Analysis of Genetic Characterization and Clonality of Legionella pneumophila Isolated from Cooling Towers in Japan


Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains
2.2. SBT
2.3. Data Analysis
2.4. Genome Sequencing of the 25 L. pneumophila Strains
2.5. Nucleotide Sequence Accession Number
3. Results
3.1. Distribution of the SGs of L. pneumophila
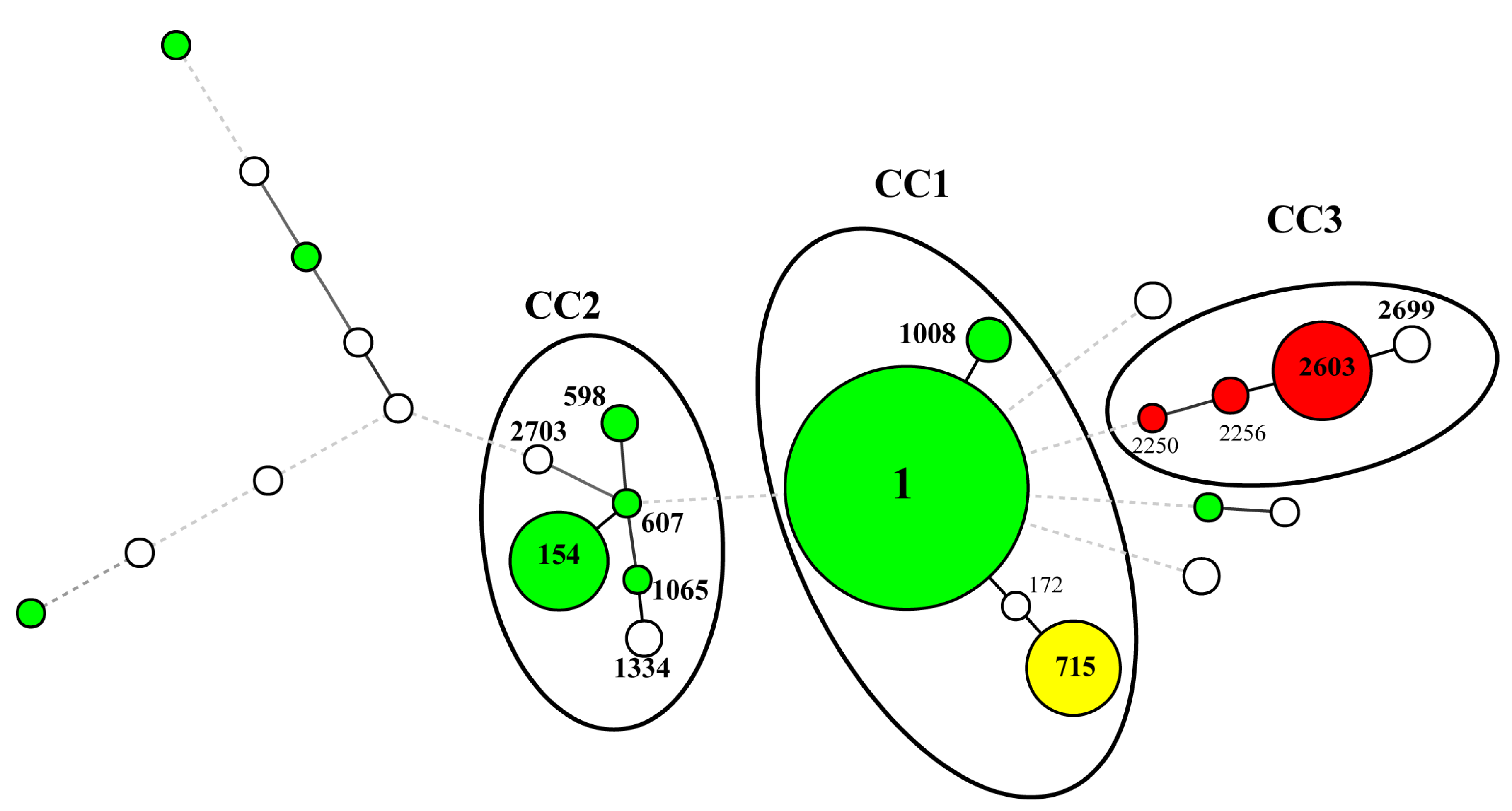
3.2. SBT Analysis of L. pneumophila
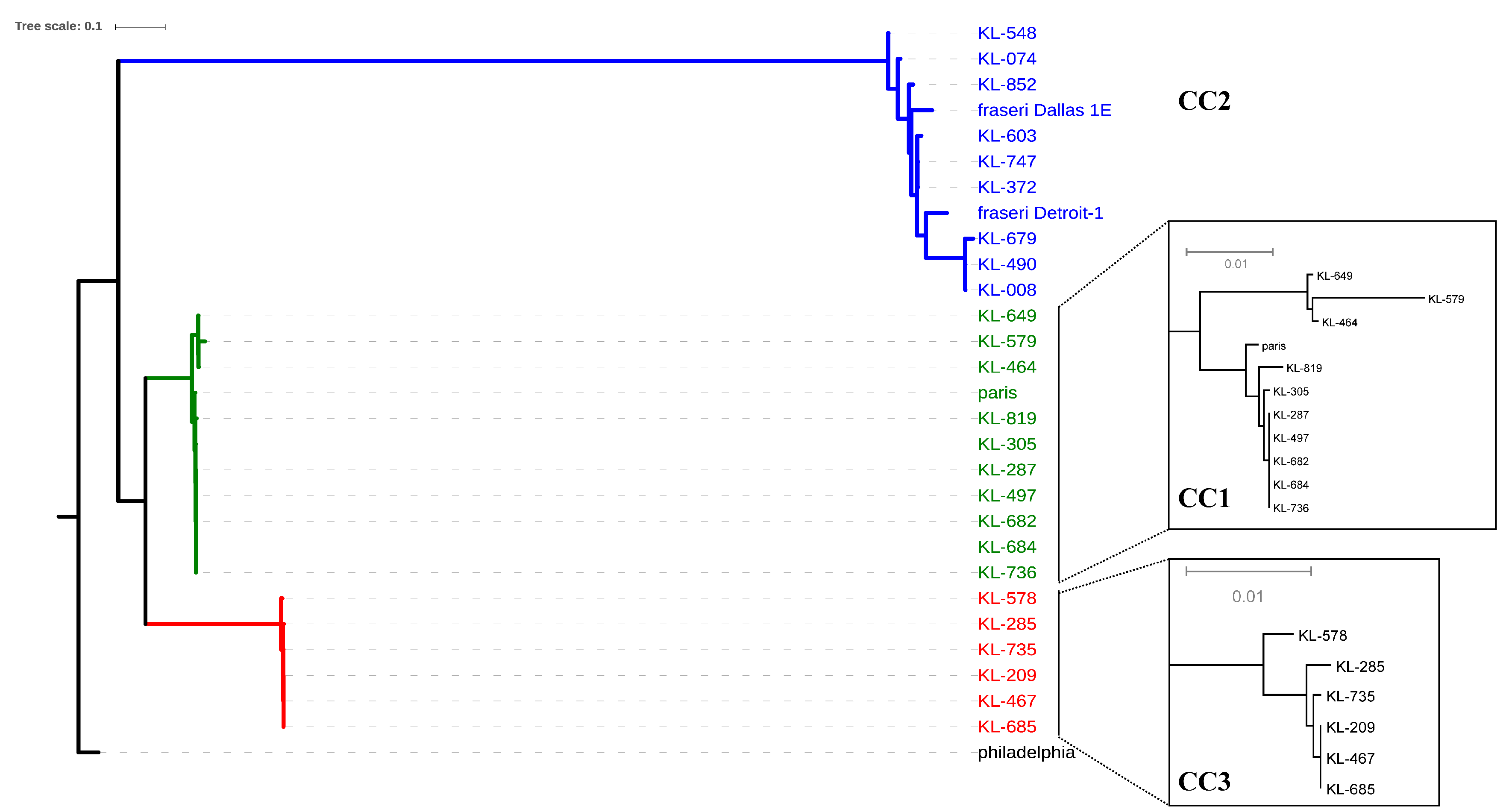
3.3. Whole Genome Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| CC | Clonal complex |
| SBT | Sequence-based typing |
| WGS | Whole genome sequencing |
| UT | Untypable |
| ST | Sequence type |
| SG | Serogroup |
| ANI | Average nucleotide identity |
| SNP | Single nucleotide polymorphism |
References
- Fields, B.S.; Benson, R.F.; Besser, R.E. Legionella and Legionnaires’ disease: 25 years of investigation. Clin. Microbiol. Rev. 2002, 15, 506–526. [Google Scholar] [CrossRef] [PubMed]
- Diederen, B.M.W. Legionella spp. and Legionnaires’ disease. J. Infect. 2008, 56, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Fliermans, C.B.; Cherry, W.B.; Orrison, L.H.; Smith, S.J.; Tison, D.L.; Pope, D.H. Ecological distribution of Legionella pneumophila. Appl. Environ. Microbiol. 1981, 41, 9–16. [Google Scholar]
- National Institute of Infectious Diseases and Tuberculosis and Infectious Diseases Control Division, Ministry of Health, Labour, and Welfare. Legionellosis, April 1999–July 2000; IASR (Infectious Agents Surveillance Report); IASR: Tokyo, Japan, 2000; pp. 186–187. [Google Scholar]
- Fitzhenry, R.; Weiss, D.; Cimini, D.; Balter, S.; Boyd, C.; Alleyne, L.; Stewart, R.; McIntosh, N.; Econome, A.; Lin, Y.; et al. Legionnaires’ Disease Outbreaks and Cooling Towers, New York City, NY, USA. Emerg. Infect. Dis. 2017, 23, 1769. [Google Scholar] [CrossRef] [PubMed]
- Bhopal, R.S.; Fallon, R.J. Variation in time and space of non-outbreak Legionnaires’ disease in Scotland. Epidemiol. Infect. 1991, 106, 45–61. [Google Scholar] [CrossRef] [PubMed]
- García-Fulgueiras, A.; Navarro, C.; Fenoll, D.; García, J.; González-Diego, P.; Jiménez-Buñuales, T.; Pelaz, C. Legionnaires’ disease outbreak in Murcia, Spain. Emerg. Infect. Dis. 2003, 9, 915–921. [Google Scholar] [CrossRef]
- Shivaji, T.; Sousa Pinto, C.; San-Bento, A.; Oliveira Serra, L.A.; Valente, J.; Machado, J.; Vasconcelos, P. A large community outbreak of Legionnaires disease in Vila Franca de Xira, Portugal, October to November 2014. Eurosurveillance 2014, 19, 20991. [Google Scholar] [CrossRef]
- Maisa, A.; Brockmann, A.; Renken, F.; Lück, C.; Pleischl, S.; Exner, M.; Daniels-Haardt, I.; Jurke, A. Epidemiological investigation and case-control study: A Legionnaires’ disease outbreak associated with cooling towers in Warstein, Germany, August–September 2013. Eurosurveillance 2015, 20, 30064. [Google Scholar] [CrossRef]
- Li, L.; Qin, T.; Li, Y.; Zhou, H.; Song, H.; Ren, H.; Li, L.; Li, Y.; Zhao, D. Prevalence and Molecular Characteristics of Waterborne Pathogen Legionella in Industrial Cooling Tower Environments. Int. J. Environ. Res. Public Health 2015, 12, 12605–12617. [Google Scholar] [CrossRef] [PubMed]
- Amemura-Maekawa, J.; Kikukawa, K.; Helbig, J.H.; Kaneko, S.; Suzuki-Hashimoto, A.; Furuhata, K.; Chang, B.; Murai, M.; Ichinose, M.; Ohnishi, M.; et al. Distribution of monoclonal antibody subgroups and sequence-based types among Legionella pneumophila serogroup 1 isolates derived from cooling tower water, bathwater, and soil in Japan. Appl. Environ. Microbiol. 2012, 78, 4263–4270. [Google Scholar] [CrossRef]
- Farhat, C.; Mentasti, M.; Jacobs, E.; Fry, N.K.; Lück, C. The N-acylneuraminate cytidyltransferase gene, neuA, is heterogenous in Legionella pneumophila strains but can be used as a marker for epidemiological typing in the consensus sequence-based typing scheme. J. Clin. Microbiol. 2011, 49, 4052–4058. [Google Scholar] [CrossRef] [PubMed]
- Gaia, V.; Fry, N.K.; Afshar, B.; Lück, P.C.; Meugnier, H.; Etienne, J.; Peduzzi, R.; Harrison, T.G. Consensus sequence-based scheme for epidemiological typing of clinical and environmental isolates of Legionella pneumophila. J. Clin. Microbiol. 2005, 43, 2047–2052. [Google Scholar] [CrossRef] [PubMed]
- Ratzow, S.; Gaia, V.; Helbig, J.H.; Fry, N.K.; Lück, P.C. Addition of neuA, the gene encoding N-acylneuraminate cytidylyl transferase, increases the discriminatory ability of the consensus sequence-based scheme for typing Legionella pneumophila serogroup 1 strains. J. Clin. Microbiol. 2007, 45, 1965–1968. [Google Scholar] [CrossRef]
- Coil, D.; Jospin, G.; Darling, A.E. A5-miseq: An updated pipeline to assemble microbial genomes from Illumina MiSeq data. Bioinformatics 2015, 31, 587–589. [Google Scholar] [CrossRef]
- Gardner, S.N.; Slezak, T.; Hall, B.G. kSNP3.0: SNP detection and phylogenetic analysis of genomes without genome alignment or reference genome. Bioinformatics 2015, 31, 2877–2878. [Google Scholar] [CrossRef]
- Richter, M.; Rosselló-Móra, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef] [Green Version]
- Goris, J.; Konstantinidis, K.T.; Klappenbach, J.A.; Coenye, T.; Vandamme, P.; Tiedje, J.M. DNA–DNA hybridization values and their relationship to whole-genome sequence similarities. Int. J. Syst. Evol. Microbiol. 2007, 57, 81–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuroki, T.; Ishihara, T.; Ito, K.; Kura, F. Bathwater-associated cases of legionellosis in Japan, with a special focus on Legionella concentrations in water. Jpn. J. Infect. Dis. 2009, 62, 201–205. [Google Scholar]
- Amemura-Maekawa, J.; Kura, F.; Chida, K.; Ohya, H.; Kanatani, J.I.; Isobe, J.; Tanaka, S.; Nakajima, H.; Hiratsuka, T.; Yoshino, S.; et al. Legionella pneumophila and Other Legionella Species Isolated from Legionellosis Patients in Japan between 2008 and 2016. Appl. Environ. Microbiol. 2018, 84, e00721-18. [Google Scholar] [CrossRef] [PubMed]
- Kuroki, T.; Amemura-Maekawa, J.; Ohya, H.; Furukawa, I.; Suzuki, M.; Masaoka, T.; Aikawa, K.; Hibi, K.; Morita, M.; Lee, K.I.; et al. Outbreak of Legionnaire’s Disease Caused by Legionella pneumophila Serogroups 1 and 13. Emerg. Infect. Dis. 2017, 23, 349–351. [Google Scholar] [CrossRef] [PubMed]
- Kanatani, J.I.; Isobe, J.; Norimoto, S.; Kimata, K.; Mitsui, C.; Amemura-Maekawa, J.; Kura, F.; Sata, T.; Watahiki, M. Prevalence of Legionella species isolated from shower water in public bath facilities in Toyama Prefecture, Japan. J. Infect. Chemother. 2017, 23, 265–270. [Google Scholar] [CrossRef]
- Amemura-Maekawa, J.; Kura, F.; Chang, B.; Suzuki-Hashimoto, A.; Ichinose, M.; Endo, T.; Watanabe, H. Distinct difference of flaA genotypes of Legionella pneumophila between isolates from bath water and cooling tower water. Microbiol. Immunol. 2008, 52, 460–464. [Google Scholar] [CrossRef]
- Qin, T.; Zhou, H.; Ren, H.; Guan, H.; Li, M.; Zhu, B.; Shao, Z. Distribution of sequence-based types of legionella pneumophila serogroup 1 strains isolated from cooling towers, hot springs, and potable water systems in China. Appl. Environ. Microbiol. 2014, 80, 2150–2157. [Google Scholar] [CrossRef]
- Lee, H.K.; Shim, J.I.; Kim, H.E.; Yu, J.Y.; Kang, Y.H. Distribution of Legionella species from environmental water sources of public facilities and genetic diversity of L. pneumophila serogroup 1 in South Korea. Appl. Environ. Microbiol. 2010, 76, 6547–6554. [Google Scholar] [CrossRef]
- Kozak-Muiznieks, N.A.; Lucas, C.E.; Brown, E.; Pondo, T.; Taylor, T.H., Jr.; Frace, M.; Miskowski, D.; Winchell, J.M. Prevalence of sequence types among clinical and environmental isolates of Legionella pneumophila serogroup 1 in the United States from 1982 to 2012. J. Clin. Microbiol. 2014, 52, 201–211. [Google Scholar] [CrossRef]
- Harrison, T.G.; Afshar, B.; Doshi, N.; Fry, N.K.; Lee, J.V. Distribution of Legionella pneumophila serogroups, monoclonal antibody subgroups and DNA sequence types in recent clinical and environmental isolates from England and Wales (2000–2008). Eur. J. Clin. Microbiol. Infect. Dis. 2009, 28, 781–791. [Google Scholar] [CrossRef] [PubMed]
- David, S.; Sánchez-Busó, L.; Harris, S.R.; Marttinen, P.; Rusniok, C.; Buchrieser, C.; Harrison, T.G.; Parkhill, J. Dynamics and impact of homologous recombination on the evolution of Legionella pneumophila. PLoS Genet. 2017, 13, e1006855. [Google Scholar] [CrossRef] [PubMed]
- Kozak, N.A.; Benson, R.F.; Brown, E.; Alexander, N.T.; Taylor, T.H., Jr.; Shelton, B.G.; Fields, B.S. Distribution of lag-1 alleles and sequence-based types among Legionella pneumophila serogroup 1 clinical and environmental isolates in the United States. J. Clin. Microbiol. 2009, 47, 2525–2535. [Google Scholar] [CrossRef]
- Kozak-Muiznieks, N.A.; Morrison, S.S.; Mercante, J.W.; Ishaq, M.K.; Johnson, T.; Caravas, J.; Lucas, C.E.; Brown, E.; Raphael, B.H.; Winchell, J.M. Comparative genome analysis reveals a complex population structure of Legionella pneumophila subspecies. Infect. Genet. Evol. 2018, 59, 172–185. [Google Scholar] [CrossRef] [PubMed]
- Raphael, B.H.; Baker, D.J.; Nazarian, E.; Lapierre, P.; Bopp, D.; Kozak-Muiznieks, N.A.; Morrison, S.S.; Lucas, C.E.; Mercante, J.W.; Musser, K.A.; et al. Genomic resolution of outbreak-associated Legionella pneumophila serogroup 1 isolates from New York State. Appl. Environ. Microbiol. 2016, 82, 3582–3590. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| SG | No. of Isolates (%) | STs (No. of Isolates) | |
|---|---|---|---|
| SG1 | 114 | (70.8) | ST1 (88), ST154 (15), ST1008 (3), ST598 (2), ST2 (1), ST40 (1), ST45 (1), ST59 (1), ST607 (1), ST1065 (1) |
| SG13 | 18 | (11.2) | ST2603 (15), ST2256 (2), ST2250 (1) |
| SG7 | 13 | (8.1) | ST715 (13) |
| SG UT | 7 | (4.3) | ST1334 (2), ST2699 * (2), ST172 (1), ST1916 (1), ST2701 * (1) |
| SG5 | 4 | (2.5) | ST2700 * (2), ST2704 * (2) |
| SG6 | 2 | (1.2) | ST68 (1), ST242 (1) |
| SG2 | 1 | (0.6) | ST2702 * (1) |
| SG8 | 1 | (0.6) | ST1324 (1) |
| SG9 | 1 | (0.6) | ST2703 * (1) |
| Complexes | No. of Strain | Years | Site in CT a | SG | ST | DDBJ Accession No. of Read Data |
|---|---|---|---|---|---|---|
| CC1 | KL287 | 2006 | F | 1 | 1 | DRR163670 |
| KL305 | 2006 | A | 1 | 1 | DRR163671 | |
| KL464 | 2008 | A | 1 | 1 | DRR163673 | |
| KL497 | 2008 | E | 1 | 1 | DRR163675 | |
| KL579 | 2009 | H | 1 | 1 | DRR163677 | |
| KL649 | 2010 | A | 1 | 1 | DRR163679 | |
| KL682 | 2010 | M | 1 | 1008 | DRR170776 | |
| KL684 | 2010 | D | 1 | 1 | DRR163680 | |
| KL736 | 2011 | J | 1 | 1 | DRR163683 | |
| KL819 | 2012 | Q | 7 | 715 | DRR170777 | |
| CC2 | KL008 | 2003 | K | UT | 1334 | DRR170771 |
| KL074 | 2004 | L | 1 | 607 | DRR170772 | |
| KL372 | 2007 | C | 1 | 154 | DRR163672 | |
| KL490 | 2008 | N | UT | 1334 | DRR170773 | |
| KL548 | 2009 | O | 1 | 598 | DRR170774 | |
| KL603 | 2009 | I | 1 | 154 | DRR163678 | |
| KL679 | 2010 | P | 1 | 1065 | DRR170775 | |
| KL747 | 2011 | G | 1 | 154 | DRR163684 | |
| KL852 b | 2012 | L | 9 | 2703 | DRR170778 | |
| CC3 | KL209 | 2005 | B | 13 | 2603 | DRR163668 |
| KL285 | 2006 | F | UT | 2699 | DRR163669 | |
| KL467 | 2008 | A | 13 | 2603 | DRR163674 | |
| KL578 | 2009 | H | 13 | 2603 | DRR163676 | |
| KL685 | 2010 | D | 13 | 2603 | DRR163681 | |
| KL735 | 2011 | J | 13 | 2603 | DRR163682 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakanishi, N.; Nomoto, R.; Tanaka, S.; Arikawa, K.; Iwamoto, T. Analysis of Genetic Characterization and Clonality of Legionella pneumophila Isolated from Cooling Towers in Japan. Int. J. Environ. Res. Public Health 2019, 16, 1664. https://doi.org/10.3390/ijerph16091664
Nakanishi N, Nomoto R, Tanaka S, Arikawa K, Iwamoto T. Analysis of Genetic Characterization and Clonality of Legionella pneumophila Isolated from Cooling Towers in Japan. International Journal of Environmental Research and Public Health. 2019; 16(9):1664. https://doi.org/10.3390/ijerph16091664
Chicago/Turabian StyleNakanishi, Noriko, Ryohei Nomoto, Shinobu Tanaka, Kentaro Arikawa, and Tomotada Iwamoto. 2019. "Analysis of Genetic Characterization and Clonality of Legionella pneumophila Isolated from Cooling Towers in Japan" International Journal of Environmental Research and Public Health 16, no. 9: 1664. https://doi.org/10.3390/ijerph16091664
APA StyleNakanishi, N., Nomoto, R., Tanaka, S., Arikawa, K., & Iwamoto, T. (2019). Analysis of Genetic Characterization and Clonality of Legionella pneumophila Isolated from Cooling Towers in Japan. International Journal of Environmental Research and Public Health, 16(9), 1664. https://doi.org/10.3390/ijerph16091664