Effect of Climate and Land Use on the Spatio-Temporal Variability of Tick-Borne Bacteria in Europe

 ,
,  ,
,  , , , , ,
, , , , , 
 ,
,  add
Show full author list
add
Show full author list
Abstract
:1. Introduction
2. Materials and Methods
2.1. Questing Tick Data
2.2. Molecular Analyses
2.3. Climatic and Environmental Data
2.4. Statistical Analyses
3. Results
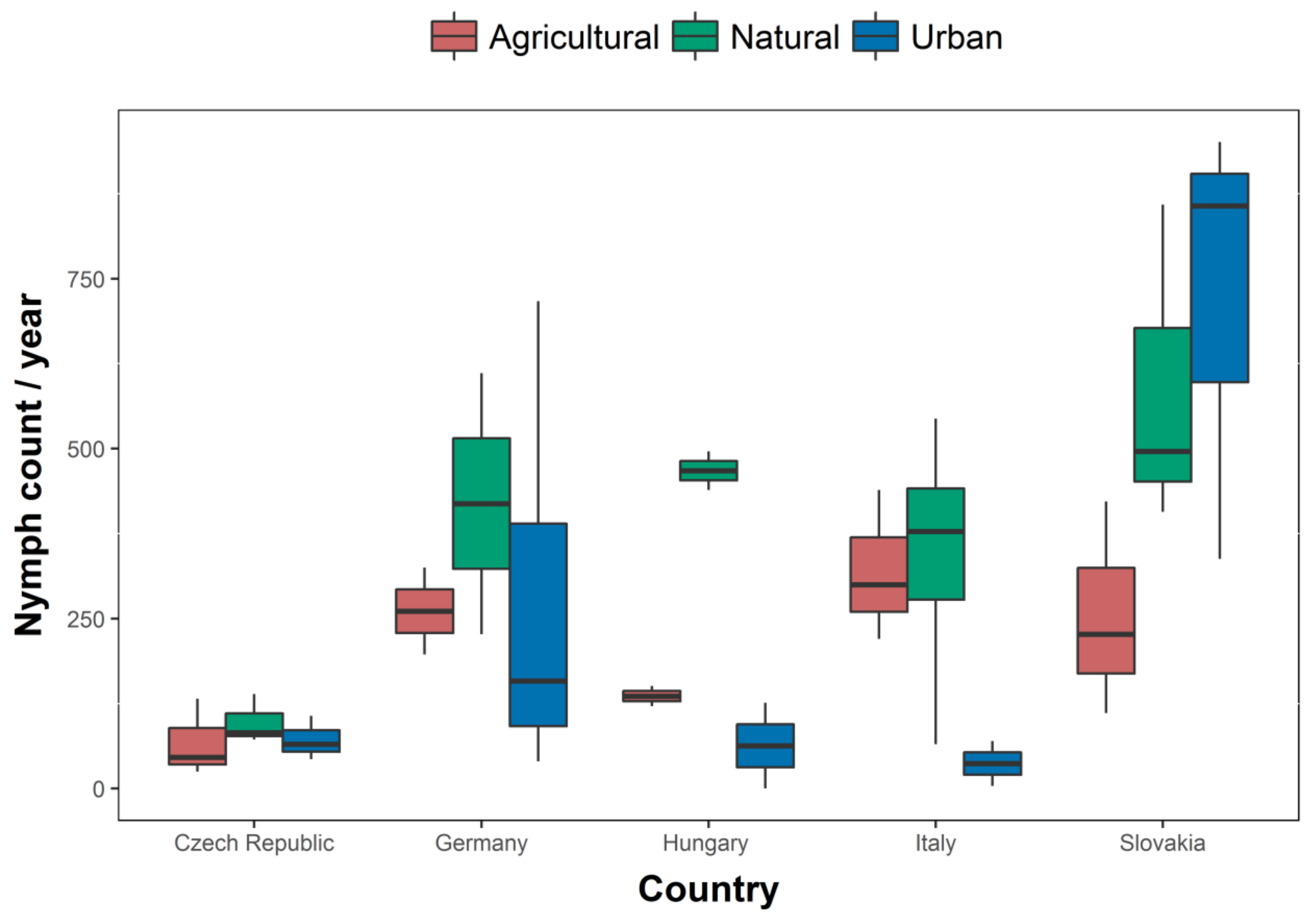
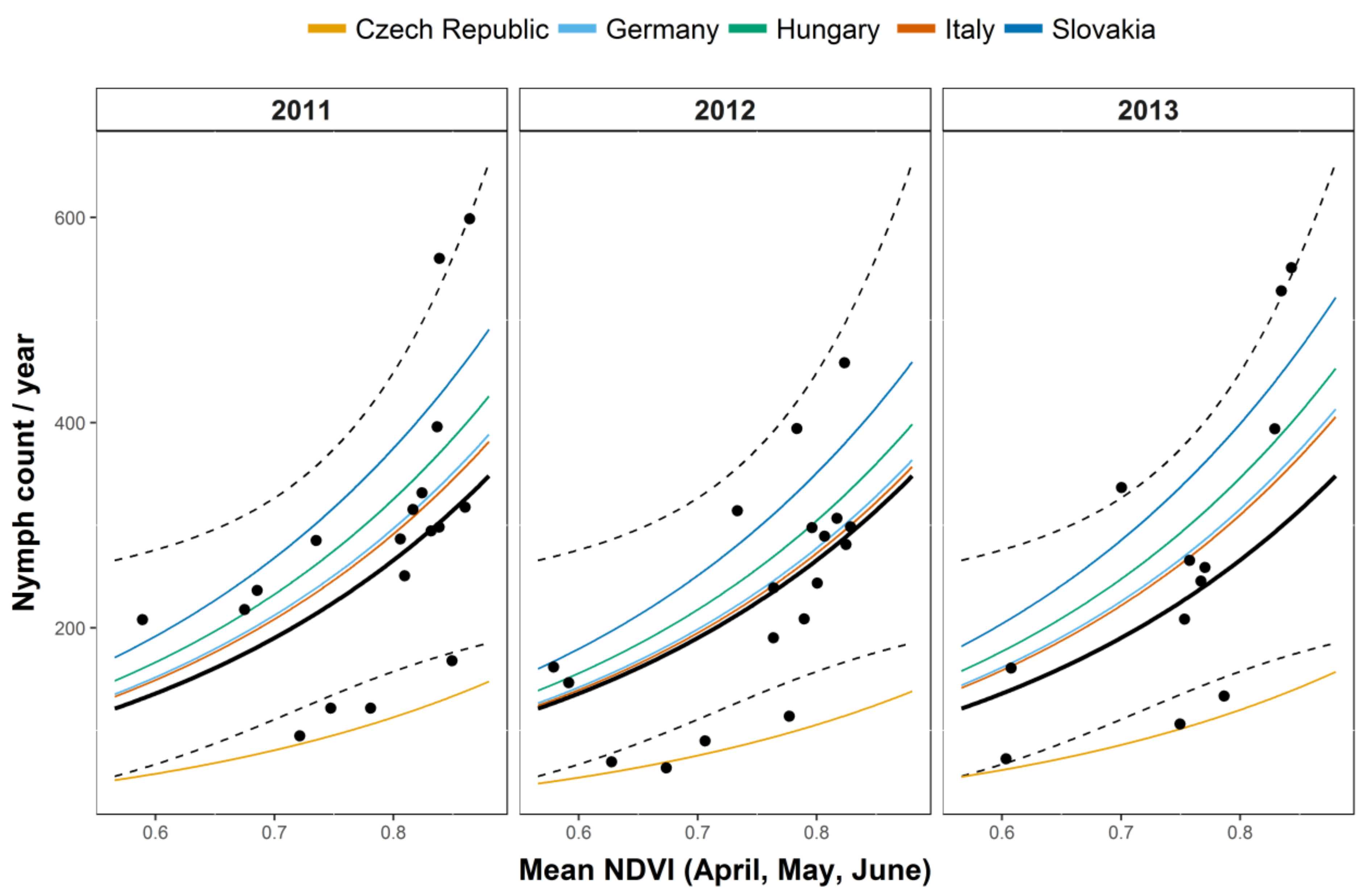
3.1. Questing Nymph Abundance
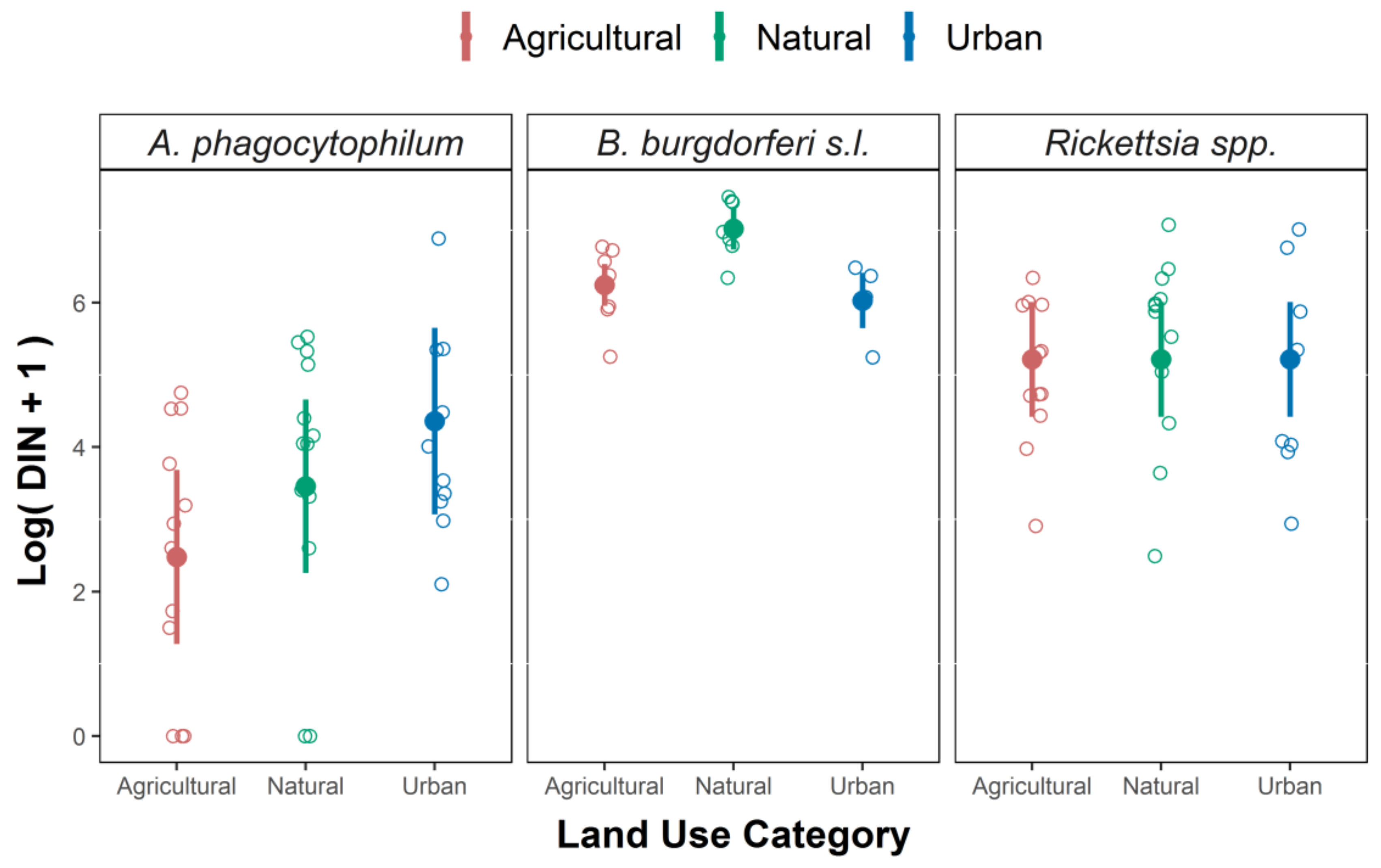
3.2. Infection Prevalence and DIN
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ellis, E.C.; Ramankutty, N. Putting People in the Map: Anthropogenic Biomes of the World. Front. Ecol. Environ. 2008, 6, 439–447. [Google Scholar] [CrossRef]
- Bradley, C.A.; Altizer, S. Urbanization and the Ecology of Wildlife Diseases. Trends Ecol. Evol. 2007, 22, 95–102. [Google Scholar] [CrossRef] [PubMed]
- United Nations, Department of Economic and Social Affairs, Population Division (2014). World Urbanization Prospects: The 2014 Revision, Highlights (ST/ESA/SER.A/352); United Nations: New York, NY, USA, 2014. [Google Scholar]
- Allen, T.; Murray, K.A.; Zambrana-Torrelio, C.; Morse, S.S.; Rondinini, C.; Di Marco, M.; Breit, N.; Olival, K.J.; Daszak, P. Global Hotspots and Correlates of Emerging Zoonotic Diseases. Nat. Commun. 2017, 8, 1124. [Google Scholar] [CrossRef] [PubMed]
- Medlock, J.M.; Hansford, K.M.; Bormane, A.; Derdakova, M.; Estrada-Peña, A.; George, J.-C.; Golovljova, I.; Jaenson, T.G.T.; Jensen, J.-K.; Jensen, P.M.; et al. Driving Forces for Changes in Geographical Distribution of Ixodes ricinus Ticks in Europe. Parasites Vectors 2013, 6, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heyman, P.; Cochez, C.; Hofhuis, A.; van der Giessen, J.; Sprong, H.; Porter, S.R.; Losson, B.; Saegerman, C.; Donoso-Mantke, O.; Niedrig, M.; et al. A Clear and Present Danger: Tick-Borne Diseases in Europe. Expert Rev. Anti-Infect. Ther. 2010, 8, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Rizzoli, A.; Hauffe, H.; Carpi, G.; Vourc, H.G.; Neteler, M.; Rosa, R. Lyme Borreliosis in Europe. Euro Surveill. 2011, 16. [Google Scholar] [CrossRef]
- Brouqui, P.; Parola, P.; Fournier, P.E.; Raoult, D. Spotted Fever Rickettsioses in Southern and Eastern Europe. FEMS Immunol. Med. Microbiol. 2007, 49, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Takken, W.; van Vliet, A.J.H.; Verhulst, N.O.; Jacobs, F.H.H.; Gassner, F.; Hartemink, N.; Mulder, S.; Sprong, H. Acarological Risk of Borrelia burgdorferi Sensu Lato Infections across Space and Time in The Netherlands. Vector Borne Zoonotic Dis. 2017, 17, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Mannelli, A.; Boggiatto, G.; Grego, E.; Cinco, M.; Murgia, R.; Stefanelli, S.; De Meneghi, D.; Rosati, S. Acarological Risk of Exposure to Agents of Tick-Borne Zoonoses in the First Recognized Italian Focus of Lyme Borreliosis. Epidemiol. Infect. 2003, 131, 1139–1147. [Google Scholar] [CrossRef] [PubMed]
- Randolph, S.E.; EDEN-TBD Sub-Project Team. Human Activities Predominate in Determining Changing Incidence of Tick-Borne Encephalitis in Europe. Euro. Surveill. 2010, 15, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Cagnacci, F.; Bolzoni, L.; Rosà, R.; Carpi, G.; Hauffe, H.C.; Valent, M.; Tagliapietra, V.; Kazimirova, M.; Koci, J.; Stanko, M.; et al. Effects of Deer Density on Tick Infestation of Rodents and the Hazard of Tick-Borne Encephalitis. I: Empirical Assessment. Int. J. Parasitol. 2012, 42, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Ostfeld, R.S.; Levi, T.; Jolles, A.E.; Martin, L.B.; Hosseini, P.R.; Keesing, F. Life History and Demographic Drivers of Reservoir Competence for Three Tick-Borne Zoonotic Pathogens. PLoS ONE 2014, 9, e107387. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Fons, F.; Fernández-de-Mera, I.G.; Acevedo, P.; Gortázar, C.; de la Fuente, J. Factors Driving the Abundance of Ixodes ricinus Ticks and the Prevalence of Zoonotic I. Ricinus-Borne Pathogens in Natural Foci. Appl. Environ. Microbiol. 2012, 78, 2669–2676. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Peña, A.; Estrada-Sánchez, A.; de la Fuente, J. A Global Set of Fourier-Transformed Remotely Sensed Covariates for the Description of Abiotic Niche in Epidemiological Studies of Tick Vector Species. Parasites Vectors 2014, 7, 302. [Google Scholar] [CrossRef] [PubMed]
- Daniel, M.; Vráblík, T.; Valter, J.; Kríz, B.; Danielová, V. The TICKPRO Computer Program for Predicting Ixodes ricinus Host-Seeking Activity and the Warning System Published on Websites. Cent. Eur. J. Public Health 2010, 18, 230–236. [Google Scholar] [PubMed]
- Estrada-Peña, A. Geostatistics and Remote Sensing Using NOAA-AVHRR Satellite Imagery as Predictive Tools in Tick Distribution and Habitat Suitability Estimations for Boophilus microplus (Acari: Ixodidae) in South America. National Oceanographic and Atmosphere Administration-Advanced Very High Resolution Radiometer. Vet. Parasitol. 1999, 81, 73–82. [Google Scholar] [PubMed]
- Dobson, A.D.M.; Finnie, T.J.R.; Randolph, S.E. A Modified Matrix Model to Describe the Seasonal Population Ecology of the European Tick Ixodes ricinus. J. Appl. Ecol. 2011, 48, 1017–1028. [Google Scholar] [CrossRef]
- Palniyandi, M. The Role of Remote Sensing and GIS for Spatial Prediction of Vector-Borne Diseases Transmission: A Systematic Review. J. Vector Borne Dis. 2012, 49, 197–204. [Google Scholar]
- Estrada-Peña, A.; Estrada-Sánchez, D. Deconstructing Ixodes ricinus: A Partial Matrix Model Allowing Mapping of Tick Development, Mortality and Activity Rates. Med. Vet. Entomol. 2014, 28, 35–49. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Peña, A.; Estrada-Sánchez, A.; Estrada-Sánchez, D. Methodological Caveats in the Environmental Modelling and Projections of Climate Niche for Ticks, with Examples for Ixodes ricinus (Ixodidae). Vet. Parasitol. 2015, 208, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Peña, A.; Alexander, N.; Wint, G.R.W. Perspectives on Modelling the Distribution of Ticks for Large Areas: So Far so Good? Parasites Vectors 2016, 9, 179. [Google Scholar] [CrossRef] [PubMed]
- CLC Illustrated Nomenclature Guidelines. Available online: https://land.copernicus.eu/user-corner/technical-library/corine-land-cover-nomenclature-guidelines/html/ (accessed on 27 February 2018).
- Global Multi-Resolution Terrain Elevation Data 2010 (GMTED2010). Available online: https://lta.cr.usgs.gov/GMTED2010 (accessed on 27 February 2018).
- Filippova, N.A. Ixodid Ticks of the Subfamily Ixodinae; Fauna USSR-Acarina; Nauka Leningrad: Moscow, Russia, 1977; Volume IV. [Google Scholar]
- Siuda, K. Kleszcze Polski (Acari: Ixodida) Systematyka i Rozmieszczenie; Polskie Towarzystwo Parazytologiczne: Warszawa, Poland, 1993. [Google Scholar]
- Hillyard, P.D. Ticks of North-West Europe; The Dorset Press: Dorchester, UK, 1996. [Google Scholar]
- Estrada-Pena, A.; Bouattour, A.; Camicas, J.-L.; Walker, A.R. Ticks of Domestic Animals in the Mediterranean Region. A Guide to Identification of Species; University of Zaragoza: Zaragoza, Spain, 2004. [Google Scholar]
- Rijpkema, S.G.; Molkenboer, M.J.; Schouls, L.M.; Jongejan, F.; Schellekens, J.F. Simultaneous Detection and Genotyping of Three Genomic Groups of Borrelia burgdorferi Sensu Lato in Dutch Ixodes ricinus Ticks by Characterization of the Amplified Intergenic Spacer Region between 5S and 23S rRNA Genes. J. Clin. Microbiol. 1995, 33, 3091–3095. [Google Scholar] [PubMed]
- Massung, R.F.; Slater, K.; Owens, J.H.; Nicholson, W.L.; Mather, T.N.; Solberg, V.B.; Olson, J.G. Nested PCR Assay for Detection of Granulocytic Ehrlichiae. J. Clin. Microbiol. 1998, 36, 1090–1095. [Google Scholar] [PubMed]
- Reye, A.L.; Stegniy, V.; Mishaeva, N.P.; Velhin, S.; Hübschen, J.M.; Ignatyev, G.; Muller, C.P. Prevalence of Tick-Borne Pathogens in Ixodes ricinus and Dermacentor reticulatus Ticks from Different Geographical Locations in Belarus. PLoS ONE 2013, 8, e54476. [Google Scholar] [CrossRef] [PubMed]
- Baráková, I.; Derdáková, M.; Selyemová, D.; Chvostáč, M.; Špitalská, E.; Rosso, F.; Collini, M.; Rosà, R.; Tagliapietra, V.; Girardi, M.; et al. Tick-Borne Pathogens and Their Reservoir Hosts in Northern Italy. Ticks Tick-Borne Dis. 2017, 9, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Derdakova, M.; Beati, L.; Pet’ko, B.; Stanko, M.; Fish, D. Genetic Variability within Borrelia burgdorferi Sensu Lato Genospecies Established by PCR-Single-Strand Conformation Polymorphism Analysis of the rrfA-rrlB Intergenic Spacer in Ixodes ricinus Ticks from the Czech Republic. Appl. Environ. Microbiol. 2003, 69, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Courtney, J.W.; Kostelnik, L.M.; Zeidner, N.S.; Massung, R.F. Multiplex Real-Time PCR for Detection of Anaplasma phagocytophilum and Borrelia burgdorferi. J. Clin. Microbiol. 2004, 42, 3164–3168. [Google Scholar] [CrossRef] [PubMed]
- Regnery, R.L.; Spruill, C.L.; Plikaytis, B.D. Genotypic Identification of Rickettsiae and Estimation of Intraspecies Sequence Divergence for Portions of Two Rickettsial Genes. J. Bacteriol. 1991, 173, 1576–1589. [Google Scholar] [CrossRef] [PubMed]
- Overzier, E.; Pfister, K.; Thiel, C.; Herb, I.; Mahling, M.; Silaghi, C. Diversity of Babesia and Rickettsia Species in Questing Ixodes ricinus: A Longitudinal Study in Urban, Pasture, and Natural Habitats. Vector Borne Zoonotic Dis. 2013, 13, 559–564. [Google Scholar] [CrossRef] [PubMed]
- Overzier, E.; Pfister, K.; Thiel, C.; Herb, I.; Mahling, M.; Silaghi, C. Anaplasma phagocytophilum in Questing Ixodes ricinus Ticks: Comparison of Prevalences and Partial 16S rRNA Gene Variants in Urban, Pasture, and Natural Habitats. Appl. Environ. Microbiol. 2013, 79, 1730–1734. [Google Scholar] [CrossRef] [PubMed]
- Blaňarová, L.; Stanko, M.; Carpi, G.; Miklisová, D.; Víchová, B.; Mošanský, L.; Bona, M.; Derdáková, M. Distinct Anaplasma phagocytophilum Genotypes Associated with Ixodes Trianguliceps Ticks and Rodents in Central Europe. Ticks Tick-Borne Dis. 2014, 5, 928–938. [Google Scholar] [CrossRef] [PubMed]
- Svitálková, Z.; Haruštiaková, D.; Mahríková, L.; Berthová, L.; Slovák, M.; Kocianová, E.; Kazimírová, M. Anaplasma phagocytophilum Prevalence in Ticks and Rodents in an Urban and Natural Habitat in South-Western Slovakia. Parasites Vectors 2015, 8, 276. [Google Scholar] [CrossRef] [PubMed]
- Špitalská, E.; Stanko, M.; Mošanský, L.; Kraljik, J.; Miklisová, D.; Mahríková, L.; Bona, M.; Kazimírová, M. Seasonal Analysis of Rickettsia Species in Ticks in an Agricultural Site of Slovakia. Exp. Appl. Acarol. 2016, 68, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Minichová, L.; Hamšíková, Z.; Mahríková, L.; Slovák, M.; Kocianová, E.; Kazimírová, M.; Škultéty, Ľ.; Štefanidesová, K.; Špitalská, E. Molecular Evidence of Rickettsia spp. in Ixodid Ticks and Rodents in Suburban, Natural and Rural Habitats in Slovakia. Parasites Vectors 2017, 10, 158. [Google Scholar] [CrossRef] [PubMed]
- Venclíková, K.; Mendel, J.; Betášová, L.; Blažejová, H.; Jedličková, P.; Straková, P.; Hubálek, Z.; Rudolf, I. Neglected Tick-Borne Pathogens in the Czech Republic, 2011–2014. Ticks Tick-Borne Dis. 2016, 7, 107–112. [Google Scholar] [CrossRef] [PubMed]
- Hornok, S.; Meli, M.L.; Gönczi, E.; Halász, E.; Takács, N.; Farkas, R.; Hofmann-Lehmann, R. Occurrence of Ticks and Prevalence of Anaplasma phagocytophilum and Borrelia burgdorferi s.l. in Three Types of Urban Biotopes: Forests, Parks and Cemeteries. Ticks Tick-Borne Dis. 2014, 5, 785–789. [Google Scholar] [CrossRef] [PubMed]
- Schulze, T.L.; Jordan, R.A. Meteorologically Mediated Diurnal Questing of Ixodes scapularis and Amblyomma americanum (Acari: Ixodidae) Nymphs. J. Med. Entomol. 2003, 40, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Perret, J.-L.; Rais, O.; Gern, L. Influence of Climate on the Proportion of Ixodes ricinus Nymphs and Adults Questing in a Tick Population. J. Med. Entomol. 2004, 41, 361–365. [Google Scholar] [CrossRef] [PubMed]
- Barrios, J.M.; Verstraeten, W.W.; Maes, P.; Clement, J.; Aerts, J.M.; Farifteh, J.; Lagrou, K.; Van Ranst, M.; Coppin, P. Remotely Sensed Vegetation Moisture as Explanatory Variable of Lyme Borreliosis Incidence. Int. J. Appl. Earth Obs. Geoinf. 2012, 18, 1–12. [Google Scholar] [CrossRef]
- Neteler, M.; Hamish Bowman, M.; Landa, M.; Metz, M. GRASS GIS: A Multi-Purpose Open Source GIS. Environ. Model. Softw. 2012, 31, 124–130. [Google Scholar] [CrossRef]
- Metz, M.; Rocchini, D.; Neteler, M. Surface Temperatures at the Continental Scale: Tracking Changes with Remote Sensing at Unprecedented Detail. Remote Sens. 2014, 6, 3822–3840. [Google Scholar] [CrossRef]
- Rosà, R.; Marini, G.; Bolzoni, L.; Neteler, M.; Metz, M.; Delucchi, L.; Chadwick, E.A.; Balbo, L.; Mosca, A.; Giacobini, M.; et al. Early Warning of West Nile Virus Mosquito Vector: Climate and Land Use Models Successfully Explain Phenology and Abundance of Culex Pipiens Mosquitoes in North-Western Italy. Parasites Vectors 2014, 7, 269. [Google Scholar] [CrossRef] [PubMed]
- Randolph, S.E.; Green, R.M.; Peacey, M.F.; Rogers, D.J. Seasonal Synchrony: The Key to Tick-Borne Encephalitis Foci Identified by Satellite Data. Parasitology 2000, 121 (Pt 1), 15–23. [Google Scholar] [CrossRef] [PubMed]
- The European Climate Assessment & Dataset. Available online: https://www.knmi.nl/kennis-en-datacentrum/project/the-european-climate-assessment-dataset (accessed on 27 February 2018).
- Haylock, M.R.; Hofstra, N.; Klein, A.M.; Klok, E.J.; Jones, P.D.; New, M. A European Daily High-Resolution Gridded Data Set of Surface Temperature and Precipitation for 1950–2006. J. Geophys. Res. 2008, 113. [Google Scholar] [CrossRef]
- Roerink, G.J.; Menenti, M.; Verhoef, W. Reconstructing Cloudfree NDVI Composites Using Fourier Analysis of Time Series. Int. J. Remote Sens. 2000, 21, 1911–1917. [Google Scholar] [CrossRef]
- R: The R Project for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 12 October 2017).
- IJERPH_Rosa_2018. Available online: www.geodati.fmach.it/IJERPH_Rosa_2018 (accessed on 27 February 2018).
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Maechler, M.; Bolker, B.M. Modeling Zero-Inflated Count Data with glmmTMB. bioRxiv 2017. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Usinglme4. J. Stat. Softw. 2015, 67. [Google Scholar] [CrossRef]
- Pan, Y.; Jackson, R.T. Ethnic Difference in the Relationship between Acute Inflammation and Serum Ferritin in US Adult Males. Epidemiol. Infect. 2008, 136, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, S.; Schielzeth, H. A general and simple method for obtaining R2 from Generalized Linear Mixed-effects Models. Methods Ecol. Evol. 2013, 4, 133–142. [Google Scholar] [CrossRef]
- Trout Fryxell, R.T.; Moore, J.E.; Collins, M.D.; Kwon, Y.; Jean-Philippe, S.R.; Schaeffer, S.M.; Odoi, A.; Kennedy, M.; Houston, A.E. Habitat and Vegetation Variables Are Not Enough When Predicting Tick Populations in the Southeastern United States. PLoS ONE 2015, 10, e0144092. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, R.; Rossini, P. On the Use of NDVI Profiles as a Tool for Agricultural Statistics: The Case Study of Wheat Yield Estimate and Forecast in Emilia Romagna. Remote Sens. Environ. 1993, 45, 311–326. [Google Scholar] [CrossRef]
- Hubálek, Z.; Halouzka, J.; Juricová, Z. Host-Seeking Activity of Ixodid Ticks in Relation to Weather Variables. J. Vector Ecol. 2003, 28, 159–165. [Google Scholar] [PubMed]
- Alonso-Carné, J.; García-Martín, A.; Estrada-Peña, A. Assessing the Statistical Relationships among Water-Derived Climate Variables, Rainfall, and Remotely Sensed Features of Vegetation: Implications for Evaluating the Habitat of Ticks. Exp. Appl. Acarol. 2015, 65, 107–124. [Google Scholar] [CrossRef] [PubMed]
- Bisanzio, D.; Amore, G.; Ragagli, C.; Tomassone, L.; Bertolotti, L.; Mannelli, A. Temporal Variations in the Usefulness of Normalized Difference Vegetation Index as a Predictor for Ixodes ricinus (Acari: Ixodidae) in a Borrelia Lusitaniae Focus in Tuscany, Central Italy. J. Med. Entomol. 2008, 45, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Olwoch, J.M.; Rautenbach, C.D.W.; Erasmus, B.F.N.; Engelbrecht, F.A.; Van Jaarsveld, A.S. Simulating Tick Distributions over Sub-Saharan Africa: The Use of Observed and Simulated Climate Surfaces. J. Biogeogr. 2003, 30, 1221–1232. [Google Scholar] [CrossRef]
- Adejinmi, J.O. Effect of Water Flooding on the Oviposition Capacity of Engorged Adult Females and Hatchability of Eggs of Dog Ticks: Rhipicephalus Sanguineus and Haemaphysalis Leachi Leachi. J. Parasitol. Res. 2011, 2011, 824162. [Google Scholar] [CrossRef] [PubMed]
- Sutherst, R.W. An Experimental Investigation into the Effects of Flooding on the Ixodid Tick Boophilus microplus (Canestrini). Oecologia 1971, 6, 208–222. [Google Scholar] [CrossRef] [PubMed]
- Rosef, O.; Paulauskas, A.; Radzijevskaja, J. Prevalence of Borrelia burgdorferi Sensu Lato and Anaplasma phagocytophilum in Questing Ixodes ricinus Ticks in Relation to the Density of Wild Cervids. Acta Vet. Scand. 2009, 51, 47. [Google Scholar] [CrossRef] [PubMed]
- Rosà, R.; Pugliese, A.; Norman, R.; Hudson, P.J. Thresholds for Disease Persistence in Models for Tick-Borne Infections Including Non-Viraemic Transmission, Extended Feeding and Tick Aggregation. J. Theor. Biol. 2003, 224, 359–376. [Google Scholar] [CrossRef]
- Kazimírová, M.; Hamšíková, Z.; Kocianová, E.; Marini, G.; Mojšová, M.; Mahríková, L.; Berthová, L.; Slovák, M.; Rosá, R. Relative Density of Host-Seeking Ticks in Different Habitat Types of South-Western Slovakia. Exp. Appl. Acarol. 2016, 69, 205–224. [Google Scholar] [CrossRef] [PubMed]
- Földvári, G.; Jahfari, S.; Rigó, K.; Jablonszky, M.; Szekeres, S.; Majoros, G.; Tóth, M.; Molnár, V.; Coipan, E.C.; Sprong, H. Candidatus Neoehrlichia Mikurensis and Anaplasma phagocytophilumin Urban Hedgehogs. Emerg. Infect. Dis. 2014, 20, 496–498. [Google Scholar] [CrossRef] [PubMed]
- Rizzoli, A.P.; Silaghi, C.; Obiegala, A.; Rudolf, I.; Hubálek, Z.; Földvári, G.; Plantard, O.; Vayssier-Taussat, M.; Bonnet, S.; Špitalská, E.; et al. Ixodes ricinus and its transmitted pathogens in urban and peri-urban areas in Europe: New hazards and relevance for public health. Front. Public Health 2014, 2, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Raoult, D.; Roux, V. Rickettsioses as Paradigms of New or Emerging Infectious Diseases. Clin. Microbiol. Rev. 1997, 10, 694–719. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
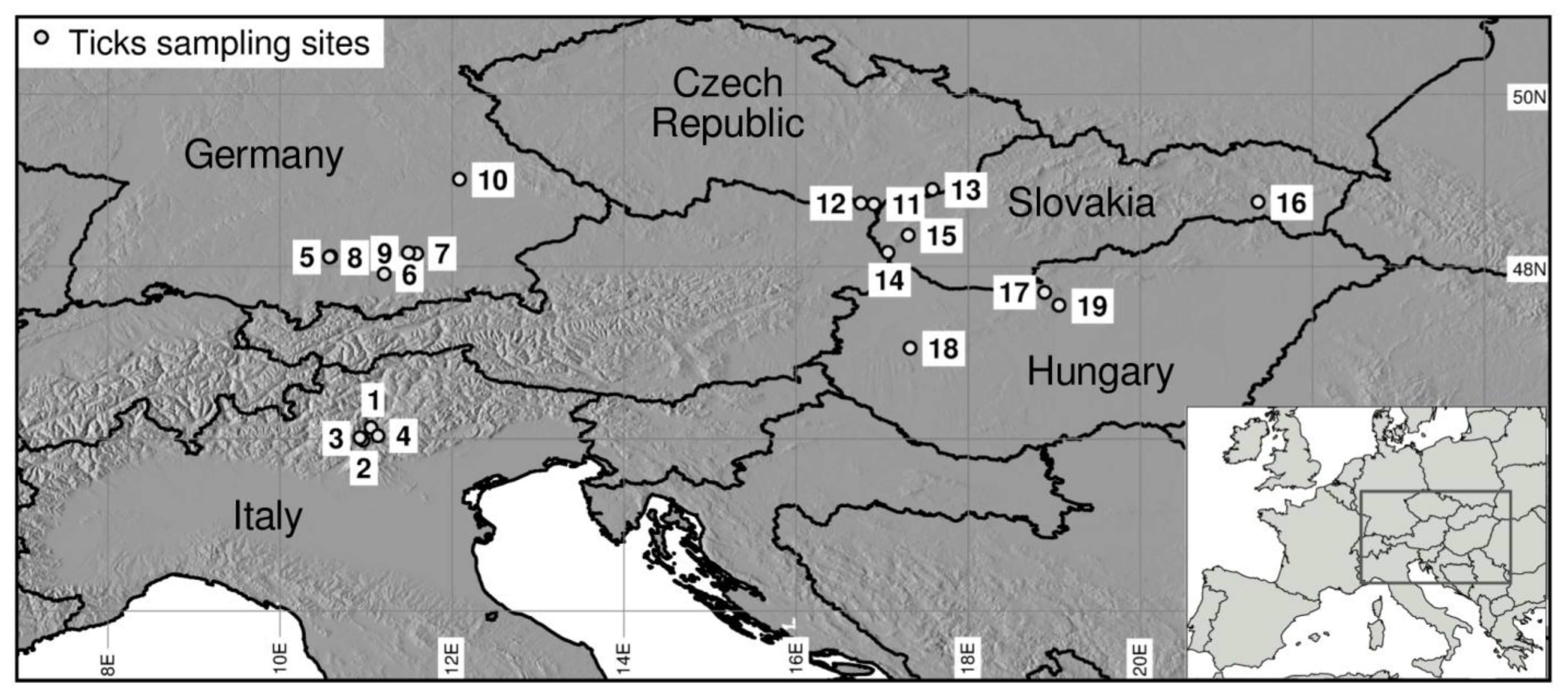
| Country | Site Number | Sampling Site | Land Use Category | Altitude (m a.s.l.) | Latitude | Longitude |
|---|---|---|---|---|---|---|
| Italy | 1 | Lamar | Natural | 784 | 46.128726 | 11.058944 |
| Italy | 2 | Cavedine | Agricultural | 717 | 45.985402 | 10.963142 |
| Italy | 3 | Pietramurata | Natural | 468 | 46.013258 | 10.927981 |
| Italy | 4 | Trento | Urban | 285 | 46.035187 | 11.139236 |
| Germany | 5 | Tussenhausen | Natural | 640 | 48.118279 | 10.589147 |
| Germany | 6 | Kerschlach | Agricultural | 724 | 47.917142 | 11.212342 |
| Germany | 7 | Englischer Garten | Urban | 514 | 48.150481 | 11.590053 |
| Germany | 8 | Berg Starnberg | Urban | 659 | 48.110117 | 10.575944 |
| Germany | 9 | Nymphenburger Schlosspark | Urban | 522 | 48.160814 | 11.492586 |
| Germany | 10 | Dörnbergpark Regensburg | Urban | 345 | 49.015478 | 12.085803 |
| Czech Republic | 11 | Pohansko | Natural | 162 | 48.727133 | 16.902319 |
| Czech Republic | 12 | Valtice | Urban | 215 | 48.734911 | 16.753142 |
| Czech Republic | 13 | Suchov | Agricultural | 426 | 48.897442 | 17.581928 |
| Slovakia | 14 | Bratislava | Urban | 184 | 48.166667 | 17.066667 |
| Slovakia | 15 | Fúgeľka | Natural | 386 | 48.366667 | 17.300000 |
| Slovakia | 16 | Rozhanovce | Agricultural | 280 | 48.750000 | 21.366667 |
| Hungary | 17 | Pilisszentkereszt | Natural | 468 | 47.700833 | 18.884722 |
| Hungary | 18 | Csabrendek | Agricultural | 159 | 47.053889 | 17.323333 |
| Hungary | 19 | Budapest | Urban | 105 | 47.550278 | 19.052778 |
| Explanatory Variable | Estimate | Std. Error | z-Value | Pr(>|z|) |
|---|---|---|---|---|
| Intercept | 5.457 | 0.260 | 21.026 | <0.001 *** |
| NDVI (Apr–May–Jun) | 0.264 | 0.124 | 2.131 | 0.033 * |
| Accumulated precipitation (Oct–Nov–Dec, previous year) | −0.265 | 0.149 | −1.779 | 0.075 |
| Model | Explanatory Variable | Estimate | Std. Error | t Value | Pr(>|t|) |
|---|---|---|---|---|---|
| DIN for Borrelia | Intercept | 6.245 | 0.148 | 42.216 | <0.001 *** |
| Habitat type Natural | 0.779 | 0.209 | 1.724 | 0.002 ** | |
| Habitat type Urban | −0.215 | 0.245 | −0.878 | 0.395 | |
| Accumulated Precipitation (Jan-Feb-Mar, previous year) | −0.310 | 0.095 | −3.268 | 0.006 ** | |
| DIN for Anaplasma | Intercept | 2.478 | 0.615 | 4.026 | 0.006 ** |
| Habitat type Natural | 0.980 | 0.540 | 1.814 | 0.080 | |
| Habitat type Urban | 1.882 | 0.591 | 3.185 | 0.003 ** | |
| NDVI (January) | 0.740 | 0.261 | 2.831 | 0.008 ** | |
| DIN for Rickettsia | Intercept | 5.214 | 0.405 | 4.267 | <0.001 *** |
| NDVI (March) | 0.488 | 0.153 | 29.603 | 0.003 ** |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosà, R.; Andreo, V.; Tagliapietra, V.; Baráková, I.; Arnoldi, D.; Hauffe, H.C.; Manica, M.; Rosso, F.; Blaňarová, L.; Bona, M.; et al. Effect of Climate and Land Use on the Spatio-Temporal Variability of Tick-Borne Bacteria in Europe. Int. J. Environ. Res. Public Health 2018, 15, 732. https://doi.org/10.3390/ijerph15040732
Rosà R, Andreo V, Tagliapietra V, Baráková I, Arnoldi D, Hauffe HC, Manica M, Rosso F, Blaňarová L, Bona M, et al. Effect of Climate and Land Use on the Spatio-Temporal Variability of Tick-Borne Bacteria in Europe. International Journal of Environmental Research and Public Health. 2018; 15(4):732. https://doi.org/10.3390/ijerph15040732
Chicago/Turabian StyleRosà, Roberto, Veronica Andreo, Valentina Tagliapietra, Ivana Baráková, Daniele Arnoldi, Heidi Christine Hauffe, Mattia Manica, Fausta Rosso, Lucia Blaňarová, Martin Bona, and et al. 2018. "Effect of Climate and Land Use on the Spatio-Temporal Variability of Tick-Borne Bacteria in Europe" International Journal of Environmental Research and Public Health 15, no. 4: 732. https://doi.org/10.3390/ijerph15040732
APA StyleRosà, R., Andreo, V., Tagliapietra, V., Baráková, I., Arnoldi, D., Hauffe, H. C., Manica, M., Rosso, F., Blaňarová, L., Bona, M., Derdáková, M., Hamšíková, Z., Kazimírová, M., Kraljik, J., Kocianová, E., Mahríková, L., Minichová, L., Mošanský, L., Slovák, M., ... Rizzoli, A. (2018). Effect of Climate and Land Use on the Spatio-Temporal Variability of Tick-Borne Bacteria in Europe. International Journal of Environmental Research and Public Health, 15(4), 732. https://doi.org/10.3390/ijerph15040732