Mercury Exposure and Poor Nutritional Status Reduce Response to Six Expanded Program on Immunization Vaccines in Children: An Observational Cohort Study of Communities Affected by Gold Mining in the Peruvian Amazon

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
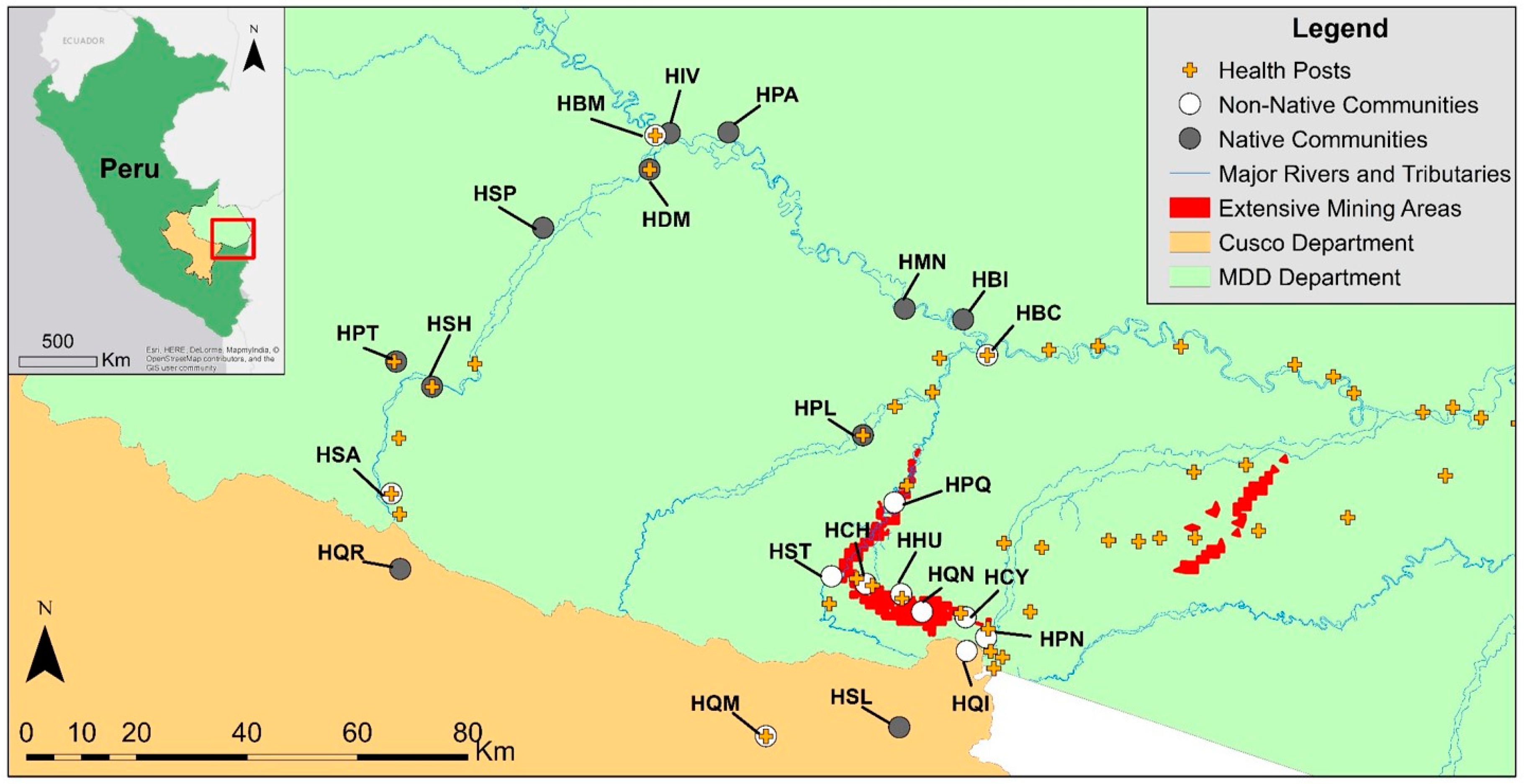
2.1. Study Background and Population
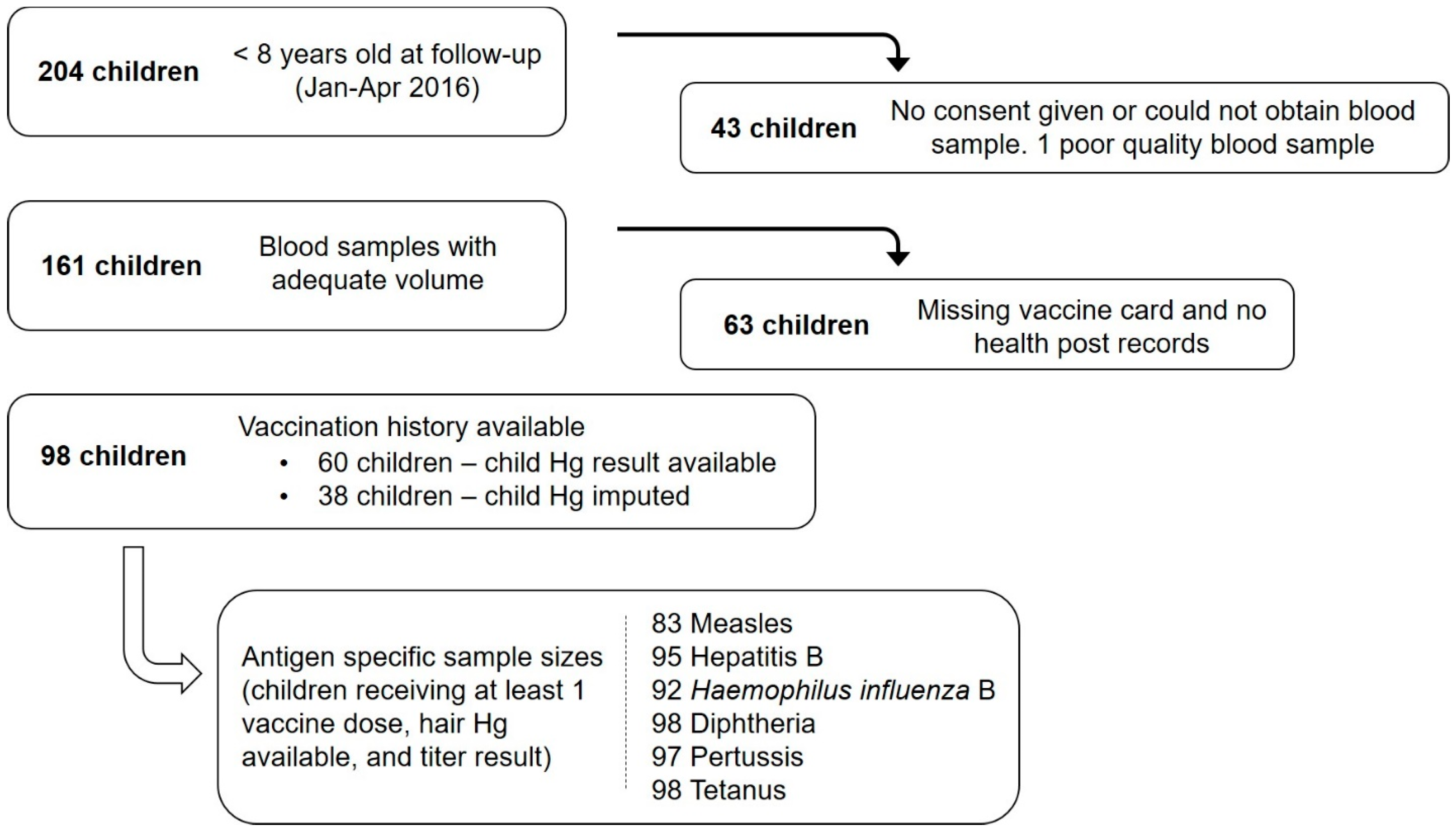
2.2. Study Design and Data Collection
2.3. Hair Mercury Analysis
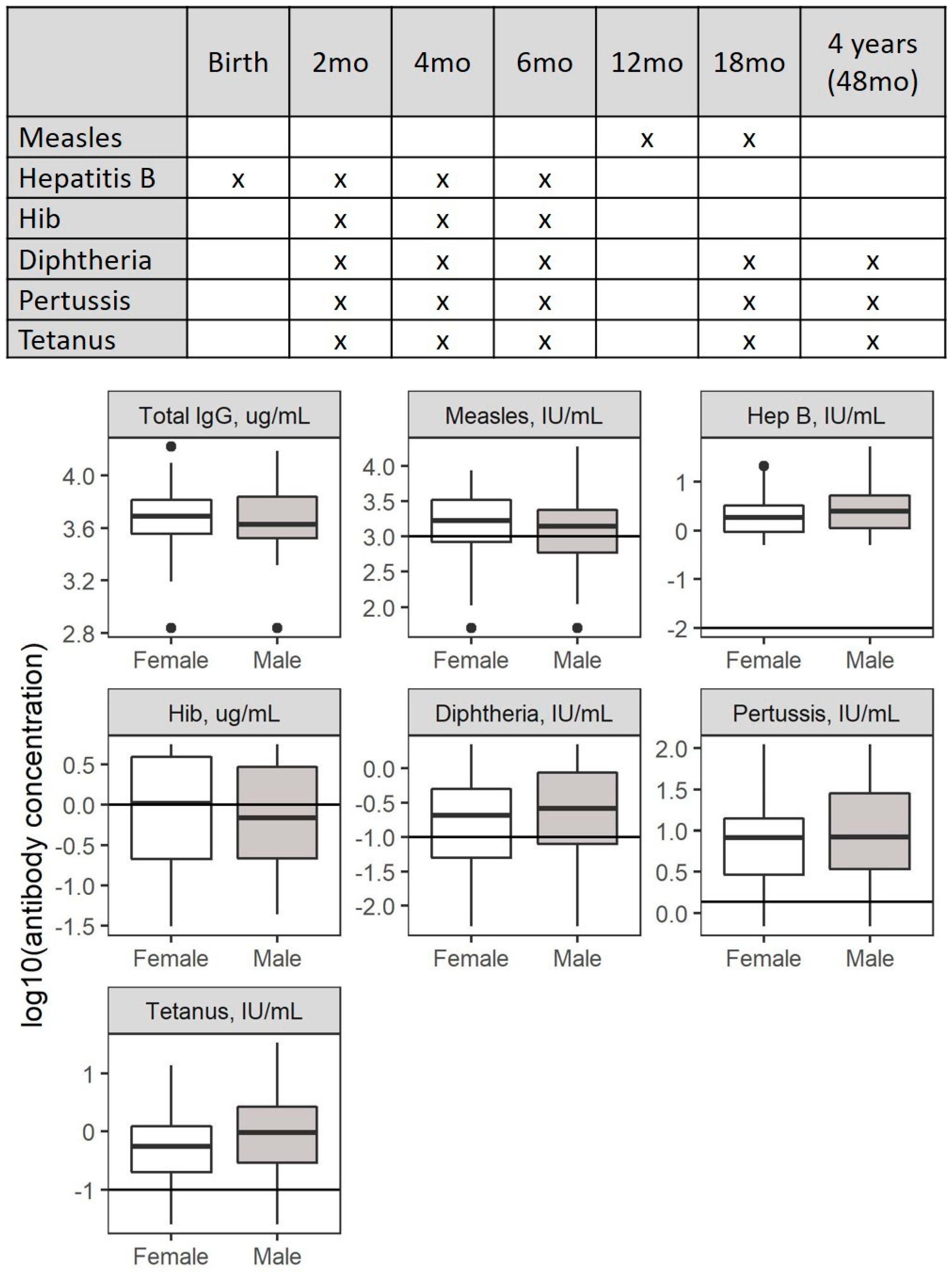
2.4. Antibody Analysis
2.5. Statistical Analysis
Risk Factors Associated with Changes in Antibody Concentrations and Levels That Are Indicative of Being Not Protected or a Non-Responder
3. Results
3.1. Population Characteristics
3.1.1. Child and Parental Hair Mercury Content
3.1.2. Nutritional Status
3.1.3. Vaccination Coverage and Response
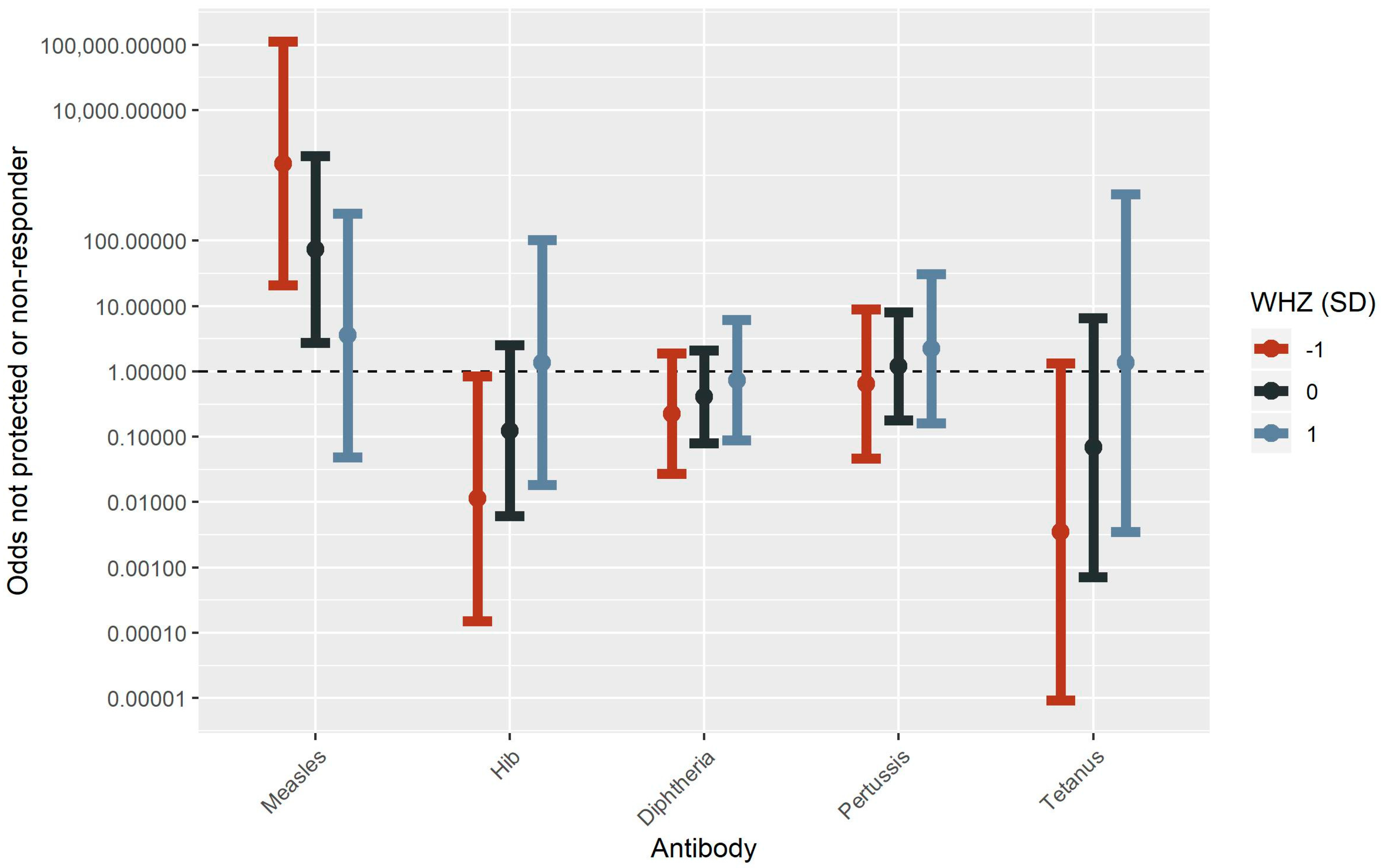
3.2. Risk Factors Associated with Changes in Antibody Concentrations
3.2.1. Community Effects
3.2.2. Malnutrition
3.2.3. Hair Mercury
3.2.4. Interaction between Mercury and Malnutrition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Heilmann, C.; Grandjean, P.; Weihe, P.; Nielsen, F.; Budtz-Jorgensen, E. Reduced antibody responses to vaccinations in children exposed to polychlorinated biphenyls. PLoS Med. 2006, 3, e311. [Google Scholar] [CrossRef]
- Jusko, T.A.; De Roos, A.J.; Lee, S.Y.; Thevenet-Morrison, K.; Schwartz, S.M.; Verner, M.A.; Murinova, L.P.; Drobna, B.; Kocan, A.; Fabisikova, A.; et al. A Birth Cohort Study of Maternal and Infant Serum PCB-153 and DDE Concentrations and Responses to Infant Tuberculosis Vaccination. Environ. Health Perspect. 2016, 124, 813–821. [Google Scholar] [CrossRef] [Green Version]
- Feingold, B.J.; Vegosen, L.; Davis, M.; Leibler, J.; Peterson, A.; Silbergeld, E.K. A Niche for Infectious Disease in Environmental Health: Rethinking the Toxicological Paradigm. Environ. Health Perspect. 2010, 118, 1165–1172. [Google Scholar] [CrossRef] [Green Version]
- Suk, W.; Ruchirawat, M.; Stein, R.T.; Diaz-Barriga, F.; Carpenter, D.O.; Neira, M.; Sly, P.D. Health Consequences of Environmental Exposures in Early Life: Coping with a Changing World in the Post-MDG Era. Ann. Glob. Health 2016, 82, 20–27. [Google Scholar] [CrossRef]
- Kordas, K.; Lonnerdal, B.; Stoltzfus, R.J. Interactions between nutrition and environmental exposures: Effects on health outcomes in women and children. J. Nutr. 2007, 137, 2794–2797. [Google Scholar] [CrossRef]
- Malm, O.; Branches, F.J.P.; Akagi, H.; Castro, M.B.; Pfeiffer, W.C.; Harada, M.; Bastos, W.R.; Kato, H. Mercury and methylmercury in fish and human hair from the Tapajos river basin, Brazil. Sci. Total Environ. 1995, 175, 141–150. [Google Scholar] [CrossRef]
- Bidone, E.D.; Castilhos, Z.C.; Santos, T.J.S.; Souza, T.M.C.; Lacerda, L.D. Fish contamination and human exposure to mercury in Tartarugalzinho River, Amapa State, Northern Amazon, Brazil. A screening approach. Water Air Soil Pollut. 1997, 97, 9–15. [Google Scholar] [CrossRef]
- Grandjean, P.; Cardoso, B.; Guimarães, G. Mercury poisoning. Lancet 1993, 342, 991. [Google Scholar] [CrossRef]
- Passos, C.J.; Mergler, D.; Gaspar, E.; Morais, S.; Lucotte, M.; Larribe, F.; Davidson, R.; de Grosbois, S. Eating tropical fruit reduces mercury exposure from fish consumption in the Brazilian Amazon. Environ. Res. 2003, 93, 123–130. [Google Scholar] [CrossRef]
- Wyatt, L.; Ortiz, E.J.; Feingold, B.; Berky, A.; Diringer, S.; Morales, A.M.; Jurado, E.R.; Hsu-Kim, H.; Pan, W. Spatial, Temporal, and Dietary Variables Associated with Elevated Mercury Exposure in Peruvian Riverine Communities Upstream and Downstream of Artisanal and Small-Scale Gold Mining. Int. J. Environ. Res. Public Health 2017, 14, 1582. [Google Scholar] [CrossRef]
- Fallacara, D.M.; Halbrook, R.S.; French, J.B. Toxic effects of dietary methylmercury in immune system development in nestling American kestrels (Falco sparverius). Environ. Toxicol. Chem. 2011, 30, 1328–1337. [Google Scholar] [CrossRef]
- Koller, L.D. Immunosuppression produced by lead, cadmium, and mercury. Am. J. Vet. Res. 1973, 34, 1457–1458. [Google Scholar]
- Koller, L.D.; Exon, J.H.; Brauner, J.A. Methylmercury: Decreased Antibody Formation in Mice. Proc. Soc. Exp. Biol. Med. 1977, 155, 602–604. [Google Scholar] [CrossRef] [Green Version]
- Pyszel, A.; Wrobel, T.; Szuba, A.; Andrzejak, R. Effect of metals, benzene, pesticides and ethylene oxide on the haematopoietic system. Med. Pr. 2005, 56, 249–255. [Google Scholar]
- Weinhouse, C.; Ortiz, E.J.; Berky, A.J.; Bullins, P.; Hare-Grogg, J.; Rogers, L.; Morales, A.M.; Hsu-Kim, H.; Pan, W.K. Hair Mercury Level is Associated with Anemia and Micronutrient Status in Children Living Near Artisanal and Small-Scale Gold Mining in the Peruvian Amazon. Am. J. Trop. Med. Hyg. 2017, 97, 1886–1897. [Google Scholar] [CrossRef]
- Ahamed, M.; Singh, S.; Behari, J.R.; Kumar, A.; Siddiqui, M.K.J. Interaction of lead with some essential trace metals in the blood of anemic children from Lucknow, India. Clin. Chim. Acta 2007, 377, 92–97. [Google Scholar] [CrossRef]
- Shah, F.; Kazi, T.G.; Afridi, H.I.; Baig, J.A.; Khan, S.; Kolachi, N.F.; Wadhwa, S.K.; Shah, A.Q. Environmental exposure of lead and iron deficit anemia in children age ranged 1–5 years: A cross sectional study. Sci. Total Environ. 2010, 408, 5325–5330. [Google Scholar] [CrossRef]
- Wright, R.O.; Tsaih, S.W.; Schwartz, J.; Wright, R.J.; Hu, H. Association between iron deficiency and blood lead level in a longitudinal analysis of children followed in an urban primary care clinic. J. Pediatr. 2003, 142, 9–14. [Google Scholar] [CrossRef]
- Bradman, A.; Eskenazi, B.; Sutton, P.; Athanasoulis, M.; Goldman, L.R. Iron deficiency associated with higher blood lead in children living in contaminated environments. Environ. Health Perspect. 2001, 109, 1079–1084. [Google Scholar] [CrossRef]
- Gaayeb, L.; Sarr, J.B.; Cames, C.; Pinçon, C.; Hanon, J.-B.; Ndiath, M.O.; Seck, M.; Herbert, F.; Sagna, A.B.; Schacht, A.-M.; et al. Effects of Malnutrition on Children’s Immunity to Bacterial Antigens in Northern Senegal. Am. J. Trop. Med. Hyg. 2014, 90, 566–573. [Google Scholar] [CrossRef]
- Rey-Cuille, M.-A.; Seck, A.; Njouom, R.; Chartier, L.; Sow, H.D.; Mamadou; Ka, A.S.; Njankouo, M.; Rousset, D.; Giles-Vernick, T.; Unal, G.; et al. Low Immune Response to Hepatitis B Vaccine among Children in Dakar, Senegal. PLoS ONE 2012, 7, e38153. [Google Scholar] [CrossRef]
- el-Gamal, Y.; Aly, R.H.; Hossny, E.; Afify, E.; el-Taliawy, D. Response of Egyptian infants with protein calorie malnutrition to hepatitis B vaccination. J. Trop. Pediatr. 1996, 42, 144–145. [Google Scholar] [CrossRef] [Green Version]
- Suskind, R.; Sirishinha, S.; Vithayasai, V.; Edelman, R.; Damrongsak, D.; Charupatana, C.; Olson, R.E. Immunoglobulins and antibody response in children with protein-calorie malnutrition. Am. J. Clin. Nutr. 1976, 29, 836–841. [Google Scholar] [CrossRef] [Green Version]
- Wesley, A.; Coovadia, H.M.; Watson, A.R. Immunization against measles in children at risk for severe disease. Trans. R. Soc. Trop. Med. Hyg. 1979, 73, 710–715. [Google Scholar] [CrossRef]
- Heilmann, C.; Budtz-Jorgensen, E.; Nielsen, F.; Heinzow, B.; Weihe, P.; Grandjean, P. Serum Concentrations of Antibodies against Vaccine Toxoids in Children Exposed Perinatally to Immunotoxicants. Environ. Health Perspect. 2010, 118, 1434–1438. [Google Scholar] [CrossRef]
- Gallagher, C.M.; Smith, D.M.; Golightly, M.G.; Meliker, J.R. Total blood mercury and rubella antibody concentrations in US children aged 6–11 years, NHANES 2003–2004. Sci. Total Environ. 2013, 442, 48–55. [Google Scholar] [CrossRef]
- Gallagher, C.M.; Smith, D.M.; Meliker, J.R. Total blood mercury and serum measles antibodies in US children, NHANES 2003–2004. Sci. Total Environ. 2011, 410, 65–71. [Google Scholar] [CrossRef]
- Ilback, N.G.; Wesslen, L.; Fohlman, J.; Friman, G. Effects of methyl mercury on cytokines, inflammation and virus clearance in a common infection (Coxsackie B3 myocarditis). Toxicol. Lett. 1996, 89, 19–28. [Google Scholar] [CrossRef]
- Christensen, M.M.; EllermannEriksen, S.; Rungby, J.; Mogensen, S.C. Influence of mercuric chloride on resistance to generalized infection with herpes simplex virus type 2 in mice. Toxicology 1996, 114, 57–66. [Google Scholar] [CrossRef]
- Sheehan, M.C.; Burke, T.A.; Breysse, P.N.; Navas-Acien, A.; McGready, J.; Fox, M.A. Association of markers of chronic viral hepatitis and blood mercury levels in US reproductive-age women from NHANES 2001–2008: A cross-sectional study. Environ. Health 2012, 11, 62. [Google Scholar] [CrossRef]
- National Research Council. Toxicological Effects of Methylmercury; National Academy Press: Washington, DC, USA, 2000. [Google Scholar]
- U.S. Environmental Protection Agency. Integrated Risk Information System: Methylmercury (MeHg). (CASRN 22967-92-6); 2001. Available online: https://cfpub.epa.gov/ncea/iris/iris_documents/documents/subst/0073_summary.pdf (accessed on 14 January 2019).
- Oken, E.; Wright, R.O.; Kleinman, K.P.; Bellinger, D.; Amarasiriwardena, C.J.; Hu, H.; Rich-Edwards, J.W.; Gillman, M.W. Maternal fish consumption, hair mercury, and infant cognition in a US cohort. Environ. Health Perspect. 2005, 113, 1376–1380. [Google Scholar] [CrossRef] [PubMed]
- Instituto Nacional de Estadística e Informática (INEI). Estado de la Población Peruana 2015. 2015; 56p. Available online: https://www.inei.gob.pe/media/MenuRecursivo/publicaciones_digitales/Est/Lib1251/Libro.pdf (accessed on 14 January 2019).
- Asner, G.P.; Llactayo, W.; Tupayachi, R.; Luna, E.R. Elevated rates of gold mining in the Amazon revealed through high-resolution monitoring. Proc. Natl. Acad. Sci. USA 2013, 110, 18454–18459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swenson, J.J.; Carter, C.E.; Domec, J.C.; Delgado, C.I. Gold Mining in the Peruvian Amazon: Global Prices, Deforestation, and Mercury Imports. PLoS ONE 2011, 6, e18875. [Google Scholar] [CrossRef] [PubMed]
- Caballero Espejo, J.; Messinger, M.; Román-Dañobeytia, F.; Ascorra, C.; Fernandez, L.; Silman, M. Deforestation and Forest Degradation Due to Gold Mining in the Peruvian Amazon: A 34-Year Perspective. Remote Sens. 2018, 10, 1903. [Google Scholar] [CrossRef]
- Diringer, S.E.; Feingold, B.J.; Ortiz, E.J.; Gallis, J.A.; Araujo-Flores, J.M.; Berky, A.; Pan, W.K.Y.; Hsu-Kim, H. River transport of mercury from artisanal and small-scale gold mining and risks for dietary mercury exposure in Madre de Dios, Peru. Environ. Sci. Processes Impacts 2015, 17, 478–487. [Google Scholar] [CrossRef] [PubMed]
- Fidel Suarez, M.; Aramayo Berdeja, J.L. Atlas de Salud 2008—Departamento de Pando; OPS/OMS, SEDES: Pando, Bolivia, 2010; 101p. [Google Scholar]
- Pan, W.; Erlien, C.; Bilsborrow, R.E. Morbidity and mortality disparities among colonist and indigenous populations in the Ecuadorian Amazon. Soc. Sci. Med. 2010, 70, 401–411. [Google Scholar] [Green Version]
- Instituto Nacional de Estadística e Informática (INEI). Encuesta Demográfica y de Salud Familiar—ENDES 2014; Nacional y Departamental: Lima, Peru, 2014.
- Instituto Nacional de Estadística e Informática (INEI). Encuesta Demográfica y de Salud Familiar—ENDES 2017; Nacional y Departamental: Lima, Peru, 2017.
- Instituto Nacional de Estadística e Informática (INEI). Encuesta Demografia y de Salud Familiar—ENDES 2015; Nacional y Departamental: Lima, Peru, 2015.
- Itell, H.L.; McGuire, E.P.; Muresan, P.; Cunningham, C.K.; McFarland, E.J.; Borkowsky, W.; Permar, S.R.; Fouda, G.G. Development and application of a multiplex assay for the simultaneous measurement of antibody responses elicited by common childhood vaccines. Vaccine 2018, 36, 5600–5608. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Haemoglobin Concentrations for the Diagnosis of Anaemia and Assessment of Severity; WHO: Geneva, Switzerland, 2011. [Google Scholar]
- Myatt, M.; Guevarra, E. zscorer: Child Anthropometry z-Score Calculator. 2018. Available online: https://rdrr.io/cran/zscorer/ (accessed on 14 January 2019).
- Centers for Disease Control and Prevention (CDC). Recommendations of the Advisory Committee on Immunization Practices (ACIP): Prevention of Pertussis, Tetanus, and Diphtheria among Pregnant and Postpartum Women and Their Infants. MMWR Recomm. Rep. 2008, 57, 1–47. [Google Scholar]
- Centers for Disease Control and Prevention (CDC). Recommendations of the Advisory Committee on Immunization Practices (ACIP) Part II: Immunization of Adults: A Comprehensive Immunization Strategy to Eliminate Transmission of Hepatitis B Virus Infection in the United States. MMWR Recomm. Rep. 2006, 54, 1–25. [Google Scholar]
- Centers for Disease Control and Prevention (CDC). Prevention and Control of Haemophilus influenzae Type b Disease: Recommendations of the Advisory Committee on Immunization Practices (ACIP). MMWR Recomm. Rep. 2014, 63, 1–14. [Google Scholar]
- Plotkin, S.A. Correlates of Protection Induced by Vaccination. Clin. Vaccine Immunol. 2010, 17, 1055–1065. [Google Scholar] [CrossRef] [Green Version]
- Mkhize, N.N.; Durgiah, R.; Ashley, V.; Archary, D.; Garrett, N.J.; Karim, Q.A.; Karim, S.S.A.; Moore, P.L.; Yates, N.; Passmore, J.-A.S.; et al. Broadly neutralizing antibody specificities detected in the genital tract of HIV-1 infected women. AIDS 2016, 30, 1005–1014. [Google Scholar] [CrossRef] [PubMed]
- Permar, S.R.; Fong, Y.; Vandergrift, N.; Fouda, G.G.; Gilbert, P.; Parks, R.; Jaeger, F.H.; Pollara, J.; Martelli, A.; Liebl, B.E.; et al. Maternal HIV-1 envelope-specific antibody responses and reduced risk of perinatal transmission. J. Clin. Investig. 2015, 125, 2702–2706. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: http://www.R-project.org/ (accessed on 14 January 2019).
- Chang, Y.T.; Guo, C.Y.; Tsai, M.S.; Cheng, Y.Y.; Lin, M.T.; Chen, C.H.; Shen, D.; Wang, J.R.; Sung, J.M. Poor immune response to a standard single dose non-adjuvanted vaccination against 2009 pandemic H1N1 influenza virus A in the adult and elder hemodialysis patients. Vaccine 2012, 30, 5009–5018. [Google Scholar] [CrossRef] [PubMed]
- Hassan, K.; Shternberg, L.; Alhaj, M.; Giron, R.; Reshef, R.; Barak, M.; Kristal, B. The effect of erythropoietin therapy and hemoglobin levels on the immune response to engerix-B vaccination in chronic kidney disease. Ren. Fail. 2003, 25, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Miyagawa, K.; Hayashi, Y.; Kurihara, S.; Maeda, A. Co-administration of l-cystine and l-theanine enhances efficacy of influenza vaccination in elderly persons: Nutritional status-dependent immunogenicity. Geriatr. Gerontol. Int. 2008, 8, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.J.; Lee, S.H.; Byun, Y.H.; Yun, G.Y.; Kim, S.K.; Seong, B.L.; Kim, A.R.; Park, E.S.; Kim, H.J.; Lee, J.E.; et al. Risk factors affecting seroconversion after influenza A/H1N1 vaccination in hemodialysis patients. BMC Nephrol. 2012, 13, 165. [Google Scholar] [CrossRef] [PubMed]
- Nyland, J.F.; Wang, S.B.; Shirley, D.L.; Santos, E.O.; Ventura, A.M.; de Souza, J.M.; Silbergeld, E.K. Fetal and maternal immune responses to methylmercury exposure: A cross-sectional study. Environ. Res. 2011, 111, 584–589. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, M.; Khan, N.; Muniroh, M.; Motomura, E.; Yanagisawa, R.; Matsuyama, T.; Vogel, C.F.A. Activation of interleukin-6 and-8 expressions by methylmercury in human U937 macrophages involves RelA and p50. J. Appl. Toxicol. 2017, 37, 611–620. [Google Scholar] [CrossRef]
- Grandjean, P.; Weihe, P.; White, R.F.; Debes, F.; Araki, S.; Yokoyama, K.; Murata, K.; Sorensen, N.; Dahl, R.; Jorgensen, P.J. Cognitive deficit in 7-year-old children with prenatal exposure to methylmercury. Neurotoxicol. Teratol. 1997, 19, 417–428. [Google Scholar] [CrossRef]
- Feeney, A.J.; Atkinson, M.J.; Cowan, M.J.; Escuro, G.; Lugo, G. A defective V-kappa A2 allele in Navajos which play play a role in increased susceptibility to Haemophilus influenzae type disease. J. Clin. Investig. 1996, 97, 2277–2282. [Google Scholar] [CrossRef] [PubMed]
- Siber, G.R.; Santosham, M.; Reid, G.R.; Thompson, C.; Almeido-Hill, J.; Morell, A.; deLange, G.; Ketcham, J.K.; Callahan, E.H. Impaired Antibody Response to Haemophilus influenzae Type b Polysaccharide and Low IgG2 and IgG4 Concentrations in Apache Children. N. Engl. J. Med. 1990, 323, 1387–1392. [Google Scholar] [CrossRef] [PubMed]
- Guthridge, S.; McIntyre, P.; Isaacs, D.; Hanlon, M.; Patel, M. Differing serologic responses to an Haemophilus influenzae type b polysaccharide-Neisseria meningitidis outer membrane protein conjugate (PRP-OMPC) vaccine in Australian Aboriginal and Caucasian infants—Implications for disease epidemiology. Vaccine 2000, 18, 2584–2591. [Google Scholar] [CrossRef]
- Menzies, R.; Turnour, C.; Chiu, C.; McIntyre, P. Vaccine preventable diseases and vaccination coverage in Aboriginal and Torres Strait Islander people, Australia 2003 to 2006. Commun. Dis. Intell. Q. Rep. 2008, 32, S2–S67. [Google Scholar] [PubMed]
- Millar, E.V.; O’Brien, K.L.; Levine, O.S.; Kvamme, S.; Reid, R.; Santosham, M. Toward elimination of Haemophilus influenzae type B carriage and disease among high-risk American Indian children. Am. J. Public Health 2000, 90, 1550–1554. [Google Scholar] [PubMed]
- Singleton, R.; Hammitt, L.; Hennessy, T.; Bulkow, L.; DeByle, C.; Parkinson, A.; Cottle, T.E.; Peters, H.; Butler, J.C. The Alaska Haemophilus influenzae type b experience: Lessons in controlling a vaccine-preventable disease. Pediatrics 2006, 118, e421–e429. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Immunization in Practice: A Practical Guide for Health Staff—2015 Update. 2015. Available online: https://apps.who.int/iris/bitstream/handle/10665/193412/9789241549097_eng.pdf?sequence=1 (accessed on 14 January 2019).
- Davila, S.; Froeling, F.E.M.; Tan, A.; Bonnard, C.; Boland, G.J.; Snippe, H.; Hibberd, M.L.; Seielstad, M. New genetic associations detected in a host response study to hepatitis B vaccine. Genes Immun. 2010, 11, 232–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jepson, A.; Banya, W.; SisayJoof, F.; HassanKing, M.; Nunes, C.; Bennett, S.; Whittle, H. Quantification of the relative contribution of major histocompatibility complex (MHC) and non-MHC genes to human immune responses to foreign antigens. Infect. Immun. 1997, 65, 872–876. [Google Scholar]
- Milet, J.; Sabbagh, A.; Migot-Nabias, F.; Luty, A.J.F.; Gaye, O.; Garcia, A.; Courtin, D. Genome-wide association study of antibody responses to Plasmodium falciparum candidate vaccine antigens. Genes Immun. 2016, 17, 110–117. [Google Scholar] [CrossRef]
- Lee, Y.C.; Newport, M.J.; Goetghebuer, T.; Siegrist, C.A.; Weiss, H.A.; Pollard, A.J.; Marchant, A.; the MRC Twin Study Group. Influence of genetic and environmental factors on the immunogenicity of Hib vaccine in Gambian twins. Vaccine 2006, 24, 5335–5340. [Google Scholar] [CrossRef]
- Ward, J.; Brenneman, G.; Letson, G.W.; Heyward, W.L.; the Alaska H. influenzae Vaccine Study Group. Limited Efficacy of a Haemophilus influenzae Type b Conjugate Vaccine in Alaska Native Infants. N. Engl. J. Med. 1990, 323, 1393–1401. [Google Scholar] [CrossRef] [PubMed]
- Tsang, R.S.; Bruce, M.G.; Lem, M.; Barreto, L.; Ulanova, M. A review of invasive Haemophilus influenzae disease in the Indigenous populations of North America. Epidemiol. Infect. 2014, 142, 1344–1354. [Google Scholar] [CrossRef] [PubMed]
- Jacups, S.P.; Morris, P.S.; Leach, A.J. Haemophilus influenzae type b carriage in Indigenous children and children attending childcare centers in the Northern Territory, Australia, spanning pre- and post-vaccine eras. Vaccine 2011, 29, 3083–3088. [Google Scholar] [CrossRef] [PubMed]
- Petersen, G.M.; Silimperi, D.R.; Rotter, J.I.; Terasaki, P.I.; Schanfield, M.S.; Park, M.S.; Ward, J.I. Genetic factors in Haemophilus influenzae type b disease susceptibility and antibody acquisition. J. Pediatr. 1987, 110, 228–233. [Google Scholar] [CrossRef]
- Bulkow, L.R.; Wainwright, R.B.; Letson, G.W.; Chang, S.J.; Ward, J.I. Compariative Immunogenicity of Four Haemophilus-influenzae Type B Conjugate Vaccines in Alaksa Native Infants. Pediatr. Infect. Dis. J. 1993, 12, 484–492. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Population Characteristic | Native Communities | Non-Native Communities | p-Value | All Study Children | NHANES | p-Value | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| N | Mean (SD) | N | Mean (SD) | N | Mean (SD) | N | Mean (SD) | ||||
| Age, years | |||||||||||
| <4 | 22 | 2.5 (1.0) | 46 | 2.6 (1.0) | 0.978 | 68 | 2.6 (1.0) | 239 | 2.4 (0.5) | 0.184 | |
| 4 to <8 | 29 | 5.7 (1.1) | 64 | 5.6 (0.9) | 0.581 | 93 | 5.6 (1.0) | 214 | 4.5 (0.5) | <0.001 | |
| Sex | |||||||||||
| Female | 25 | - | 59 | - | - | 84 | - | 238 | - | - | |
| Male | 26 | - | 51 | - | - | 77 | - | 215 | - | - | |
| Child hair Hg | |||||||||||
| Hg content, µg/g | 35 | 3.8 (2.6) | 78 | 1.0 (2.7) | <0.001 | 113 | 1.5 (3.2) | 453 | 0.2 (2.7) | <0.001 | |
| Exceed 1.2 µg/g, % | 88.6 | - | 42.3 | - | - | 56.6 | - | 2.9 | - | - | |
| Exceed 2.0 µg/g, % | 82.9 | - | 24.3 | - | - | 42.5 | - | 1.5 | - | - | |
| Parental average hair Hg | |||||||||||
| Hg content, µg/g | 51 | 5.1 (2.1) | 107 | 1.6 (2.4) | <0.001 | 158 | 2.3 (2.7) | - | - | - | |
| Nutritional status | |||||||||||
| Hemoglobin, g/dL | 50 | 10.9 (1.2) | 108 | 11.6 (1.2) | 0.001 | 158 | 11.3 (1.2) | 453 | 12.6 (0.8) | <0.001 | |
| Anemic, % | 52 | - | 27.8 | - | - | 35.4 | - | 1.5 | - | - | |
| Stunted, % | 18.4 | - | 6.8 | - | - | 10.5 | - | 1.3 | - | - | |
| HAZ | 49 | −1.3 (1.0) | 103 | −0.6 (1.0) | <0.001 | 152 | −0.8 (1.0) | 453 | 1.0 (1.4) | <0.001 | |
| WAZ | 50 | −0.6 (1.1) | 107 | 0.1 (1.2) | 0.001 | 157 | −0.1 (1.2) | 453 | 0.9 (1.3) | <0.001 | |
| WHZ | 48 | 0.5 (1.0) | 99 | 0.7 (1.1) | 0.189 | 147 | 0.6 (1.1) | 453 | 0.4 (1.1) | 0.026 | |
| Vaccination Coverage, % a | |||||||||||
| Measles | 68.6 | - | 60.0 | - | - | 62.7 | - | - | - | - | |
| Hepatitis B | 74.5 | - | 70.0 | - | - | 71.4 | - | - | - | - | |
| Hib | 74.5 | - | 66.4 | - | - | 68.9 | - | - | - | - | |
| Diphtheria | 76.5 | - | 73.6 | - | - | 74.5 | - | - | - | - | |
| Pertussis | 76.5 | - | 72.7 | - | - | 73.9 | - | - | - | - | |
| Tetanus | 76.5 | - | 73.6 | - | - | 74.5 | - | - | - | - | |
| Vaccine Coverage, % b | |||||||||||
| Hepatitis B (birth dose) | 62.2 | - | 73.8 | - | - | 69.4 | - | - | - | - | |
| Pentavalent | 89.2 | - | 86.9 | - | - | 87.8 | - | - | - | - | |
| DPT | 64.9 | - | 63.9 | - | - | 64.3 | - | - | - | - | |
| MMR | 45.9 | - | 44.2 | - | - | 44.9 | - | - | - | - | |
| Antibody Titer c | |||||||||||
| Total IgG (µg/mL) | 51 | 4881.9 (1.8) | 110 | 4464.0 (1.7) | 0.181 | 161 | 4592.4 (1.7) | - | - | - | |
| Measles (IU/mL) | 51 | 1034.6 (3.0) | 110 | 1400.7 (3.4) | 0.029 | 161 | 1272.5 (3.3) | - | - | - | |
| Hepatitis B (IU/mL) | 51 | 2.0 (3.2) | 110 | 2.1 (2.9) | 0.943 | 161 | 2.1 (3.0) | - | - | - | |
| Hib (µg/mL) | 51 | 0.4 (4.5) | 110 | 1.0 (4.4) | 0.004 | 161 | 0.8 (4.6) | - | - | - | |
| Diphtheria (IU/mL) | 51 | 0.1 (5.8) | 110 | 0.2 (5.4) | 0.045 | 161 | 0.2 (5.6) | - | - | - | |
| Pertussis (IU/mL) | 51 | 7.1 (4.3) | 110 | 7.6 (3.9) | 0.800 | 161 | 7.4 (4.0) | - | - | - | |
| Tetanus (IU/mL) | 51 | 0.5 (4.5) | 110 | 0.6 (4.8) | 0.201 | 161 | 0.6 (4.7) | - | - | - | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wyatt, L.; Permar, S.R.; Ortiz, E.; Berky, A.; Woods, C.W.; Amouou, G.F.; Itell, H.; Hsu-Kim, H.; Pan, W. Mercury Exposure and Poor Nutritional Status Reduce Response to Six Expanded Program on Immunization Vaccines in Children: An Observational Cohort Study of Communities Affected by Gold Mining in the Peruvian Amazon. Int. J. Environ. Res. Public Health 2019, 16, 638. https://doi.org/10.3390/ijerph16040638
Wyatt L, Permar SR, Ortiz E, Berky A, Woods CW, Amouou GF, Itell H, Hsu-Kim H, Pan W. Mercury Exposure and Poor Nutritional Status Reduce Response to Six Expanded Program on Immunization Vaccines in Children: An Observational Cohort Study of Communities Affected by Gold Mining in the Peruvian Amazon. International Journal of Environmental Research and Public Health. 2019; 16(4):638. https://doi.org/10.3390/ijerph16040638
Chicago/Turabian StyleWyatt, Lauren, Sallie Robey Permar, Ernesto Ortiz, Axel Berky, Christopher W. Woods, Genevieve Fouda Amouou, Hannah Itell, Heileen Hsu-Kim, and William Pan. 2019. "Mercury Exposure and Poor Nutritional Status Reduce Response to Six Expanded Program on Immunization Vaccines in Children: An Observational Cohort Study of Communities Affected by Gold Mining in the Peruvian Amazon" International Journal of Environmental Research and Public Health 16, no. 4: 638. https://doi.org/10.3390/ijerph16040638
APA StyleWyatt, L., Permar, S. R., Ortiz, E., Berky, A., Woods, C. W., Amouou, G. F., Itell, H., Hsu-Kim, H., & Pan, W. (2019). Mercury Exposure and Poor Nutritional Status Reduce Response to Six Expanded Program on Immunization Vaccines in Children: An Observational Cohort Study of Communities Affected by Gold Mining in the Peruvian Amazon. International Journal of Environmental Research and Public Health, 16(4), 638. https://doi.org/10.3390/ijerph16040638