
Use of Walnut Shell Powder to Inhibit Expression of Fe2+-Oxidizing Genes of Acidithiobacillus Ferrooxidans

Abstract
:1. Introduction
2. Materials and Methods
2.1. Determination of Walnut Shell Composition
2.2. Strains, Medium, and Culture Conditions
2.3. Analytical Methods for the Culture
2.4. Total RNA Extraction and Real-Time Polymerase Chain Reaction
2.5. Growth Curves of A. ferrooxidans in 9K Medium Containing Different Components of Walnut Shell
2.6. Pyrite Bioleaching Solution Preparation and Bioleaching Experiment
3. Results and Discussion
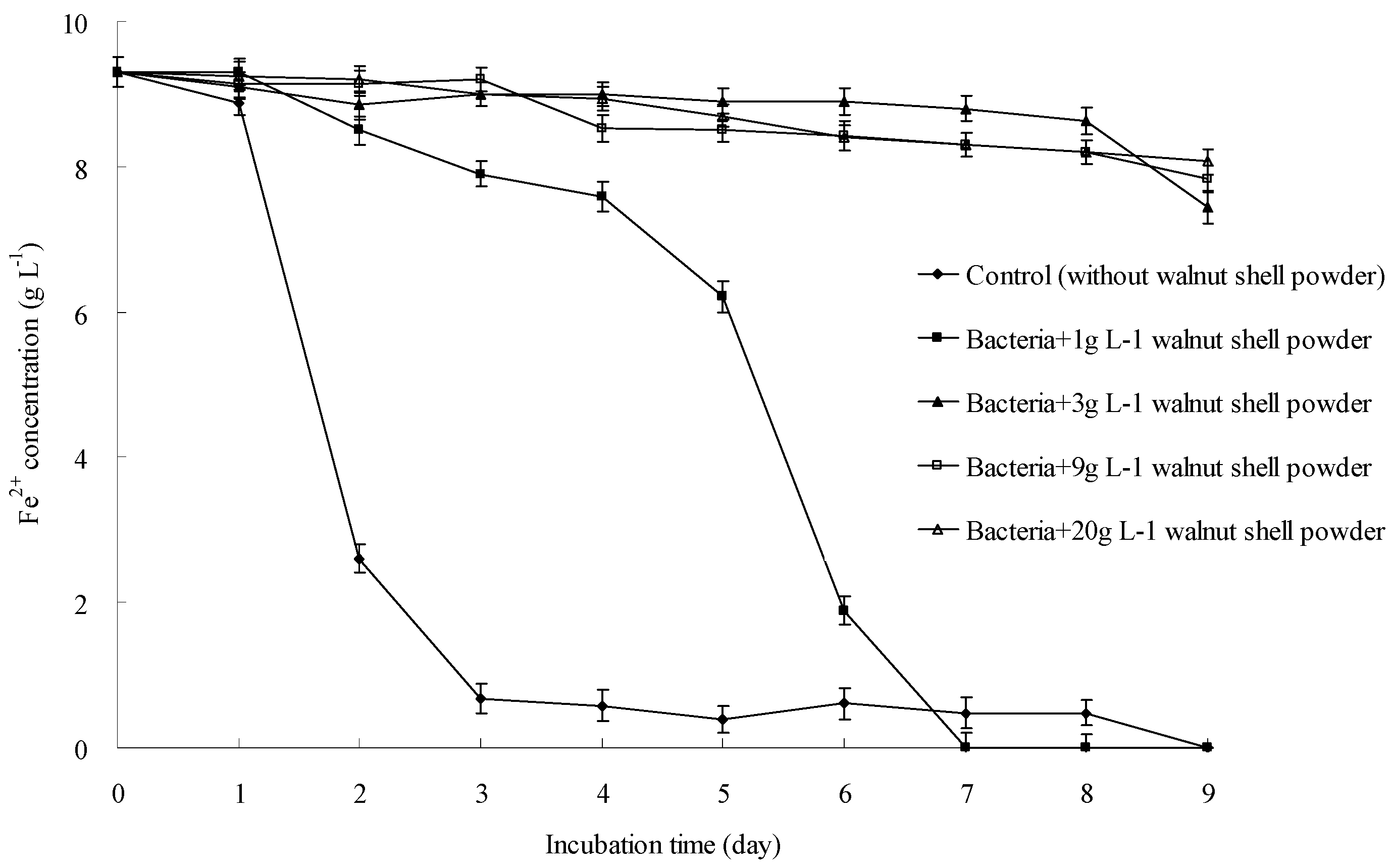
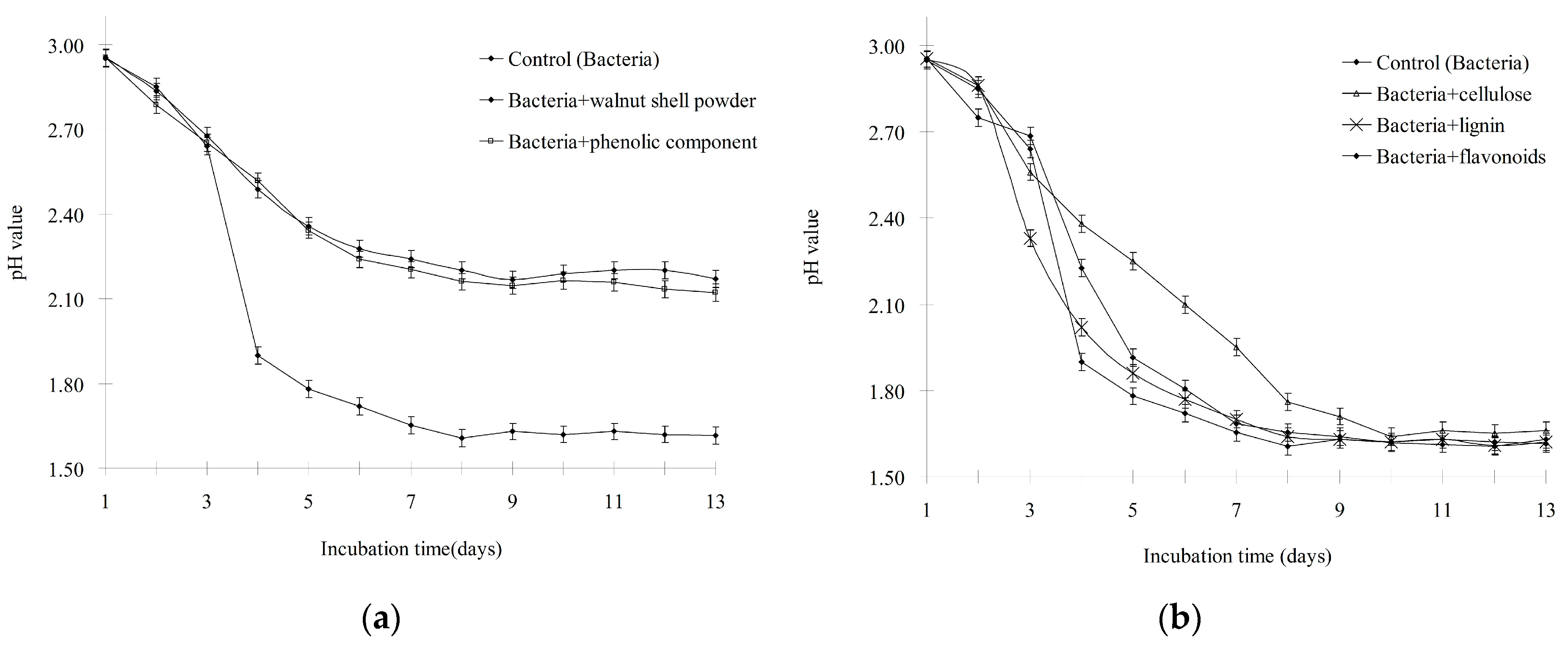
3.1. Walnut Shell Powder Inhibited the Oxidative Activity of A. ferrooxidans
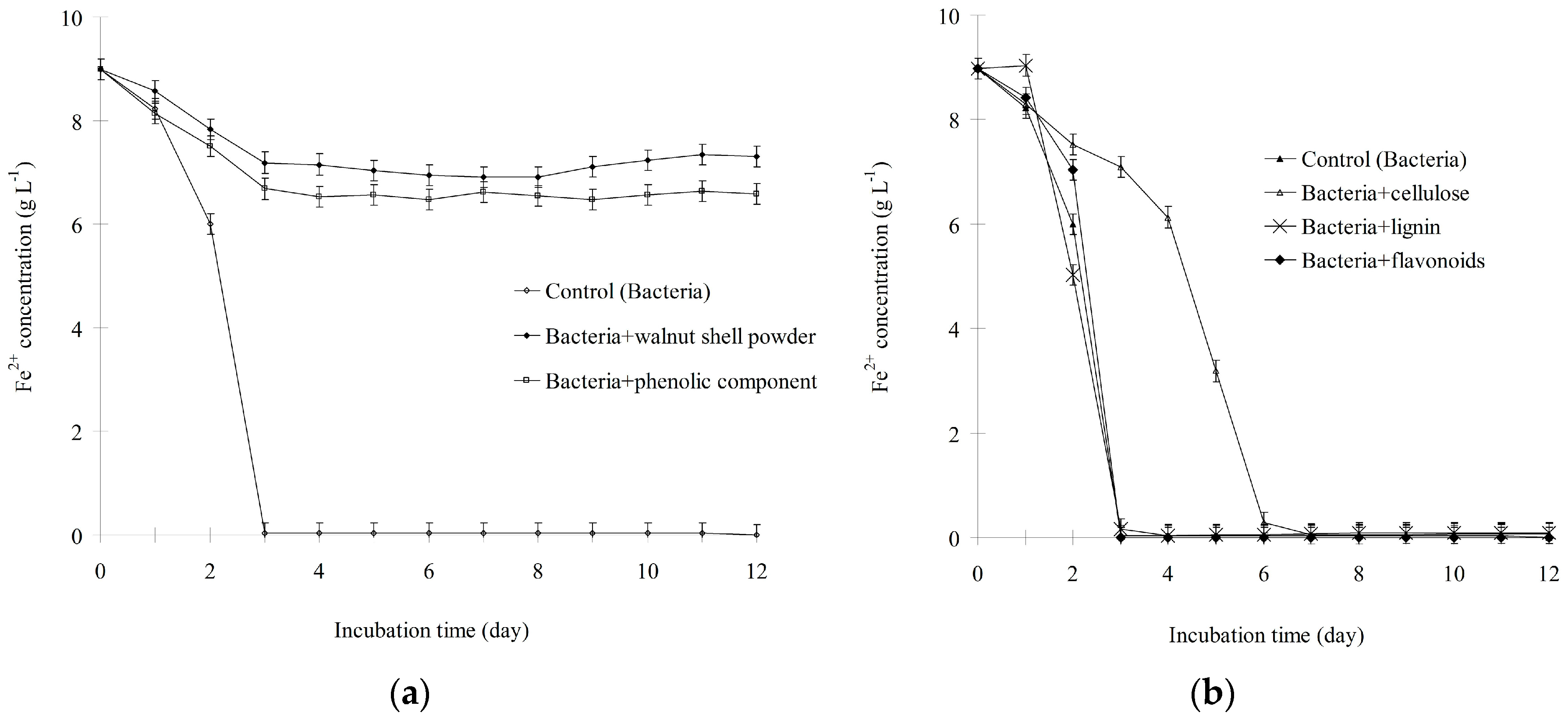
3.2. Only the Phenolic Component Could Inhibit the Oxidative Activity of A. ferrooxidans
3.3. Relative Expression of Fe2+-Oxidizing Genes Was Inhibited by the Phenolic Components
3.4. Expression of Rus Operon Genes Was Suppressed by the Phenolic Fraction
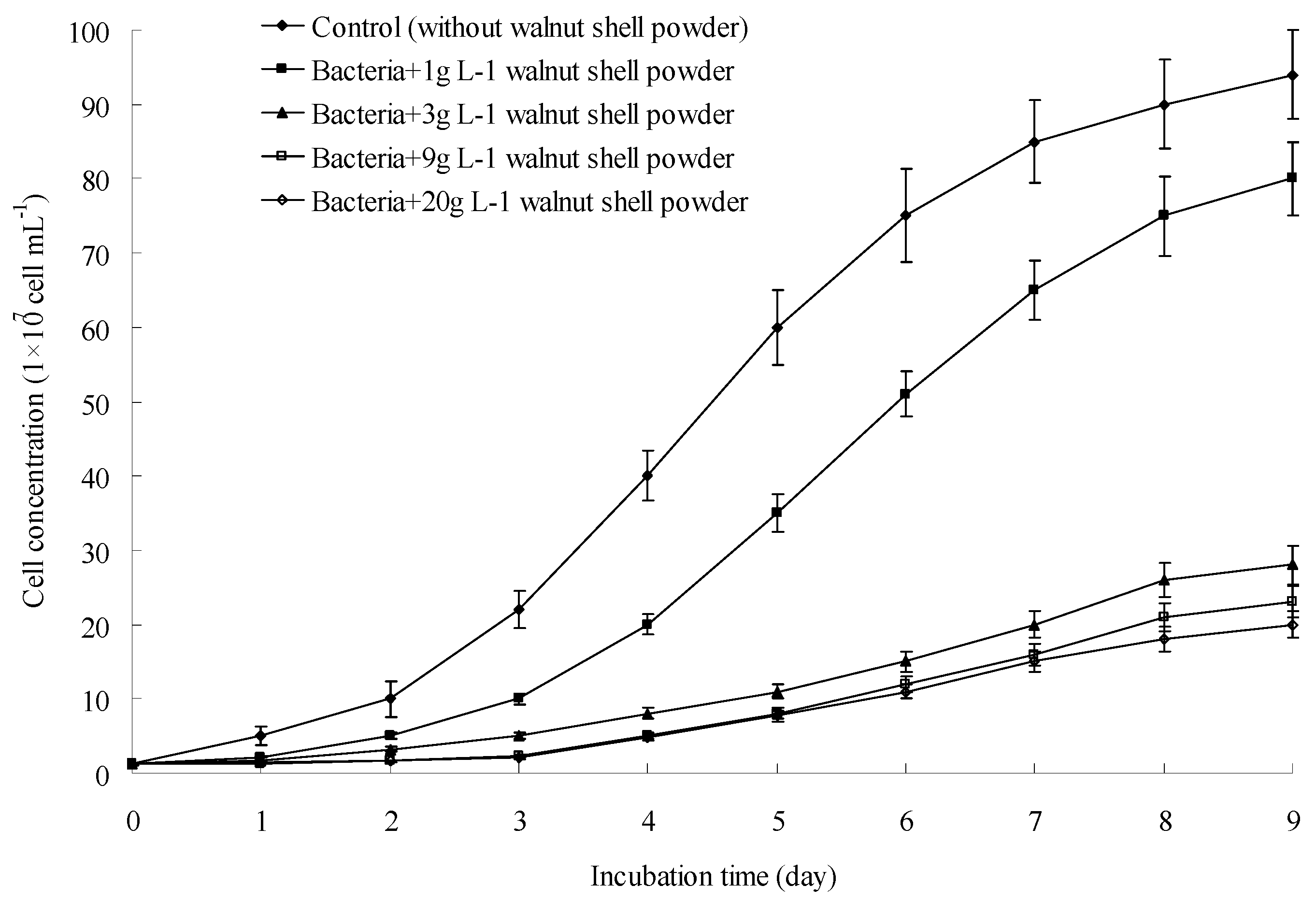
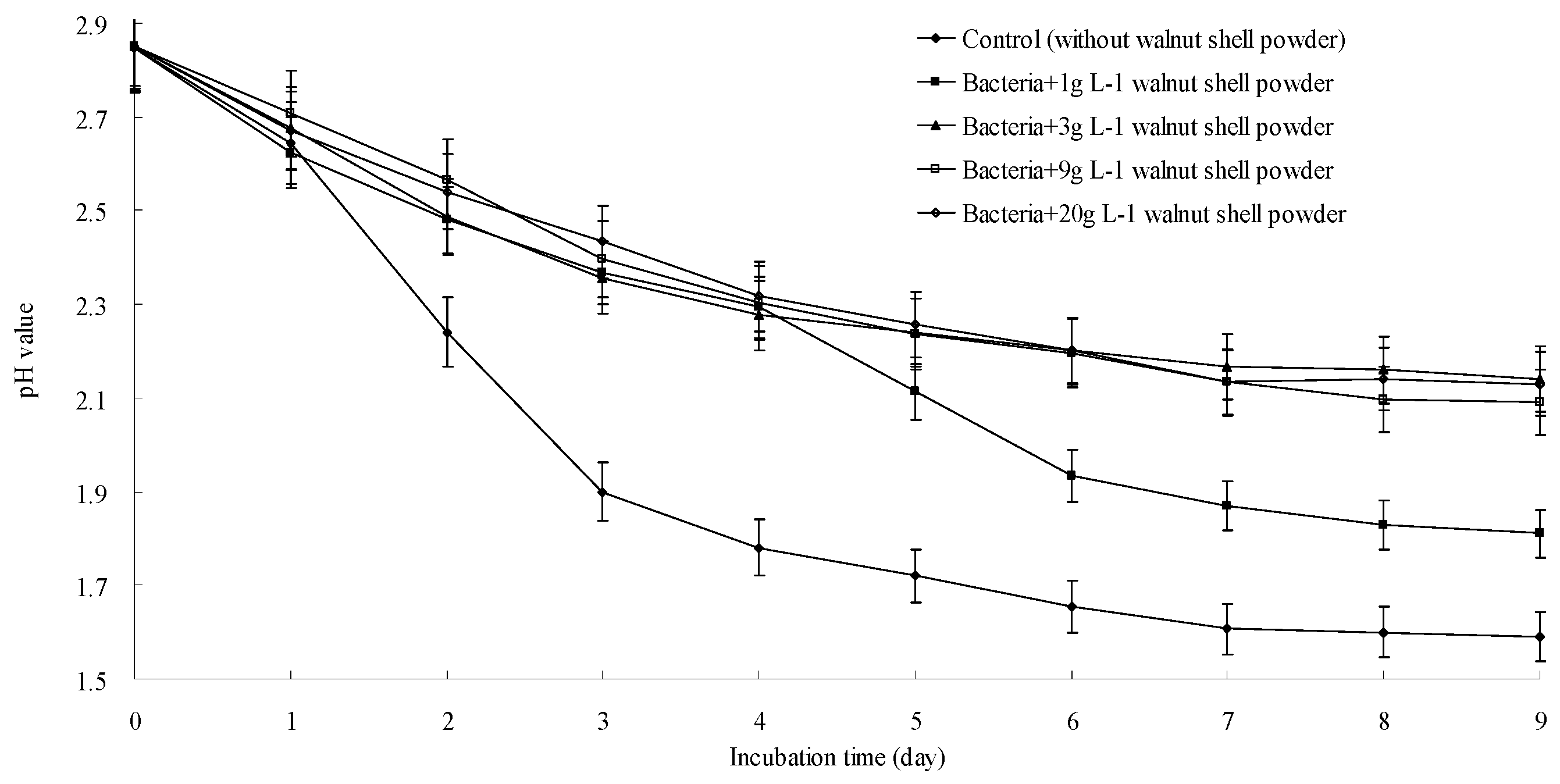
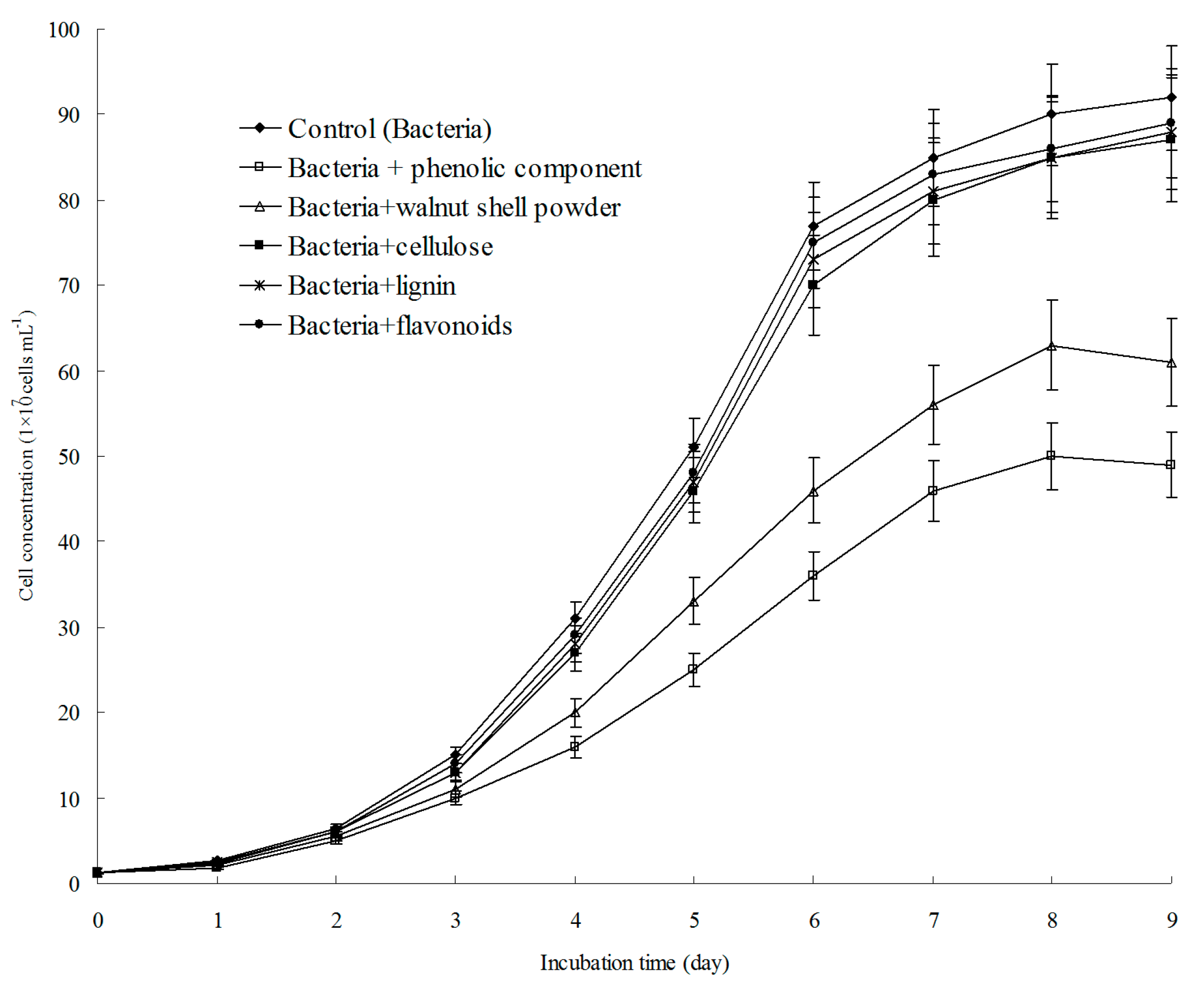
3.5. A. ferrooxidans Growth Could Be Inhibited by Walnut Shell Powder and the Phenolic Components
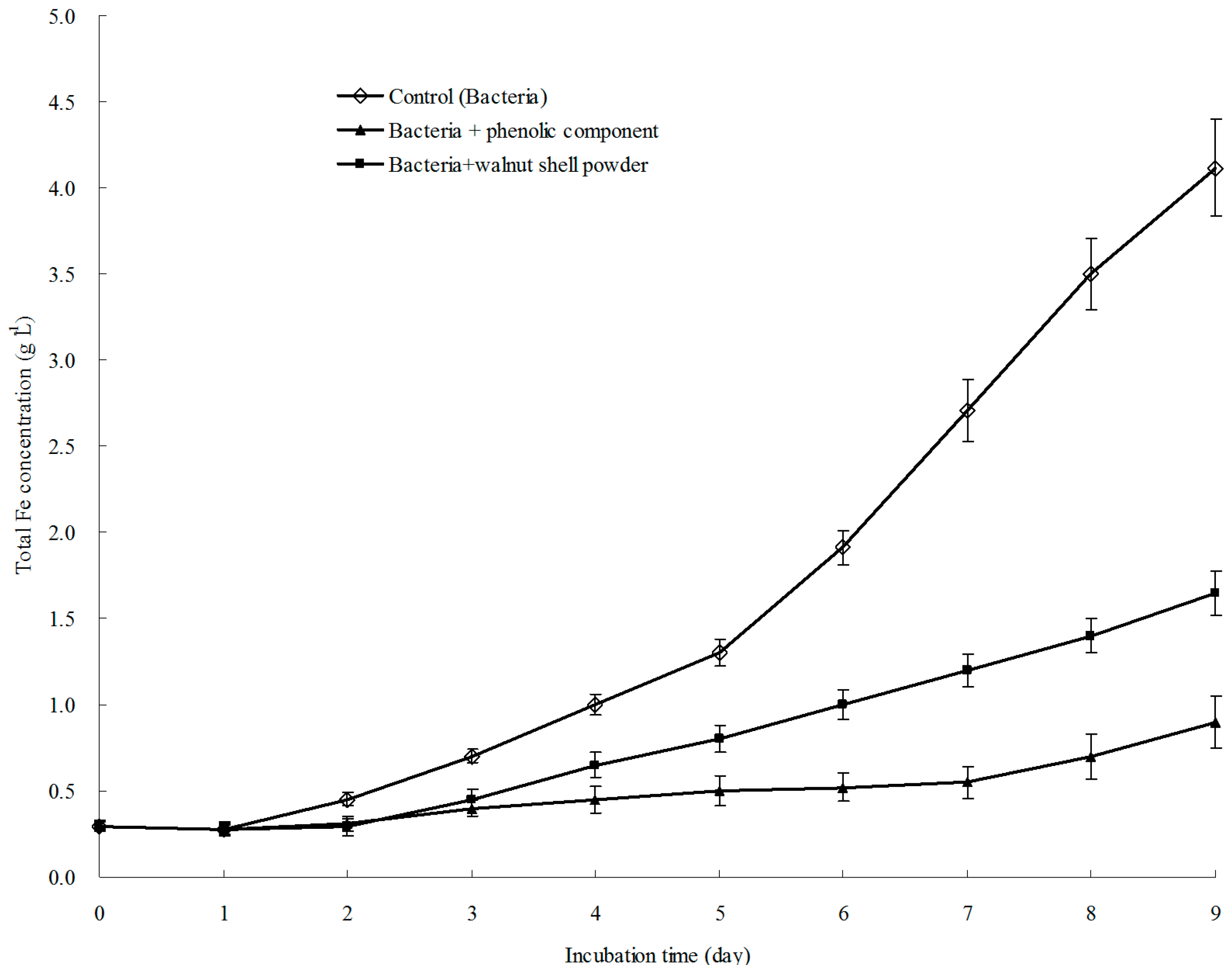
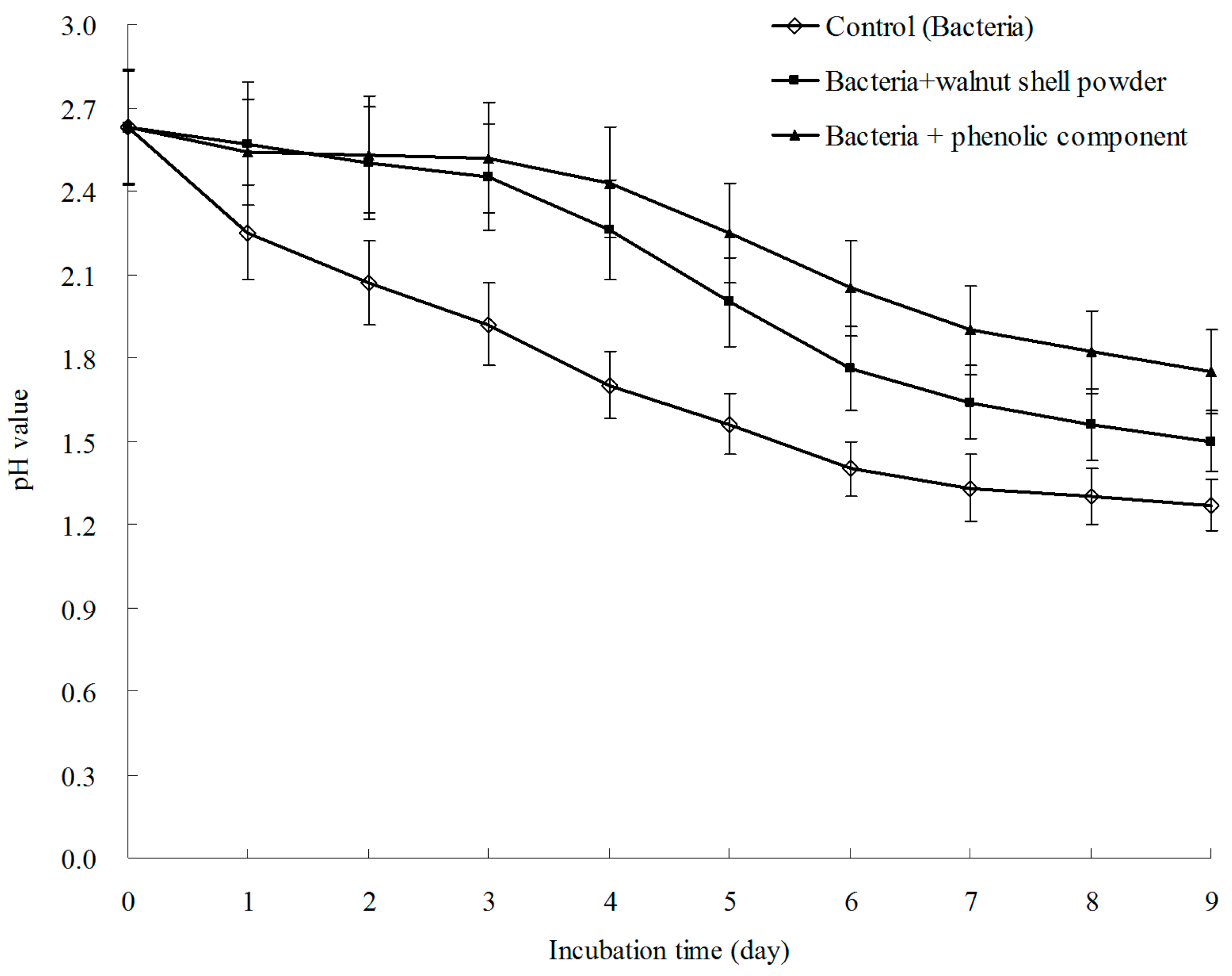
3.6. Result of Bioleaching Experiment Showed that the Phenolic Fraction Was the Active Ingredient
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Vicente-Beckett, V.A.; Taylor McCauley, G.J.; Duivenvoorden, L.J. Metal speciation in sediments and soils associated with acid-mine drainage in Mount Morgan (Queensland, Australia). J. Environ. Sci. Health A Tox. Hazard. Subst. Environ. Eng. 2015, 51, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.B.; Hallberg, K.B. Acid mine drainage remediation options: A review. Sci. Total Environ. 2005, 338, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Kucera, J.; Bouchal, P.; Lochman, J.; Potesil, D.; Janiczek, O.; Zdrahal, Z.; Mandl, M. Ferrous iron oxidation by sulfur-oxidizing Acidithiobacillus ferrooxidans and analysis of the process at the levels of transcription and protein synthesis. Antonie Van Leeuwenhoek 2013, 103, 905–919. [Google Scholar] [CrossRef] [PubMed]
- Xie, X.; Xiao, S.; Liu, J. Microbial communities in acid mine drainage and their interaction with pyrite surface. Curr. Microbiol. 2009, 59, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Alakangas, L.; Andersson, E.; Mueller, S. Neutralization/prevention of acid rock drainage using mixtures of alkaline by-products and sulfidic mine wastes. Environ. Sci. Pollut. Res. Int. 2013, 20, 7907–7916. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, P.K.; Kim, K.; Equeenuddin, S.M.; Powell, M.A. Current approaches for mitigating acid mine drainage. Rev. Environ. Contam. Toxicol. 2013, 226, 1–32. [Google Scholar] [PubMed]
- DellaGreca, M.; Zarrelli, A.; Fergola, P.; Cerasuolo, M.; Pollio, A.; Pinto, G. Fatty acids released by chlorella vulgaris and their role in interference with pseudokirchneriella subcapitata: Experiments and modelling. J. Chem. Ecol. 2010, 36, 339–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brune, K.D.; Bayer, T.S. Engineering microbial consortia to enhance biomining and bioremediation. Front. Microbiol. 2012, 3, 203. [Google Scholar] [CrossRef] [PubMed]
- Roberts, D.W.; Roberts, J.F.; Hodges, G.; Gutsell, S.; Ward, R.S.; Llewellyn, C. Aquatic toxicity of cationic surfactants to daphnia magna. SAR QSAR Environ. Res. 2013, 24, 417–427. [Google Scholar] [CrossRef] [PubMed]
- Din, M.I.; Mirza, M.L. Biosorption potentials of a novel green biosorbent saccharum bengalense containing cellulose as carbohydrate polymer for removal of Ni (II) ions from aqueous solutions. Int. J. Biol. Macromol. 2013, 54, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Mahboubi, A.; Kamalinejad, M.; Ayatollahi, A.M.; Babaeian, M. Total phenolic content and antibacterial activity of five plants of labiatae against four foodborne and some other bacteria. Iran. J. Pharm. Res. 2014, 13, 559–566. [Google Scholar] [PubMed]
- Morais-Braga, M.F.; Souza, T.M.; Santos, K.K.; Guedes, G.M.; Andrade, J.C.; Tintino, S.R.; Sobral-Souza, C.E.; Costa, J.G.; Saraiva, A.A.; Coutinho, H.D. Phenolic compounds and interaction between aminoglycosides and natural products of lygodium venustum SW against multiresistant bacteria. Chemotherapy 2012, 58, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Ding, D.; Lei, Z.; Yang, Y.; Feng, C.; Zhang, Z. Selective removal of cesium from aqueous solutions with nickel (II) hexacyanoferrate (III) functionalized agricultural residue-walnut shell. J. Hazard. Mater. 2014, 270, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Ding, D.; Zhao, Y.; Yang, S.; Shi, W.; Zhang, Z.; Lei, Z.; Yang, Y. Adsorption of cesium from aqueous solution using agricultural residue-walnut shell: Equilibrium, kinetic and thermodynamic modeling studies. Water Res. 2013, 47, 2563–2571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giovanelli, G.; Buratti, S. Comparison of polyphenolic composition and antioxidant activity of wild Italian blueberries and some cultivated varieties. Food Chem. 2009, 112, 903–908. [Google Scholar] [CrossRef]
- Torun, M.; Dincer, C.; Topuz, A.; Sahin-Nadeem, H.; Ozdemir, F. Aqueous extraction kinetics of soluble solids, phenolics and flavonoids from sage (salvia fruticosa miller) leaves. J. Food Sci. Technol. 2015, 52, 2797–2805. [Google Scholar] [CrossRef] [PubMed]
- Ni, Q.; Xu, G.; Wang, Z.; Gao, Q.; Wang, S.; Zhang, Y. Seasonal variations of the antioxidant composition in ground bamboo sasa argenteastriatus leaves. Int. J. Mol. Sci. 2012, 13, 2249–2262. [Google Scholar] [CrossRef] [PubMed]
- Ross, K.; Mazza, G. Characteristics of lignin from flax shives as affected by extraction conditions. Int. J. Mol. Sci. 2010, 11, 4035–4050. [Google Scholar] [CrossRef] [PubMed]
- Al-Hoqbani, A.A.; Abdel-Halim, E.S.; Al-Deyab, S.S. Extraction of palm tree cellulose and its functionalization via graft copolymerization. Int. J. Biol. Macromol. 2014, 70, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yue, M. Isolation and identification of acidophiles in acid mine drainage of tongling xinqiao coal mine. In Proceedings of the 3rd International Conference on Environmental Technology and Knowledge Transfer, Hefei, China, 22 January 2010; Nelles, M., Cai, J., Wu, K., Eds.; pp. 807–811.
- Silverman, M.P.; Lundgren, D.G. Studies on the chemoautotrophic iron bacterium ferrobacillus ferrooxidans: I: An improved medium and a harvesting procedure for securing high cell yields. J. Bacteriol. 1959, 77, 642–647. [Google Scholar] [PubMed]
- Yang, X.; Wang, S.; Liu, Y.; Zhang, Y. Identification and characterization of Acidithiobacillus ferrooxidans YY2 and its application in the biodesulfurization of coal. Can. J. Microbiol. 2015, 61, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Yarzabal, A.; Appia-Ayme, C.; Ratouchniak, J.; Bonnefoy, V. Regulation of the expression of the Acidithiobacillus ferrooxidans rus operon encoding two cytochromes C, a cytochrome oxidase and rusticyanin. Microbiology (Reading Engl.) 2004, 150, 2113–2123. [Google Scholar] [CrossRef] [PubMed]
- Carlos, C.; Reis, F.C.; Vicentini, R.; Madureira, D.J.; Ottoboni, L.M. The rus operon genes are differentially regulated when Acidithiobacillus ferrooxidans LR is kept in contact with metal sulfides. Curr. Microbiol. 2008, 57, 375–380. [Google Scholar] [CrossRef] [PubMed]
- Kchaou, W.; Abbes, F.; Mansour, R.B.; Blecker, C.; Attia, H.; Besbes, S. Phenolic profile, antibacterial and cytotoxic properties of second grade date extract from Tunisian cultivars (Phoenix dactylifera L.). Food Chem. 2016, 194, 1048–1055. [Google Scholar] [CrossRef] [PubMed]
- Mahboubi, A.; Asgarpanah, J.; Sadaghiyani, P.N.; Faizi, M. Total phenolic and flavonoid content and antibacterial activity of Punica granatum L. var. Pleniflora flowers (Golnar) against bacterial strains causing foodborne diseases. BMC Complement. Altern. Med. 2015, 15, 366. [Google Scholar] [CrossRef] [PubMed]
- Liljeqvist, M.; Rzhepishevska, O.I.; Dopson, M. Gene identification and substrate regulation provide insights into sulfur accumulation during bioleaching with the psychrotolerant acidophile Acidithiobacillus ferrivorans. Appl. Environ. Microbiol. 2013, 79, 951–957. [Google Scholar] [CrossRef] [PubMed]
- Bruscella, P.; Appia-Ayme, C.; Levican, G.; Ratouchniak, J.; Jedlicki, E.; Holmes, D.S.; Bonnefoy, V. Differential expression of two BC1 complexes in the strict acidophilic chemolithoautotrophic bacterium Acidithiobacillus ferrooxidans suggests a model for their respective roles in iron or sulfur oxidation. Microbiology (Reading Engl.) 2007, 153, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Quatrini, R.; Appia-Ayme, C.; Denis, Y.; Jedlicki, E.; Holmes, D.S.; Bonnefoy, V. Extending the models for iron and sulfur oxidation in the extreme acidophile Acidithiobacillus ferrooxidans. BMC Genomics 2009, 10, 394. [Google Scholar] [CrossRef] [PubMed]
- Rohwerder, T.; Gehrke, T.; Kinzler, K.; Sand, W. Bioleaching review part A: Progress in bioleaching: Fundamentals and mechanisms of bacterial metal sulfide oxidation. Appl. Microbiol. Biotechnol. 2003, 63, 239–248. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Peng, Q.; Selimi, D.; Wang, Q.; Charkowski, A.O.; Chen, X.; Yang, C.H. The plant phenolic compound p-coumaric acid represses gene expression in the dickeya dadantii type III secretion system. Appl. Environ. Microbiol. 2009, 75, 1223–1228. [Google Scholar] [CrossRef] [PubMed]
- Myszka, K.; Schmidt, M.T.; Olejnik-Schmidt, A.K.; Leja, K.; Czaczyk, K. Influence of phenolic acids on indole acetic acid production and on the type III secretion system gene transcription in food-associated Pseudomonas fluorescens KM05. J. Biosci. Bioeng. 2014, 118, 651–656. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Chen, K.M.; Zhi, D.J.; Xie, Q.J.; Li, H.Y. Effects of pyrite bioleaching solution of Acidithiobacillus ferrooxidans on viability, differentiation and mineralization potentials of rat osteoblasts. Arch. Pharm. Res. 2015, 38, 2228–2240. [Google Scholar] [CrossRef] [PubMed]
- Hong, F.F.; He, H.; Liu, J.Y.; Tao, X.X.; Zheng, L.; Zhao, Y.D. Comparison analysis of coal biodesulfurization and coal’s pyrite bioleaching with Acidithiobacillus ferrooxidans. Sci. World J. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Medium | The Walnut Shell Component Added |
|---|---|---|
| Control (Bacteria) | 0 K + pyrite | None |
| Bacteria + walnut shell powder | 0 K + pyrite | Walnut shell powder |
| Bacteria + phenolic fraction | 0 K + pyrite | Phenolics |
| Component | Content (g·kg−1 Walnut Shell) | The Dosage Used in the Experiments (g) |
|---|---|---|
| Cellulose | 350.7 ± 10.5 | 1.05 |
| Lignin | 314.7 ± 8.5 | 0.94 |
| Phenolics | 16.9 ± 1.9 | 0.051 |
| Flavonoids | 3.7 ± 0.5 | 0.011 |
| Operon Name | Gene | Control | Phenolic Fraction | Ratio of Control to Total Phenolics |
|---|---|---|---|---|
| res operon | hyp | 20.3 ± 2.4 | 7.5 ± 0.2 | 2.7 |
| resC | 11.4 ± 1.3 | 4.1 ± 0.8 | 2.8 | |
| resB | 16.2 ± 2.1 | 1.1 ± 0.2 | 14.7 | |
| petI operon | petC-1 | 7.2 ± 1.1 | 3.2 ± 0.4 | 2.3 |
| petB-1 | 16.2 ± 2.1 | 4.2 ± 0.2 | 3.9 | |
| petA-1 | 26.2 ± 3.1 | 8.3 ± 0.6 | 3.2 | |
| sdrA-1 | 17.9 ± 1.6 | 1.5 ± 0.1 | 11.9 | |
| cycA-1 | 26.9 ± 3.2 | 12.1 ± 1.1 | 2.2 | |
| iron regulators | regA | 6.2 ± 1.9 | 2.8 ± 0.7 | 2.2 |
| regB | 30.8 ± 2.7 | 16.8 ± 1.9 | 1.8 | |
| ctaR | 17.2 ± 2.2 | 3.5 ± 0.4 | 4.9 | |
| fur | 5.4 ± 1.1 | 1.2 ± 0.3 | 4.5 |
| Gene | Control | Total Phenolics | Ratio of Control to Total Phenolics |
|---|---|---|---|
| cyc2 | 27.3 ± 2.2 | 4.7 ± 0.3 | 5.8 |
| orf | 80.1 ± 7.6 | 7.5 ± 0.2 | 10.7 |
| cyc1 | 10.4 ± 0.9 | 0.6 ± 0.04 | 17.3 |
| coxB | 142.9 ± 8.3 | 8.7 ± 0.9 | 16.4 |
| coxA | 75.6 ± 6.6 | 8.1 ± 0.5 | 9.3 |
| coxC | 57.3 ± 4.3 | 9.4 ± 0.6 | 6.1 |
| coxD | 155.4 ± 7.3 | 13.6 ± 1.3 | 11.4 |
| rus | 227.5 ± 13.7 | 10.3 ± 0.5 | 22.1 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Liu, Y.; Tan, H.; Zhang, Y.; Yue, M. Use of Walnut Shell Powder to Inhibit Expression of Fe2+-Oxidizing Genes of Acidithiobacillus Ferrooxidans. Int. J. Environ. Res. Public Health 2016, 13, 461. https://doi.org/10.3390/ijerph13050461
Li Y, Liu Y, Tan H, Zhang Y, Yue M. Use of Walnut Shell Powder to Inhibit Expression of Fe2+-Oxidizing Genes of Acidithiobacillus Ferrooxidans. International Journal of Environmental Research and Public Health. 2016; 13(5):461. https://doi.org/10.3390/ijerph13050461
Chicago/Turabian StyleLi, Yuhui, Yehao Liu, Huifang Tan, Yifeng Zhang, and Mei Yue. 2016. "Use of Walnut Shell Powder to Inhibit Expression of Fe2+-Oxidizing Genes of Acidithiobacillus Ferrooxidans" International Journal of Environmental Research and Public Health 13, no. 5: 461. https://doi.org/10.3390/ijerph13050461
APA StyleLi, Y., Liu, Y., Tan, H., Zhang, Y., & Yue, M. (2016). Use of Walnut Shell Powder to Inhibit Expression of Fe2+-Oxidizing Genes of Acidithiobacillus Ferrooxidans. International Journal of Environmental Research and Public Health, 13(5), 461. https://doi.org/10.3390/ijerph13050461