Impact of Marine Drugs on Cytoskeleton-Mediated Reproductive Events


Abstract
:1. Introduction
2. Microfilaments or Actin Filaments
2.1. Nucleation and Function of Microfilaments
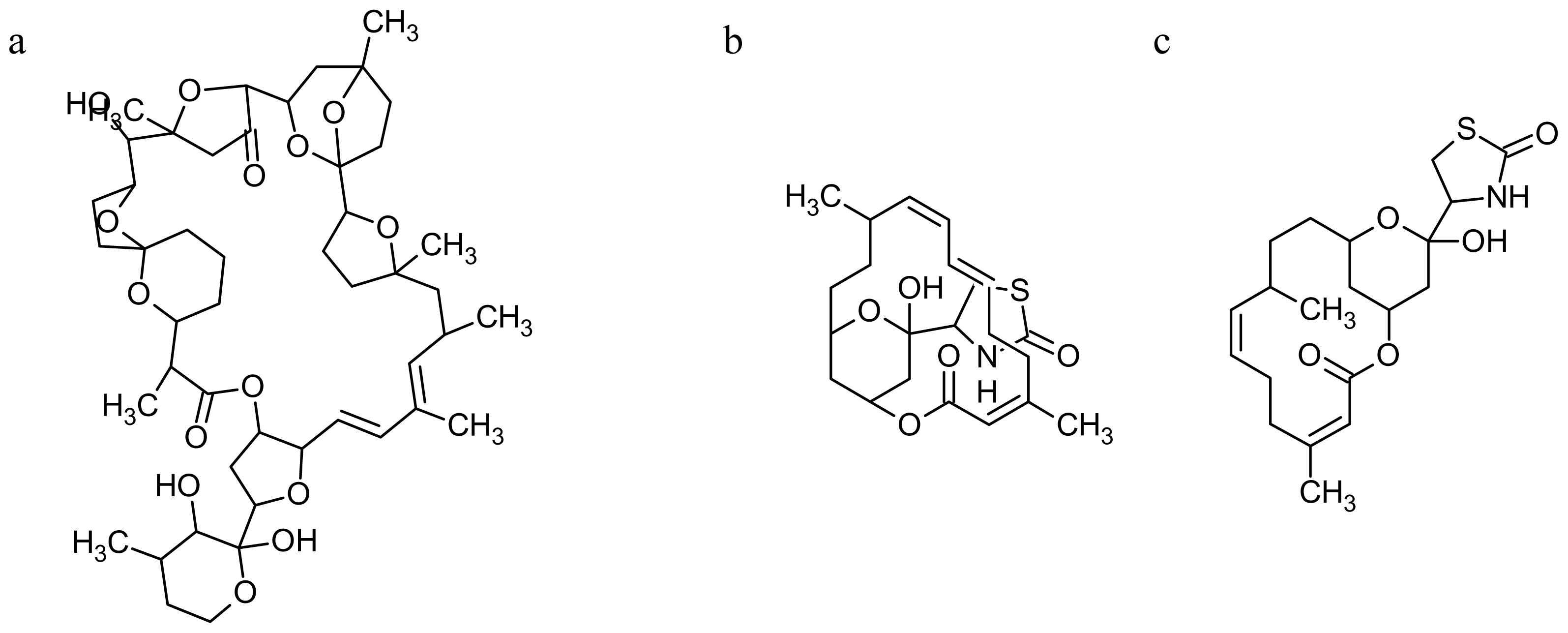
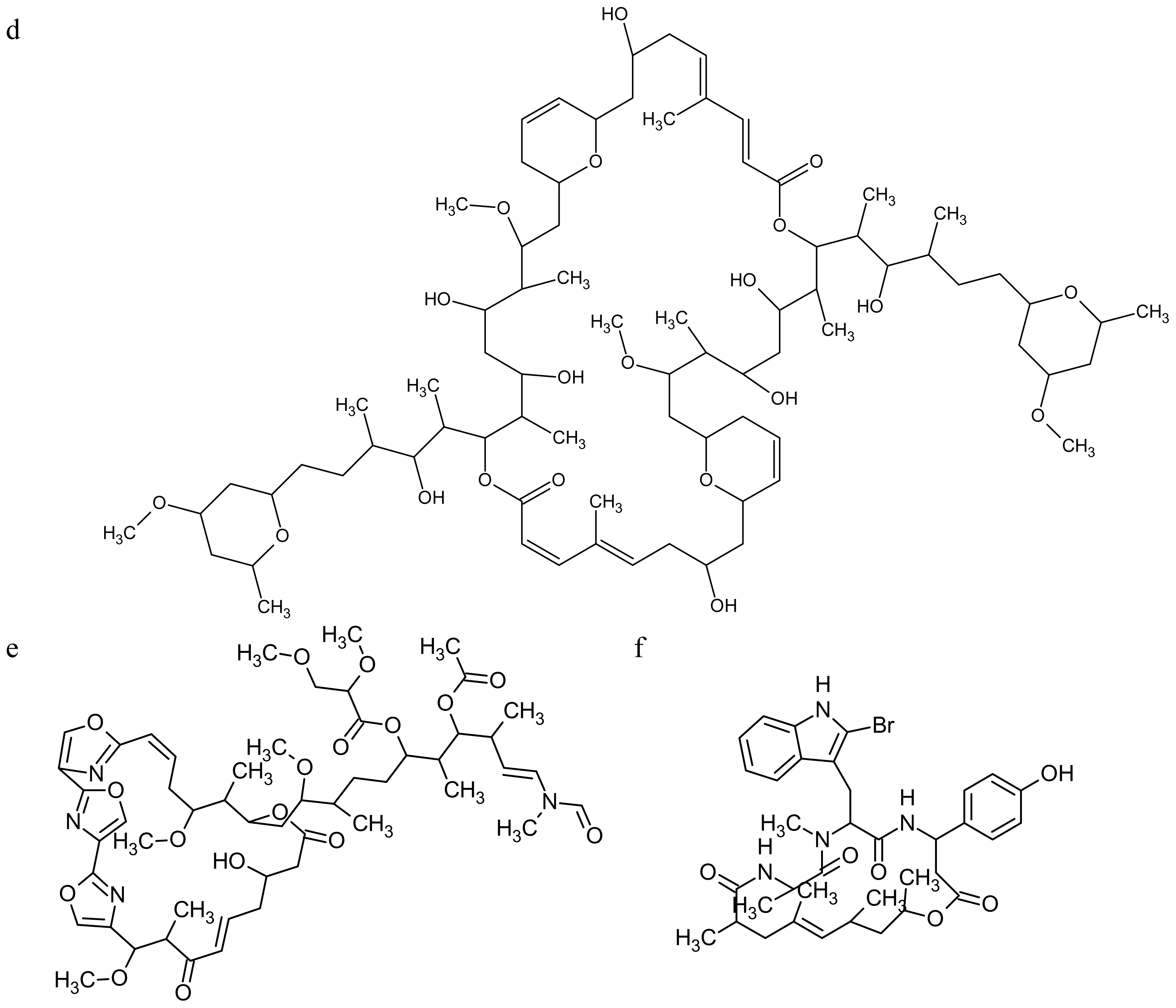
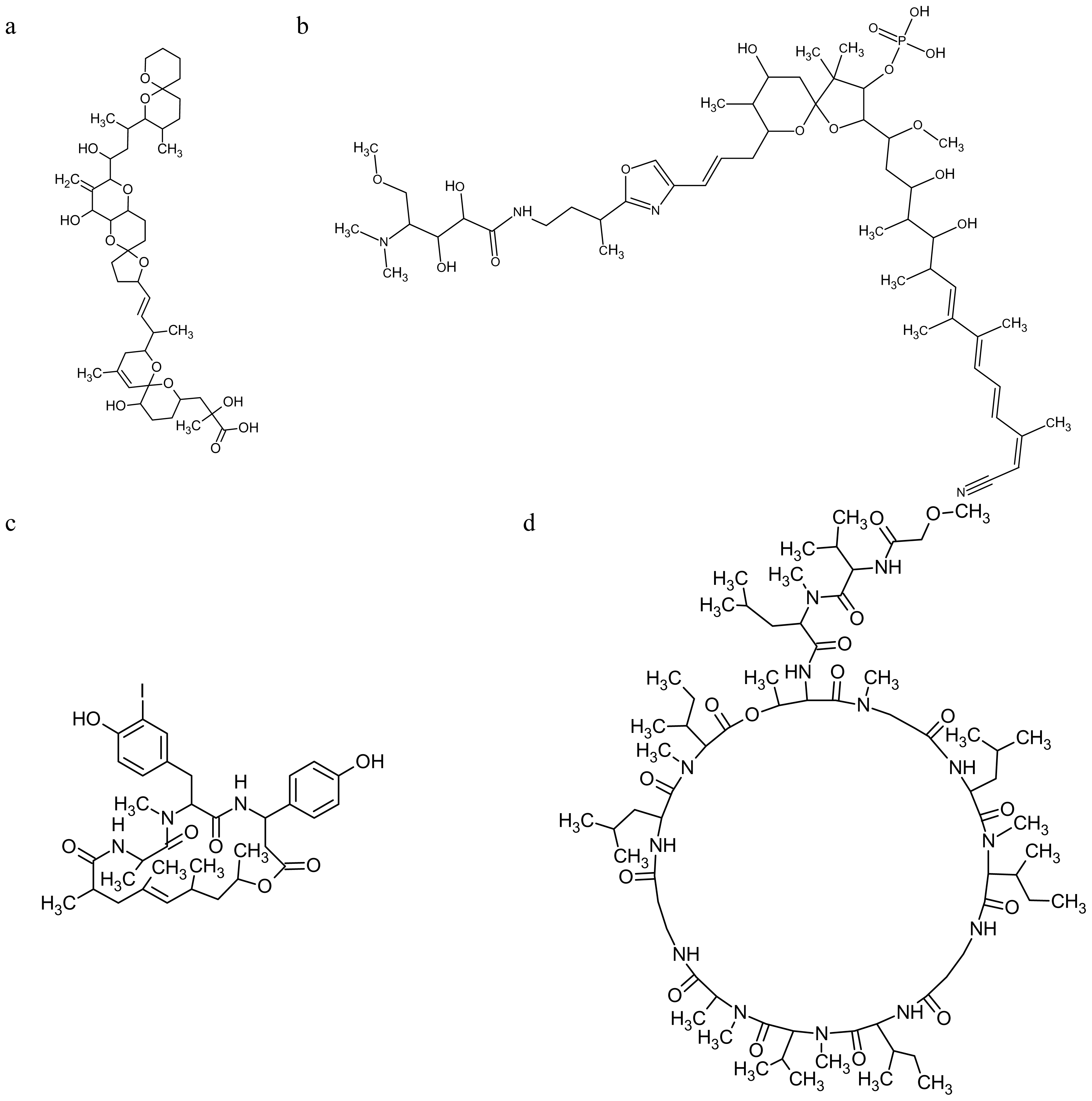
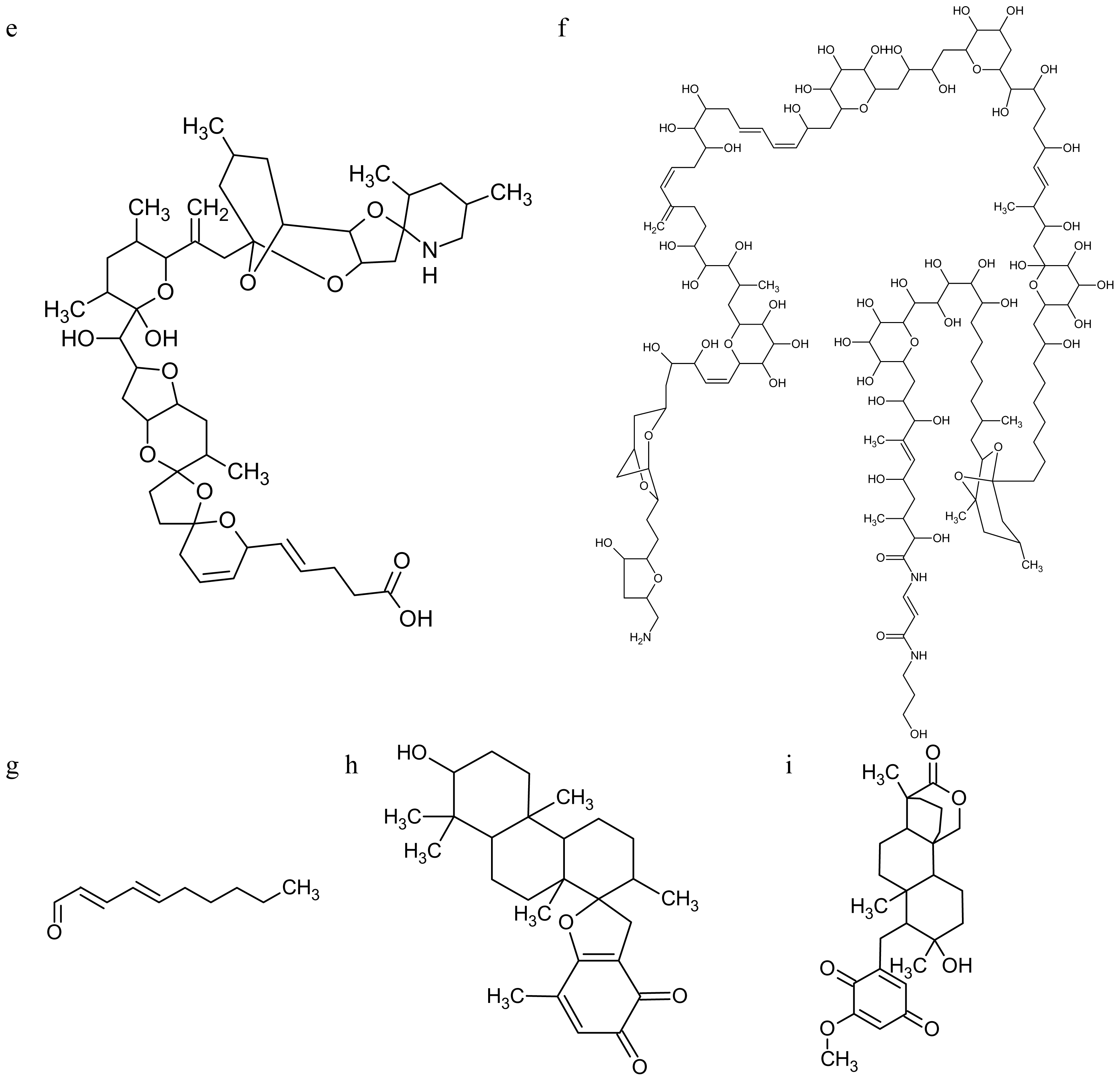
2.2. Marine Toxins Interfering with Microfilament Network
3. Microtubules
3.1. Microtubule Formation, Functions and Dynamics
3.2. Marine Toxins Interfering with Microtubules
4. Reproduction
4.1. Effect of Marine Drugs on Cytoskeletal Dynamics Involved in Reproductive Events
4.1.1. Spermatogenesis
4.1.2. Sperm motility
4.1.3. Oogenesis
4.2. Gamete Activation and Fertilization
4.2.1. Sperm capacitation and acrosome reaction
4.2.2. Fertilization
4.3. Early Development
5. Conclusions
Acknowledgments
References
- Silvestre, F; Tosti, E. Impact of marine drugs on animal reproductive processes. Mar Drugs 2009, 7, 539–564. [Google Scholar]
- Wegner, A. Head to tail polymerization of actin. J Mol Biol 1976, 108, 139–150. [Google Scholar]
- Kueh, HY; Mitchison, TJ. Structural plasticity in actin and tubulin polymer dynamics. Science 2009, 325, 960–963. [Google Scholar]
- Giganti, A; Friederich, E. The actin cytoskeleton as a therapeutic target: state of the art and future directions. Prog Cell Cycle Res 2003, 5, 511–525. [Google Scholar]
- Diez, S; Gerisch, G; Anderson, K; Muller-Taubenberger, A; Bretschneider, T. Subsecond reorganization of the actin network in cell motility and chemotaxis. Proc Natl Acad Sci USA 2005, 102, 7601–7606. [Google Scholar]
- Xiao, X; Yang, WX. Actin-based dynamics during spermatogenesis and its significance. J Zhejiang Univ Sci B 2007, 8, 498–506. [Google Scholar]
- Gunst, SJ. Actions by actin: reciprocal regulation of cortactin activity by tyrosine kinases and F-actin. Biochem J 2004, 380, e7–e8. [Google Scholar]
- Vartiainen, MK; Machesky, LM. The WASP-Arp2/3 pathway: genetic insights. Curr Opin Cell Biol 2004, 16, 174–181. [Google Scholar]
- Zigmond, SH. Beginning and ending an actin filament: control at the barbed end. Curr Top Dev Biol 2004, 63, 145–188. [Google Scholar]
- Gohla, A; Birkenfeld, J; Bokoch, GM. Chronophin, a novel HAD-type serine protein phosphatase, regulates cofilin-dependent actin dynamics. Nat Cell Biol 2005, 7, 21–29. [Google Scholar]
- Sun, QY; Schatten, H. Regulation of dynamic events by microfilaments during oocyte maturation and fertilization. Reproduction 2006, 131, 193–205. [Google Scholar]
- Didry, D; Carlier, MF; Pantaloni, D. Synergy between actin depolymerizing factor/cofilin and profilin in increasing actin filament turnover. J Biol Chem 1998, 273, 25602–25611. [Google Scholar]
- Loisel, TP; Boujemaa, R; Pantaloni, D; Carlier, MF. Reconstitution of actin-based motility of Listeria and Shigella using pure proteins. Nature 1999, 401, 613–616. [Google Scholar]
- Balasubramanian, MK; Hirani, BR; Burke, JD; Gould, KL. The Schizosaccharomyces pombe cdc3+ gene encodes a profilin essential for cytokinesis. J Cell Biol 1994, 125, 1289–1301. [Google Scholar]
- Gunsalus, KC; Bonaccorsi, S; Williams, E; Verni, F; Gatti, M; Goldberg, ML. Mutations in twinstar, a Drosophila gene encoding a cofilin/ADF homologue, result in defects in centrosome migration and cytokinesis. J Cell Biol 1995, 131, 1243–1259. [Google Scholar]
- Somma, MP; Fasulo, B; Cenci, G; Cundari, E; Gatti, M. Molecular dissection of cytokinesis by RNA interference in Drosophila cultured cells. Mol Biol Cell 2002, 13, 2448–2460. [Google Scholar]
- Nusco, GA; Chun, JT; Ercolano, E; Lim, D; Gragnaniello, G; Kyozuka, K; Santella, L. Modulation of calcium signalling by the actin-binding protein cofilin. Biochem Biophys Res Commun 2006, 348, 109–114. [Google Scholar]
- Spector, I; Braet, F; Shochet, NR; Bubb, MR. New anti-actin drugs in the study of the organization and function of the actin cytoskeleton. Microsc Res Tech 1999, 47, 18–37. [Google Scholar]
- Fenteany, G; Zhu, S. Small-molecule inhibitors of actin dynamics and cell motility. Curr Top Med Chem 2003, 3, 593–616. [Google Scholar]
- Jordan, MA; Wilson, L. Microtubules and actin filaments: dynamic targets for cancer chemotherapy. Curr Opin Cell Biol 1998, 10, 123–130. [Google Scholar]
- Allingham, JS; Klenchin, VA; Rayment, I. Actin-targeting natural products: structures, properties and mechanisms of action. Cell Mol Life Sci 2006, 63, 2119–2134. [Google Scholar]
- Yasumoto, T; Murata, M; Oshima, Y; Sano, M; Matsumoto, GK; Clardy, J. Diarrheic shellfish toxins. Tetrahedron 1985, 41, 1019–1025. [Google Scholar]
- Espina, B; Rubiolo, JA. Marine toxins and the cytoskeleton: pectenotoxins, unusual macrolides that disrupt actin. Febs J 2008, 275, 6082–6088. [Google Scholar]
- Spector, I; Shochet, NR; Kashman, Y; Groweiss, A. Latrunculins: novel marine toxins that disrupt microfilament organization in cultured cells. Science 1983, 219, 493–495. [Google Scholar]
- Yarmola, EG; Somasundaram, T; Boring, TA; Spector, I; Bubb, MR. Actin-latrunculin A structure and function. Differential modulation of actin-binding protein function by latrunculin A. J Biol Chem 2000, 275, 28120–28127. [Google Scholar]
- Coue, M; Brenner, SL; Spector, I; Korn, ED. Inhibition of actin polymerization by latrunculin A. FEBS Lett 1987, 213, 316–318. [Google Scholar]
- Hertzog, M; Yarmola, EG; Didry, D; Bubb, MR; Carlier, MF. Control of actin dynamics by proteins made of beta-thymosin repeats: the actobindin family. J Biol Chem 2002, 277, 14786–14792. [Google Scholar]
- Ayscough, KR; Stryker, J; Pokala, N; Sanders, M; Crews, P; Drubin, DG. High rates of actin filament turnover in budding yeast and roles for actin in establishment and maintenance of cell polarity revealed using the actin inhibitor latrunculin-A. J Cell Biol 1997, 137, 399–416. [Google Scholar]
- Morton, WM; Ayscough, KR; McLaughlin, PJ. Latrunculin alters the actin-monomer subunit interface to prevent polymerization. Nat Cell Biol 2000, 2, 376–378. [Google Scholar]
- Saito, SY. Toxins affecting actin filaments and microtubules. Prog Mol Subcell Biol 2009, 46, 187–219. [Google Scholar]
- Spector, I; Shochet, NR; Blasberger, D; Kashman, Y. Latrunculins--novel marine macrolides that disrupt microfilament organization and affect cell growth: I. Comparison with cytochalasin D. Cell Motil Cytoskeleton 1989, 13, 127–144. [Google Scholar]
- Kobayashi, M; Tanaka, J; Katori, T; Kitagawa, I. Marine natural products. XXIII. Three new cytotoxic dimeric macrolides, swinholides B and C and isoswinholide A, congeners of swinholide A, from the Okinawan marine sponge Theonella swinhoei. Chem Pharm Bull (Tokyo) 1990, 38, 2960–2966. [Google Scholar]
- Bubb, MR; Spector, I; Bershadsky, AD; Korn, ED. Swinholide A is a microfilament disrupting marine toxin that stabilizes actin dimers and severs actin filaments. J Biol Chem 1995, 270, 3463–3466. [Google Scholar]
- Fusetani, N; Yasumuro, K; Matsunaga, S; Hashimoto, K. Mycalolides A–C, hybrid macrolides of ulapualides and halichondramide, from a sponge of the genus. Mycale Tetrahedron Lett 1989, 30, 2809–2812. [Google Scholar]
- Saito, S; Watabe, S; Ozaki, H; Fusetani, N; Karaki, H. Mycalolide B, a novel actin depolymerizing agent. J Biol Chem 1994, 269, 29710–29714. [Google Scholar]
- Saito, SY; Watabe, S; Ozaki, H; Kobayashi, M; Suzuki, T; Kobayashi, H; Fusetani, N; Karaki, H. Actin-depolymerizing effect of dimeric macrolides, bistheonellide A and swinholide A. J Biochem 1998, 123, 571–578. [Google Scholar]
- Hori, M; Saito, S; Shin, YZ; Ozaki, H; Fusetani, N; Karaki, H. Mycalolide-B, a novel and specific inhibitor of actomyosin ATPase isolated from marine sponge. FEBS Lett 1993, 322, 151–154. [Google Scholar]
- Ohizumi, Y. Application of physiologically active substances isolated from natural resources to pharmacological studies. Jpn J Pharmacol 1997, 73, 263–289. [Google Scholar]
- Bubb, MR; Senderowicz, AM; Sausville, EA; Duncan, KL; Korn, ED. Jasplakinolide, a cytotoxic natural product, induces actin polymerization and competitively inhibits the binding of phalloidin to F-actin. J Biol Chem 1994, 269, 14869–14871. [Google Scholar]
- Bubb, MR; Spector, I; Beyer, BB; Fosen, KM. Effects of jasplakinolide on the kinetics of actin polymerization. An explanation for certain in vivo observations. J Biol Chem 2000, 275, 5163–5170. [Google Scholar]
- Botana, LM. Marine toxins and the cytoskeleton. Febs J 2008, 275, 6059. [Google Scholar]
- Vale, C; Botana, LM. Marine toxins and the cytoskeleton: okadaic acid and dinophysistoxins. Febs J 2008, 275, 6060–6066. [Google Scholar]
- Heideker, J; Lis, ET; Romesberg, FE. Phosphatases, DNA damage checkpoints and checkpoint deactivation. Cell Cycle 2007, 6, 3058–3064. [Google Scholar]
- Eriksson, JE; Brautigan, DL; Vallee, R; Olmsted, J; Fujiki, H; Goldman, RD. Cytoskeletal integrity in interphase cells requires protein phosphatase activity. Proc Natl Acad Sci USA 1992, 89, 11093–11097. [Google Scholar]
- Ishihara, H; Martin, BL; Brautigan, DL; Karaki, H; Ozaki, H; Kato, Y; Fusetani, N; Watabe, S; Hashimoto, K; Uemura, D; et al. Calyculin A and okadaic acid: inhibitors of protein phosphatase activity. Biochem Biophys Res Commun 1989, 159, 871–877. [Google Scholar]
- Smith, GD; Sadhu, A; Wolf, DP. Transient exposure of rhesus macaque oocytes to calyculin-A and okadaic acid stimulates germinal vesicle breakdown permitting subsequent development and fertilization. Biol Reprod 1998, 58, 880–886. [Google Scholar]
- Rangel, M; Prado, MP; Konno, K; Naoki, H; Freitas, JC; Machado-Santelli, GM. Cytoskeleton alterations induced by Geodia corticostylifera depsipeptides in breast cancer cells. Peptides 2006, 27, 2047–2057. [Google Scholar]
- Mayer, AM; Gustafson, KR. Marine pharmacology in 2005–2006: antitumour and cytotoxic compounds. Eur J Cancer 2008, 44, 2357–2387. [Google Scholar]
- Kitagawa, I; Ohashi, K; Kawanishi, H; Shibuya, H; Shinkai, K; Akedo, H. Ionophoretic activities of oligopeptide lactones and resin-glycosides in human erythrocytes. Chem Pharm Bull (Tokyo) 1989, 37, 1679–1681. [Google Scholar]
- Kobayashi, M; Lee, NK; Shibuya, H; Momose, T; Kitagawa, I. Marine natural products. XXVI. Biologically active tridecapeptide lactones from the Okinawan marine sponge Theonella swinhoei (Theonellidae). (2). Structures of theonellapeptolides Ia, Ib, Ic, and Ie. Chem Pharm Bull (Tokyo) 1991, 39, 1177–1184. [Google Scholar]
- Ohta, E; Okada, H; Ohta, S; Kobayashi, M; Kitagawa, I; Horiike, S; Takahashi, T; Hosoya, H; Yamamoto, K; Ikegami, S. Malformation of immature starfish oocytes by theonellapeptolide Ie, a Tridecapeptide lactone from a marine sponge Petrosia species, through disturbance of cortical F-actin distribution. Biosci Biotechnol Biochem 2003, 67, 1908–1915. [Google Scholar]
- Satake, M; Ofuji, K; Naoki, H; James, K; Furey, A; McMahon, T; Silke, J; Yasumoto, T. Azaspiracid, a new marine toxin having unique spiro ring assemblies, isolated from Irish mussels, Mytilus edulis. J Am Chem Soc 1998, 120, 9967–9968. [Google Scholar]
- Ofuji, K; Satake, M; McMahon, T; Silke, J; James, KJ; Naoki, H; Oshima, Y; Yasumoto, T. Two analogs of azaspiracid isolated from mussels, Mytilus edulis, involved in human intoxication in Ireland. Nat Toxins 1999, 7, 99–102. [Google Scholar]
- Ofuji, K; Satake, M; McMahon, T; James, KJ; Naoki, H; Oshima, Y; Yasumoto, T. Structures of azaspiracid analogs, azaspiracid-4 and azaspiracid-5, causative toxins of azaspiracid poisoning in Europe. Biosci Biotechnol Biochem 2001, 65, 740–742. [Google Scholar]
- Volmer, DA; Brombacher, S; Whitehead, B. Studies on azaspiracid biotoxins. I. Ultrafast high-resolution liquid chromatography/mass spectrometry separations using monolithic columns. Rapid Commun Mass Spectrom 2002, 16, 2298–2305. [Google Scholar]
- James, KJ; Sierra, MD; Lehane, M; Brana Magdalena, A; Furey, A. Detection of five new hydroxyl analogues of azaspiracids in shellfish using multiple tandem mass spectrometry. Toxicon 2003, 41, 277–283. [Google Scholar]
- Rehmann, N; Hess, P; Quilliam, MA. Discovery of new analogs of the marine biotoxin azaspiracid in blue mussels (Mytilus edulis) by ultra-performance liquid chromatography/tandem mass spectrometry. Rapid Commun Mass Spectrom 2008, 22, 549–558. [Google Scholar]
- Vilarino, N. Marine toxins and the cytoskeleton: azaspiracids. Febs J 2008, 275, 6075–6081. [Google Scholar]
- Usami, M; Satake, M; Ishida, S; Inoue, A; Kan, Y; Yasumoto, T. Palytoxin analogs from the dinoflagellate Ostreopsis siamensis. J Am Chem Soc 1995, 117, 5389–5390. [Google Scholar]
- Taniyama, S; Arakawa, O; Terada, M; Nishio, S; Takatani, T; Mahmud, Y; Noguchi, T. Ostreopsis sp., a possible origin of palytoxin (PTX) in parrotfish Scarus ovifrons. Toxicon 2003, 42, 29–33. [Google Scholar]
- Louzao, MC; Ares, IR; Cagide, E. Marine toxins and the cytoskeleton: a new view of palytoxin toxicity. Febs J 2008, 275, 6067–6074. [Google Scholar]
- Miralto, A; Barone, G; Romano, G; Poulet, SA; Ianora, A; Russo, GL; Buttino, I; Mazzarella, G; Laabir, M; Cabrini, M; Giacobbe, MG. The insidious effect of diatoms on copepod reproduction. Nature 1999, 402, 173–176. [Google Scholar]
- Caldwell, GS. The influence of bioactive oxylipins from marine diatoms on invertebrate reproduction and development. Mar Drugs 2009, 7, 367–400. [Google Scholar]
- Caldwell, GS; Bentley, MG; Olive, PJW. First evidence of sperm motility inhibition by the diatom aldehyde 2E,4E-decadienal. Mar Ecol Prog Ser 2004, 273, 97–108. [Google Scholar]
- McHardy, LM; Warabi, K; Andersen, RJ; Roskelley, CD; Roberge, M. Strongylophorine-26, a Rho-dependent inhibitor of tumor cell invasion that reduces actin stress fibers and induces nonpolarized lamellipodial extensions. Mol Cancer Ther 2005, 4, 772–778. [Google Scholar]
- Mitchison, T; Kirschner, M. Dynamic instability of microtubule growth. Nature 1984, 312, 237–242. [Google Scholar]
- Margolis, RL; Wilson, L. Opposite end assembly and disassembly of microtubules at steady state in vitro. Cell 1978, 13, 1–8. [Google Scholar]
- Margolis, RL; Wilson, L. Microtubule treadmills--possible molecular machinery. Nature 1981, 293, 705–711. [Google Scholar]
- Wang, YL. Exchange of actin subunits at the leading edge of living fibroblasts: possible role of treadmilling. J Cell Biol 1985, 101, 597–602. [Google Scholar]
- Theriot, JA; Mitchison, TJ. Actin microfilament dynamics in locomoting cells. Nature 1991, 352, 126–131. [Google Scholar]
- Mountain, V; Compton, DA. Dissecting the role of molecular motors in the mitotic spindle. Anat Rec 2000, 261, 14–24. [Google Scholar]
- Alberti, C. Cytoskeleton structure and dynamic behaviour: quick excursus from basic molecular mechanisms to some implications in cancer chemotherapy. Eur Rev Med Pharmacol Sci 2009, 13, 13–21. [Google Scholar]
- Wade, RH. On and around microtubules: an overview. Mol Biotechnol 2009, 43, 177–191. [Google Scholar]
- Hartwell, LH; Weinert, TA. Checkpoints: controls that ensure the order of cell cycle events. Science 1989, 246, 629–634. [Google Scholar]
- Murray, AW. Creative blocks: cell-cycle checkpoints and feedback controls. Nature 1992, 359, 599–604. [Google Scholar]
- Vale, RD. The molecular motor toolbox for intracellular transport. Cell 2003, 112, 467–480. [Google Scholar]
- Vallee, RB; Williams, JC; Varma, D; Barnhart, LE. Dynein: An ancient motor protein involved in multiple modes of transport. J Neurobiol 2004, 58, 189–200. [Google Scholar]
- Howard, J; Hudspeth, AJ; Vale, RD. Movement of microtubules by single kinesin molecules. Nature 1989, 342, 154–158. [Google Scholar]
- Block, SM; Goldstein, LS; Schnapp, BJ. Bead movement by single kinesin molecules studied with optical tweezers. Nature 1990, 348, 348–352. [Google Scholar]
- Wang, Z; Khan, S; Sheetz, MP. Single cytoplasmic dynein molecule movements: characterization and comparison with kinesin. Biophys J 1995, 69, 2011–2023. [Google Scholar]
- Mallik, R; Carter, BC; Lex, SA; King, SJ; Gross, SP. Cytoplasmic dynein functions as a gear in response to load. Nature 2004, 427, 649–652. [Google Scholar]
- Toba, S; Watanabe, TM; Yamaguchi-Okimoto, L; Toyoshima, YY; Higuchi, H. Overlapping hand-over-hand mechanism of single molecular motility of cytoplasmic dynein. Proc Natl Acad Sci USA 2006, 103, 5741–5745. [Google Scholar]
- Reck-Peterson, SL; Yildiz, A; Carter, AP; Gennerich, A; Zhang, N; Vale, RD. Single-molecule analysis of dynein processivity and stepping behavior. Cell 2006, 126, 335–348. [Google Scholar]
- Gennerich, A; Carter, AP; Reck-Peterson, SL; Vale, RD. Force-induced bidirectional stepping of cytoplasmic dynein. Cell 2007, 131, 952–965. [Google Scholar]
- Gennerich, A; Vale, RD. Walking the walk: how kinesin and dynein coordinate their steps. Curr Opin Cell Biol 2009, 21, 59–67. [Google Scholar]
- Hirokawa, N; Takemura, R. Kinesin superfamily proteins and their various functions and dynamics. Exp Cell Res 2004, 301, 50–59. [Google Scholar]
- King, SM. The dynein microtubule motor. Biochim Biophys Acta 2000, 1496, 60–75. [Google Scholar]
- Kamal, A; Goldstein, LS. Connecting vesicle transport to the cytoskeleton. Curr Opin Cell Biol 2000, 12, 503–508. [Google Scholar]
- Alexandre, H; van Cauwenberge, A; Tsukitani, Y; Mulnard, J. Pleiotropic effect of okadaic acid on maturing mouse oocytes. Development 1991, 112, 971–980. [Google Scholar]
- Paul, VJ; Arthur, KE; Ritson-Williams, R; Ross, C; Sharp, K. Chemical defenses: from compounds to communities. Biol Bull 2007, 213, 226–251. [Google Scholar]
- Hodgson, LM. Antimicrobial and antineoplastic activity in some south Florida seaweeds. Bot Mar 1984, 27, 387–390. [Google Scholar]
- Paul, VJ; Fenical, W. Chemical defense in tropical green algae, order Caulerpales. Mar Ecol Prog Ser 1986, 34, 157–169. [Google Scholar]
- Smyrniotopoulos, V; Abatis, D; Tziveleka, LA; Tsitsimpikou, C; Roussis, V; Loukis, A; Vagias, C. Acetylene sesquiterpenoid esters from the green alga Caulerpa prolifera. J Nat Prod 2003, 66, 21–24. [Google Scholar]
- Freile-Pelegrin, Y; Morales, JL. Antibacterial activity in marine algae from the coast of Yucatan, Mexico. Bot Mar 2004, 47, 140–146. [Google Scholar]
- Fischel, JL; Lemee, R; Formento, P; Caldani, C; Moll, JL; Pesando, D; Meinesz, A; Grelier, P; Pietra, P; Guerriero, A; et al. Cell growth inhibitory effects of caulerpenyne, a sesquiterpenoid from the marine algae Caulerpa taxifolia. Anticancer Res 1995, 15, 2155–2160. [Google Scholar]
- Cavas, L; Baskin, Y; Yurdakoc, K; Olgun, N. Antiproliferative and newly attributed apoptotic activities from an invasive marine alga: Caulerpa racemosa var. cylindracea. J Exp Mar Biol Ecol 2006, 339, 111–119. [Google Scholar]
- Mozzachiodi, R; Scuri, R; Roberto, M; Brunelli, M. Caulerpenyne, a toxin from the seaweed Caulerpa taxifolia, depresses afterhyperpolarization in invertebrate neurons. NeuroScience 2001, 107, 519–526. [Google Scholar]
- Barbier, P; Guise, S; Huitorel, P; Amade, P; Pesando, D; Briand, C; Peyrot, V. Caulerpenyne from Caulerpa taxifolia has an antiproliferative activity on tumor cell line SK-N-SH and modifies the microtubule network. Life Sci 2001, 70, 415–429. [Google Scholar]
- Pettit, GR. The dolastatins. Fortschr Chem Org Naturst 1997, 70, 1–79. [Google Scholar]
- Bai, RL; Pettit, GR; Hamel, E. Binding of dolastatin 10 to tubulin at a distinct site for peptide antimitotic agents near the exchangeable nucleotide and vinca alkaloid sites. J Biol Chem 1990, 265, 17141–17149. [Google Scholar]
- Bai, R; Friedman, SJ; Pettit, GR; Hamel, E. Dolastatin 15, a potent antimitotic depsipeptide derived from Dolabella auricularia. Interaction with tubulin and effects of cellular microtubules. Biochem Pharmacol 1992, 43, 2637–2645. [Google Scholar]
- Cruz-Monserrate, Z; Mullaney, JT; Harran, PG; Pettit, GR; Hamel, E. Dolastatin 15 binds in the vinca domain of tubulin as demonstrated by Hummel-Dreyer chromatography. Eur J Biochem 2003, 270, 3822–3828. [Google Scholar]
- Simon-Levert, A; Arrault, A; Bontemps-Subielos, N; Canal, C; Banaigs, B. Meroterpenes from the ascidian Aplidium aff. densum. J Nat Prod 2005, 68, 1412–1415. [Google Scholar]
- Simon-Levert, A; Aze, A; Bontemps-Subielos, N; Banaigs, B; Geneviere, AM. Antimitotic activity of methoxyconidiol, a meroterpene isolated from an ascidian. Chem Biol Interact 2007, 168, 106–116. [Google Scholar]
- O’Brien, ET; Jacobs, RS; Wilson, L. Inhibition of bovine brain microtubule assembly in vitro by stypoldione. Mol Pharmacol 1983, 24, 493–499. [Google Scholar]
- White, SJ; Jacobs, RS. Effect of stypoldione on cell cycle progression, DNA and protein synthesis, and cell division in cultured sea urchin embryos. Mol Pharmacol 1983, 24, 500–508. [Google Scholar]
- O’Brien, ET; Asai, DJ; Jacobs, RS; Wilson, L. Selective inhibition of cytokinesis in sea urchin embryos by low concentrations of stypoldione, a marine natural product that reacts with sulfhydryl groups. Mol Pharmacol 1989, 35, 635–642. [Google Scholar]
- Yanagimachi, R. Knobil, E, Neil, J, Eds.; Mammalian fertilization. In The Physiology of Reproduction; Raven press: New York, NY, USA, 1994; pp. 189–317. [Google Scholar]
- Grudzinskas, JG; Yovich, JL. Gametes: The Spermatozoon; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Stratmhann, RR. Why life histories evolve differently in the sea. Soc Integ Comp Biol 1990, 30, 197–207. [Google Scholar]
- Tosti, E. Sperm activation in species with external fertilisation. Zygote 1994, 2, 359–361. [Google Scholar]
- Sanders, S; Debuse, M. Endocrine and Reproductive Systems, 2nd ed; Elsevier Science Ltd: London, UK, 2003; pp. 149–156. [Google Scholar]
- Yan, HH; Mruk, DD; Lee, WM; Cheng, CY. Ectoplasmic specialization: a friend or a foe of spermatogenesis. Bioessays 2007, 29, 36–48. [Google Scholar]
- Cheng, CY; Mruk, DD. Cell junction dynamics in the testis: Sertoli-germ cell interactions and male contraceptive development. Physiol Rev 2002, 82, 825–874. [Google Scholar]
- Lee, NP; Cheng, CY. Ectoplasmic specialization, a testis-specific cell-cell actin-based adherens junction type: is this a potential target for male contraceptive development. Hum Reprod Update 2004, 10, 349–369. [Google Scholar]
- Cheng, CY; Mruk, D; Silvestrini, B; Bonanomi, M; Wong, CH; Siu, MK; Lee, NP; Lui, WY; Mo, MY. AF-2364 [1-(2,4-dichlorobenzyl)-1H-indazole-3-carbohydrazide] is a potential male contraceptive: a review of recent data. Contraception 2005, 72, 251–261. [Google Scholar]
- Redenbach, DM; Vogl, AW. Microtubule polarity in Sertoli cells: a model for microtubule-based spermatid transport. Eur J Cell Biol 1991, 54, 277–290. [Google Scholar]
- Redenbach, DM; Boekelheide, K; Vogl, AW. Binding between mammalian spermatid-ectoplasmic specialization complexes and microtubules. Eur J Cell Biol 1992, 59, 433–448. [Google Scholar]
- Beach, SF; Vogl, AW. Spermatid translocation in the rat seminiferous epithelium: coupling membrane trafficking machinery to a junction plaque. Biol Reprod 1999, 60, 1036–1046. [Google Scholar]
- Guttman, JA; Kimel, GH; Vogl, AW. Dynein and plus-end microtubule-dependent motors are associated with specialized Sertoli cell junction plaques (ectoplasmic specializations). J Cell Sci 2000, 113 Pt 12, 2167–2176. [Google Scholar]
- Velichkova, M; Guttman, J; Warren, C; Eng, L; Kline, K; Vogl, AW; Hasson, T. A human homologue of Drosophila kelch associates with myosin-VIIa in specialized adhesion junctions. Cell Motil Cytoskeleton 2002, 51, 147–164. [Google Scholar]
- Collins, CA; Vallee, RB. Preparation of microtubules from rat liver and testis: cytoplasmic dynein is a major microtubule associated protein. Cell Motil Cytoskeleton 1989, 14, 491–500. [Google Scholar]
- Hall, ES; Eveleth, J; Jiang, C; Redenbach, DM; Boekelheide, K. Distribution of the microtubule-dependent motors cytoplasmic dynein and kinesin in rat testis. Biol Reprod 1992, 46, 817–828. [Google Scholar]
- Miller, MG; Mulholland, DJ; Vogl, AW. Rat testis motor proteins associated with spermatid translocation (dynein) and spermatid flagella (kinesin-II). Biol Reprod 1999, 60, 1047–1056. [Google Scholar]
- Silverman-Gavrila, RV; Forer, A. Evidence that actin and myosin are involved in the poleward flux of tubulin in metaphase kinetochore microtubules of crane-fly spermatocytes. J Cell Sci 2000, 113 Pt 4, 597–609. [Google Scholar]
- Ianora, A; Miralto, A; Buttino, I; Romano, G; Poulet, SA. First evidence of some dinoflagellates reducing male copepod fertilization capacity. Limnol Oceanogr 1999, 44, 147–153. [Google Scholar]
- Ishijima, S; Mohri, H. A quantitative description of flagellar movement in golden hamster spermatozoa. J Exp Biol 1985, 114, 463–475. [Google Scholar]
- Mohri, H; Awano, M; Ishijima, S. Maturation and capacitation of mammalian spermatozoa. Prog Clin Biol Res 1989, 294, 53–62. [Google Scholar]
- Mohri, H. Role of tubulin and dynein in spermatozoan motility. Mol Reprod Dev 1993, 36, 221–223. [Google Scholar]
- Summers, KE; Gibbons, IR. Adenosine triphosphate-induced sliding of tubules in trypsin-treated flagella of sea-urchin sperm. Proc Natl Acad Sci USA 1971, 68, 3092–3096. [Google Scholar]
- Azamar, Y; Uribe, S; Mujica, A. F-actin involvement in guinea pig sperm motility. Mol Reprod Dev 2007, 74, 312–320. [Google Scholar]
- Tosti, E; Romano, G; Buttino, I; Cuomo, A; Ianora, A; Miralto, A. Bioactive aldehydes from diatoms block the fertilization current in ascidian oocytes. Mol Reprod Dev 2003, 66, 72–80. [Google Scholar]
- Su, YH; Vacquier, VD. A flagellar K(+)-dependent Na(+)/Ca(2+) exchanger keeps Ca(2+) low in sea urchin spermatozoa. Proc Natl Acad Sci USA 2002, 99, 6743–6748. [Google Scholar]
- Brokaw, CJ; Josslin, R; Bobrow, L. Calcium ion regulation of flagellar beat symmetry in reactivated sea urchin spermatozoa. Biochem Biophys Res Commun 1974, 58, 795–800. [Google Scholar]
- Brokaw, CJ. Calcium-induced asymmetrical beating of triton-demembranated sea urchin sperm flagella. J Cell Biol 1979, 82, 401–411. [Google Scholar]
- Gibbons, BH; Gibbons, IR. Calcium-induced quiescence in reactivated sea urchin sperm. J Cell Biol 1980, 84, 13–27. [Google Scholar]
- Boni, R; Gualtieri, R; Talevi, R; Tosti, E. Calcium and other ion dynamics during gamete maturation and fertilization. Theriogenology 2007, 68(Suppl 1), S156–S164. [Google Scholar]
- Morton, BE; Fraser, CF; Thenawidjaja, M; Albagli, L; Rayner, MD. Potent inhibition of sperm motility by palytoxin. Exp Cell Res 1982, 140, 261–265. [Google Scholar]
- Monroy, A. Processes controlling sperm-egg fusion. Eur J Biochem 1985, 152, 51–56. [Google Scholar]
- Voronina, E; Wessel, GM. The regulation of oocyte maturation. Curr Top Dev Biol 2003, 58, 53–110. [Google Scholar]
- Grudzinskas, JG; Yovich, JL. Gametes: The Oocyte; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Bates, WR; Nishida, H. Developmental roles of nuclear complex factors released during oocyte maturation in the ascidians Halocynthia roretzi and Boltenia villosa. Zoolog Sci 1998, 15, 69–76. [Google Scholar]
- Elder, K; Dale, B. In Vitro Fertilization, 2nd ed; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Mehlmann, LM. Stops and starts in mammalian oocytes: recent advances in understanding the regulation of meiotic arrest and oocyte maturation. Reproduction 2005, 130, 791–799. [Google Scholar]
- Yamashita, M; Mita, K; Yoshida, N; Kondo, T. Molecular mechanisms of the initiation of oocyte maturation: general and species-specific aspects. Prog Cell Cycle Res 2000, 4, 115–129. [Google Scholar]
- Cuomo, A; Di Cristo, C; Paolucci, M; Di Cosmo, A; Tosti, E. Calcium currents correlate with oocyte maturation during the reproductive cycle in Octopus vulgaris. J Exp Zoolog A Comp Exp Biol 2005, 303, 193–202. [Google Scholar]
- Cuomo, A; Silvestre, F; De Santis, R; Tosti, E. Ca2+ and Na+ current patterns during oocyte maturation, fertilization, and early developmental stages of Ciona intestinalis. Mol Reprod Dev 2006, 73, 501–511. [Google Scholar]
- Tosti, E. Calcium ion currents mediating oocyte maturation events. Reprod Biol Endocrinol 2006, 4, 26. [Google Scholar]
- Cui, XS; Kim, NH. Maternally derived transcripts: identification and characterisation during oocyte maturation and early cleavage. Reprod Fertil Dev 2007, 19, 25–34. [Google Scholar]
- Silvestre, F; Cuomo, A; Tosti, E. Ion current activity and molecules modulating maturation and growth stages of ascidian (Ciona intestinalis) oocytes. Mol Reprod Dev 2009, 76, 1084–1093. [Google Scholar]
- Prodon, F; Chenevert, J; Sardet, C. Establishment of animal-vegetal polarity during maturation in ascidian oocytes. Dev Biol 2006, 290, 297–311. [Google Scholar]
- Gard, DL. Ectopic spindle assembly during maturation of Xenopus oocytes: evidence for functional polarization of the oocyte cortex. Dev Biol 1993, 159, 298–310. [Google Scholar]
- Kloc, M; Etkin, LD. RNA localization mechanisms in oocytes. J Cell Sci 2005, 118, 269–282. [Google Scholar]
- Terasaki, M. Redistribution of cytoplasmic components during germinal vesicle breakdown in starfish oocytes. J Cell Sci 1994, 107 Pt 7, 1797–1805. [Google Scholar]
- Bobinnec, Y; Marcaillou, C; Morin, X; Debec, A. Dynamics of the endoplasmic reticulum during early development of Drosophila melanogaster. Cell Motil Cytoskeleton 2003, 54, 217–225. [Google Scholar]
- Sun, QY; Wu, GM; Lai, L; Park, KW; Cabot, R; Cheong, HT; Day, BN; Prather, RS; Schatten, H. Translocation of active mitochondria during pig oocyte maturation, fertilization and early embryo development in vitro. Reproduction 2001, 122, 155–163. [Google Scholar]
- Schatten, H; Walter, M; Biessmann, H; Schatten, G. Microtubules are required for centrosome expansion and positioning while microfilaments are required for centrosome separation in sea urchin eggs during fertilization and mitosis. Cell Motil Cytoskeleton 1988, 11, 248–259. [Google Scholar]
- Calarco, PG. The role of microfilaments in early meiotic maturation of mouse oocytes. Microsc Microanal 2005, 11, 146–153. [Google Scholar]
- Wessel, GM; Conner, SD; Berg, L. Cortical granule translocation is microfilament mediated and linked to meiotic maturation in the sea urchin oocyte. Development 2002, 129, 4315–4325. [Google Scholar]
- Connors, SA; Kanatsu-Shinohara, M; Schultz, RM; Kopf, GS. Involvement of the cytoskeleton in the movement of cortical granules during oocyte maturation, and cortical granule anchoring in mouse eggs. Dev Biol 1998, 200, 103–115. [Google Scholar]
- Verlhac, MH; Dumont, J. Interactions between chromosomes, microfilaments and microtubules revealed by the study of small GTPases in a big cell, the vertebrate oocyte. Mol Cell Endocrinol 2008, 282, 12–17. [Google Scholar]
- Pielak, RM; Gaysinskaya, VA; Cohen, WD. Formation and function of the polar body contractile ring in Spisula. Dev Biol 2004, 269, 421–432. [Google Scholar]
- Miyazaki, A; Kamitsubo, E; Nemoto, SI. Premeiotic aster as a device to anchor the germinal vesicle to the cell surface of the presumptive animal pole in starfish oocytes. Dev Biol 2000, 218, 161–171. [Google Scholar]
- Gard, DL. Organization, nucleation, and acetylation of microtubules in Xenopus laevis oocytes: a study by confocal immunofluorescence microscopy. Dev Biol 1991, 143, 346–362. [Google Scholar]
- Miyazaki, A; Kato, KH; Nemoto, S. Role of microtubules and centrosomes in the eccentric relocation of the germinal vesicle upon meiosis reinitiation in sea-cucumber oocytes. Dev Biol 2005, 280, 237–247. [Google Scholar]
- Sardet, C; Paix, A; Prodon, F; Dru, P; Chenevert, J. From oocyte to 16-cell stage: cytoplasmic and cortical reorganizations that pattern the ascidian embryo. Dev Dyn 2007, 236, 1716–1731. [Google Scholar]
- Yang, HY; Mains, PE; McNally, FJ. Kinesin-1 mediates translocation of the meiotic spindle to the oocyte cortex through KCA-1, a novel cargo adapter. J Cell Biol 2005, 169, 447–457. [Google Scholar]
- Lutz, DA; Hamaguchi, Y; Inoue, S. Micromanipulation studies of the asymmetric positioning of the maturation spindle in Chaetopterus sp. oocytes: I. Anchorage of the spindle to the cortex and migration of a displaced spindle. Cell Motil Cytoskeleton 1988, 11, 83–96. [Google Scholar]
- Weber, KL; Sokac, AM; Berg, JS; Cheney, RE; Bement, WM. A microtubule-binding myosin required for nuclear anchoring and spindle assembly. Nature 2004, 431, 325–329. [Google Scholar]
- Maro, B; Verlhac, MH. Polar body formation: new rules for asymmetric divisions. Nat Cell Biol 2002, 4, E281–283. [Google Scholar]
- Terada, Y; Simerly, C; Schatten, G. Microfilament stabilization by jasplakinolide arrests oocyte maturation, cortical granule exocytosis, sperm incorporation cone resorption, and cell-cycle progression, but not DNA replication, during fertilization in mice. Mol Reprod Dev 2000, 56, 89–98. [Google Scholar]
- Terada, Y; Fukaya, T; Yajima, A. Localization of microfilaments during oocyte maturation of golden hamster. Mol Reprod Dev 1995, 41, 486–492. [Google Scholar]
- Kim, NH; Cho, SK; Choi, SH; Kim, EY; Park, SP; Lim, JH. The distribution and requirements of microtubules and microfilaments in bovine oocytes during in vitro maturation. Zygote 2000, 8, 25–32. [Google Scholar]
- Kim, NH; Chung, HM; Cha, KY; Chung, KS. Microtubule and microfilament organization in maturing human oocytes. Hum Reprod 1998, 13, 2217–2222. [Google Scholar]
- Ryabova, LV; Betina, MI; Vassetzky, SG. Influence of cytochalasin B on oocyte maturation in Xenopus laevis. Cell Differ 1986, 19, 89–96. [Google Scholar]
- Gard, DL; Cha, BJ; Roeder, AD. F-actin is required for spindle anchoring and rotation in Xenopus oocytes: a re-examination of the effects of cytochalasin B on oocyte maturation. Zygote 1995, 3, 17–26. [Google Scholar]
- Poulet, SA; Ianora, A; Miralto, A; Meijer, L. Do diatoms arrest embryonic development in copepods. Mar Ecol Prog Ser 1994, 111, 79–86. [Google Scholar]
- Kyozuka, K; Chun, JT; Puppo, A; Gragnaniello, G; Garante, E; Santella, L. Guanine nucleotides in the meiotic maturation of starfish oocytes: regulation of the actin cytoskeleton and of Ca(2+) signaling. PLoS One 2009, 4, e6296. [Google Scholar]
- Liu, H; Namikoshi, M; Akano, K; Kobayashi, H; Nagai, H; Yao, X. Seven new meroditerpenoids, from the marine sponge Strongylophora strongylata, that inhibited the maturation of starfish oocytes. J Asian Nat Prod Res 2005, 7, 661–670. [Google Scholar]
- Pondaven, P; Meijer, L; Bialojan, C. La microinjection d’acide okadaique, un inhibiteur de phosphoproteine phosphatase, induit la maturation de l’ovocyte de l’etoile de mer. C r hebd Seances Acad Sci (Paris) 1989, 309, 563–569. [Google Scholar]
- Picard, A; Capony, JP; Brautigan, DL; Doree, M. Involvement of protein phosphatases 1 and 2A in the control of M phase-promoting factor activity in starfish. J Cell Biol 1989, 109, 3347–3354. [Google Scholar]
- Rime, H; Huchon, D; Jessus, C; Goris, J; Merlevede, W; Ozon, R. Characterization of MPF activation by okadaic acid in Xenopus oocyte. Cell Differ Dev 1990, 29, 47–58. [Google Scholar]
- Rime, H; Ozon, R. Protein phosphatases are involved in the in vivo activation of histone H1 kinase in mouse oocyte. Dev Biol 1990, 141, 115–122. [Google Scholar]
- Gavin, AC; Tsukitani, Y; Schorderet-Slatkine, S. Induction of M-phase entry of prophase-blocked mouse oocytes through microinjection of okadaic acid, a specific phosphatase inhibitor. Exp Cell Res 1991, 192, 75–81. [Google Scholar]
- Sasseville, M; Cote, N; Guillemette, C; Richard, FJ. New insight into the role of phosphodiesterase 3A in porcine oocyte maturation. BMC Dev Biol 2006, 6, 47. [Google Scholar]
- Swain, JE; Ding, J; Brautigan, DL; Villa-Moruzzi, E; Smith, GD. Proper chromatin condensation and maintenance of histone H3 phosphorylation during mouse oocyte meiosis requires protein phosphatase activity. Biol Reprod 2007, 76, 628–638. [Google Scholar]
- Breitbart, H; Cohen, G; Rubinstein, S. Role of actin cytoskeleton in mammalian sperm capacitation and the acrosome reaction. Reproduction 2005, 129, 263–268. [Google Scholar]
- Lin, M; Hess, R; Aitken, RJ. Induction of sperm maturation in vitro in epididymal cell cultures of the tammar wallaby (Macropus eugenii): disruption of motility initiation and sperm morphogenesis by inhibition of actin polymerization. Reproduction 2002, 124, 107–117. [Google Scholar]
- Talbot, P; Kleve, MG. Hamster sperm cross react with antiactin. J Exp Zool 1978, 204, 131–136. [Google Scholar]
- Camatini, M; Anelli, G; Casale, A. Identification of actin in boar spermatids and spermatozoa by immunoelectron microscopy. Eur J Cell Biol 1986, 42, 311–318. [Google Scholar]
- Flaherty, SP; Winfrey, VP; Olson, GE. Localization of actin in human, bull, rabbit, and hamster sperm by immunoelectron microscopy. Anat Rec 1988, 221, 599–610. [Google Scholar]
- Olson, GE; Winfrey, VP. Changes in actin distribution during sperm development in the opossum, Monodelphis domestica. Anat Rec 1991, 230, 209–217. [Google Scholar]
- Moreno-Fierros, L; Hernandez, EO; Salgado, ZO; Mujica, A. F-actin in guinea pig spermatozoa: its role in calmodulin translocation during acrosome reaction. Mol Reprod Dev 1992, 33, 172–181. [Google Scholar]
- Yagi, A; Paranko, J. Actin, alpha-actinin, and spectrin with specific associations with the postacrosomal and acrosomal domains of bovine spermatozoa. Anat Rec 1995, 241, 77–87. [Google Scholar]
- Sanchez-Gutierrez, M; Contreras, RG; Mujica, A. Cytochalasin-D retards sperm incorporation deep into the egg cytoplasm but not membrane fusion with the egg plasma membrane. Mol Reprod Dev 2002, 63, 518–528. [Google Scholar]
- Kumakiri, J; Oda, S; Kinoshita, K; Miyazaki, S. Involvement of Rho family G protein in the cell signaling for sperm incorporation during fertilization of mouse eggs: inhibition by Clostridium difficile toxin B. Dev Biol 2003, 260, 522–535. [Google Scholar]
- Schatten, G; Schatten, H; Spector, I; Cline, C; Paweletz, N; Simerly, C; Petzelt, C. Latrunculin inhibits the microfilament-mediated processes during fertilization, cleavage and early development in sea urchins and mice. Exp Cell Res 1986, 166, 191–208. [Google Scholar]
- Garbers, DL. Molecular basis of fertilization. Annu Rev Biochem 1989, 58, 719–742. [Google Scholar]
- Johnson, MH; Eager, D; Muggleton-Harris, A; Grave, HM. Mosaicism in organisation concanavalin A receptors on surface membrane of mouse egg. Nature 1975, 257, 321–322. [Google Scholar]
- Jegou, A; Pincet, F; Perez, E; Wolf, JP; Ziyyat, A; Gourier, C. Mapping mouse gamete interaction forces reveal several oocyte membrane regions with different mechanical and adhesive properties. Langmuir 2008, 24, 1451–1458. [Google Scholar]
- Ebensperger, C; Barros, DC. Changes at the hamster oocyte surface from the germinal vesicle stage to ovulation. Gamete Res 1984, 9, 387–397. [Google Scholar]
- Tilney, LG; Hatano, S; Ishikawa, H; Mooseker, MS. The polymerization of actin: its role in the generation of the acrosomal process of certain echinoderm sperm. J Cell Biol 1973, 59, 109–126. [Google Scholar]
- Tilney, LG; Mooseker, MS. Actin filament-membrane attachment: are membrane particles involved. J Cell Biol 1976, 71, 402–416. [Google Scholar]
- Stein, KK; Primakoff, P; Myles, D. Sperm-egg fusion: events at the plasma membrane. J Cell Sci 2004, 117, 6269–6274. [Google Scholar]
- Schatten, G; Schatten, H. Effects of motility inhibitors during sea urchin fertilization: microfilament inhibitors prevent sperm incorporation and restructuring of fertilized egg cortex, whereas microtubule inhibitors prevent pronuclear migrations. Exp Cell Res 1981, 135, 311–330. [Google Scholar]
- Hart, NH; Becker, KA; Wolenski, JS. The sperm entry site during fertilization of the zebrafish egg: localization of actin. Mol Reprod Dev 1992, 32, 217–228. [Google Scholar]
- Wilson, SM; Pickett-Heaps, JD; West, JA. Fertilization and the cytoskeleton in the red alga Bostrychia moritziana (Rhodomelaceae, Rhodophyta). Eur J Phycol 2002, 37, 509–522. [Google Scholar]
- Puppo, A; Chun, JT; Gragnaniello, G; Garante, E; Santella, L. Alteration of the cortical actin cytoskeleton deregulates Ca2+ signaling, monospermic fertilization, and sperm entry. PLoS One 2008, 3, e3588. [Google Scholar]
- McAvey, BA; Wortzman, GB; Williams, CJ; Evans, JP. Involvement of calcium signaling and the actin cytoskeleton in the membrane block to polyspermy in mouse eggs. Biol Reprod 2002, 67, 1342–1352. [Google Scholar]
- Misamore, MJ; Lynn, JW. Role of the cytoskeleton in sperm entry during fertilization in the freshwater bivalve Dreissena polymorpha. Biol Bull 2000, 199, 144–156. [Google Scholar]
- Xu, Z; Kopf, GS; Schultz, RM. Involvement of inositol 1,4,5-trisphosphate-mediated Ca2+ release in early and late events of mouse egg activation. Development 1994, 120, 1851–1859. [Google Scholar]
- Tosti, E; Boni, R. Electrical events during gamete maturation and fertilization in animals and humans. Hum Reprod Update 2004, 10, 53–65. [Google Scholar]
- Lim, D; Lange, K; Santella, L. Activation of oocytes by latrunculin A. Faseb J 2002, 16, 1050–1056. [Google Scholar]
- Satoh, N. Developmental Biology of Ascidians; Cambridge University Press: New York, NY, USA, 1994. [Google Scholar]
- Speksnijder, JE; Corson, DW; Sardet, C; Jaffe, LF. Free calcium pulses following fertilization in the ascidian egg. Dev Biol 1989, 135, 182–190. [Google Scholar]
- Speksnijder, JE; Sardet, C; Jaffe, LF. The activation wave of calcium in the ascidian egg and its role in ooplasmic segregation. J Cell Biol 1990, 110, 1589–1598. [Google Scholar]
- McDougall, A; Sardet, C. Function and characteristics of repetitive calcium waves associated with meiosis. Curr Biol 1995, 5, 318–328. [Google Scholar]
- Roegiers, F; McDougall, A; Sardet, C. The sperm entry point defines the orientation of the calcium-induced contraction wave that directs the first phase of cytoplasmic reorganization in the ascidian egg. Development 1995, 121, 3457–3466. [Google Scholar]
- Sardet, C; Speksnijder, J; Inoue, S; Jaffe, L. Fertilization and ooplasmic movements in the ascidian egg. Development 1989, 105, 237–249. [Google Scholar]
- Chiba, S; Miki, Y; Ashida, K; Wada, MR; Tanaka, KJ; Shibata, Y; Nakamori, R; Nishikata, T. Interactions between cytoskeletal components during myoplasm rearrangement in ascidian eggs. Dev Growth Differ 1999, 41, 265–272. [Google Scholar]
- Glotzer, M. The molecular requirements for cytokinesis. Science 2005, 307, 1735–1739. [Google Scholar]
- Zhong, ZS; Huo, LJ; Liang, CG; Chen, DY; Sun, QY. Small GTPase RhoA is required for ooplasmic segregation and spindle rotation, but not for spindle organization and chromosome separation during mouse oocyte maturation, fertilization, and early cleavage. Mol Reprod Dev 2005, 71, 256–261. [Google Scholar]
- Maro, B; Johnson, MH; Pickering, SJ; Flach, G. Changes in actin distribution during fertilization of the mouse egg. J Embryol Exp Morphol 1984, 81, 211–237. [Google Scholar]
- Le Guen, P; Crozet, N; Huneau, D; Gall, L. Distribution and role of microfilaments during early events of sheep fertilization. Gamete Res 1989, 22, 411–425. [Google Scholar]
- Caldwell, GS; Olive, PJ; Bentley, MG. Inhibition of embryonic development and fertilization in broadcast spawning marine invertebrates by water soluble diatom extracts and the diatom toxin 2-trans,4-trans decadienal. Aquat Toxicol 2002, 60, 123–137. [Google Scholar]
- Buttino, I; Miralto, A; Ianora, A; Romano, G; Poulet, SA. Water-soluble extracts of the diatom Thalassiosira rotula induce aberrations in embryonic tubulin organisation of the sea urchin Paracentrotus lividus. Mar Biol 1999, 134, 147–154. [Google Scholar]
- Van Blerkom, J; Davis, P; Merriam, J; Sinclair, J. Nuclear and cytoplasmic dynamics of sperm penetration, pronuclear formation and microtubule organization during fertilization and early preimplantation development in the human. Hum Reprod Update 1995, 1, 429–461. [Google Scholar]
- Sutovsky, P; Navara, CS; Schatten, G. Fate of the sperm mitochondria, and the incorporation, conversion, and disassembly of the sperm tail structures during bovine fertilization. Biol Reprod 1996, 55, 1195–1205. [Google Scholar]
- Kim, NH; Chung, KS; Day, BN. The distribution and requirements of microtubules and microfilaments during fertilization and parthenogenesis in pig oocytes. J Reprod Fertil 1997, 111, 143–149. [Google Scholar]
- Schatten, H; Schatten, G. Motility and centrosomal organization during sea urchin and mouse fertilization. Cell Motil Cytoskeleton 1986, 6, 163–175. [Google Scholar]
- Mabuchi, I. Cleavage furrow: timing of emergence of contractile ring actin filaments and establishment of the contractile ring by filament bundling in sea urchin eggs. J Cell Sci 1994, 107 Pt 7, 1853–1862. [Google Scholar]
- Noguchi, T; Mabuchi, I. Reorganization of actin cytoskeleton at the growing end of the cleavage furrow of Xenopus egg during cytokinesis. J Cell Sci 2001, 114, 401–412. [Google Scholar]
- Noguchi, T; Arai, R; Motegi, F; Nakano, K; Mabuchi, I. Contractile ring formation in Xenopus egg and fission yeast. Cell Struct Funct 2001, 26, 545–554. [Google Scholar]
- Takayama, M; Noguchi, T; Yamashiro, S; Mabuchi, I. Microtuble organization in Xenopus eggs during the first cleavage and its role in cytokinesis. Cell Struct Funct 2002, 27, 163–171. [Google Scholar]
- Murthy, K; Wadsworth, P. Dual role for microtubules in regulating cortical contractility during cytokinesis. J Cell Sci 2008, 121, 2350–2359. [Google Scholar]
- d’Ippolito, G; Romano, G; Iadicicco, O; Miralto, A; Ianora, A; Cimino, G; Fontana, A. New birth-control aldehydes from the marine diatom Skeletonema costatum: characterization and biogenesis. Tetrahedron Lett 2002, 43, 6133–6136. [Google Scholar]
- Hansen, E; Eilertsen, HC; Ernstsen, A; Geneviere, AM. Anti-mitotic activity towards sea urchin embryos in extracts from the marine haptophycean Phaeocystis pouchetii (Hariot) Lagerheim collected along the coast of northern Norway. Toxicon 2003, 41, 803–812. [Google Scholar]
- Hansen, E; Even, Y; Geneviere, AM. The α,β,γ,δ-Unsaturated Aldehyde 2-trans-4-trans-Decadienal Disturbs DNA Replication and Mitotic Events in Early Sea Urchin Embryos. Toxicol Sci 2004, 81, 190–197. [Google Scholar]
- Grace, KJ; Medina, M; Jacobs, RS; Wilson, L. Selective inhibition of cytokinesis in sea urchin embryos by the marine natural product pseudopterolide. Mol Pharmacol 1992, 41, 631–638. [Google Scholar]
- Colman, JR; Twiner, MJ; Hess, P; McMahon, T; Satake, M; Yasumoto, T; Doucette, GJ; Ramsdell, JS. Teratogenic effects of azaspiracid-1 identified by microinjection of Japanese medaka (Oryzias latipes) embryos. Toxicon 2005, 45, 881–890. [Google Scholar]
- Chae, HD; Choi, TS; Kim, BM; Jung, JH; Bang, YJ; Shin, DY. Oocyte-based screening of cytokinesis inhibitors and identification of pectenotoxin-2 that induces Bim/Bax-mediated apoptosis in p53-deficient tumors. Oncogene 2005, 24, 4813–4819. [Google Scholar]
- Pesando, D; Lemee, R; Ferrua, C; Amade, P; Girard, JP. Effects of caulerpenyne, the major toxin from Caulerpa taxifolia on mechanisms related to sea urchin egg cleavage. Aquat Toxicol 1996, 35, 139–155. [Google Scholar]
- Pesando, D; Huitorel, P; Dolcini, V; Amade, P; Girard, JP. Caulerpenyne interferes with microtubule-dependent events during the first mitotic cycle of sea urchin eggs. Eur J Cell Biol 1998, 77, 19–26. [Google Scholar]
- Semenova, MN; Kiselyov, A; Semenov, VV. Sea urchin embryo as a model organism for the rapid functional screening of tubulin modulators. Biotechniques 2006, 40, 765–774. [Google Scholar]
- Bhakuni, DS; Rawat, DS. Bioactive Marine Natural Products; Springer-Verlag and Anamaya Publishers: New York, NY, USA, 2005. [Google Scholar]
- Blunt, JW; Copp, BR; Munro, MH; Northcote, PT; Prinsep, MR. Marine natural products. Nat Prod Rep 2005, 22, 15–61. [Google Scholar]
- Amador, ML; Jimeno, J; Paz-Ares, L; Cortes-Funes, H; Hidalgo, M. Progress in the development and acquisition of anticancer agents from marine sources. Ann Oncol 2003, 14, 1607–1615. [Google Scholar]
- Singh, R; Sharma, M; Joshi, P; Rawat, DS. Clinical status of anti-cancer agents derived from marine sources. Anticancer Agents Med Chem 2008, 8, 603–617. [Google Scholar]
- Haefner, B. Drugs from the deep: marine natural products as drug candidates. Drug Discov Today 2003, 8, 536–544. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
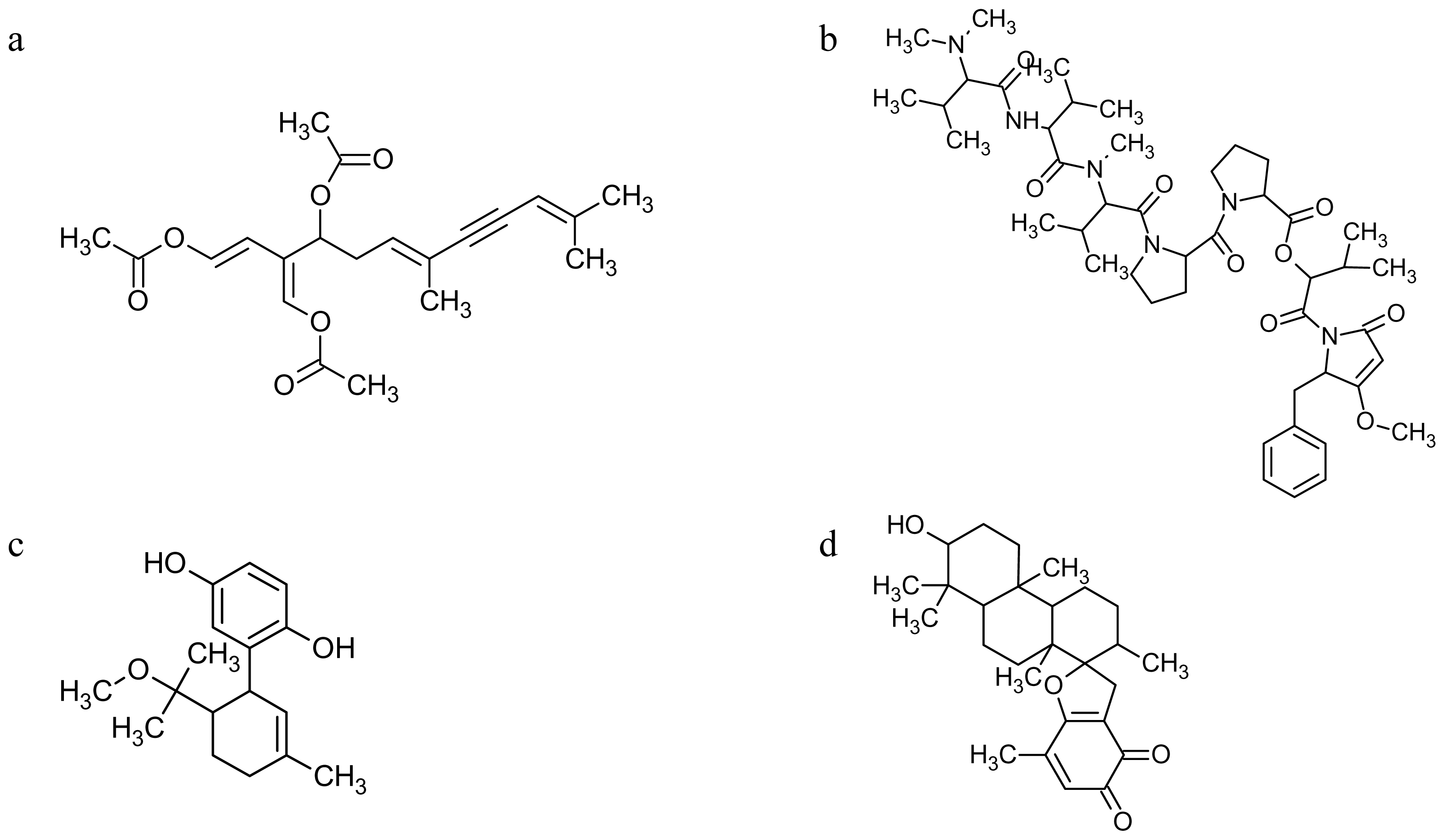
| Drug Name | Drug Source | Cellular Target | Stage/Event Affected | References |
|---|---|---|---|---|
| Azaspiracid | Dinoflagellates | Unknown | Early development | [240] |
| Calyculin | Sponges | Protein phosphatases | Oogenesis | [46] |
| Caulerpenyne | Algae | Microtubules | Early development | [92,242,243] |
| 2E,4E-Decadienal | Diatoms | Cytoskeleton, calcium signaling etc. | Sperm motility, oogenesis, fertilization, early development | [64,132,177,225–238] |
| Dolastatin | Molluscs | Microtubules | Early development | [244] |
| Geodiamolide | Sponges | Microfilaments | Early development | [47] |
| Jasplakinolide | Sponges | Microfilaments | Oogenesis | [171,178,207,208] |
| Latrunculin | Sponges | Microfilaments | Spermatogenesis, acrosome reaction, fertilization, early development | [125,197, 205–208] |
| Methoxyconidiol | Ascidians | Unknown | Early development | [104] |
| Mycalolide | Sponges | Microfilaments | Fertilization | [207] |
| Okadaic acid | Dinoflagellates | Protein phosphatases | Oogenesis | [89,180–186] |
| Palytoxin | Dinoflagellates? | Sodium pump | Sperm motility | [138] |
| Pectenotoxin | Dinoflagellates | Microfilaments | Early development | [241] |
| Pseudopterolide | Soft corals | Unknown | Early development | [239] |
| Strongylophorine | Sponges | Rho-GTPases | Oogenesis | [179] |
| Stypoldione | Algae | Microtubules (sulfhydryl groups) | Early development | [239] |
| Swinholide | Sponges | Microfilaments | Spermatogenesis | [125] |
| Theonellapeptolide | Sponges | Microfilaments | Oogenesis | [51] |
Abbreviations
| (AZA) | Azaspiracid |
| (CLA) | Calyculin-A |
| (CYN) | Caulerpenyne |
| (DD) | 2E, 4E-Decadienal |
| (DOL) | Dolastatin |
| (GEO) | Geodiamolide |
| (JAS) | Jasplakinolide |
| (LAT) | Latrunculin |
| (MET) | Methoxyconidiol |
| (MYC) | Mycalolide-B |
| (OA) | Okadaic acid |
| (PAL) | Palytoxin |
| (PTX) | Pectenotoxin |
| (PSE) | Pseudopterolide |
| (STR) | Strongylophorine |
| (STY) | Stypoldione |
| (SWI) | Swinholide-A |
| (TEO) | Theonellapeptolide Ie |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Silvestre, F.; Tosti, E. Impact of Marine Drugs on Cytoskeleton-Mediated Reproductive Events. Mar. Drugs 2010, 8, 881-915. https://doi.org/10.3390/md8040881
Silvestre F, Tosti E. Impact of Marine Drugs on Cytoskeleton-Mediated Reproductive Events. Marine Drugs. 2010; 8(4):881-915. https://doi.org/10.3390/md8040881
Chicago/Turabian StyleSilvestre, Francesco, and Elisabetta Tosti. 2010. "Impact of Marine Drugs on Cytoskeleton-Mediated Reproductive Events" Marine Drugs 8, no. 4: 881-915. https://doi.org/10.3390/md8040881
APA StyleSilvestre, F., & Tosti, E. (2010). Impact of Marine Drugs on Cytoskeleton-Mediated Reproductive Events. Marine Drugs, 8(4), 881-915. https://doi.org/10.3390/md8040881