The Occurrence of Bioactive Micromonosporae in Aquatic Habitats of the Sunshine Coast in Australia


Abstract
:Introduction
Results
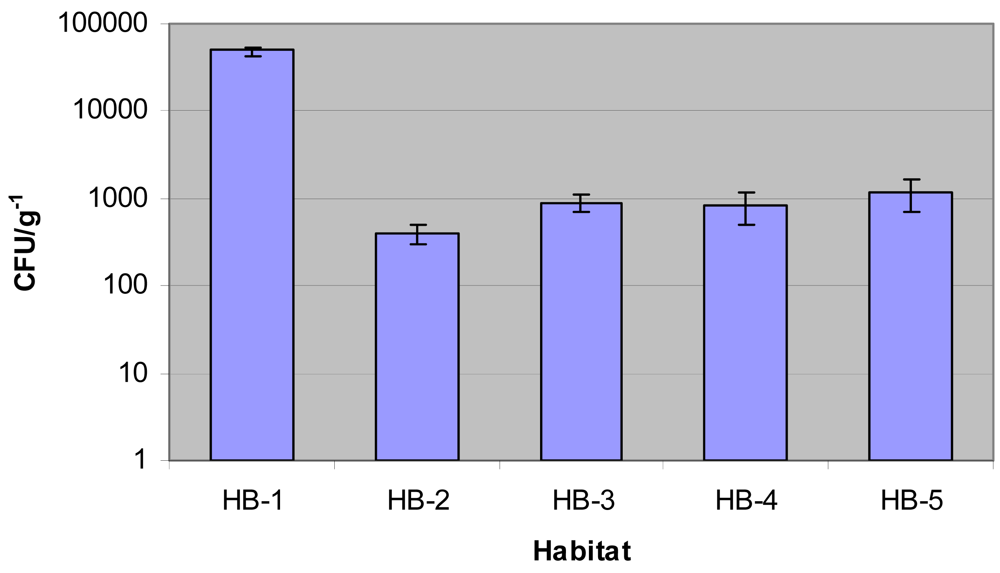
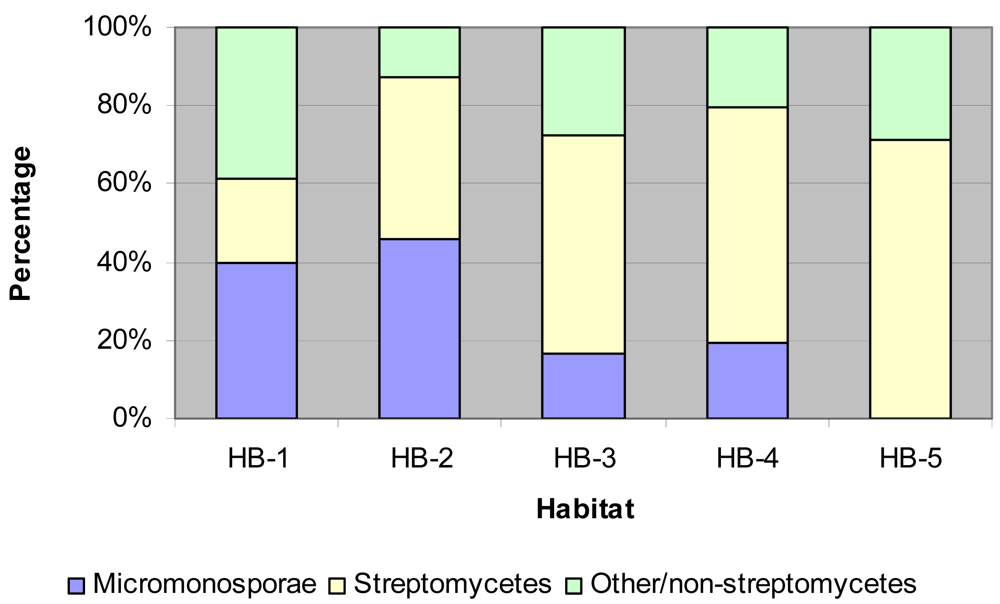
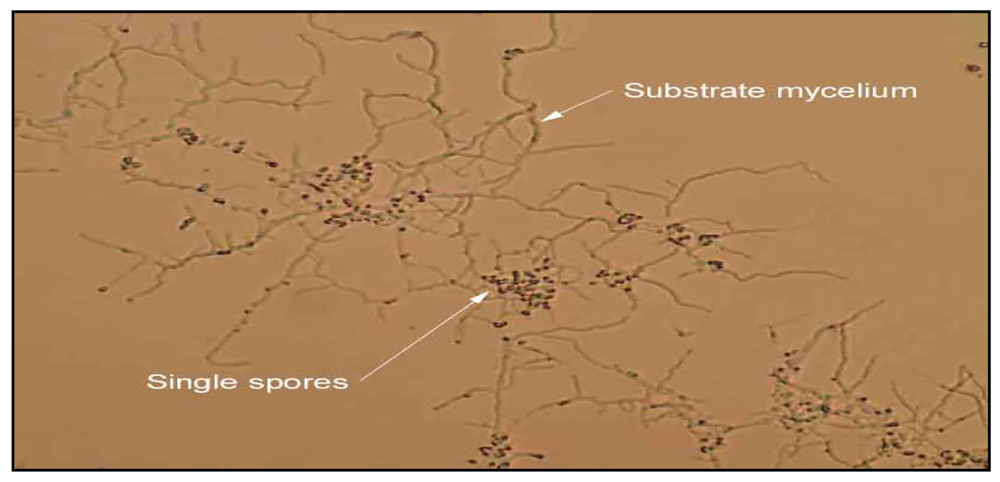
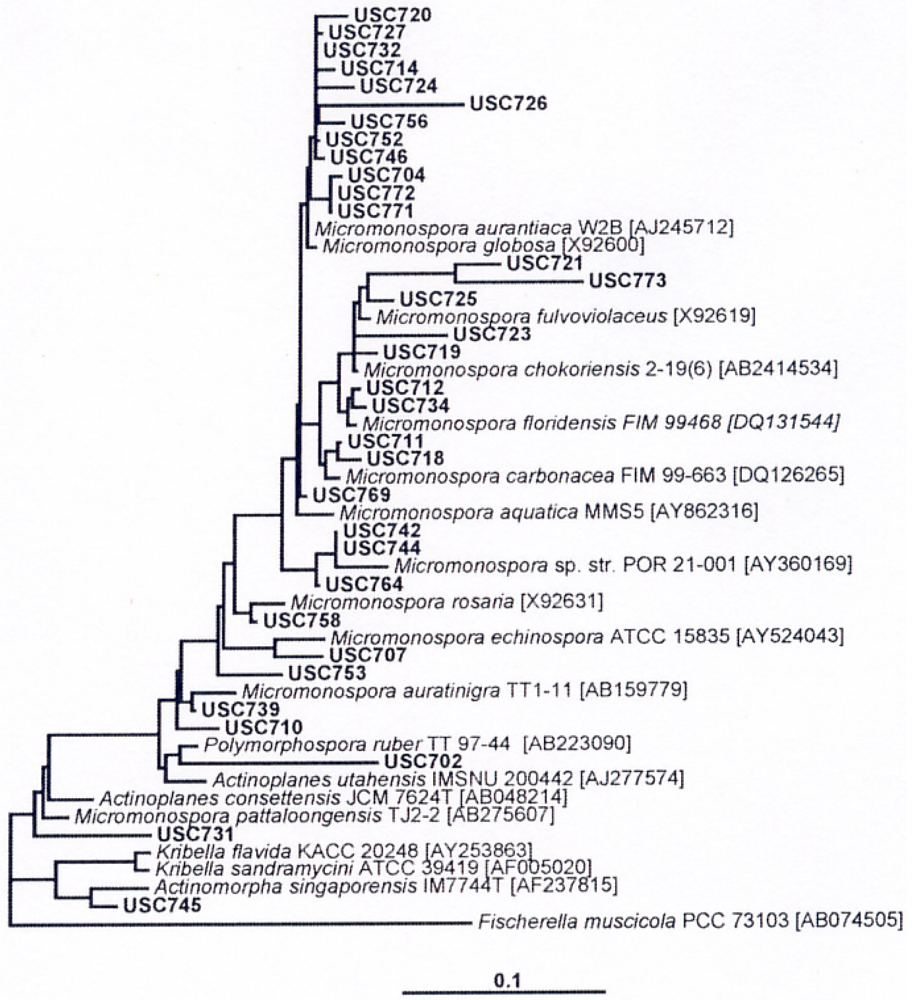
Isolation and identification of Micromonosporae
Testing for antimicrobial activity
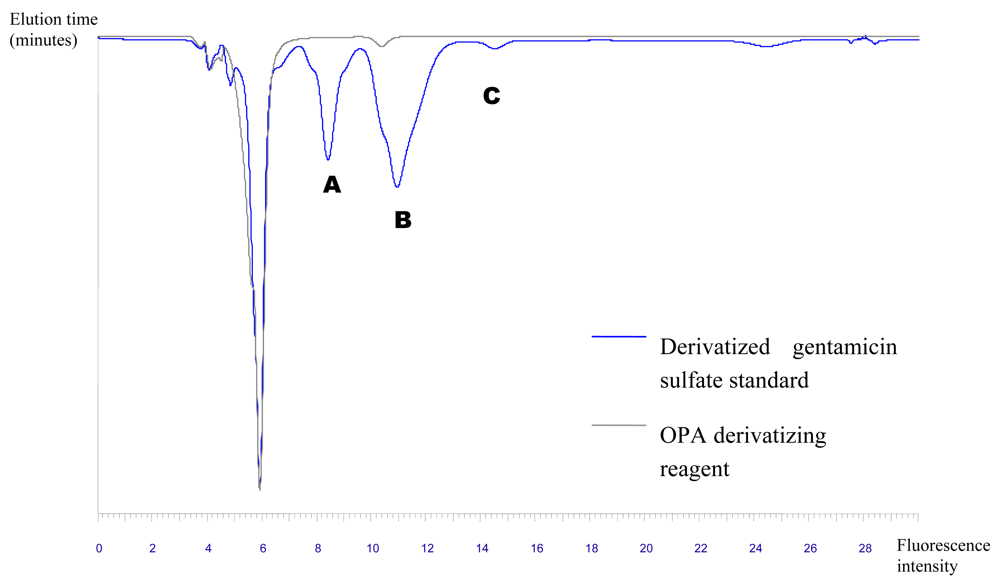
Chemical characterization of the bioactive compounds produced by the isolates
Discussion
Experimental Section
Sampling Sites
Selective Isolation of Micromonosporae
Identification of the isolates
Testing for antimicrobial activity
Chemical characterization of the bioactive compounds
Testing of crude extracts for retention of antimicrobial activity
HPLC analysis of crude extracts
Acknowledgements
References
- Ashton, EC; Hogarth, PJ; Ormond, R. Breakdown of mangrove leaf litter in a managed mangrove forest in Peninsular Malaysia. Hydrobiologia 1999, 413, 77–88. [Google Scholar]
- Ara, I; Kudo, T; Matsumoto, A; Takahashi, Y; Omura, S. Nonomurae maheshkhaliensis sp., nov., a novel actinomycete isolated from mangrove rhizosphere mud. J Gen Appl Microbiol 2007, 53, 159–166. [Google Scholar]
- Basik, M; Mousses, S; Trent, J. Integration of genomic technologies for accelerated cancer drug development. Biotechniques 2003, 35, 580–586. [Google Scholar]
- Barrow, RA. Kurtböke, DI, Swings, J, Eds.; Australian microfungi as producers of bioactive compounds for drug discovery. In Microbial Genetic Resources and Biodiscovery; Queensland Complete Printing Services: Nambour, Australia, 2004; pp. 133–149. [Google Scholar]
- Baker, JT; Bell, JD; Murphy, PT. Australian deliberations on access to its terrestrial and marine biodiversity. J Ethnopharmacol 1996, 15, 229–237. [Google Scholar]
- Barker, KF. Antibiotic resistance: a current perspective. Brit J Clin Pharmaco 1999, 48, 109–124. [Google Scholar]
- Berdy, J. Bioactive microbial metabolites. J Antibiot 2005, 58, 1–26. [Google Scholar]
- Bibikova, MV; Ivanitskaia, LP; Singal, EM. Directed screening of aminoglycoside antibiotic producers on selective media with gentamicin. Antibiotiki 1981, 26, 488–92. [Google Scholar]
- Bredholt, H; Fjærvik, E; Johnsen, G; Zotchev, SB. Actinomycetes from sediments in the Trondheim Fjord, Norway: Diversity and biological activity. Mar Drugs 2008, 6, 12–24. [Google Scholar]
- Boon, PI. Relationships between actinomycete populations and organic matter degradation in Lake Mulwala, Southeastern Australia. Regul River 1989, 4, 409–418. [Google Scholar]
- Bourne, DG; Evans-Illidge, E; Llewellyn, LE. Kurtböke, DI, Swings, J, Eds.; Marine microbes for biodiscovery: Just the tip of an iceberg. In Microbial Genetic Resources and Biodiscovery; Queensland Complete Printing Services: Nambour, Australia, 2004; pp. 185–205. [Google Scholar]
- Bulina, TI; Alferova, IV; Terekhova, LP. A novel approach to isolation of actinomycetes involving irradiation of soil samples with microwaves. Microbiology-USSR 1997, 66, 231–234. [Google Scholar]Bull, AT; Stach, JE. Marine actinobacteria: new opportunities for natural product search and discovery. Trends Microbiol 2007, 15, 491–499. [Google Scholar]
- Cross, T. Aquatic actinomycetes: A critical survey of the occurrence, growth and role of actinomycetes in aquatic habitats. J Appl Bacteriol 1981, 50, 397–423. [Google Scholar]
- Cross, T. Williams, ST, Sharpe, ME, Holt, JG, Eds.; The Actinomycetes II: Growth and examination of actinomycetes - some guidelines. In Bergey's Manual of Systematic Bacteriology; Williams and Wilkins: Baltimore, USA, 1989; pp. 4–2340. 2343. [Google Scholar]
- Demain, AL. From natural products discovery to commercialization: a success story. J Ind Microbiol Biotech 2006, 33, 486–495. [Google Scholar]
- Ensign, JC; Normand, P; Burden, JP; Yallop, CA. Physiology of some actinomycete genera. Res Microbiol 1993, 144, 657–660. [Google Scholar]
- Fenical, W; Jensen, PR. Developing a new resource for drug discovery: marine actinomycete bacteria. Nature Chem Biol 2006, 2, 666–673. [Google Scholar]
- Gause, GF; Maksimova, TS; Olkhovatova, OL. Schaal, KP, Pulverer, G, Eds.; Resistance of actinomycetes to their own antibiotics and its possible significance to ecology. In Actinomycetes: Proceedings from the 4th International Symposium on Actinomycete Biology; Gustav Fischer Verlag: Stuttgart, Germany, 1981; pp. 181–184. [Google Scholar]
- Gershon, D. Microarray technology: an array of opportunities. Nature 2002, 416, 885–891. [Google Scholar]
- Gontang, EA; Fenical, W; Jensen, PR. Phylogenetic diversity of Gram-positive bacteria cultured from marine sediments. Appl Environ Microb 2007, 73, 3272–3282. [Google Scholar]
- Goodfellow, M; Cross, T. Goodfellow, M, Mordarski, M, Williams, ST, Eds.; Classification. In The Biology of Actinomycetes; Academic Press: London, UK, 1984; pp. 7–164. [Google Scholar]
- Goodfellow, M; Haynes, JA. Ortiz-Ortiz, L, Bojalil, LF, Yakoleff, V, Eds.; Actinomycetes in marine sediments. In Biological, Biochemical and Biomedical Aspects of Actinomycetes; Academic Press, Inc.: Orlando, USA, 1984; pp. 453–472. [Google Scholar]
- Goodfellow, M; Williams, ST. Ecology of actinomycetes. Ann Rev Microbiol 1983, 37, 189–216. [Google Scholar]
- Gullo, VP; McAlpine, J; Lam, KS; Baker, D; Petersen, F. Drug discovery from natural products. J Ind Microbiol Biotechnol 2006, 33, 523–531. [Google Scholar]
- Hayakawa, M. Kurtböke, DI, Ed.; Selective isolation of rare actinomycete genera using pretreatment techniques. In Selective Isolation of Rare Actinomycetes; Queensland Complete Printing Services: Nambour, Australia, 2003; pp. 56–81. [Google Scholar]
- Hayakawa, M; Sadakata, T; Kajiura, T; Nonomura, H. New methods for the highly selective isolation of Micromonospora and Microbiospora from soil. J Ferment Bioeng 1991, 72, 320–326. [Google Scholar]
- Hopwood, D (Ed.) Streptomyces in Nature and Medicine: The Antibiotic Makers; Oxford University Press: Oxford, UK, 2007.
- Horan, AC. Gullo, VP, Ed.; Aerobic actinomycetes: sources of novel natural products. In The Discovery of Natural Products with Therapeutic Potential; Butterworth-Heinemann: Boston, USA, 1994; pp. 30–3. [Google Scholar]
- Horan, AC; Brodsky, BC. Micromonospora rosaria sp. nov., nom. rev., the rosaramicin producer. Int J Syst Evol Micr 1986, 36, 478–480. [Google Scholar]
- Hotta, K; Takahashi, A; Okami, Y; Umezawa, H. Relationship between antibiotic resistance and antibiotic productivity in actinomycetes which produce aminoglycoside antibiotics. J Antibiot 1983, 36, 1789–1791. [Google Scholar]
- Huang, H; Lv, J; Hu, Y; Fang, Z; Zhang, K; Bao, S. Micromonospora rifamycinica sp. nov., a novel actinomycete from mangrove sediment. Int J Syst Evol Micr 2008, 58, 17–20. [Google Scholar]
- Jensen, PR; Dwight, R; Fenical, W. Distribution of Actinomycetes in near-shore tropical marine sediments. Appl Environ Microb 1991, 57, 1102–1108. [Google Scholar]
- Jensen, PR; Gontag, E; Mafnas, C; Mincer, TJ; Fenical, W. Culturable marine actinomycete diversity from tropical Pacific Ocean sediments. Environ Microbiol 2005, 7, 1039–1048. [Google Scholar]
- Johnston, DW; Cross, T. The occurrence and distribution of actinomycetes in lakes of the English Lake District. Freshwater Biol 1976, 6, 457–463. [Google Scholar]
- Kim, TK; Garson, MJ; Fuerst, JA. Marine actinomycetes related to the “Salinispora” group from the Great Barrier Reef sponge Pseudoceratina clavata. Environ Microbiol 2005, 7, 509–518. [Google Scholar]
- Kurtböke, DI. Australian actinomycetes: An unexhausted source for biotechnological applications. Actinomycetologica 2000, 14, 17–27. [Google Scholar]
- Kurtböke, DI (Ed.) Selective Isolation of Rare Actinomycetes; Queensland Complete Printing Services: Nambour, Australia, 2003.
- Kurtböke, DI. Watanabe, MM, Suzuki, K, Seki, T, Eds.; Actino-Rush on the Sunshine Coast: Prospects for Bioprospecting. In Proceedings of the 10th International Congress on Culture Collections; Japan Society for Culture Collections and World Federation of Culture Collections Publication, Tokyo, Japan, 2004a; pp. 319–322. [Google Scholar]
- Kurtböke, DI. Kurtböke, DI, Swings, J, Eds.; Uniqueness of the “Smart State’s” microbial diversity from a microbial library to biodiscovery: An academic initiative complementing Queensland Government’s Action Plan. In Microbial Genetic Resources and Biodiscovery; Queensland Complete Printing Services: Nambour, Australia, 2004b; pp. 236–248. [Google Scholar]
- Kurtböke, DI; Cardillo, R; Nasini, G; Petrolini, B; Sardi, P; Locci, R. Lechevalier, H, Ensign, JC, Kalakoutskii, LV, Locci, R, Eds.; Anti-candida metabolites produced by streptomycetes isolated from the atmosphere of agricultural environments. In Actinomycetes; Waksman Institute of Microbiology, Rutgers and The State University of New Jersey, 1986; pp. 223–231. [Google Scholar]
- Lazzarini, A; Cavaletti, L; Toppo, G; Marinelli, F. Rare genera of actinomycetes as potential producers of new antibiotics. Antonie van Leeuwenhoek 2000, 78, 388–405. [Google Scholar]
- Long, RA; Azam, F. Antagonistic Interactions among Marine Pelagic Bacteria. Appl Environ Microb 2001, 67, 4975–4983. [Google Scholar]
- Ludwig, W; Strunk, O; Klugbauer, S; Klugbauer, N; Weizenegger, M; Neumaier, J. Bacterial phylogeny based on comparative sequence analysis. Electrophoresis 1998, 19, 554–568. [Google Scholar]
- Ludwig, W; Strunk, O; Westram, R; Richter, L; Meier, H; Yadhukumar Bucher, A; Lai, T; Steppi, S; Jobb, G; Föster, W; Brettske, I; Gerber, S; Ginhart, AW; Gross, O; Grumann, S; Hermann, S; Jost, R; König, A; Liss, T; Lüssmann, R; May, M; Nonhoff, B; Reichel, B; Strehlow, R; Stamatakis, A; Stuckmann, N; Vilbig, A; Lenke, M; Ludwig, T; Bode, A; Schleifer, KH. ARB: a software environment for sequence data. Nuc Acid Res 2004, 32, 1363–1371. [Google Scholar]
- Magarvey, NA; Keller, JM; Bernan, V; Dworkin, M; Sherman, DH. Isolation and characterization of novel marine-derived actinomycete taxa rich in bioactive metabolites. Appl Environ Microbiol 2004, 70, 7520–7529. [Google Scholar]
- Mincer, TJ; Jensen, PR; Kauffman, CA; Fenical, W. Widespread and persistent populations of a major new marine actinomycete taxon in the ocean sediments. Appl Environ Microb 2002, 68, 5005–5011. [Google Scholar]
- Okazaki, T. Kurtböke, DI, Ed.; Selective Isolation of Rare Actinomycetes; Queensland Complete Printing Services: Nambour, Australia, 2003; pp. 102–122. [Google Scholar]
- Proksch, P; Edrada, RA; Ebel, R. Drugs from the seas: current status and microbiological implications. Appl Microbiol Biotech 2002, 59, 125–134. [Google Scholar]
- Radajewski, S; Webster, G; Reay, DS; Morris, SA; Ineson, P; Nedwell, DB; Prosser, JI; Murrell, JC. Identification of active methylotroph populations in an acidic forest soil by stable isotope probing. Microbiology 2002, 148, 2331–2342. [Google Scholar]
- Rowbotham, TJ; Cross, T. Ecology of Rhodococcus coprophilus and associated actinomycetes in fresh water and agricultural habitats. J Gen Microbiol 1977, 100, 231–240. [Google Scholar]
- Rusnak, K; Troyanovich, J; Mierzwa, R; Chu, M; Patel, M; Weistein, M. An antibiotic with activity against gram-positive bacteria from the gentamicin-producing strain of Micromonospora purpurea. Appl Microbiol Biotechnol 2001, 56, 502–503. [Google Scholar]
- Seiler, N. Blau, K, Halket, JM, Eds.; Fluorescent Derivatives. In Handbook of Derivatives for Chromatography; John Wiley and Sons: New York, USA, 1993; pp. 175–213. [Google Scholar]
- Shimizu, M; Nakagawa, Y; Sato, Y; Furumai, T; Igarashi, Y; Onaka, H; Yoshida, R; Kunoh, H. Studies on endophytic Actinomycetes (I) Streptomyces sp. isolated from rhododendron and its antifungal activity. J Gen Plant Pathol 2000, 66, 360–366. [Google Scholar]
- Terekhova, LP. Kurtböke, DI, Ed.; Isolation of actinomycetes with the use of microwaves and electric pulses. In Selective Isolation of Rare Actinomycetes; Queensland Complete Printing Services: Nambour, Australia, 2003; pp. 82–101. [Google Scholar]
- Vickers, JC; Williams, ST. An assessment of plate inoculation procedures for the enumeration and isolation of soil streptomycetes. Microbios Lett 1987, 35, 113–117. [Google Scholar]
- Waksman, SA. Streptomycin: background, isolation properties, and utilization. Nobel Lecture, December 12, 1952. In Nobel Lectures, Physiology and Medicine 1942–1962; Elsevier: Amsterdam, Netherlands, 1964; pp. 370–388. [Google Scholar]
- Watanabe, Y; Shinzato, N; Fukatsu, T. Isolation of Actinomycetes from termites’ guts. Biosci Biotechnol Biochem 2003, 67, 1797–1801. [Google Scholar]
- Weinstein, MJ; Luedemann, GM; Oden, EM; Wagman, GH. Gentamicin, a new broad-spectrum antibiotic complex. Antimicrob Agents Ch 1964, 1–7. [Google Scholar]
- Wellington, EMH; Williams, ST. Preservation of actinomycete inoculum in frozen glycerol. Microbios Lett 1978, 6, 151–157. [Google Scholar]
- Williams, PG; Buchanan, GO; Feling, RH; Kauffman, CA; Jensen, PR; Fenical, W. New cytotoxic salinosporamides from marine actinomycete Salinispora tropica. J Org Chem 2005, 70, 6196–6203. [Google Scholar]
- Williams, ST. Are antibiotics produced in soil? Pedobiologia 1982, 23, 427–435. [Google Scholar]
- Williams, ST; Davies, FL. Use of antibiotics for selective isolation and enumeration of actinomycetes in soil. J Gen Microbiol 1965, 38, 251–261. [Google Scholar]
- Williams, ST; Vickers, JC. Ecology of antibiotic production. Microb Ecol 1986, 12, 43–52. [Google Scholar]
- Williams, ST; Wellington, EMH. Nisbet, LJ, Winstanley, DJ, Eds.; Principles and problems of selective isolation of microbes. In Bioactive Microbial Products: Search and Discovery; Academic Press: London, UK, 1982a; pp. 9–26. [Google Scholar]
- Williams, ST; Wellington, EMH. Page, AL, Miller, RH, Keeney, DR, Eds.; Actinomycetes. In Methods of Soil Analysis, Part 2, Chemical and Microbiological Properties; American Society of Agronomy/Soil Science Society of America Publications: Madison, USA, 1982b; pp. 969–987. [Google Scholar]
- Williams, ST; Goodfellow, M; Vickers, JC. Kelly, DP, Carr, NG, Eds.; New microbes from old habitats? In The Microbe 1984, II: Prokaryotes and Eukaryotes; Cambridge University Press: Cambridge, UK, 1984a; pp. 219–256. [Google Scholar]
- Williams, ST; Lanning, S; Wellington, EMH. Goodfellow, M, Mordarski, M, Williams, ST, Eds.; Ecology of actinomycetes. In The Biology of Actinomycetes; Academic Press, Inc.: London, UK, 1984b; pp. 481–528. [Google Scholar]
- Wohl, DL; McArthur, JV. Actinomycete-flora associated with submersed freshwater macrophytes. FEMS Microbiol Ecol 1998, 26, 135–140. [Google Scholar]
- Zheng, Z; Zeng, W; Huang, Y; Yang, Z; Li, J; Cai, H; Su, W. Detection of antitumor and antimicrobial activities in marine organism associated actinomycetes isolated from the Taiwan Strait, China. FEMS Microbiol Lett 2000, 188, 87–91. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Locations | Habitat types | Habitat Descriptive Image |
|---|---|---|
| Petrie Ck, McDonald Rd, Diddillibah (S1)
Paynter Ck, Diddillibah Rd, Diddillibah (S2) Maroochy R, River Store Rd, Maroochy (S3) | HB-1
Mangrove mud sediments derived from the Maroochy River or tributaries |  |
| Maroochy R, Muller Park, Bli Bli (S4)
Maroochy R, Oysterbank Rd, Bli Bli (S5) Maroochy R, Nojoor Rd, Mudjimba (S6) Eudlo Ck, Fishermans Rd, Maroochydore (S7) | HB-2
Sand sediments derived from the Maroochy River or tributaries |  |
| Stumers Ck, David Low Way, Coolum (S19)
Coondibah Ck, Flinders St, Currumundi (S20) Currumundi Lake, Noel Burns Park, Currumundi (S21) Mooloolah R, Marra Ct, Mountain Creek (S22) | HB-3
Marine sand sediments not derived from the Maroochy River or tributaries |  |
| Lillyponds, Delicia Rd, Mapleton (S11)
Tesch Park, Coral St, Maleny (S16) Ewen Maddock Dam, Glass House Mountains Rd, Landsborough (S17) Lake Macdonald, Noosa Shire Botanical Gardens, Pomona (S18) Wetland adjacent to David Low Way, Marcoola (S23) | HB-4
Fine, silty sediment from freshwater habitats |  |
| Obi Obi Creek Crossing no. 2, Obi Obi Rd, Kidaman Creek (S12)
Little Yabba Ck, Little Yabba Creek Picnic Area, Conondale- Kenilworth Rd, Kenilworth (S13) Boolumba Ck, Dayuse Area - Area 2, Booloomba Creek Rd, Kenilworth (S14) Mary River, Maleny-Kenilworth Rd, Conondale (S15) | HB-5
Coarse grained sediment from freshwater habitats |  |
| Wappa Dam, Wappa Dam Rd, Kiamba (L24)
Wetland adjacent to David Low Way, Marcoola (L25) Hyatt Regency Lakes, David Low Way, Coolum (L26) Nelson Park Lakes, Alexandra Headland (L27) Lake Macdonald, Noosa Shire Botanical Gardens, Pomona (S18) Ewen Maddock Dam, Glass House Mountains Rd, Landsborough (S17) Wetland adjacent to David Low Way, Marcoola (S23) | HB-6
Lily-leaf samples from freshwater habitats |  |
| DSM No. | Organism | Antibiotic produced | Isolated from |
|---|---|---|---|
| DSM 803 | M. rosaria | Rosaramycin | Soil |
| DSM 43168 | M. carbonacea | Everninomicins | Soil |
| DSM 43171 | M. halophytica | Halomycin | Salt pool |
| DSM 43816 | M. echinospora | Gentamicin | Soil |
| DSM 43817 | M. pallida | Gentamicin | Soil |
| DSM 43818 | M. nigra | Halomycin | Salt pool |
| DSM 43868 | M. olivasterospora | Fortamicin B | Soil, paddy field |
| Original Isolation Media | Number of isolates screened for antibacterial activity | Antibacterial activity | Percent (%) of total |
|---|---|---|---|
| With gentamicin | 59 | 6 | 10.2% |
| Without gentamicin | 12 | 6 | 50.0% |
| Total | 71 | 12 | 16.9% |
| Isolate no. | Antimicrobial activity | ||
|---|---|---|---|
| S. aureus | E. coli | VRE | |
| USC-702 a | ✓ | X | X |
| USC-703 a | ✓ | X | X |
| USC-705 b | ✓ | X | X |
| USC-706 b | ✓ | X | X |
| USC-707 b | ✓ | X | X |
| USC-708 a | ✓ | X | X |
| USC-710 b | ✓ | ✓ | ✓ |
| USC-712 b | ✓ | X | X |
| USC-713 b | ✓ | X | X |
| USC-731 a | ✓ | ✓ | X |
| USC-757 a | ✓ | X | X |
| USC-758 a | ✓ | ✓ | ✓ |
Share and Cite
Eccleston, G.P.; Brooks, P.R.; Kurtböke, D.I. The Occurrence of Bioactive Micromonosporae in Aquatic Habitats of the Sunshine Coast in Australia. Mar. Drugs 2008, 6, 243-261. https://doi.org/10.3390/md6020243
Eccleston GP, Brooks PR, Kurtböke DI. The Occurrence of Bioactive Micromonosporae in Aquatic Habitats of the Sunshine Coast in Australia. Marine Drugs. 2008; 6(2):243-261. https://doi.org/10.3390/md6020243
Chicago/Turabian StyleEccleston, Glen P., Peter R. Brooks, and D. Ipek Kurtböke. 2008. "The Occurrence of Bioactive Micromonosporae in Aquatic Habitats of the Sunshine Coast in Australia" Marine Drugs 6, no. 2: 243-261. https://doi.org/10.3390/md6020243
APA StyleEccleston, G. P., Brooks, P. R., & Kurtböke, D. I. (2008). The Occurrence of Bioactive Micromonosporae in Aquatic Habitats of the Sunshine Coast in Australia. Marine Drugs, 6(2), 243-261. https://doi.org/10.3390/md6020243