Reconciling the Entomological Hazard and Disease Risk in the Lyme Disease System

Abstract
:1. Introduction
2. Materials and Methods
2.1. LD Risk Model
2.2. Entomological Risk and Human Exposure Functions
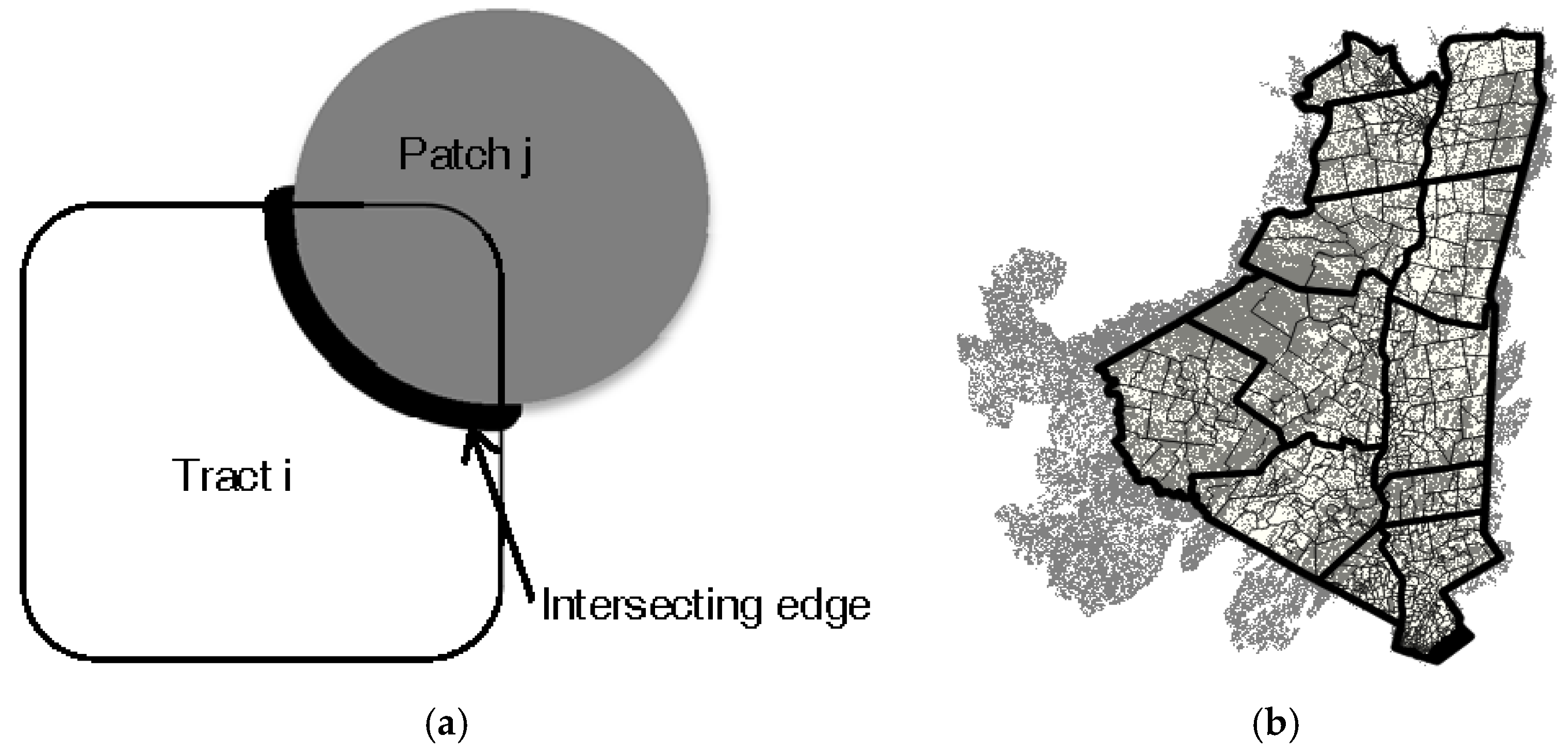
- We considered Expj as either proportional to the perimeter of patch j that falls within range of i (proportional to the probability of entering patch j, assuming humans in population unit i move by random walk), proportional to the area of patch j that falls within the range of i (proportional to the relative amount of time spent in patch j), or as a constant.
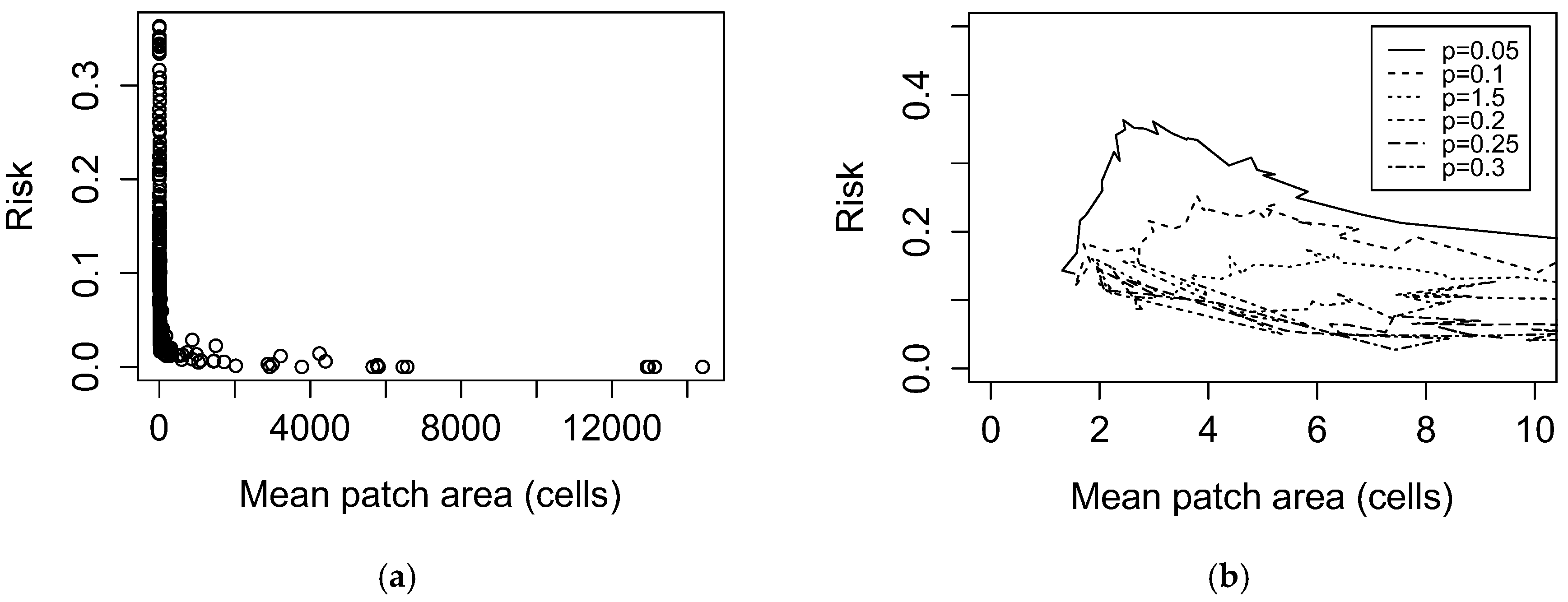
- We considered Entj as either a negative exponential function of the area of patch j (as hypothesized in Allan et al., 2003 [9]), a linear function of the area of patch j, or as a constant.
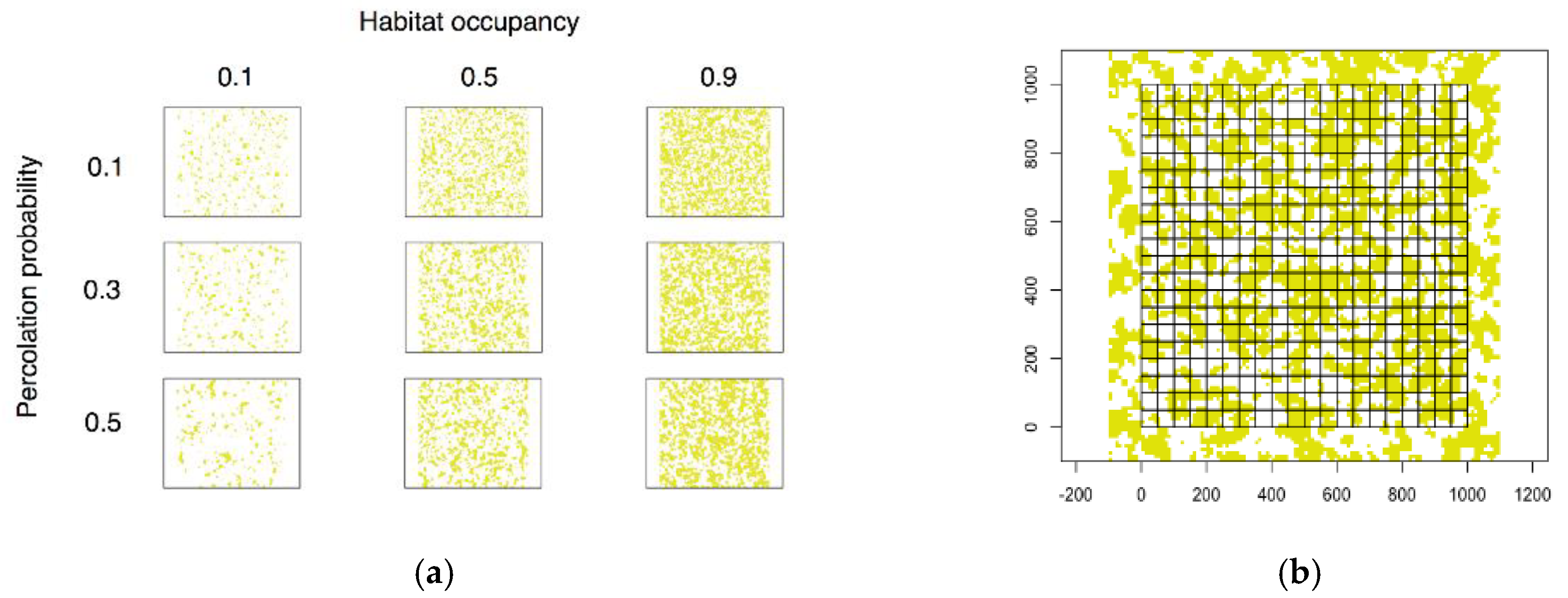
2.3. Simulated Landscapes
2.4. Landscape Analysis
2.5. Risk Index Calculation
- exposure constant, entomological risk as a negative exponential function of patch area:
- exposure directly related to intersecting patch perimeter, entomological risk as a negative exponential function of patch area:
- exposure directly related to intersecting patch area, entomological risk as a negative exponential function of patch area:
- exposure directly related to intersecting patch perimeter, entomological risk constant:
- exposure directly related to intersecting patch area, entomological risk constant:
- exposure constant, entomological risk directly related to area:
- exposure directly related to intersecting patch perimeter, entomological risk directly related to area:
- exposure directly related to intersecting patch area, entomological risk directly related to area:
2.6. County LDI
2.7. Model Evaluation
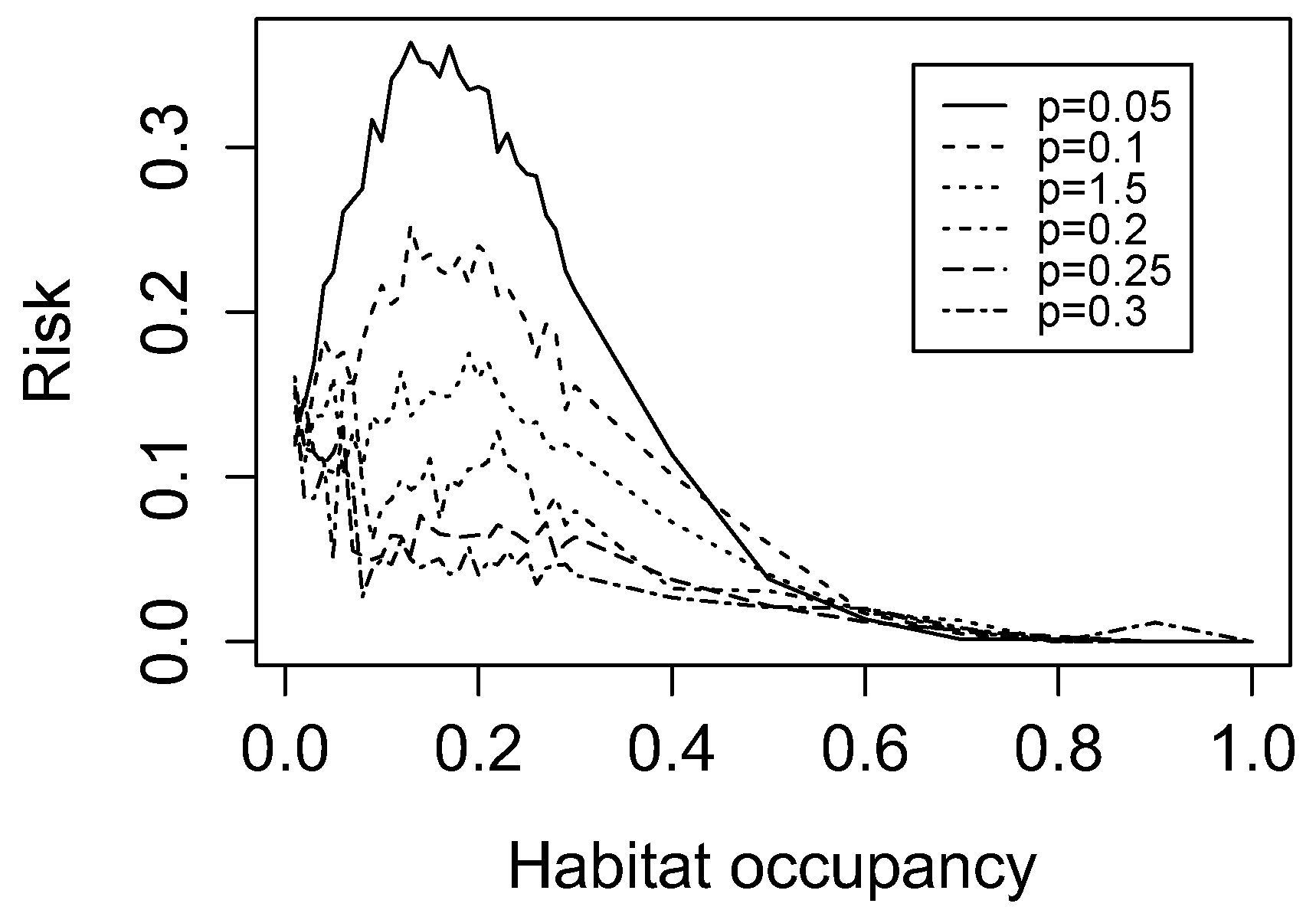
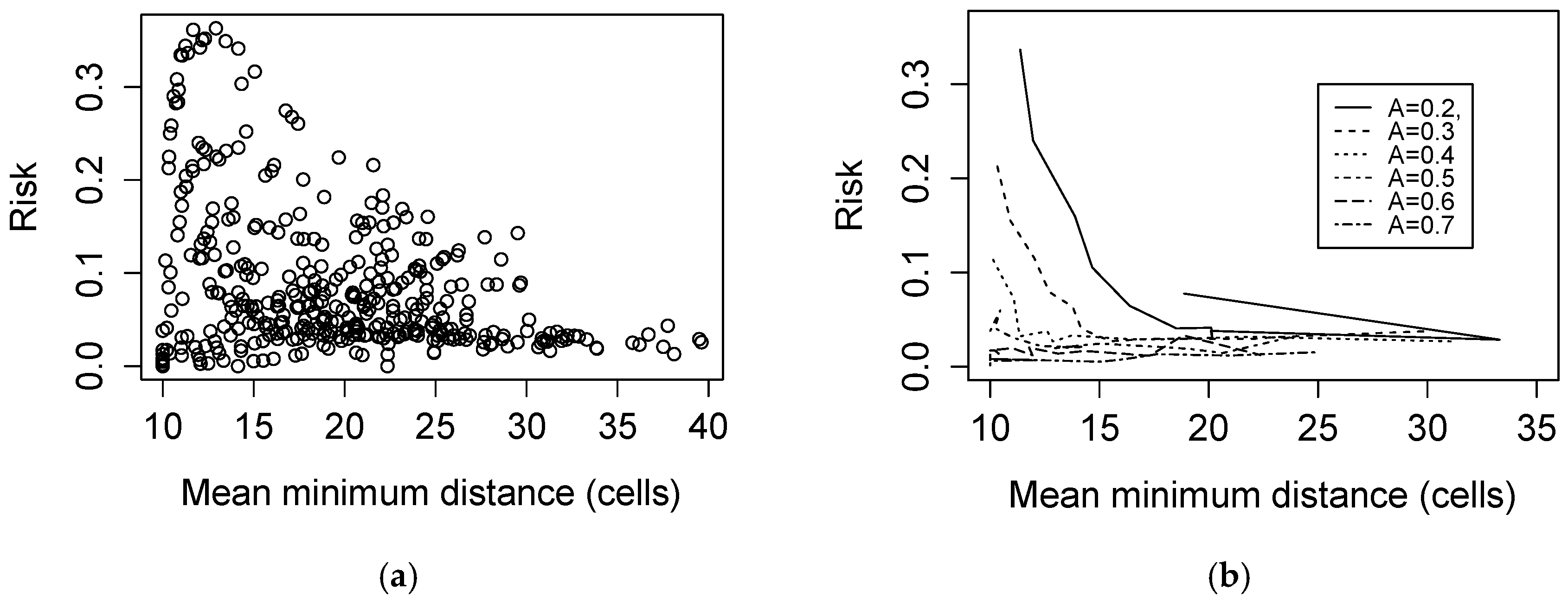
3. Results
3.1. Simulated Landscapes
3.2. Landscape Characterization
3.3. Spatial Structure of LDI
3.4. Model Evaluation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Centers for Disease Control and Prevention. County-Level Lyme Disease Data from 2000–2016. Available online: https://www.cdc.gov/lyme/stats/index.html (accessed on 1 December 2017).
- Steere, A.C.; Coburn, J.; Glickstein, L. The emergence of Lyme disease. J. Clin. Investig. 2004, 113, 1093. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.A. Summary of Notifiable Infectious Diseases and Conditions—United States, 2015. Morb. Mortal. Wkly. Rep. 2017, 64, 1–143. [Google Scholar] [CrossRef] [PubMed]
- Ogden, N.; Koffi, J.; Pelcat, Y.; Lindsay, L. Environmental risk from Lyme disease in central and eastern Canada: A summary of recent surveillance information. Can. Commun. Dis. Rep. 2014, 40, 74–82. [Google Scholar] [PubMed]
- Kilpatrick, A.M.; Dobson, A.D.; Levi, T.; Salkeld, D.J.; Swei, A.; Ginsberg, H.S.; Kjemtrup, A.; Padgett, K.A.; Jensen, P.M.; Fish, D.; et al. Lyme disease ecology in a changing world: Consensus, uncertainty and critical gaps for improving control. Philos. Trans. R. Soc. B 2017, 372, 20160117. [Google Scholar] [CrossRef] [PubMed]
- Piesman, J. Ecology of Borrelia burgdorferi sensu lato in North America. In Lyme Borreliosis: Biology, Epidemiology, and Control; Gray, J.S., Kahl, O., Lane, R.S., Stanek, G., Eds.; CAB International: Wallingford, UK, 2002; pp. 223–249. [Google Scholar]
- Davis, S.; Bent, S.J. Loop analysis for pathogens: Niche partitioning in the transmission graph for pathogens of the North American tick Ixodes scapularis. J. Theor. Biol. 2011, 269, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Ripoche, M.; Gasmi, S.; Adam-Poupart, A.; Koffi, J.K.; Lindsay, L.R.; Ludwig, A.; Milord, F.; Ogden, N.H.; Thivierge, K.; Leighton, P.A. Passive Tick Surveillance Provides an Accurate Early Signal of Emerging Lyme Disease Risk and Human Cases in Southern Canada. J. Med. Entomol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Allan, B.F.; Keesing, F.; Ostfeld, R.S. Effect of Forest Fragmentation on Lyme Disease Risk. Conserv. Biol. 2003, 17, 267–272. [Google Scholar] [CrossRef]
- Brownstein, J.S.; Skelly, D.K.; Holford, T.R.; Fish, D. Forest fragmentation predicts local scale heterogeneity of Lyme disease risk. Oecologia 2005, 146, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Zolnik, C.P.; Falco, R.C.; Kolokotronis, S.-O.; Daniels, T.J. No observed effect of landscape fragmentation on pathogen infection prevalence in blacklegged ticks (Ixodes scapularis) in the Northeastern United States. PLoS ONE 2015, 10, e0139473. [Google Scholar] [CrossRef] [PubMed]
- LoGiudice, K.; Duerr, S.T.; Newhouse, M.J.; Schmidt, K.A.; Killilea, M.E.; Ostfeld, R.S. Impact of host community composition on Lyme disease risk. Ecology 2008, 89, 2841–2849. [Google Scholar] [CrossRef] [PubMed]
- Frank, D.H.; Fish, D.; Moy, F.H. Landscape features associated with Lyme disease risk in a suburban residential environment. Landsc. Ecol. 1998, 13, 27–36. [Google Scholar] [CrossRef]
- Li, S.; Hartemink, N.; Speybroeck, N.; Vanwambeke, S.O. Consequences of Landscape Fragmentation on Lyme Disease Risk: A Cellular Automata Approach. PLoS ONE 2012, 7, e39612. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, P.R.; Mills, J.N.; Prieur-Richard, A.-H.; Ezenwa, V.O.; Bailly, X.; Rizzoli, A.; Suzán, G.; Vittecoq, M.; García-Peña, G.E.; Daszak, P. Does the impact of biodiversity differ between emerging and endemic pathogens? The need to separate the concepts of hazard and risk. Philos. Trans. R. Soc. B 2017, 372, 20160129. [Google Scholar] [CrossRef] [PubMed]
- Finch, C.; Al-Damluji, M.S.; Krause, P.J.; Niccolai, L.; Steeves, T.; O’Keefe, C.F.; Diuk-Wasser, M.A. Integrated assessment of behavioral and environmental risk factors for Lyme disease infection on Block Island, Rhode Island. PLoS ONE 2014, 9, e84758. [Google Scholar] [CrossRef] [PubMed]
- Aenishaenslin, C.; Bouchard, C.; Koffi, J.K.; Ogden, N.H. Exposure and preventive behaviours toward ticks and Lyme disease in Canada: Results from a first national survey. Ticks Tick Borne Dis. 2017, 8, 112–118. [Google Scholar] [CrossRef] [PubMed]
- Nelson, C.A.; Saha, S.; Kugeler, K.J.; Delorey, M.J.; Shankar, M.B.; Hinckley, A.F.; Mead, P.S. Incidence of clinician-diagnosed Lyme disease, United States, 2005–2010. Emerg. Infect. Dis. 2015, 21, 1625. [Google Scholar] [CrossRef] [PubMed]
- Diuk-Wasser, M.A.; Liu, Y.; Steeves, T.K.; Folsom-O’Keefe, C.; Dardick, K.R.; Lepore, T.; Bent, S.J.; Usmani-Brown, S.; Telford, S.R., III; Fish, D. Monitoring human babesiosis emergence through vector surveillance New England, USA. Emerg. Infect. Dis. 2014, 20, 225. [Google Scholar] [CrossRef] [PubMed]
- Pepin, K.M.; Eisen, R.J.; Mead, P.S.; Piesman, J.; Fish, D.; Hoen, A.G.; Barbour, A.G.; Hamer, S.; Diuk-Wasser, M.A. Geographic variation in the relationship between human Lyme disease incidence and density of infected host-seeking Ixodes scapularis nymphs in the Eastern United States. Am. J. Trop. Med. Hyg. 2012, 86, 1062–1071. [Google Scholar] [CrossRef] [PubMed]
- Connally, N.P.; Ginsberg, H.S.; Mather, T.N. Assessing peridomestic entomological factors as predictors for Lyme disease. J. Vector Ecol. 2006, 31, 364–370. [Google Scholar] [CrossRef]
- Wood, C.L.; Lafferty, K.D. Biodiversity and disease: A synthesis of ecological perspectives on Lyme disease transmission. Trends Ecol. Evol. 2013, 28, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Jackson, L.E.; Hilborn, E.D.; Thomas, J.C. Towards landscape design guidelines for reducing Lyme disease risk. Int. J. Epidemiol. 2006, 35, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Falco, R.C.; Fish, D. Ticks parasitizing humans in a Lyme disease endemic area of southern New York State. Am. J. Epidemiol. 1988, 128, 1146–1152. [Google Scholar] [CrossRef] [PubMed]
- Saura, S.; Martinez-Millán, J. Landscape patterns simulation with a modified random clusters method. Landsc. Ecol. 2000, 15, 661–678. [Google Scholar] [CrossRef]
- With, K.A. Using percolation theory to assess landscape connectivity and effects of habitat fragmentation. In Applying Landscape Ecology in Biological Conservation; Gutzwiller, K., Ed.; Springer Science+Business Media: New York, NY, USA, 2002; pp. 105–130. [Google Scholar]
- Homer, C.; Dewitz, J.; Yang, L.; Jin, S.; Danielson, P.; Xian, G.; Coulston, J.; Herold, N.; Wickham, J.; Megown, K. Completion of the 2011 National Land Cover Database for the conterminous United States–representing a decade of land cover change information. Photogramm. Eng. Remote Sens. 2015, 81, 345–354. [Google Scholar]
- Larsen, A.E.; MacDonald, A.J.; Plantinga, A.J. Lyme Disease Risk Influences Human Settlement in the Wildland–Urban Interface: Evidence from a Longitudinal Analysis of Counties in the Northeastern United States. Am. J. Trop. Med. Hyg. 2014, 91, 747–755. [Google Scholar] [CrossRef] [PubMed]
- NYS. Civil Boundaries Data. Available online: http://gis.ny.gov/gisdata/inventories/details.cfm?DSID=927 (accessed on 1 December 2017).
- NNDSS. National Notifiable Disease Surveillance System Surveillance Case Definitions, Lyme Disease (Borrelia burgdorferi). Available online: http://www.cdc.gov/nndss/conditions/lyme-disease/ (accessed on 1 December 2017).
- Waller, L.A.; Gotway, C.A. Applied Spatial Statistics for Public Health Data; John Wiley & Sons: Hoboken, NJ, USA, 2004; Volume 368. [Google Scholar]
- Dale, M.R.; Fortin, M.-J. Spatial Analysis: A Guide for Ecologists; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Ostfeld, R.S.; Keesing, F. Biodiversity and disease risk: The case of Lyme disease. Conserv. Biol. 2000, 14, 722–728. [Google Scholar] [CrossRef]
- Schmidt, K.A.; Ostfeld, R.S. Biodiversity and the dilution effect in disease ecology. Ecology 2001, 82, 609–619. [Google Scholar] [CrossRef]
- LoGiudice, K.; Ostfeld, R.S.; Schmidt, K.A.; Keesing, F. The ecology of infectious disease: Effects of host diversity and community composition on Lyme disease risk. Proc. Natl. Acad. Sci. USA 2003, 100, 567–571. [Google Scholar] [CrossRef] [PubMed]
- MacArthur, R.H.; Wilson, E.O. An equilibrium theory of insular zoogeography. Evolution 1963, 17, 373–387. [Google Scholar] [CrossRef]
- Lafferty, K.D.; Wood, C.L. It’s a myth that protection against disease is a strong and general service of biodiversity conservation: Response to Ostfeld and Keesing. Conserv. Biol. 2013, 14, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Randolph, S. Commentary on ‘A Candide response to Panglossian accusations by Randolph and Dobson: Biodiversity buffers disease’ by Dr. R. Ostfeld (Parasitology 2013, in press). Parasitology 2013, 140, 1199–1200. [Google Scholar] [PubMed]
- Randolph, S.E.; Dobson, A.D.M. Pangloss revisited: A critique of the dilution effect and the biodiversity-buffers-disease paradigm. Parasitology 2012, 139, 847–863. [Google Scholar] [CrossRef] [PubMed]
- Ostfeld, R.S.; Keesing, F. Straw men don’t get Lyme disease: Response to Wood and Lafferty. Trends Ecol. Evol. 2013, 28, 502–503. [Google Scholar] [CrossRef] [PubMed]
- Linske, M.A.; Williams, S.C.; Stafford, K.C.; Ortega, I.M. Ixodes scapularis (Acari: Ixodidae) Reservoir Host Diversity and Abundance Impacts on Dilution of Borrelia burgdorferi (Spirochaetales: Spirochaetaceae) in Residential and Woodland Habitats in Connecticut, United States. J. Med. Entomol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Bouchard, C.; Aenishaenslin, C.; Rees, E.E.; Koffi, J.K.; Pelcat, Y.; Ripoche, M.; Milord, F.; Lindsay, L.R.; Ogden, N.H.; Leighton, P.A. Integrated Social-Behavioral and Ecological Risk Maps to Prioritize Local Public Health Responses to Lyme Disease. Environ. Health Perspect. 2018, 126, 47008. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| County | Population 2010 | Mean LDI 2000–2015 | Forest Area (ha) 2011 | Forest Perimeter (km) 2011 | No. Patches 2011 |
|---|---|---|---|---|---|
| Albany | 304,204 | 7.40 × 10−4 | 65.1426 | 378.75 | 1374 |
| Columbia | 63,096 | 6.76 × 10−3 | 83.0822 | 407.078 | 1637 |
| Dutchess | 297,488 | 2.49 × 10−3 | 111.7104 | 516.086 | 1835 |
| Greene | 49,221 | 3.12 × 10−3 | 125.3981 | 410.046 | 747 |
| Orange | 372,813 | 1.06 × 10−3 | 118.2992 | 446.79 | 1853 |
| Putnam | 99,710 | 1.66 × 10−3 | 43.4138 | 157.504 | 349 |
| Rensselaer | 159,429 | 1.44 × 10−3 | 91.6401 | 455.258 | 1560 |
| Rockland | 311,687 | 5.30 × 10−4 | 21.4342 | 71.91 | 548 |
| Schenectady | 154,727 | 1.70 × 10−4 | 26.1887 | 142.528 | 548 |
| Sullivan | 77,547 | 4.62 × 10−4 | 204.5404 | 604.91 | 539 |
| Ulster | 182,493 | 1.73 × 10−3 | 228.011 | 598.846 | 1241 |
| Westchester | 949,113 | 3.06 × 10−4 | 47.8913 | 267.724 | 1150 |
| Formula | Expj | Entj ~ Area | Coefficient | SE | p | λ | pλ | L | R2 | |
|---|---|---|---|---|---|---|---|---|---|---|
 | Const. | Neg. Exp. | 5.47 × 10−2 | 1.01 × 10−2 | 5.70 × 10−8 *** | −2.43 × 10−2 | 9.39 × 10−1 | −10.0 | 0.68 | |
 | Perim. | Neg. Exp. | 2.55 × 10−2 | 5.01 × 10−3 | 3.66 × 10−7 *** | −7.85 × 10−3 | 9.80 × 10−1 | −10.5 | 0.67 | |
 | Area | Neg. Exp. | 8.43 × 10−1 | 1.67 × 10−1 | 4.76 × 10−7 *** | −9.00 × 10−4 | 9.98 × 10−1 | −10.5 | 0.67 | |
 | Perim. | Const. | 1.95 × 10−2 | 1.31 × 10−2 | 1.38 × 10−1 | 3.76 × 10−1 | 5.56 × 10−1 | −15.1 | 0.16 | |
 | Area | Const. | 5.83 × 10−4 | 3.94 × 10−4 | 1.39 × 10−1 | 3.77 × 10−1 | 5.75 × 10−1 | −15.1 | 0.16 | |
 | Const. | Lin. | 1.53 × 10−5 | 2.96 × 10−4 | 9.59 × 10−1 | 6.45 × 10−1 | 1.32 × 10−1 | −15.7 | 0.016 | |
 | Perim. | Lin. | −1.56 × 10−8 | 1.82 × 10−7 | 9.32 × 10−1 | 6.59 × 10−1 | 1.24 × 10−1 | −15.7 | 0.025 | |
 | Area | Lin. | −5.21 × 10−7 | 6.07 × 10−6 | 9.32 × 10−1 | 6.59 × 10−1 | 1.24 × 10−1 | −15.7 | 0.025 | |
| No. patches < 2 ha | 1.35 × 10−1 | 2.62 × 10−2 | 2.46 × 10−7 *** | −1.36 × 10−2 | 9.66 × 10−1 | −10.3 | 0.67 |
| Formula | Coefficient | SE | p | λ | pλ | L |
|---|---|---|---|---|---|---|
| −2.92 × 100 | 8.61 × 10−1 | 6.95 × 10−4 *** | −1.20 × 10−1 | 7.62 × 10−1 | −13.8 | |
| −2.57 × 100 | 3.54 × 100 | 4.67 × 10−1 | 6.85 × 10−1 | 5.34 × 10−2 | −15.5 | |
| 1.25 × 101 | 2.98 × 100 | 2.60 × 10−5 *** | −6.98 × 10−1 | 3.45 × 10−1 | −15.3 | |
| −1.72 × 100 | 1.26 × 100 | 1.73 × 10−1 | 5.41 × 10−1 | 1.94 × 10−1 | −14.9 | |
| −4.25 × 100 | 5.96 × 100 | 4.76 × 10−1 | 6.57 × 10−1 | 6.43 × 10−2 | −15.5 | |
| 7.26 × 100 | 3.53 × 100 | 3.98 × 10−2 * | 6.50 × 10−1 | 6.26 × 10−2 | −13.9 | |
| 1.03 × 10−1 | 3.4 × 10−1 | 7.64 × 10−1 | 6.27 × 10−1 | 1.14 × 10−1 | −15.7 | |
| A | 8.74 × 10−4 | 4.07 × 10−4 | 3.17 × 10−2 * | 1.47 × 10−1 | 7.39 × 10−1 | −14.8 |
| % Forest cover | 3.62 × 103 | 1.57 × 103 | 2.14 × 10−2 * | 4.79 × 10−1 | 2.60 × 10−1 | −13.7 |
| (% Forest cover) 2 | 2.79 × 106 | 1.47 × 106 | 5.79 × 10−1 | 5.05 × 10−1 | 2.33 × 10−1 | −14.3 |
| B | 3.89 × 10−3 | 1.04 × 10−3 | 1.96 × 10−4 *** | −2.94 × 10−1 | 4.40 × 10−1 | −14.0 |
| No. patches | 1.75 × 10−3 | 3.40 × 10−4 | 2.93 × 10−7 *** | −7.70 × 10−1 | 9.47 × 10−2 | −14.2 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
McClure, M.; Diuk-Wasser, M. Reconciling the Entomological Hazard and Disease Risk in the Lyme Disease System. Int. J. Environ. Res. Public Health 2018, 15, 1048. https://doi.org/10.3390/ijerph15051048
McClure M, Diuk-Wasser M. Reconciling the Entomological Hazard and Disease Risk in the Lyme Disease System. International Journal of Environmental Research and Public Health. 2018; 15(5):1048. https://doi.org/10.3390/ijerph15051048
Chicago/Turabian StyleMcClure, Max, and Maria Diuk-Wasser. 2018. "Reconciling the Entomological Hazard and Disease Risk in the Lyme Disease System" International Journal of Environmental Research and Public Health 15, no. 5: 1048. https://doi.org/10.3390/ijerph15051048
APA StyleMcClure, M., & Diuk-Wasser, M. (2018). Reconciling the Entomological Hazard and Disease Risk in the Lyme Disease System. International Journal of Environmental Research and Public Health, 15(5), 1048. https://doi.org/10.3390/ijerph15051048