hepatic Cholesterol Transport. Annu. Rev. Physiol. 2011, 73, 239–259. [Google Scholar] [CrossRef]
- Jóźwiak, M.; Filipowska, A.; Fiorino, F.; Struga, M. Anticancer activities of fatty acids and their heterocyclic derivatives. Eur. J. Pharmacol. 2020, 871, 172937. [Google Scholar] [CrossRef] [PubMed]
- Fibulin-1 (FIBL-1)—Function. Available online: https://www.nextprot.org/entry/NX_P23142/ (accessed on 17 April 2024).
- Sushi Domain-Containing Protein 2 (SUSD2)—Function. Available online: https://www.nextprot.org/entry/NX_Q9UGT4/ (accessed on 17 April 2024).
- FABP1—Fatty Acid-Binding Protein, Liver—Function. Available online: https://www.nextprot.org/entry/NX_P07148/ (accessed on 4 May 2023).
- Zhang, J.R.; Coleman, T.; Langmade, S.J.; Scherrer, D.E.; Lane, L.; Lanier, M.H.; Feng, C.; Sands, M.S.; Schaffer, J.E.; Semenkovich, C.F.; et al. Niemann-Pick C1 protects against atherosclerosis in mice via regulation of macrophage intracellular cholesterol trafficking. J. Clin. Investig. 2008, 118, 2281–2290. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.H.; Jiang, N.; Yao, P.B.; Zheng, X.L.; Cayabyab, F.S.; Tang, C.K. NPC1, intracellular cholesterol trafficking and atherosclerosis. Clin. Chim. Acta 2014, 429, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Marques, L.R.; Diniz, T.A.; Antunes, B.M.; Rossi, F.E.; Caperuto, E.C.; Lira, F.S.; Gonçalves, D.C. Reverse cholesterol transport: Molecular mechanisms and the non-medical approach to enhance HDL cholesterol. Front. Physiol. 2018, 9, 526. [Google Scholar] [CrossRef]
- Shin, H.-C.; Kim, S.H.; Park, Y.; Lee, B.H.; Hwang, H.J. Effects of 12-week oral supplementation of Ecklonia cava polyphenols on anthropometric and blood lipid parameters in overweight Korean individuals: A double-blind randomized clinical trial. Phytother. Res. 2012, 26, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Yeo, A.-R.; Lee, J.; Tae, I.H.; Park, S.-R.; Cho, Y.H.; Lee, B.H.; Shin, H.C.; Kim, S.H.; Yoo, Y.C. Anti-hyperlipidemic Effect of Polyphenol Extract (Seapolynol(TM)) and Dieckol Isolated from Ecklonia cava in in vivo and in vitro Models. Prev. Nutr. Food Sci. 2012, 17, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Falé, P.; Amaral, F.; Amorim Madeira, P.J.; Sousa Silva, M.; Florêncio, M.H.; Frazão, F.N.; Serralheiro, M.L.M. Acetylcholinesterase inhibition, antioxidant activity and toxicity of Peumus boldus water extracts on HeLa and Caco-2 cell lines. Food Chem. Toxicol. 2012, 50, 2656–2662. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, E.; Araújo, J.E.; Gómez-Meire, S.; Lodeiro, C.; Perez-Melon, C.; Iglesias-Lamas, E.; Otero-Glez, A.; Capelo, J.L.; Santos, H.M. Proteomics analysis of the peritoneal dialysate effluent reveals the presence of calcium-regulation proteins and acute inflammatory response. Clin. Proteom. 2014, 11, 17. [Google Scholar] [CrossRef] [PubMed]
- Jorge, S.; Capelo, J.L.; LaFramboise, W.; Dhir, R.; Lodeiro, C.; Santos, H.M. Development of a Robust Ultrasonic-Based Sample Treatment to Unravel the Proteome of OCT-Embedded Solid Tumor Biopsies. J. Proteome Res. 2019, 18, 2979–2986. [Google Scholar] [CrossRef]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef]
- Cox, J.; Neuhauser, N.; Michalski, A.; Scheltema, R.A.; Olsen, J.V.; Mann, M. Andromeda: A Peptide Search Engine Integrated into the MaxQuant Environment. J. Proteome Res. 2011, 10, 1794–1805. [Google Scholar] [CrossRef] [PubMed]
- Tyanova, S.; Cox, J. Perseus: A Bioinformatics Platform for Integrative Analysis of Proteomics Data in Cancer Research. In Methods in Molecular Biology; von Stechow, L., Ed.; Springer: New York, NY, USA, 2018; pp. 133–148. ISBN 978-1-4939-7493-1. [Google Scholar]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.-H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Rt | [M − H]+ m/z | Molecular Formula | Δ m/z (Da) | p-Value | FC (Extract/Control) |
|---|---|---|---|---|---|---|
| Heptadec-2-enamide * | 17.54 | 268.2636 | C17H33NO | 0.0001 | 1.24 × 10−3 | 1.91 |
| (Z)-2-ketoctadec-9-enamide * | 14.57 | 296.2583 | C18H33NO2 | −0.00008 | 0.50 × 10−4 | 1.78 |
| Linoleic acid hydroxamate * | 13.11 | 278.2478 | C18H33NO2 | −0.00008 | 8.42 × 10−3 | 1.64 |
| Choline * | 1.12 | 104.10703 | C5H13NO | −0.16 | 1.50 × 10−5 | 1.61 |
| 3-Hetosphingosine * | 15.47 | 298.2739 | C18H35NO2 | −0.00010 | 1.05 × 10−3 | 1.59 |
| Palmitoleoyl ethanolamide * | 13.62 | 280.26334 | C18H35NO2 | −0.00006 | 0.1 × 10−5 | 1.54 |
| Myristamide * | 16.28 | 228.23232 | C14H29NO | 0.00012 | 2.5 × 10−5 | 1.49 |
| Linoleamide * | 14.33 | 280.2633 | C18H33NO | −0.00001 | 1.73 × 10−4 | 1.47 |
| Dodecanamide * | 14.13 | 200.2008 | C12H25NO | 0.00013 | 4.68 × 10−4 | 1.40 |
| Glutathione oxidized * | 1.92 | 613.1532 | C20H32N6O12S2 | 1.61 | 2.35 × 10−3 | 1.35 |
| C16 phytosphingosine * | 11.47 | 290.2691 | C16H35NO3 | −1.07 | 0.10 × 10−5 | 1.22 |
| Glutathione | 1.94 | 308.0908 | C10H17N3O6S | 0.0016 | 1.17 × 10−3 | −50.45 |
| Band | Fold Change (Extract/Control) | p-Value |
|---|---|---|
| 1 | −0.051 | 0.306 |
| 2 | 0.157 | 0.369 |
| 3 | −0.715 | 0.009 |
| 4 | −0.236 | 3.26 × 10−4 |
| 5 | −0.198 | 0.057 |
| 6 | −0.044 | 0.390 |
| 7 | −0.393 | 0.011 |
| 8 | −0.357 | 0.044 |
| 9 | −0.527 | 1.18 × 10−4 |
| 10 | −0.415 | 0.001 |
| 11 | 0.024 | 0.412 |
| 12 | 0.146 | 0.231 |
| 13 | −0.362 | 0.008 |
| 14 | −0.317 | 0.002 |
| 15 | −0.158 | 0.124 |
| 16 | −0.065 | 0.349 |
| 17 | −0.099 | 0.273 |
| 18 | −0.359 | 0.005 |
| 19 | 1.382 | 4.97 × 10−6 |
| 20 | 0.028 | 0.334 |
| 21 | 0.040 | 0.232 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
André, R.; Pacheco, R.; Santos, H.M.; Serralheiro, M.L. Exploring the Hypocholesterolemic Potential of a Fucus vesiculosus Extract: Omic Insights into Molecular Mechanisms at the Intestinal Level. Mar. Drugs 2024, 22, 187. https://doi.org/10.3390/md22040187
André R, Pacheco R, Santos HM, Serralheiro ML. Exploring the Hypocholesterolemic Potential of a Fucus vesiculosus Extract: Omic Insights into Molecular Mechanisms at the Intestinal Level. Marine Drugs. 2024; 22(4):187. https://doi.org/10.3390/md22040187
Chicago/Turabian StyleAndré, Rebeca, Rita Pacheco, Hugo M. Santos, and Maria Luísa Serralheiro. 2024. "Exploring the Hypocholesterolemic Potential of a Fucus vesiculosus Extract: Omic Insights into Molecular Mechanisms at the Intestinal Level" Marine Drugs 22, no. 4: 187. https://doi.org/10.3390/md22040187
APA StyleAndré, R., Pacheco, R., Santos, H. M., & Serralheiro, M. L. (2024). Exploring the Hypocholesterolemic Potential of a Fucus vesiculosus Extract: Omic Insights into Molecular Mechanisms at the Intestinal Level. Marine Drugs, 22(4), 187. https://doi.org/10.3390/md22040187