A Review on Equine Piroplasmosis: Epidemiology, Vector Ecology, Risk Factors, Host Immunity, Diagnosis and Control

 ,
, 
 ,
, 
Abstract
:1. Introduction
2. Historical Perspective
3. Host Range
4. Geographical Distribution
5. Basic Biology
5.1. Morphology
5.2. Life Cycle of Equine Piroplasms
5.2.1. Theileria equi
5.2.2. Babesia caballi
6. Transmission of Equine Piroplasms
7. Immunity in Equine Proplasmosis
8. Clinical Manifestations
8.1. Clinical Signs
8.2. Haematological Changes during Equine Piroplasmosis
8.3. Serum Biochemical Changes
8.4. Gross Pathological Findings
8.5. Histopathological Findings
8.6. Pathophysiology of Anemia in Equine Piroplasmosis
9. Ticks as Vector of Equine Piroplasmosis
10. Risk Factors Associated with Equine Piroplasmosis
10.1. Intrinsic Risk Factors
10.1.1. Equine Species
10.1.2. Age, Gender, and Breeds
10.1.3. Host Activity and Castration Status
10.2. Environmental or Extrinsic Risk Factors
Altitude, Grazing and Climatic Factors
11. Epidemiology of Equine Piroplasmosis
12. Genetic Diversity of Equine Piroplasmosis Parasites
13. Notes on the Detection of Equine Piroplasms in Non-Equine Hosts
14. Diagnosis of Equine Piroplasmosis
14.1. Microscopy and in Vitro Culture
14.2. Serology
14.3. Complement Fixation Test (CFT)
14.4. Indirect Immunofluorescent Antibody Test (IFAT)
14.5. Enzyme-Linked Immunosorbent Assay (ELISA)
14.6. Immunochromatographic Test (ICT)
14.7. DNA Based Diagnostic Techniques
15. Treatment
16. Prevention of Equine Piroplasmosis
17. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Qablan, M.A.; Obornik, M.; Petrželková, K.J.; Sloboda, M.; Shudiefat, M.; Hořín, P.; Modrý, D. Infections by Babesia caballi and Theileria equi in Jordanian equids: Epidemiology and genetic diversity. Parasitology 2013, 140, 1096–1103. [Google Scholar] [CrossRef]
- De Waal, D.T. Equine piroplasmosis: A review. Br. Vet. J. 1992, 148, 6–14. [Google Scholar] [CrossRef]
- Sumbria, D.; Moudgil, A.D.; Singla, L.D. Equine Piroplasmosis: Current status. Veterinaria 2014, 1, 9–14. [Google Scholar]
- Friedhoff, K.T.; Soule, C. An account on equine babesiosis. Rev. Sci. Tech. (Int. Off. Epizoot.) 1996, 15, 1191–1201. [Google Scholar] [CrossRef]
- Rothschild, C.M. Equine piroplasmosis. J. Equine Vet. Sci. 2013, 33, 497–508. [Google Scholar] [CrossRef]
- Knowles, J. Equine Babesiosis (piroplasmosis): A problem in the international movement of horses. Br. Vet. J. 1996, 152, 123–126. [Google Scholar] [CrossRef]
- Roberts, E.D.; Morehouse, L.G.; Griner, J.H.; Mcdaniel, H.A. Equine piroplasmosis. J. Am. Vet. Med. Ass 1962, 141, 1323–1329. [Google Scholar]
- Erbsloh, J.K. Babesiosis in the newborn foal. Supplement 1975, 23, 725–726. [Google Scholar]
- Mehlhorn, H.; Schein, E. Redescription of Babesia equi Laveran, 1901 as Theileria equi. Parasitol. Res. 1998, 84, 467–475. [Google Scholar] [CrossRef]
- Henning, M.W. Animal Diseases in South Africa, 2nd ed.; Central News Agency Ltd.: Cape town, South Africa, 1949; pp. 10–12. [Google Scholar]
- De Waal, D.T.; Potgieter, F.T. The transstadial transmission of Babesia caballi by Rhipicephalus evertsi evertsi. Onderstepoort J. Vet. Res. 1987, 54, 655–666. [Google Scholar] [PubMed]
- Laveran, M. Contribution a l’etude de Piroplasma equi. CR Soc. Biol. 1901, 53, 385–388. [Google Scholar]
- Schein, H. Equine piroplasmosis in South Annam, French Indo-China. Bull. Soc. Exot. Pathol. 1917, 10, 871–873. [Google Scholar]
- Stoute, R.A. Piroplasmosis equine malaria. J. Am. Vet. Med. Asso. 1917, 51, 239. [Google Scholar]
- Theiler, A. Equine malaria and its sequelae. J. Comp. Pathol. Ther. 1903, 16, 97–127. [Google Scholar] [CrossRef]
- Koch, R. Contributions to the history of the development of piroplasma. J. Hyg. Infect. Dis. 1906, 54, 1–9. [Google Scholar]
- Salim, B.; Bakheit, M.A.; Kamau, J.; Sugimoto, C. Current status of equine piroplasmosis in the Sudan. Infect. Genet. Evol. 2013, 16, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Brüning, A. Equine piroplasmosis an update on diagnosis, treatment and prevention. Br. Vet. J. 1996, 152, 139–151. [Google Scholar] [CrossRef]
- Levine, N.D. Veterinary Protozoology, 1st ed.; Iowa State University Press: Ames, IA, USA, 1985. [Google Scholar]
- Scoles, G.A.; Ueti, M.W. Vector ecology of equine piroplasmosis. Annu. Rev. Entomol. 2015, 60, 561–580. [Google Scholar] [CrossRef]
- Malekifard, F.; Tavassoli, M.; Yakhchali, M.; Darvishzadeh, R. Detection of Theileria equi and Babesia caballi using microscopic and molecular methods in horses in suburb of Urmia, Iran. Vet. Res. Forum 2014, 5, 129–133. [Google Scholar]
- Bhoora, R. Molecular Characterization of Babesia caballi and Theileria equi, the Aetiological Agents of Equine Piroplasmosis, in South Africa. Ph.D. Dissertation, University of Pretoria, Pretoria, South Africa, 2009. [Google Scholar]
- Ueti, M.W.; Knowles, D.P. Equine Piroplasmids. In Parasitic Protozoa of Farm Animals and Pets; Florin-Christensen, M., Schnittger, L., Eds.; Springer: Heidelberg, Germany, 2018; pp. 259–269. [Google Scholar]
- Ueti, M.W.; Palmer, G.H.; Scoles, G.A.; Kappmeyer, L.S.; Knowles, D.P. Persistently infected horses are reservoirs for intrastadial tick-borne transmission of the apicomplexan parasite Babesia equi. Infect. Immun. 2008, 76, 3525–3529. [Google Scholar] [CrossRef]
- Short, M.A.; Clark, C.K.; Harvey, J.W.; Wenzlow, N.; Hawkins, I.K.; Allred, D.R.; Kitchen, D.L. Outbreak of equine piroplasmosis in Florida. J. Am. Vet. Med. Assoc. 2012, 240, 588–595. [Google Scholar] [CrossRef]
- Jalovecka, M.; Hajdusek, O.; Sojka, D.; Kopacek, P.; Malandrin, L. The complexity of piroplasms life cycles. Front. Cell. Infect. Microbiol. 2018, 8, 248. [Google Scholar] [CrossRef]
- Holbrook, A.A.; Frerichs, W.M. Equine piroplasmosis. In Proceedings of the Annual Meeting of the United States Animal Health Association, New Orleans, LA, USA, 6–11 October 1968; pp. 220–222. [Google Scholar]
- Allsopp, M.T.E.P.; Lewis, B.D.; Penzhorn, B.L. Molecular evidence for transplacental transmission of Theileria equi from carrier mares to their apparently healthy foals. Vet. Parasitol. 2007, 148, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Sant, C.; d’Abadie, R.; Pargass, I.; Basu, A.K.; Asgarali, Z.; Charles, R.A.; Georges, K.C. Prospective study investigating transplacental transmission of equine piroplasmosis in thoroughbred foals in Trinidad. Vet. Parasitol. 2016, 226, 132–137. [Google Scholar] [CrossRef]
- De Waal, D.T.; van Heerden, J. Equine piroplasmosis. In Infectious Diseases of Livestock; Coetzer, J.A.W., Tustin, R.C., Eds.; Oxford University Press: Cape Town, South Africa, 2004; pp. 425–433. [Google Scholar]
- Françoso, R.; Riccio, A.V.; Fernandes, C.B.; Alonso, M.A.; Belli, C.B. Transplacental transmission of Theileria equi in mules: Should we worry? Vet. Parasitol. 2018, 264, 39–41. [Google Scholar] [CrossRef] [PubMed]
- Wise, L.N.; Kappmeyer, L.S.; Mealey, R.H.; Knowles, D.P. Review of equine piroplasmosis. J. Vet. Intern. Med. 2013, 27, 1334–1346. [Google Scholar] [CrossRef] [PubMed]
- Maurer, F.D. Equine piroplasmosis--another emerging disease. J. Am. Vet. Med. Assoc. 1962, 141, 699. [Google Scholar]
- Ahmed, J. The role of cytokines in immunity and immunopathogenesis of pirolasmoses. Parasitol. Res. 2002, 88, S48–S50. [Google Scholar] [CrossRef]
- Hanafusa, Y.; Cho, K.O.; Kanemaru, T.; Wada, R.; Sugimoto, C.; Onuma, M. Pathogenesis of Babesia caballi infection in experimental horses. J. Vet. Med. Sci. 1998, 60, 1127–1132. [Google Scholar] [CrossRef] [PubMed]
- Ambawat, H.K.; Malhotra, D.V.; Kumar, S.; Dhar, S. Erythrocyte associated haemato-biochemical changes in Babesia equi infection experimentally produced in donkeys. Vet. Parasitol. 1999, 85, 319–324. [Google Scholar] [CrossRef]
- Knowles, D.P.; Kappmeyer, L.S.; Perryman, L.E. Specific immune responses are required to control parasitemia in Babesia equi infection. Infect. Immun. 1994, 62, 1909–1913. [Google Scholar]
- Kumar, S.; Malhotra, D.V.; Dhar, S.; Nichani, A.K. Vaccination of donkeys against Babesia equi using killed merozoite immunogen. Vet. Parasitol. 2002, 106, 19–33. [Google Scholar] [CrossRef]
- Zweygarth, E.; Ahmed, J.S.; Rehbein, G.; Voigt, W.P. Cell-mediated immune response to Babesia equi-transformed lymphoblastoid cells in vitro. Zentralblatt für Bakteriologie Mikrobiologie und Hygiene 1 Abt. Originale. A Medizinische Mikrobiologie Infektionskrankheiten und Parasitologie 1983, 254, 281–289. [Google Scholar] [CrossRef]
- Banerjee, D.P.; Singh, B.; Gautam, O.P.; Sarup, S. Cell-mediated immune response in equine babesiosis. Trop. Anim. Health Prod. 1977, 9, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Cunha, C.W.; McGuire, T.C.; Kappmeyer, L.S.; Hines, S.A.; Lopez, A.M.; Dellagostin, O.A.; Knowles, D.P. Development of specific immunoglobulin Ga (IgGa) and IgGb antibodies correlates with control of parasitemia in Babesia equi infection. Clin. Vaccine Immunol. 2006, 13, 297–300. [Google Scholar] [CrossRef] [PubMed]
- Frerichs, W.M.; Johnson, A.J.; Holbrook, A.A. Equine piroplasmosis: Attempts to infect laboratory animals with Babesia equi. Am. J. Vet. Res. 1969, 30, 1333–1336. [Google Scholar] [PubMed]
- Kouam, M.K.; Kantzoura, V.; Gajadhar, A.A.; Theis, J.H.; Papadopoulos, E.; Theodoropoulos, G. Seroprevalence of equine piroplasms and host-related factors associated with infection in Greece. Vet. Parasitol. 2010, 169, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Laus, F.; Spaterna, A.; Faillace, V.; Veronesi, F.; Ravagnan, S.; Beribé, F.; Tesei, B. Clinical investigation on Theileria equi and Babesia caballi infections in Italian donkeys. BMC Vet. Res. 2015, 11, 100. [Google Scholar] [CrossRef]
- Mahoney, D.F.; Wright, I.G.; Frerichs, W.M.; Groenendyk, S.; O’Sullivan, B.M.; Roberts, M.C.; Waddell, A.H. The identification of Babesia equi in Australia. Aust. Vet. J. 1977, 53, 461–464. [Google Scholar] [CrossRef]
- Osman, S.A. Clinical, haematological and therapeutic studies on babesiosis in Arabian horses in the Qassim region, central of Saudi Arabia. J. Appl. Anim. Res. 2017, 45, 118–121. [Google Scholar] [CrossRef]
- Zobba, R.; Ardu, M.; Niccolin, S.; Chessa, B.; Manna, L.; Cocco, R.; Parpaglia, M.L.P. Clinical and laboratory findings in equine piroplasmosis. J. Equine Vet. Sci. 2008, 28, 301–308. [Google Scholar] [CrossRef]
- Mahmoud, M.S.; El-Ezz, N.T.A.; Abdel-Shafy, S.; Nassar, S.A.; El Namaky, A.H.; Khalil, W.K.; Suarez, C.E. Assessment of Theileria equi and Babesia caballi infections in equine populations in Egypt by molecular, serological and hematological approaches. Parasites Vectors 2016, 9, 260. [Google Scholar] [CrossRef] [PubMed]
- Hailat, N.Q.; Lafi, S.Q.; Al-Darraji, A.M.; Al-Ani, F.K. Equine babesiosis associated with strenuous exercise: Clinical and pathological studies in Jordan. Vet. Parasitol. 1997, 69, 1–8. [Google Scholar] [CrossRef]
- Camacho, A.T.; Guitian, F.J.; Pallas, E.; Gestal, J.J.; Olmeda, A.S.; Habela, M.A.; Spielman, A. Theilera (Babesia) equi and Babesia caballi infections in horses in Galicia, Spain. Trop. Anim. Health Prod. 2005, 37, 293–302. [Google Scholar] [CrossRef]
- Allen, P.C.; Frerichs, W.M.; Holbrook, A.A. Experimental acute Babesia caballi infections: II. Response of platelets and fibrinogen. Exp. Parasitol. 1975, 37, 373–379. [Google Scholar] [CrossRef]
- Walker, J.B.; Keirans, J.E.; Horak, I. The genus Rhipicephalus (Acari, Ixodidae). In A Guide to the Brown Ticks of the World; Cambridge University Press: New York, NY, USA, 2005. [Google Scholar]
- Stiller, D.; Goff, W.L.; Johnson, L.W.; Knowles, D.P. Dermacentor variabilis and Boophilus microplus (Acari: Ixodidae): Experimental vectors of Babesia equi to equids. J. Med. Entomol. 2002, 39, 667–670. [Google Scholar] [CrossRef] [PubMed]
- Kerber, C.E.; Labruna, M.B.; Ferreira, F.; De Waal, D.T.; Knowles, D.P.; Gennari, S.M. Prevalence of equine piroplasmosis and its association with tick infestation in the State of São Paulo, Brazil. Rev. Bras. Parasitol. Veterinária 2009, 18, 1–8. [Google Scholar] [CrossRef]
- Scoles, G.A.; Hutcheson, H.J.; Schlater, J.L.; Hennager, S.G.; Pelzel, A.M.; Knowles, D.P. Equine piroplasmosis associated with Amblyomma cajennense ticks, Texas, USA. Emerg. Infect. Dis. 2011, 17, 1903. [Google Scholar] [CrossRef] [PubMed]
- Abedi, V.; Razmi, G.; Seifi, H.; Naghibi, A. Molecular and serological detection of Theileria equi and Babesia caballi infection in horses and ixodid ticks in Iran. Ticks Tick-Borne Dis. 2014, 5, 239–244. [Google Scholar] [CrossRef]
- Ferrolho, J.; Antunes, S.; Santos, A.S.; Velez, R.; Padre, L.; Cabezas-Cruz, A.; Domingos, A. Detection and phylogenetic characterization of Theileria spp., and Anaplasma marginale in Rhipicephalus bursa in Portugal. Ticks Tick-Borne Dis. 2016, 7, 443–448. [Google Scholar] [CrossRef]
- Scoles, G.A.; Ueti, M.W. Amblyomma cajennense is an intrastadial biological vector of Theileria equi. Parasites Vectors 2013, 6, 306. [Google Scholar] [CrossRef]
- Peckle, M.; Pires, M.S.; dos Santos, T.M.; Roier, E.C.; da Silva, C.B.; Vilela, J.A.; Massard, C.L. Molecular epidemiology of Theileria equi in horses and their association with possible tick vectors in the state of Rio de Janeiro, Brazil. Parasitol. Res. 2013, 112, 2017–2025. [Google Scholar] [CrossRef]
- Duell, J.R.; Carmichael, R.; Herrin, B.H.; Holbrook, T.C.; Talley, J.; Little, S.E. Prevalence and species of ticks on horses in Central Oklahoma. J. Med. Entomol. 2013, 50, 1330–1333. [Google Scholar] [CrossRef]
- De Waal, D.T. The transovarial transmission of Babesia caballi by Hyalomma truncatum. Onderstepoort J. Vet. Res. 1990, 57, 99–100. [Google Scholar] [PubMed]
- Balkaya, I.; Utuki, A.E.; Piskin, F.C. Prevalence of Theileria equi and B. caballi in donkeys from eastern Turkey in winter season. Pak. Vet. J. 2010, 30, 245–246. [Google Scholar]
- Guidi, E.; Pradier, S.; Lebert, I.; Leblond, A. Piroplasmosis in an endemic area: Analysis of the risk factors and their implications in the control of theileriosis and babesiosis in horses. Parasitol. Res. 2015, 114, 71–83. [Google Scholar] [CrossRef]
- Santos, T.M.D.; Roier, E.C.R.; Santos, H.A.; Pires, M.S.; Vilela, J.A.R.; Moraes, L.M.D.B.; Massard, C.L. Factors associated to Theileria equi in equids of two microregions from Rio de Janeiro, Brazil. Braz. J. Vet. Parasitol. 2011, 20, 235–241. [Google Scholar] [CrossRef]
- García-Bocanegra, I.; Arenas-Montes, A.; Hernández, E.; Adaszek, Ł.; Carbonero, A.; Almería, S.; Arenas, A. Seroprevalence and risk factors associated with Babesia caballi and Theileria equi infection in equids. Vet. J. 2013, 195, 172–178. [Google Scholar] [CrossRef]
- Rüegg, S.R.; Torgerson, P.; Deplazes, P.; Mathis, A. Age-dependent dynamics of Theileria equi and Babesia caballi infections in southwest Mongolia based on IFAT and/or PCR prevalence data from domestic horses and ticks. Parasitology 2007, 134, 939–947. [Google Scholar]
- Moretti, A.; Mangili, V.; Salvatori, R.; Maresca, C.; Scoccia, E.; Torina, A.; Pietrobelli, M. Prevalence and diagnosis of Babesia and Theileria infections in horses in Italy: A preliminary study. Vet. J. 2010, 184, 346–350. [Google Scholar] [CrossRef]
- Roberts, C.W.; Walker, W.; Alexander, J. Sex-associated hormones and immunity to protozoan parasites. Clin. Microbiol. Rev. 2001, 14, 476–488. [Google Scholar] [CrossRef] [PubMed]
- Grandi, G.; Molinari, G.; Tittarelli, M.; Sassera, D.; Kramer, L.H. Prevalence of Theileria equi and Babesia caballi infection in horses from northern Italy. Vector-Borne Zoonotic Dis. 2011, 11, 955–956. [Google Scholar] [CrossRef]
- Bahrami, S.; Ghadrdan, A.R.; Pourmahdi, B.M.; Vafayi, S.M. Epidemiology of Theileria equi in Persian Arab horses from Iran. Vet. Med. 2014, 59, 409–414. [Google Scholar] [CrossRef]
- Shkap, V.; Cohen, I.; Leibovitz, B.; Pipano, E.; Avni, G.; Shofer, S.; Knowles, D. Seroprevalence of Babesia equi among horses in Israel using competitive inhibition ELISA and IFA assays. Vet. Parasitol. 1998, 76, 251–259. [Google Scholar] [CrossRef]
- Hughes, V.L.; Randolph, S.E. Testosterone increases the transmission potential of tick-borne parasites. Parasitology 2001, 123, 365–371. [Google Scholar] [CrossRef]
- Hughes, V.L.; Randolph, S.E. Testosterone depresses innate and acquired resistance to ticks in natural rodent hosts: A force for aggregated distributions of parasites. J. Parasitol. 2001, 87, 49–54. [Google Scholar] [CrossRef]
- Randolph, S.E.; Nuttall, P.A. Nearly right or precisely wrong? Natural versus laboratory studies of vector-borne diseases. Parasitol. Today 1994, 10, 458–462. [Google Scholar] [CrossRef]
- Byambaa, B.; Dash, M.; Tarasevic, I.W. New Research about Tick-Borne Rickettsiosis; University of Ulaanbaatar: Ulaanbaatar, Mongolia, 1994. (In Mongolian) [Google Scholar]
- Abutarbush, S.M.; Alqawasmeh, D.M.; Mukbel, R.M.; Al-Majali, A.M. Equine babesiosis: Seroprevalence, risk factors and comparison of different diagnostic methods in Jordan. Transbound. Emerg. Dis 2012, 59, 72–78. [Google Scholar] [CrossRef]
- Salim, B.O.M.; Hassan, S.M.; Bakheit, M.A.; Alhassan, A.; Igarashi, I.; Karanis, P.; Abdelrahman, M.B. Diagnosis of Babesia caballi and Theileria equi infections in horses in Sudan using ELISA and PCR. Parasitol. Res. 2008, 103, 1145. [Google Scholar] [CrossRef]
- Butler, C.M.; van Oldruitenborgh-Oosterbaan, M.M.S.; Stout, T.A.; van der Kolk, J.H.; Van den Wollenberg, L.; Nielen, M.; Houwers, D.J. Prevalence of the causative agents of equine piroplasmosis in the South West of The Netherlands and the identification of two autochthonous clinical Theileria equi infections. Vet. J. 2012, 193, 381–385. [Google Scholar] [CrossRef]
- Del Pino, L.E.B.; Roberto, N.; Vincenzo, V.; Francesca, I.; Antonella, C.; Luca, A.G.; Teresa, S.M. Babesia caballi and Theileria equi infections in horses in Central-Southern Italy: Sero-molecular survey and associated risk factors. Ticks Tick-Borne Dis. 2016, 7, 462–469. [Google Scholar] [CrossRef]
- Motloang, M.Y.; Thekisoe, O.M.M.; Alhassan, A.; Bakheit, M.; Motheo, M.P.; Masangane, F.E.S.; Mbati, P.A. Prevalence of Theileria equi and Babesia caballi infections in horses belonging to resource-poor farmers in the north-eastern Free State Province, South Africa. Onderstepoort J. Vet. Res. 2008, 75, 141–146. [Google Scholar] [CrossRef]
- Sumbria, D.; Singla, L.D.; Sharma, A. Theileria equi and Babesia caballi infection of equids in Punjab, India: A serological and molecular survey. Trop. Anim. Health Prod. 2016, 48, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Cantú-Martínez, M.A.; Segura-Correa, J.C.; Silva-Páez, M.L.; Avalos-Ramírez, R.; Wagner, G.G. Prevalence of antibodies to Theileria equi and Babesia caballi in horses from northeastern Mexico. J. Parasitol. 2012, 98, 869–870. [Google Scholar] [CrossRef] [PubMed]
- Posada-Guzmán, M.F.; Dolz, G.; Romero-Zúñiga, J.J.; Jiménez-Rocha, A.E. Detection of Babesia caballi and Theileria equi in blood from equines from four indigenous communities in Costa Rica. Vet. Med. Int. 2015, 2015, 236278. [Google Scholar] [CrossRef]
- Nugraha, A.B.; Cahyaningsih, U.; Amrozi, A.; Ridwan, Y.; Agungpriyono, S.; Taher, D.M.; Sivakumar, T. Serological and molecular prevalence of equine piroplasmosis in Western Java, Indonesia. Vet. Parasitol. Reg. Stud. Rep. 2018, 14, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Turaki, U.A.; Kumsha, H.A.; Biu, A.A.; Bokko, P.B. Prevalence of P7roplasmosis amongst local horses in Northeastern Nigeria. J. Agric. Vet. Sci. 2014, 7, 4–7. [Google Scholar]
- Sanusi, M.; Ahmed, I.A.; Tahir, I.; Mai, H.M.; Kalla, D.J.U.; Shuaibu, I. Survey of equine piroplasmosis in the Savanna areas, Bauchi state, North-eastern Nigeria. Ippologia Cremona 2014, 25, 3–8. [Google Scholar]
- Mshelia, W.P.; Sambo, K.W.; Adamu, S.; Edeh, E.R.; Onoja, I.I. Persistence of equine piroplasmosis in horses in Nigeria. J. Equine Vet. Sci. 2016, 39, S104–S105. [Google Scholar] [CrossRef] [Green Version]
- Al-Obaidi, Q.T.; Arshad, M.; Al-Sultan, I.I.; Azlinda, A.; Mohd-Azam, K.G.K. Comparison between microscopic examination and competitive ELISA for diagnosis of equine piroplasmosis in Kelantan, Malaysia. Malays. J. Vet. Res. 2016, 7, 23–29. [Google Scholar]
- Guven, E.; Avcioglu, H.; Deniz, A.; Balkaya, I.; Abay, U.; Yavuz, Ş.; Akyüz, M. Prevalence and molecular characterization of Theileria equi and Babesia caballi in jereed horses in Erzurum, Turkey. Acta Parasitol. 2017, 62, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Jaffer, O.; Abdishakur, F.; Hakimuddin, F.; Riya, A.; Wernery, U.; Schuster, R.K. A comparative study of serological tests and PCR for the diagnosis of equine piroplasmosis. Parasitol. Res. 2010, 106, 709–713. [Google Scholar] [CrossRef] [PubMed]
- Alanazi, A.D.; Alyousif, M.S.; Hassieb, M.M. Seroprevalence study on Theileria equi and Babesia caballi antibodies in horses from central province of Saudi Arabia. J. Parasitol. 2012, 98, 1015–1017. [Google Scholar] [CrossRef]
- Kamyingkird, K.; Yangtara, S.; Desquesnes, M.; Cao, S.; Adjou, P.K.; Jittapalapong, S.; Igarashi, I. Seroprevalence of Babesia caballi and Theileria equi in horses and mules from Northern Thailand. J. Protozool. Res. 2014, 24, 11–17. [Google Scholar]
- Sigg, L.; Gerber, V.; Gottstein, B.; Doher, M.G.; Frey, C.F. Seroprevalence of Babesia caballi and Theileria equi in the Swiss horse population. Parasitol. Int. 2012, 59, 313–317. [Google Scholar] [CrossRef] [PubMed]
- Farkas, R.; Tánczos, B.; Gyurkovszky, M.; Földvári, G.; Solymosi, N.; Edelhofer, R.; Hornok, S. Serological and molecular detection of Theileria equi infection in horses in Hungary. Vet. Parasitol. 2013, 192, 143–148. [Google Scholar] [CrossRef]
- Cortés, M.G.M.; Fernández-García, J.L.; Martínez-Estéllez, M.Á.H. Seroprevalence of Theileria equi and Babesia caballi in horses in Spain. Parasite 2017, 24, 14. [Google Scholar] [CrossRef]
- Ros-García, A.; M’ghirbi, Y.; Hurtado, A.; Bouattour, A. Prevalence and genetic diversity of piroplasm species in horses and ticks from Tunisia. Infect. Genet. Evol. 2013, 17, 33–37. [Google Scholar] [CrossRef]
- Munkhjargal, T.; Sivakumar, T.; Battsetseg, B.; Nyamjargal, T.; Aboulaila, M.; Purevtseren, B.; Igarashi, I. Prevalence and genetic diversity of equine piroplasms in Tov province, Mongolia. Infect. Genet. Evol. 2013, 16, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.G.; Yun, S.H.; Choi, S.K.; Cho, G.J.; Park, Y.S.; Cho, K.H.; Kwak, D. Molecular and phylogenetic analysis of equine piroplasms in the Republic of Korea. Res. Vet. Sci. 2013, 94, 579–583. [Google Scholar] [CrossRef]
- Díaz-Sánchez, A.A.; Pires, M.S.; Estrada, C.Y.; Cañizares, E.V.; del Castillo Domínguez, S.L.; Cabezas-Cruz, A.; Corona-González, B. First molecular evidence of Babesia caballi and Theileria equi infections in horses in Cuba. Parasitol. Res. 2018, 117, 3109–3118. [Google Scholar] [CrossRef] [PubMed]
- Rosales, R.; Rangel-Rivas, A.; Escalona, A.; Jordan, L.S.; Gonzatti, M.I.; Aso, P.M.; Mijares, A. Detection of Theileria equi and Babesia caballi infections in Venezuelan horses using Competitive-Inhibition ELISA and PCR. Vet. Parasitol. 2013, 196, 37–43. [Google Scholar] [CrossRef]
- Braga, M.D.S.C.D.; Costa, F.N.; Gomes, D.R.M.; Xavier, D.R.; André, M.R.; Gonçalves, L.R.; Machado, R.Z. Genetic diversity of piroplasmids species in equids from island of São Luís, northeastern Brazil. Rev. Bras. Parasitol. Veterinária 2017, 26, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Gallusová, M.; Qablan, M.A.; D’Amico, G.; Oborník, M.; Petrželková, K.J.; Mihalca, A.D.; Modrý, D. Piroplasms in feral and domestic equines in rural areas of the Danube Delta, Romania, with survey of dogs as a possible reservoir. Vet. Parasitol. 2014, 206, 287–292. [Google Scholar] [CrossRef]
- Seo, M.G.; Yun, S.H.; Choi, S.K.; Cho, G.J.; Park, Y.S.; Kwon, O.D.; Kwon, Y.S. Seroprevalence of equine piroplasms in the Republic of Korea. Vet. Parasitol 2011, 179, 224–226. [Google Scholar] [CrossRef]
- Wang, M.; Guo, W.; Igarashi, I.; Xuan, X.; Wang, X.; Xiang, W.; Jia, H. Epidemiological investigation of equine piroplasmosis in China by enzyme-linked immunosorbent assays. J. Vet. Med. Sci. 2014, 76, 549–552. [Google Scholar] [CrossRef] [PubMed]
- Gizachew, A.; Schuster, R.K.; Joseph, S.; Werner, R.; Georgy, N.A.; Elizabeth, S.K.; Wernery, U. Piroplasmosis in Donkeys—A Hematological and Serological Study in Central Ethiopia. J. Equine Vet. Sci. 2013, 33, 18–21. [Google Scholar] [CrossRef]
- Oduori, D.O.; Onyango, S.C.; Kimari, J.N.; MacLeod, E. A field survey for the seroprevalence of Theileria equi and Babesia caballi in donkeys from Nuu Division, Kenya. Ticks Tick-Borne Dis. 2015, 6, 683–688. [Google Scholar] [CrossRef]
- Machado, R.Z.; Toledo, C.Z.P.; Teixeira, M.C.A.; André, M.R.; Freschi, C.R.; Sampaio, P.H. Molecular and serological detection of Theileria equi and Babesia caballi in donkeys (Equus asinus) in Brazil. Vet. Parasitol. 2012, 186, 461–465. [Google Scholar] [CrossRef]
- Manna, G.; Cersini, A.; Nardini, R.; Del Pino, L.E.B.; Antognetti, V.; Zini, M.; Scicluna, M.T. Genetic diversity of Theileria equi and Babesia caballi infecting horses of Central-Southern Italy and preliminary results of its correlation with clinical and serological status. Ticks Tick-Borne Dis. 2018, 9, 1212–1220. [Google Scholar] [CrossRef] [PubMed]
- Alhassan, A.; Thekisoe, O.M.; Yokoyama, N.; Inoue, N.; Motloang, M.; Mbati, P.A.; Igarashi, I. Development of loop-mediated isothermal amplification (LAMP) method for diagnosis of equine piroplasmosis. Vet. Parasitol. 2007, 143, 155–160. [Google Scholar] [CrossRef]
- Battsetseg, B.; Lucero, S.; Xuan, X.; Claveria, F.G.; Inoue, N.; Alhassan, A.; Fujisaki, K. Detection of natural infection of Boophilus microplus with Babesia equi and Babesia caballi in Brazilian horses using nested polymerase chain reaction. Vet. Parasitol. 2002, 107, 351–357. [Google Scholar] [CrossRef]
- Cacciò, S.; Cammà, C.; Onuma, M.; Severini, C. The β-tubulin gene of Babesia and Theileria parasites is an informative marker for species discrimination. Int. J. Parasitol. 2000, 30, 1181–1185. [Google Scholar] [CrossRef]
- Katzer, F.; McKellar, S.; Kirvar, E.; Shiels, B. Phylogenetic analysis of Theileria and Babesia equi in relation to the establishment of parasite populations within novel host species and the development of diagnostic tests. Mol. Biochem. Parasitol. 1998, 95, 33–44. [Google Scholar] [CrossRef]
- Bhoora, R.; Buss, P.; Guthrie, A.J.; Penzhorn, B.L.; Collins, N. Genetic diversity of piroplasms in Plains zebra (Equus quagga burchellii) and Cape mountain zebra (Equus zebra zebra) in South Africa. Vet. Parasitol. 2010, 174, 145–149. [Google Scholar] [CrossRef]
- Salim, B.; Bakheit, M.A.; Kamau, J.; Nakamura, I.; Sugimoto, C. Nucleotide sequence heterogeneity in the small subunit ribosomal RNA gene within Theileria equi from horses in Sudan. Parasitol. Res. 2010, 106, 493. [Google Scholar] [CrossRef] [PubMed]
- Kouam, M.K.; Kantzoura, V.; Masuoka, P.M.; Gajadhar, A.A.; Theodoropoulos, G. Genetic diversity of equine piroplasms in Greece with a note on speciation within Theileria genotypes (T. equi and T. equi-like). Infect. Genet. Evol. 2010, 10, 963–968. [Google Scholar] [CrossRef] [PubMed]
- Veronesi, F.; Morganti, G.; Ravagnan, S.; Laus, F.; Spaterna, A.; Diaferia, M.; Capelli, G. Molecular and serological detection of tick-borne pathogens in donkeys (Equus asinus) in Italy. Vet. Microbiol. 2014, 173, 348–354. [Google Scholar] [CrossRef] [PubMed]
- Nagore, D.; Garcıa-Sanmartın, J.; Garcıa-Pérez, A.L.; Juste, R.A.; Hurtado, A. Detection and identification of equine Theileria and Babesia species by reverse line blotting: Epidemiological survey and phylogenetic analysis. Vet. Parasitol. 2004, 123, 41–54. [Google Scholar] [CrossRef]
- Hall, C.M.; Busch, J.D.; Scoles, G.A.; Palma-Cagle, K.A.; Ueti, M.W.; Kappmeyer, L.S.; Wagner, D.M. Genetic characterization of Theileria equi infecting horses in North America: Evidence for a limited source of US introductions. Parasites Vectors 2013, 6, 35. [Google Scholar] [CrossRef]
- Liu, Q.; Meli, M.L.; Zhang, Y.; Meili, T.; Stirn, M.; Riond, B.; Hofmann-Lehmann, R. Sequence heterogeneity in the 18S rRNA gene in Theileria equi from horses presented in Switzerland. Vet. Parasitol. 2016, 221, 24–29. [Google Scholar] [CrossRef]
- Criado-Fornelio, A.; Gónzalez-del-Rıo, M.A.; Buling-Sarana, A.; Barba-Carretero, J.C. The “expanding universe” of piroplasms. Vet. Parasitol. 2004, 119, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Uilenberg, G. Babesia—A historical overview. Vet. Parasitol. 2006, 138, 3–10. [Google Scholar] [CrossRef]
- Fritz, D. A PCR study of piroplasms in 166 dogs and 111 horses in France (March 2006 to March 2008). Parasitol. Res. 2010, 106, 1339–1342. [Google Scholar] [CrossRef]
- Qablan, M.A.; Sloboda, M.; Jirků, M.; Oborník, M.; Dwairi, S.; Amr, Z.S.; Modrý, D. Quest for the piroplasms in camels: Identification of Theileria equi and Babesia caballi in Jordanian dromedaries by PCR. Vet. Parasitol. 2012, 186, 456–460. [Google Scholar] [CrossRef] [PubMed]
- Sloboda, M.; Jirků, M.; Lukešová, D.; Qablan, M.; Batsukh, Z.; Fiala, I.; Lukeš, J. A survey for piroplasmids in horses and Bactrian camels in North-Eastern Mongolia. Vet. Parasitol. 2011, 179, 246–249. [Google Scholar] [CrossRef] [PubMed]
- de Sousa, K.C.M.; Fernandes, M.P.; Herrera, H.M.; Freschi, C.R.; Machado, R.Z.; André, M.R. Diversity of piroplasmids among wild and domestic mammals and ectoparasites in Pantanal wetland, Brazil. Ticks Tick-Borne Dis. 2018, 9, 245–253. [Google Scholar] [CrossRef]
- Inácio, E.L.; Pérez-Macchi, S.; Alabi, A.; Bittencourt, P.; Müller, A. Prevalence and molecular characterization of piroplasmids in domestic dogs from Paraguay. Ticks Tick-Borne Dis. 2019, 10, 321–327. [Google Scholar]
- Jasim, H.J.; Azzal, G.Y.; Othman, R.M. Conventional and molecular detection of Babesia caballi and Theileria equi parasites in infected camels in south of Iraq. Basrah J. Vet. Res. 2015, 14, 110–121. [Google Scholar]
- Da Silveira, A.W.; De Oliveira, G.G.; Menezes Santos, L.; da Silva Azuaga, L.B.; Macedo Coutinho, C.R.; Echeverria, J.T.; Izabel de Souza, A. Natural Infection of the South American Tapir (Tapirus terrestris) by Theileria equi. J. Wildl. Dis. 2017, 53, 411–413. [Google Scholar] [CrossRef] [PubMed]
- Beck, R.; Vojta, L.; Mrljak, V.; Marinculić, A.; Beck, A.; Živičnjak, T.; Cacciò, S.M. Diversity of Babesia and Theileria species in symptomatic and asymptomatic dogs in Croatia. Int. J. Parasitol. 2009, 39, 843–848. [Google Scholar] [CrossRef] [PubMed]
- Alhassan, A.; Govind, Y.; Tam, N.T.; Thekisoe, O.M.; Yokoyama, N.; Inoue, N.; Igarashi, I. Comparative evaluation of the sensitivity of LAMP, PCR and in vitro culture methods for the diagnosis of equine piroplasmosis. Parasitol. Res. 2007, 100, 1165–1168. [Google Scholar] [CrossRef]
- Zweygarth, E.; Just, M.C.; De Waal, D.T. In vitro cultivation of Babesia equi: Detection of carrier animals and isolation of parasites. Onderstepoort J. Vet. Res. 1997, 64, 51–56. [Google Scholar]
- Ybañez, A.P.; Ybañez, R.H.D.; Talle, M.G.; Arreglo, R.M.T.; Geens, M.J.C.; Villas, J.G.I., III; Laruga, C.L.; Cao, S.; Moumouni, F.P.A.; Liu, M.; et al. Serological and molecular detection of Theileria equi and Babesia caballi in Philippine horses. Ticks Tick-Borne Dis. 2018, 9, 1125–1128. [Google Scholar]
- Ali, S.; Sugimoto, C.; Onuma, M. Equine piroplasmosis. J. Equine Sci. 2001, 7, 67–77. [Google Scholar] [CrossRef]
- Weiland, G. Species-specific serodiagnosis of equine piroplasma infections by means of complement fixation test (CFT), immunofluorescence (IIF), and enzyme-linked immunosorbent assay (ELISA). Vet. Parasitol. 1986, 20, 43–48. [Google Scholar] [CrossRef]
- Knowles, D.P. Control of Babesia equi parasitemia. Parasitol. Today 1996, 12, 195–198. [Google Scholar] [CrossRef]
- Knowles, D.P.; Kappmeyer, L.S.; Stiller, D.; Hennager, S.G.; Perryman, L.E. Antibody to a recombinant merozoite protein epitope identifies horses infected with Babesia equi. J. Clin. Microbiol. 1992, 30, 3122–3126. [Google Scholar] [PubMed]
- Kappmeyer, L.S.; Perryman, L.E.; Hines, S.A.; Baszler, T.V.; Katz, J.B.; Hennager, S.G.; Knowles, D.P. Detection of equine antibodies to Babesia caballi by recombinant B. caballi rhoptry-associated protein 1 in a competitive-inhibition enzyme-linked immunosorbent assay. J. Clin. Microbiol. 1999, 37, 2285–2290. [Google Scholar]
- El-Moamly, A.A. Immuno-chromatographic Techniques: Benefits for the Diagnosis of Parasitic Infections. Austin Chromatogr 2014, 1, 1–8. [Google Scholar]
- Notomi, T.; Mori, Y.; Tomita, N.; Kanda, H. Loop-mediated isothermal amplification (LAMP): Principle, features, and future prospects. J. Microbiol. 2015, 53, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Ueti, M.W.; Mealey, R.H.; Kappmeyer, L.S.; White, S.N.; Kumpula-McWhirter, N.; Pelzel, A.M.; Hendrickson, A. Re-emergence of the apicomplexan Theileria equi in the United States: Elimination of persistent infection and transmission risk. PLoS ONE 2012, 7, e44713. [Google Scholar] [CrossRef] [PubMed]
- Frerichs, W.M.; Holbrook, A.A. Treatment of equine piroplasmosis (B. caballi) with imidocarb dipropionate. Vet. Rec. 1974, 95, 188–189. [Google Scholar] [CrossRef] [PubMed]
- Knowles, R.C.; Hourrigan, J.L.; Holbrook, A.A. Equine piroplasmosis. Equine Pr. 1980, 2, 10–14. [Google Scholar]
- Bork, S.; Yokoyama, N.; Matsuo, T.; Claveria, F.G.; Fujisaki, K.; Igarashi, I. Clotrimazole, ketoconazole, and clodinafop-propargyl as potent growth inhibitors of equine Babesia parasites during in vitro culture. J. Parasitol. 2003, 89, 604–606. [Google Scholar] [CrossRef]
- Nagai, A.; Yokoyama, N.; Matsuo, T.; Bork, S.; Hirata, H.; Xuan, X.; Igarashi, I. Growth-inhibitory effects of artesunate, pyrimethamine, and pamaquine against Babesia equi and Babesia caballi in in vitro cultures. Antimicrob. Agents Chemother. 2003, 47, 800–803. [Google Scholar] [CrossRef] [PubMed]
- Tayebwa, D.S.; Tuvshintulga, B.; Guswanto, A.; Nugraha, A.B.; Batiha, G.E.S.; Gantuya, S.; Igarashi, I. The effects of nitidine chloride and camptothecin on the growth of Babesia and Theileria parasites. Ticks Tick-Borne Dis. 2018, 9, 1192–1201. [Google Scholar] [CrossRef] [PubMed]
- Jongejan, F.; Uilenberg, G. The global importance of ticks. Parasitology 2004, 129, S3–S14. [Google Scholar] [CrossRef] [PubMed]
- Dantas-Torres, F.; Chomel, B.B.; Otranto, D. Ticks and tick-borne diseases: A One Health perspective. Trends Parasitol. 2012, 28, 437–446. [Google Scholar] [CrossRef]
- Patz, J.A.; Graczyk, T.K.; Geller, N.; Vittor, A.Y. Effects of environmental change on emerging parasitic diseases. Int. J. Parasitol. 2000, 30, 1395–1405. [Google Scholar] [CrossRef] [Green Version]






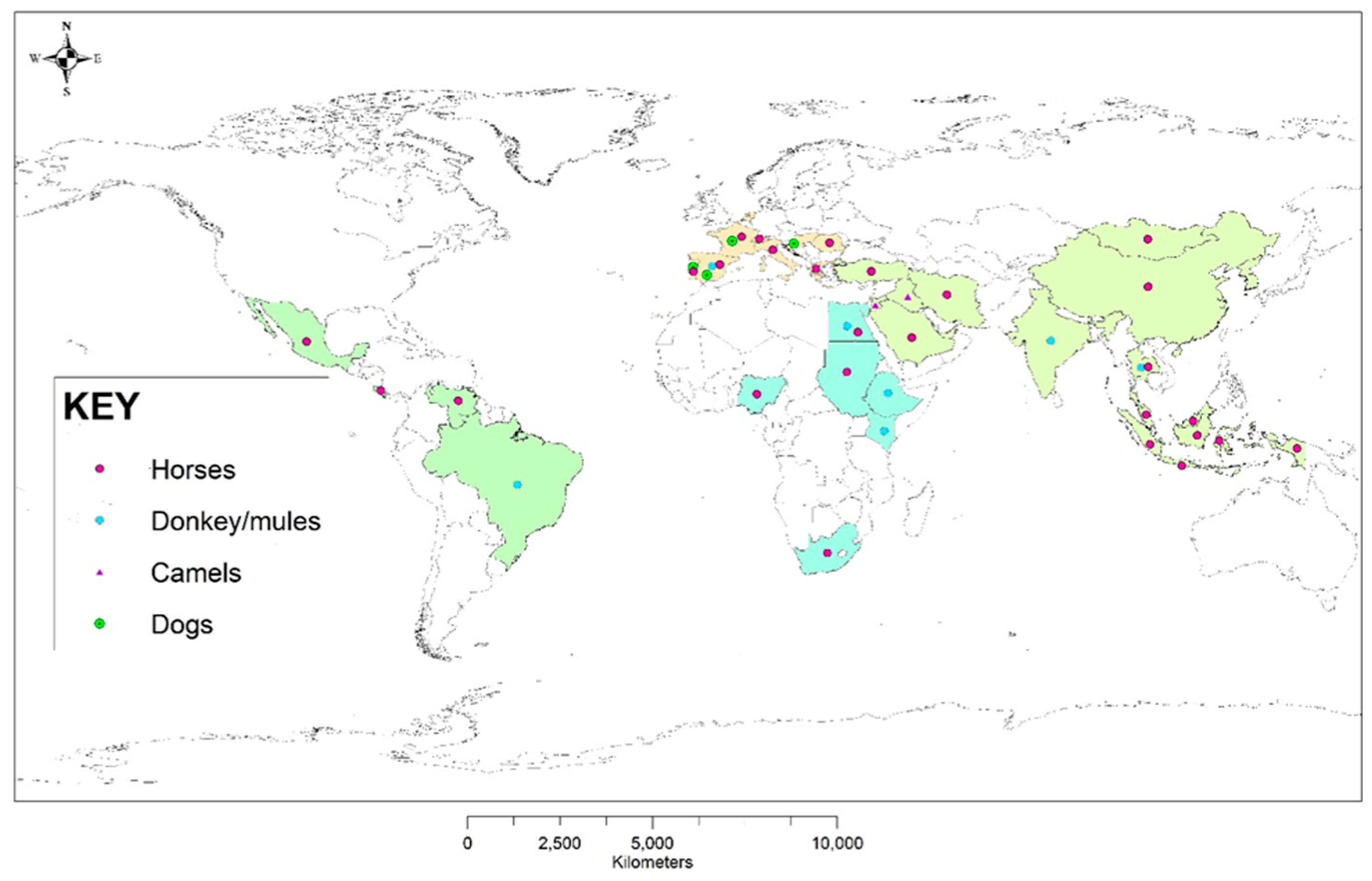{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diagnostic Technique | Year | Continent | Country | Sample Size | Prevalence (%) (B. caballi & T. equi) | Reference |
|---|---|---|---|---|---|---|
| Microscopy | 2008 | Africa | South Africa | 129 | 0 and 3.0 | [80] |
| 2014 | Africa | Nigeria | 240 | 6.0 and 94.0 | [85] | |
| 2014 | Africa | Nigeria | 400 | 83.3 and 11.1 | [86] | |
| 2016 | Africa | Nigeria | 156 | 2.7 and 10.3 | [87] | |
| 2014 | Asia | Iran | 100 | 0 and 5.0 | [56] | |
| 2016 | Asia | Malaysia | 306 | 22.2 and 16.9 | [88] | |
| 2017 | Asia | Turkey | 27 | 0 and 4.8 | [89] | |
| IFAT | 2008 | Africa | South Africa | 99 | 51.5 and 97.9 | [80] |
| 2016 | Africa | Egypt | 88 | 17.0 and 23.9 | [48] | |
| 2010 | Asia | United Arab Emirates | 105 | 10.5 and 33.3 | [90] | |
| 2012 | Asia | Saudi Arabia | 241 | 7.5 and 10.4 | [91] | |
| 2014 | Asia | Iran | 100 | 2.0 and 48.0 | [56] | |
| 2014 | Asia | Thailand | 63 | 11.1 and 3.2 | [92] | |
| 2012 | North America | Mexico | 248 | 27.4 and 45.2 | [82] | |
| 2010 | Europe | Switzerland | 689 | 1.5 and 4.4 | [93] | |
| 2012 | Europe | Netherlands | 300 | 3 and 1 | [78] | |
| 2013 | Europe | Hungry | 324 | NA and 31.8 | [94] | |
| 2017 | Europe | Spain | 3100 | 21.0 and 44.0 | [95] | |
| PCR | 2008 | Africa | Sudan | 131 | 0 and 25.2 | [77] |
| 2008 | Africa | South Africa | 99 | 0 and 12.1 | [80] | |
| 2013 | Africa | Tunisia | 104 | 0.9 and 11.5 | [96] | |
| 2016 | Africa | Egypt | 88 | 19.3 and 36.4 | [48] | |
| 2013 | Asia | Mongolia | 250 | 42.4 and 6.4 | [97] | |
| 2013 | Asia | Jordan | 288 | 7.3 and 18.8 | [1] | |
| 2013 | Asia | Korea | 224 | 0 and 0.9 | [98] | |
| 2014 | Asia | Thailand | 63 | 0 and 0 | [92] | |
| 2014 | Asia | Iran | 100 | 0 and 45.0 | [56] | |
| 2016 | Asia | India | 160 | 0 and 11.9 | [81] | |
| 2017 | Asia | Turkey | 125 | 0 and 8.8 | [89] | |
| 2018 | Asia | Indonesia | 235 | 2.1 and 6.4 | [84] | |
| 2015 | North America | Costa Rica | 130 | 20 and 46.2 | [83] | |
| 2017 | North America | Cuba | 100 | 25 and 73 | [99] | |
| 2013 | South America | Venezuela | 136 | 4.4 and 61.8 | [100] | |
| 2017 | South America | Brazil | 36 | 60 and 38.5 | [101] | |
| 2012 | Europe | Netherlands | 300 | NA and 1.6 | [78] | |
| 2013 | Europe | Hungry | 324 | NA and 15.1 | [94] | |
| 2014 | Europe | Romania | 178 | 2.2 and 20.3 | [102] | |
| 2016 | Europe | Italy | 263 | 10.3 and 70.3 | [79] | |
| ELISA | 2008 | Africa | Sudan | 158 | 4.4 and 63.5 | [77] |
| 2016 | Africa | Egypt | 88 | 14.8 and 0 | [48] | |
| 2016 | Africa | Nigeria | 252 | 4.4 and 65.6 | [87] | |
| 2010 | Asia | United Arab Emirates | 105 | 15.2 and 32.5 | [90] | |
| 2011 | Asia | Korea | 184 | 0 and 1.1 | [103] | |
| 2012 | Asia | Jordan | 253 | 0 and 14.6 | [76] | |
| 2013 | Asia | Mongolia | 250 | 51.6 and 19.6 | [97] | |
| 2014 | Asia | China | 1990 | 51.2 and 11.5 | [104] | |
| 2014 | Asia | Thailand | 63 | 1.6 and 0 | [92] | |
| 2016 | Asia | Malaysia | 306 | 63.1 and 51.3 | [88] | |
| 2016 | Asia | India | 160 | 0 and 73.1 | [81] | |
| 2018 | Asia | Indonesia | 235 | 0.4 and 1.7 | [84] | |
| 2009 | South America | Brazil | 538 | 69.6 and 26.6 | [54] | |
| 2013 | South America | Venezuela | 694 | 23.2 and 14.0 | [96] | |
| 2017 | South America | Brazil | 39 | 7.7 and 43.5 | [101] | |
| 2015 | North America | Costa Rica | 130 | 69.2 and 88.5 | [83] | |
| 2010 | Europe | Greece | 524 | 1.1 and 9.2 | [43] | |
| 2016 | Europe | Italy | 673 | 8.9 and 39.8 | [79] | |
| 2017 | Europe | Spain | 108 | 6.5 and 53.7 | [95] | |
| CFT | 2009 | South America | Brazil | 582 | 54.6 and 28.5 | [54] |
| 2015 | Europe | France | 443 | 12.9 and 58.0 | [63] |
| Diagnostic Technique | Year | Continent | Country | Equine Spec (Donkey/Mules) | Sample Size | Prevalence (%) (B. caballi & T. equi) | Reference |
|---|---|---|---|---|---|---|---|
| Microscopy | 2013 | Africa | Ethiopia | Donkeys | 393 | 1.8 and 12.2 | [105] |
| cELISA | 2015 | Africa | Kenya | Donkeys | 314 | 0 and 81.2 | [106] |
| 2016 | Africa | Egypt | Donkeys | 51 | 0 and 18.0 | [48] | |
| ELISA | 2014 | Asia | Thailand | Mules | 177 | 3.4 and 7.3 | [92] |
| 2016 | Asia | India | Donkeys | 20 | 0 and 80.0 | [81] | |
| 2013 | Europe | Spain | Mules | 56 | 32.1 and 66.1 | [65] | |
| 2013 | Europe | Spain | Donkeys | 53 | 17.0 and 47.2 | [65] | |
| 2012 | South America | Brazil | Donkeys | 88 | NA and 73.9 | [107] | |
| IFAT | 2013 | Africa | Ethiopia | Donkeys | 395 | 13.2 and 55.7 | [105] |
| 2016 | Africa | Egypt | Donkeys | 51 | 22.3 and 26.6 | [48] | |
| 2014 | Asia | Thailand | Mules | 177 | 2.8 and 10.7 | [92] | |
| 2015 | Europe | Italy | Donkey | 138 | 40.6 and 47.8 | [44] | |
| 2012 | South America | Brazil | Donkeys | 88 | 93.2 and NA | [107] | |
| PCR | 2016 | Africa | Egypt | Donkeys | 51 | 18 and 38.8 | [48] |
| 2014 | Asia | Thailand | Mules | 177 | 0 and 1.7 | [92] | |
| 2016 | Asia | India | Donkeys/mules | 20 | NA and 35.0 | [81] | |
| 2012 | South America | Brazil | Donkeys | 88 | 20.5 and 31.8 | [107] | |
| 2015 | Europe | Italy | Donkeys | 134 | 17.4 and 3.4 | [44] |
| Year | Continent | Country | Species | Diagnostic Method | Sample Size | Prevalence (%) (B. caballi & T. equi) | Reference |
|---|---|---|---|---|---|---|---|
| 2003 | Europe | Spain, Portugal and France | Dogs | PCR | 10 | NA and 40.0 | [120] |
| 2009 | Europe | Croatia | Dogs | PCR | 81 | 1.3 and 1.3 | [129] |
| 2010 | Europe | France | Dogs | PCR | 166 | 0.6 and 19.0 | [122] |
| 2019 | South America | Paraguay | Dogs | PCR | 284 | NA and 0.3 | [126] |
| 2012 | Asia | Jordan | Camels | PCR | 100 | 60.0 and 40.0 | [123] |
| 2015 | Asia | Iraq | Camels | PCR | 38 | 39.5 and 23.7 | [127] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Onyiche, T.E.; Suganuma, K.; Igarashi, I.; Yokoyama, N.; Xuan, X.; Thekisoe, O. A Review on Equine Piroplasmosis: Epidemiology, Vector Ecology, Risk Factors, Host Immunity, Diagnosis and Control. Int. J. Environ. Res. Public Health 2019, 16, 1736. https://doi.org/10.3390/ijerph16101736
Onyiche TE, Suganuma K, Igarashi I, Yokoyama N, Xuan X, Thekisoe O. A Review on Equine Piroplasmosis: Epidemiology, Vector Ecology, Risk Factors, Host Immunity, Diagnosis and Control. International Journal of Environmental Research and Public Health. 2019; 16(10):1736. https://doi.org/10.3390/ijerph16101736
Chicago/Turabian StyleOnyiche, ThankGod E., Keisuke Suganuma, Ikuo Igarashi, Naoaki Yokoyama, Xuenan Xuan, and Oriel Thekisoe. 2019. "A Review on Equine Piroplasmosis: Epidemiology, Vector Ecology, Risk Factors, Host Immunity, Diagnosis and Control" International Journal of Environmental Research and Public Health 16, no. 10: 1736. https://doi.org/10.3390/ijerph16101736
APA StyleOnyiche, T. E., Suganuma, K., Igarashi, I., Yokoyama, N., Xuan, X., & Thekisoe, O. (2019). A Review on Equine Piroplasmosis: Epidemiology, Vector Ecology, Risk Factors, Host Immunity, Diagnosis and Control. International Journal of Environmental Research and Public Health, 16(10), 1736. https://doi.org/10.3390/ijerph16101736