
Vertical Distribution of Bacterial Community Diversity and Water Quality during the Reservoir Thermal Stratification

Abstract
:1. Introduction
2. Experimental Section
2.1. Study Area and Sample Collection
2.2. Water Quality Analysis
2.3. Water Microbial DNA Extraction
2.4. Illumina Miseq Sequencing and Sequence Analysis
2.5. Nucleotide Sequence Accession Number
2.6. Data Analysis
3. Results and Discussion
3.1. Vertical Water Quality during the Thermal Stratification

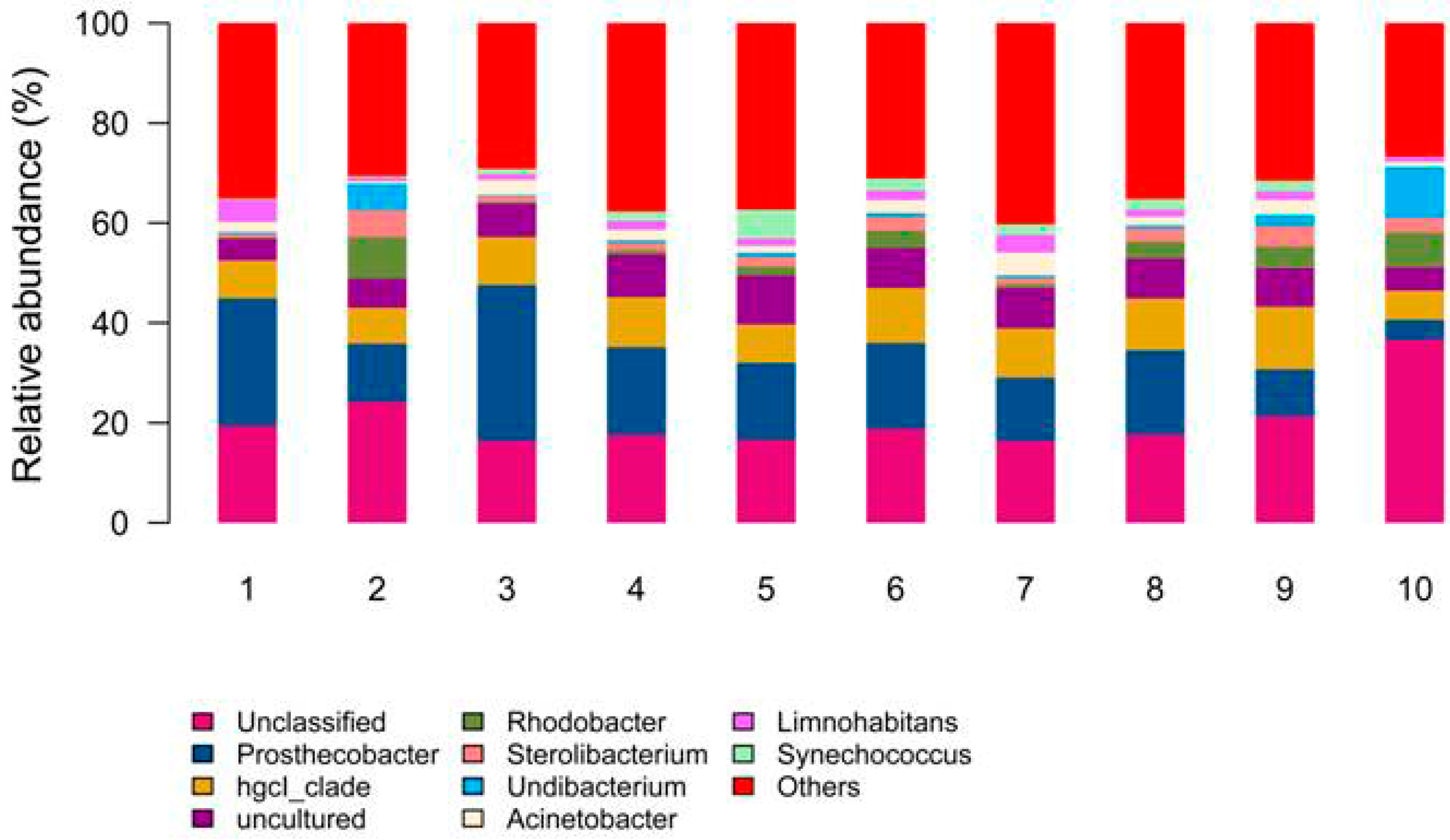
3.2. Vertical Water Bacterial Community Diversity during the Thermal Stratification

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Thermal Stratification | Water Depths | Reads Number | 0.97 Level | ||||
|---|---|---|---|---|---|---|---|
| OTUs | Diversity Estimator | Richness Estimator | |||||
| ACE | Chao1 | Shannon | Simpson | ||||
| Epilimnion | 0.5 m | 15,656 | 346 | 402 | 407 | 3.76 | 0.074 |
| 5 m | 26,128 | 476 | 507 | 506 | 4.25 | 0.038 | |
| 10 m | 24,006 | 383 | 431 | 430 | 3.47 | 0.107 | |
| Metalimnion | 20 m | 17,933 | 419 | 475 | 473 | 4.10 | 0.067 |
| 25 m | 13,199 | 401 | 451 | 454 | 4.24 | 0.046 | |
| 30 m | 37,943 | 456 | 482 | 482 | 4.04 | 0.060 | |
| Hypolimnion | 45 m | 19,711 | 438 | 466 | 462 | 4.41 | 0.038 |
| 50 m | 35,062 | 451 | 474 | 474 | 4.16 | 0.052 | |
| 55 m | 8282 | 371 | 434 | 443 | 4.33 | 0.040 | |
| 65 m | 12,197 | 386 | 445 | 456 | 4.03 | 0.044 | |




4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Röske, K.; Sachse, R.; Scheerer, C. Microbial diversity and composition of the sediment in the drinking water reservoir Saidenbach (Saxonia, Germany). Systemat. Appl. Microbiol. 2012, 35, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.H.; Huang, T.L.; Chen, S.N.; Guo, L.; Yang, X.; Liu, T.T. Spatial pattern of bacterial community functional diversity in a drinking water reservoir, Shaanxi Province, Northwest China. J. Pure Appl. Microb. 2013, 3, 1647–1654. [Google Scholar]
- Röske, K.; Roske, I.; Uhlmann, D. Characterization of the bacterial population and chemistry in the bottom sediment of a laterally subdivided drinking water reservoir system. Limnologica 2008, 38, 367–377. [Google Scholar] [CrossRef]
- Zhang, H.H.; Huang, T.L.; Chen, S.N.; Guo, L.; Yang, X. Microbial community functional diversity and enzymatic activity in the sediments of drinking water reservoirs, Northwest China. Desalin. Water Treat. 2014, 52, 1608–1614. [Google Scholar] [CrossRef]
- Zhang, H.H.; Huang, T.L.; Liu, T.T. Sediment enzyme activities and microbial community diversity in an oligotrophic drinking water reservoir, eastern China. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.Y.; Zeng, J.; Wan, W.H.; Liang, H.D.; Huang, R. Vertical distribution of ammonia-oxidizing archaea and bacteria in sediments of a Eutrophic Lake. Curr. Microbiol. 2013, 67, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Kagami, M.; Amano, Y.; Ishii, N. Community structure of planktonic fungi and the impact of parasitic Chytrids on phytoplankton in Lake Inba, Japan. Microb. Ecol. 2012, 63, 358–368. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Guo, Q.L.; Huang, T.L.; Tan, P. Response characteristics of water quality to the seasonal thermal stratification in Jin-pen reservoir along the Heihe river, Xi’an city in China. J. Hydraul. Eng. 2013, 44, 406–415. [Google Scholar]
- Qin, C.H.; Huang, T.L.; Li, X. Studies on the seasonal variation and budget of nitrogen, phosphorus of the shibianyu reservoir. J. Xi’an Univ. Architecture Technol. 2013, 45, 111–116. [Google Scholar]
- Elçi, Ş. Effects of thermal stratification and mixing on reservoir water quality. Limnology 2008, 2, 135–142. [Google Scholar] [CrossRef]
- Huang, T.L.; Li, X.; Rijnaarts, H.; Grotenhuis, T.; Ma, W.X.; Sun, X.; Xu, J.L. Effects of storm runoff on the thermal regime and water quality of a deep, stratified reservoir in a temperate monsoon zone, in Northwest China. Sci. Total Environ. 2014, 485–486, 820–827. [Google Scholar] [CrossRef] [PubMed]
- Garcia, S.L.; Salka, I.; Grossart, H.P.; Warnecke, F. Depth-discrete profiles of bacterial communities reveal pronounced spatio-temporal dynamics related to lake stratification. Environ. Microbiol. Rep. 2013, 5, 549–555. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, P.A.; Bálint, M.; Greshake, B.; Bandow, C.; Römbke, J.; Schmitt, I. Illumina metabarcoding of a soil fungal community. Soil. Biol. Biochem. 2013, 65, 128–132. [Google Scholar] [CrossRef]
- Zhang, H.H.; Huang, T.L. Archaeal community structure and quantity in the oxygen deficient sediments from three water supply reservoirs. J. Pure Appl. Microb. 2013, 4, 2783–2789. [Google Scholar]
- Wang, Y.; Sheng, H.F.; He, Y.; Wu, J.Y.; Jiang, Y.X.; Tam, N.F.Y.; Zhou, H.W. Comparison of the levels of bacterial diversity in freshwater, intertidal wetland, and marine sediments by using millions of illumina tags. Appl. Environ. Microb. 2012, 23, 8264–8271. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.T.; Peng, X.; Deng, G.H.; Sheng, H.F.; Wang, Y.; Zhou, H.W.; Tam, N.F.Y. Illumina sequencing of 16S rRNA tag revealed spatial variations of bacterial communities in a mangrove wetland. Microb. Ecol. 2013, 66, 96–104. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Yang, J.; Amalfitano, S.; Yu, X.Q.; Liu, L.M. Effects of water stratification and mixing on microbial community structure in a subtropical deep reservoir. Sci. Rep. 2014, 4. [Google Scholar] [CrossRef] [PubMed]
- Xie, W.; Yaoguang, Z.; Wei, Z.; Fang, X. The application of flow injection analysis in water quality detection. J. Environ. Hyg. 2012, 6, 20–28. [Google Scholar]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Quince, C.; Lanzen, A.; Davenport, R.; Turnbaugh, P. Removing noise from pyrosequenced amplicons. BMC Bioinform. 2011, 12. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.R.; Chai, B.; Farris, R.J.; Wang, Q.; Kulam-Syed-Mohideen, A.S. The ribosomal database project (RDP-II): Introducing myRDP space and quality controlled public data. Nucleic Acids Res. 2007, 35, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Loman, N.J.; Misra, R.V.; Dallman, T.J.; Constantinidou, C.; Gharbia, S.E. Performance comparison of benchtop high-throughput sequencing platforms. Nat. Biotechnol. 2012, 30, 434–439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R. Vegan: Community ecology package. R Package Vers. 2012, 5, 20–23. [Google Scholar]
- Brookes, J.D.; O’Brien, K.R.; Burford, M.A.; Bruesewitz, D.A.; Hodges, B.R.; McBride, C.; Hamilton, D.P. Effects of diurnal vertical mixing and stratification on phytoplankton productivity in geothermal Lake Rotowhero, New Zealand. Inland Waters. 2013, 3, 369–376. [Google Scholar] [CrossRef]
- De Hoyos, C.; Negro, A.I.; Aldasoro, J.J. Cyanobacteria distribution and abundance in the Spanish water reservoirs during thermal stratification. Limnetica 2004, 23, 119–132. [Google Scholar]
- Messyasz, B.; Czerwik-Marcinkowska, J.; Lücke, A.; Uher, B. Differences in the ultrastructure of two selected taxa of phytoplankton in a thermally stratified Lake Holzmaar (Germany). Biodiv. Res. Conserv. 2012, 28, 55–62. [Google Scholar] [CrossRef]
- Paganin, P.; Chiarini, L.; Bevivino, A.; Dalmastri, C.; Farcomeni, A.; Izzo, G.; Tabacchioni, S. Vertical distribution of bacterioplankton in Lake Averno in relation to water chemistry. FEMS Microb. Ecol. 2013, 84, 176–188. [Google Scholar] [CrossRef] [PubMed]
- Rösel, S.; Allgaier, M.; Grossart, H.P. Long-term characterization of free-living and particle-associated bacterial communities in Lake Tiefwaren reveals distinct seasonal patterns. Microb. Ecol. 2012, 3, 571–583. [Google Scholar] [CrossRef] [PubMed]
- Comeau, A.M.; Harding, T.; Galand, P.E.; Vincen, W.F.; Lovejoy, C. Vertical distribution of microbial communities in a perennially stratified Arctic lake with saline, anoxic bottom waters. Sci. Rep. 2012, 2. [Google Scholar] [CrossRef] [PubMed]
- Shade, A.; Read, J.S.; Youngblut, N.D.; Fierer, N.; Knight, R.; Kratz, T.K.; McMahon, K.D. Lake microbial communities are resilient after a whole-ecosystem disturbance. ISME. J. 2012, 12, 2153–2167. [Google Scholar] [CrossRef] [PubMed]
- Comerma, M.; Garcia, J.C.; Romero, M.; Armengol, J.; Šimek, K. Carbon flow dynamics in the pelagic community of the Sau Reservoir (Catalonia, NE Spain). Hydrobiologia 2003, 504, 87–98. [Google Scholar] [CrossRef]
- Pouliot, J.; Galand, P.E.; Lovejoy, C.; Vincent, W.F. Vertical structure of archaeal communities and the distribution of ammonia monooxygenase A gene variants in two meromictic High Arctic lakes. Environ. Microbiol. 2009, 11, 687–699. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.M.; Cupples, A.M. Diversity of five anaerobic toluene-degrading microbial communities investigated using stable isotope probing. Appl. Environ. Microbiol. 2012, 78, 972–980. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.-H.; Chen, S.-N.; Huang, T.-L.; Ma, W.-X.; Xu, J.-L.; Sun, X. Vertical Distribution of Bacterial Community Diversity and Water Quality during the Reservoir Thermal Stratification. Int. J. Environ. Res. Public Health 2015, 12, 6933-6945. https://doi.org/10.3390/ijerph120606933
Zhang H-H, Chen S-N, Huang T-L, Ma W-X, Xu J-L, Sun X. Vertical Distribution of Bacterial Community Diversity and Water Quality during the Reservoir Thermal Stratification. International Journal of Environmental Research and Public Health. 2015; 12(6):6933-6945. https://doi.org/10.3390/ijerph120606933
Chicago/Turabian StyleZhang, Hai-Han, Sheng-Nan Chen, Ting-Lin Huang, Wei-Xing Ma, Jin-Lan Xu, and Xin Sun. 2015. "Vertical Distribution of Bacterial Community Diversity and Water Quality during the Reservoir Thermal Stratification" International Journal of Environmental Research and Public Health 12, no. 6: 6933-6945. https://doi.org/10.3390/ijerph120606933
APA StyleZhang, H. -H., Chen, S. -N., Huang, T. -L., Ma, W. -X., Xu, J. -L., & Sun, X. (2015). Vertical Distribution of Bacterial Community Diversity and Water Quality during the Reservoir Thermal Stratification. International Journal of Environmental Research and Public Health, 12(6), 6933-6945. https://doi.org/10.3390/ijerph120606933