Extended Phylogeny and Extraintestinal Virulence Potential of Commensal Escherichia coli from Piglets and Sows



Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection, Isolation, and Identification of Escherichia Coli (E. coli)
2.2. Extended Phylogenetic Grouping
2.3. Virulence Genes (VGs) and Fimbrial Operons Genotyping
2.4. Statistical Analysis
3. Results
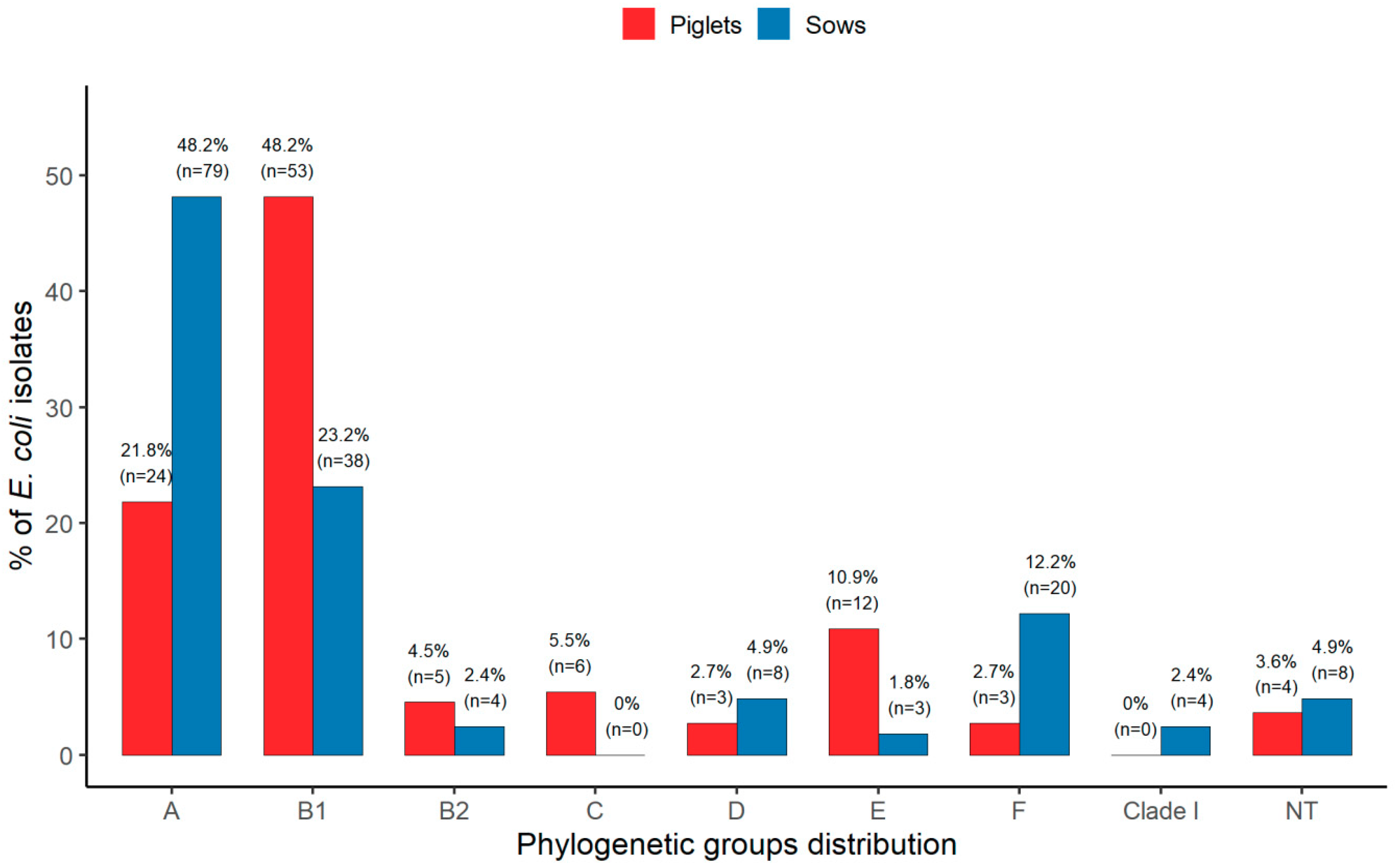
3.1. Extended Phylogenetic Structure of E. coli
3.2. Prevalence of VGs
3.3. Distribution of VGs according to Phylogenetic Groups
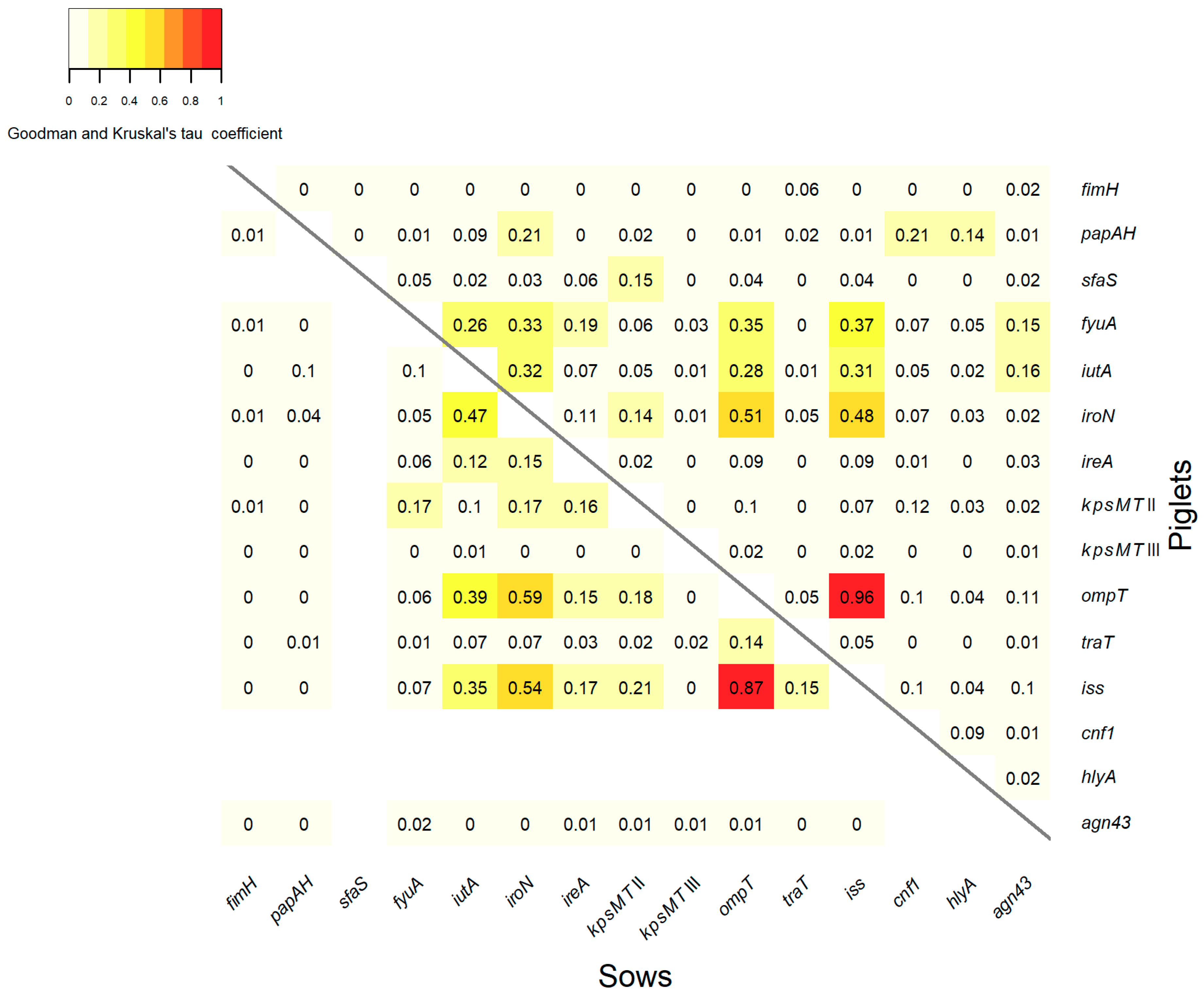
3.4. Association between VGs
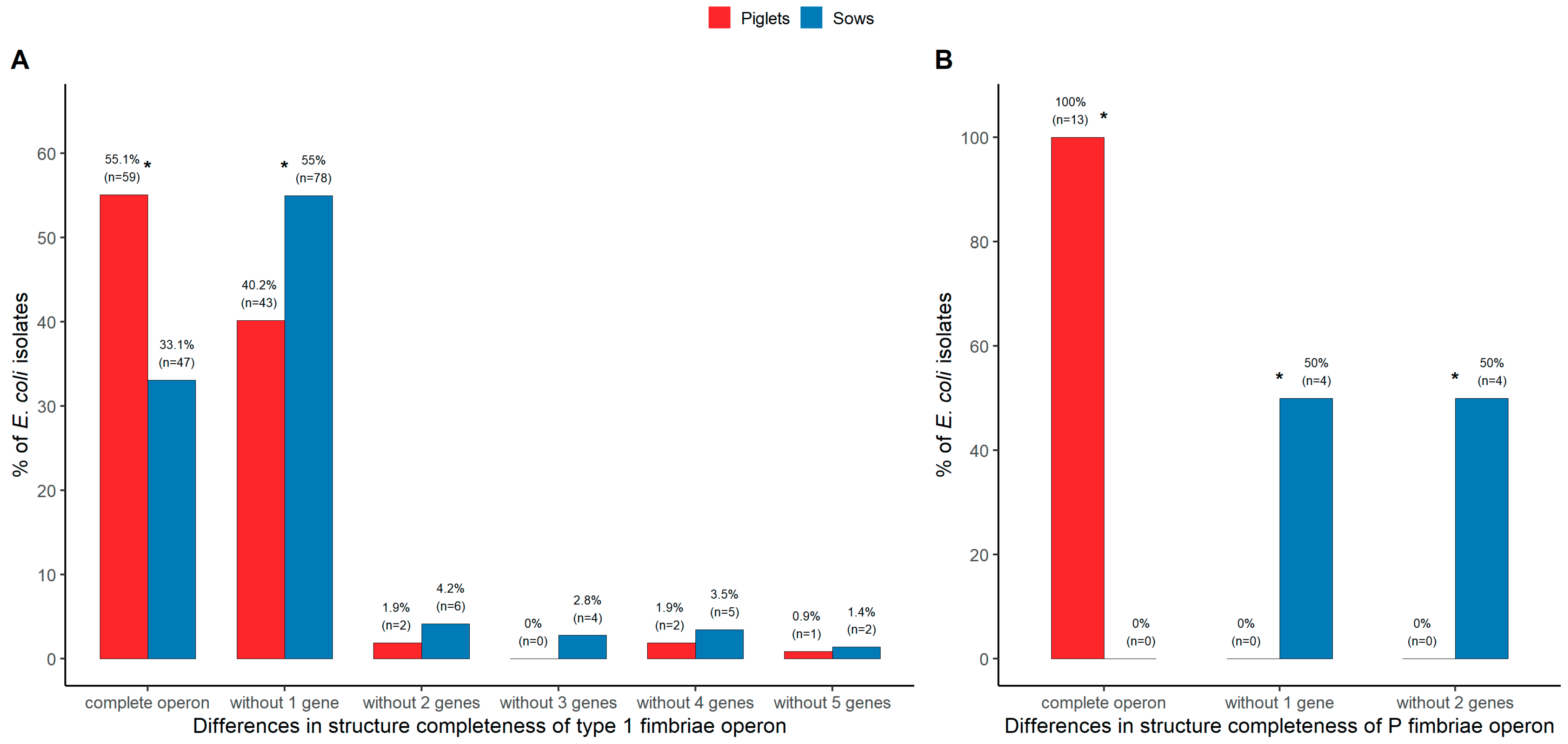
3.5. Genotypic Analysis of Fimbrial Operons for Type 1 and P Fimbriae
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tenaillon, O.; Skurnik, D.; Picard, B.; Denamur, E. The population genetics of commensal Escherichia coli. Nat. Rev. Microbiol. 2010, 8, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Köhler, C.D.; Dobrindt, U. What defines extraintestinal pathogenic Escherichia coli? Int. J. Med. Microbiol. 2011, 301, 642–647. [Google Scholar] [CrossRef] [PubMed]
- Starčič, E.M.; Žgur-Bertok, D. Virulence potential for extraintestinal infections among commensal Escherichia coli isolated from healthy humans—The Trojan horse within our gut. FEMS Microbiol. Lett. 2015, 362, fnu061. [Google Scholar]
- Johnson, J.R.; Russo, T.A. Molecular epidemiology of extraintestinal pathogenic (uropathogenic) Escherichia coli. Int. J. Med. Microbiol. 2005, 295, 383–404. [Google Scholar] [CrossRef]
- Bélanger, L.; Garenaux, A.; Harel, J.; Boulianne, M.; Nadeau, E.; Dozois, C.M. Escherichia coli from animal reservoirs as a potential source of human extraintestinal pathogenic E. coli. FEMS Immunol. Med. Microbiol. 2011, 62, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Tan, C.; Tang, X.; Zhang, X.; Ding, Y.; Zhao, Z.; Wu, B.; Cai, X.; Liu, Z.; He, Q.; Chen, H. Serotypes and virulence genes of extraintestinal pathogenic Escherichia coli isolates from diseased pigs in China. Vet. J. 2012, 192, 483–488. [Google Scholar] [CrossRef]
- Picard, B.; Garcia, J.S.; Gouriou, S.; Duriez, P.; Brahimi, N.; Bingen, E.; Elion, J.; Denamur, E. The link between phylogeny and virulence in Escherichia coli extraintestinal infection. Infect. Immun. 1999, 67, 546–553. [Google Scholar] [CrossRef] [Green Version]
- Sarowska, J.; Futoma-Koloch, B.; Jama-Kmiecik, A.; Frej-Madrzak, M.; Ksiazczyk, M.; Bugla-Ploskonska, G.; Choroszy-Krol, I. Virulence factors, prevalence and potential transmission of extraintestinal pathogenic Escherichia coli isolated from different sources: Recent reports. Gut Pathog. 2019, 11, 10. [Google Scholar] [CrossRef] [Green Version]
- Wasiński, B. Extra-intestinal pathogenic Escherichia coli–threat connected with food-borne infections. Ann. Agric. Environ. Med. 2019. [Google Scholar] [CrossRef]
- Johnson, J.R.; Murray, A.C.; Gajewski, A.; Sullivan, M.; Snippes, P.; Kuskowski, M.A.; Smith, K.E. Isolation and molecular characterization of nalidixic acid-resistant extraintestinal pathogenic Escherichia coli from retail chicken products. Antimicrob. Agents Chemother. 2003, 47, 2161–2168. [Google Scholar] [CrossRef] [Green Version]
- Venturini, C.; Hassan, K.A.; Chowdhury, P.R.; Paulsen, I.T.; Walker, M.J.; Djordjevic, S.P. Sequences of two related multiple antibiotic resistance virulence plasmids sharing a unique IS26-related molecular signature isolated from different Escherichia coli pathotypes from different hosts. PLoS ONE 2013, 8, e78862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reid, C.J.; Wyrsch, E.R.; Chowdhury, P.R.; Zingali, T.; Liu, M.; Darling, A.E.; Chapman, T.A.; Djordjevic, S.P. Porcine commensal Escherichia coli: A reservoir for class 1 integrons associated with IS26. Microb. Genom. 2017, 3, 12. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.R.; Oswald, E.; O’Bryan, T.T.; Kuskowski, M.A.; Spanjaard, L. Phylogenetic distribution of virulence-associated genes among Escherichia coli isolates associated with neonatal bacterial meningitis in the Netherlands. J. Infect. Dis. 2002, 185, 774–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowrouzian, F.L.; Wold, A.E.; Adlerberth, I. Escherichia coli strains belonging to phylogenetic group B2 have superior capacity to persist in the intestinal microflora of infants. J. Infect. Dis. 2005, 191, 1078–1083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clermont, O.; Christenson, J.K.; Denamur, E.; Gordon, D.M. The Clermont Escherichia coli phylo-typing method revisited: Improvement of specificity and detection of new phylo-groups. Environ. Microbiol. Rep. 2013, 5, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Escobar-Páramo, P.; Le Menac’h, A.; Le Gall, T.; Amorin, C.; Gouriou, S.; Picard, B.; Skurnik, D.; Denamur, E. Identification of forces shaping the commensal Escherichia coli genetic structure by comparing animal and human isolates. Environ Microbiol. 2006, 8, 1975–1984. [Google Scholar] [CrossRef]
- Gordon, D.M.; Cowling, A. The distribution and genetic structure of Escherichia coli in Australian vertebrates: Host and geographic effects. Microbiology 2003, 149, 3575–3586. [Google Scholar] [CrossRef] [Green Version]
- Carlos, C.; Pires, M.M.; Stoppe, N.C.; Hachich, E.M.; Sato, M.I.; Gomes, T.A.; Amaral, L.A.; Ottoboni, L.M. Escherichia coli phylogenetic group determination and its application in the identification of the major animal source of fecal contamination. BMC Microbiol. 2010, 10, 161. [Google Scholar] [CrossRef] [Green Version]
- Cortés, P.; Blanc, V.; Mora, A.; Dahbi, G.; Blanco, J.E.; Blanco, M.; López, C.; Andreu, A.; Navarro, F.; Alonso, M.P.; et al. Isolation and characterization of potentially pathogenic antimicrobial-resistant Escherichia coli strains from chicken and pig farms in Spain. Appl. Environ. Microbiol. 2010, 76, 2799–2805. [Google Scholar] [CrossRef] [Green Version]
- Dixit, S.M.; Gordon, D.M.; Wu, X.Y.; Chapman, T.; Kailasapathy, K.; Chin, J.J. Diversity analysis of commensal porcine Escherichia coli–associations between genotypes and habitat in the porcine gastrointestinal tract. Microbiology 2004, 150, 1735–1740. [Google Scholar] [CrossRef] [Green Version]
- Bok, E.; Mazurek, J.; Pusz, P.; Stosik, M.; Baldy-Chudzik, K. Age as a factor influencing diversity of commensal E. coli microflora in pigs. Pol. J. Microbiol. 2013, 62, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Clermont, O.; Bonacorsi, S.; Bingen, E. Rapid and simple determination of the Escherichia coli phylogenetic group. Appl. Environ. Microbiol. 2000, 66, 4555–4558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logue, C.M.; Wannemuehler, Y.; Nicholson, B.A.; Doetkott, C.; Barbieri, N.L.; Nolan, L.K. Comparative analysis of phylogenetic assignment of human and avian ExPEC and fecal commensal Escherichia coli using the (previous and revised) Clermont phylogenetic typing methods and its impact on avian pathogenic Escherichia coli (APEC) classification. Front. Microbiol. 2017, 8, 283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, T.A.; Johnson, J.R. Medical and economic impact of extraintestinal infections due to Escherichia coli: Focus on an increasingly important endemic problem. Microbes Infect. 2003, 5, 449–456. [Google Scholar] [CrossRef]
- Fairbrother, J.M.; Gyles, C. Colibacillosis. In Diseases of Swine, 10th ed.; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2012; pp. 723–749. [Google Scholar]
- Zhu, Y.; Dong, W.; Ma, J.; Yuan, L.; Hejair, H.M.; Pan, Z.; Liu, G.; Yao, H. Characterization and virulence clustering analysis of extraintestinal pathogenic Escherichia coli isolated from swine in China. BMC Vet. Res. 2017, 13, 94. [Google Scholar] [CrossRef] [Green Version]
- Spindola, M.G.; Cunha, M.P.V.; Moreno, L.Z.; Amigo, C.R.; Silva, A.P.S.; Parra, B.M.; Poor, A.P.; de Oliveira, C.H.; Perez, B.P.; Knöbl, T.; et al. Genetic diversity, virulence genotype and antimicrobial resistance of uropathogenic Escherichia coli (UPEC) isolated from sows. Vet. Q. 2018, 38, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Zong, B.; Zhang, Y.; Wang, X.; Liu, M.; Zhang, T.; Zhu, Y.; Zheng, Y.; Hu, L.; Li, P.; Chen, H.; et al. Characterization of multiple type-VI secretion system (T6SS) VgrG proteins in the pathogenicity and antibacterial activity of porcine extra-intestinal pathogenic Escherichia coli. Virulence 2019, 10, 118–132. [Google Scholar] [CrossRef] [Green Version]
- Jakobsen, L.; Spangholm, D.J.; Pedersen, K.; Jensen, L.B.; Emborg, H.D.; Agersø, Y.; Aarestrup, F.M.; Hammerum, A.M.; Frimodt-Møller, N. Broiler chickens, broiler chicken meat, pigs and pork as sources of ExPEC related virulence genes and resistance in Escherichia coli isolates from community-dwelling humans and UTI patients. Int. J. Food Microbiol. 2010, 142, 264–272. [Google Scholar] [CrossRef]
- Manges, A.R.; Johnson, J.R. Reservoirs of extraintestinal pathogenic Escherichia coli. Microbiol. Spectrum 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Mellata, M. Human and avian extraintestinal pathogenic Escherichia coli: Infections, zoonotic risks, and antibiotic resistance trends. Foodborne Pathog. Dis. 2013, 10, 916–932. [Google Scholar] [CrossRef] [Green Version]
- Tan, C.; Xu, Z.; Zheng, H.; Liu, W.; Tang, X.; Shou, J.; Wu, B.; Wang, S.; Zhao, G.P.; Chen, H. Genome sequence of a porcine extraintestinal pathogenic Escherichia coli strain. J. Bacteriol. 2011, 193, 5038. [Google Scholar] [CrossRef] [Green Version]
- Manges, A.R.; Harel, J.; Masson, L.; Edens, T.J.; Portt, A.; Reid-Smith, R.J.; Zhanel, G.G.; Kropinski, A.M.; Boerlin, P. Multilocus sequence typing and virulence gene profiles associated with Escherichia coli from human and animal sources. Foodborne Pathog. Dis. 2015, 12, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Reid, C.J.; DeMaere, M.Z.; Djordjevic, S.P. Australian porcine clonal complex 10 (CC10) Escherichia coli belong to multiple sublineages of a highly diverse global CC10 phylogeny. Microb. Genom. 2019, 5, e000225. [Google Scholar] [CrossRef] [PubMed]
- Lescat, M.; Clermont, O.; Woerther, P.L.; Glodt, J.; Dion, S.; Skurnik, D.; Djossou, F.; Dupont, C.; Perroz, G.; Picard, B.; et al. Commensal Escherichia coli strains in Guiana reveal a high genetic diversity with host-dependant population structure. Environ. Microbiol. Rep. 2013, 5, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Clermont, O.; Lescat, M.; O’Brien, C.L.; Gordon, D.M.; Tenaillon, O.; Denamur, E. Evidence for a human-specific Escherichia coli clone. Environ. Microbiol. 2008, 10, 1000–1006. [Google Scholar]
- Clermont, O.; Gordon, D.M.; Brisse, S.; Walk, S.T.; Denamur, E. Characterization of the cryptic Escherichia lineages: Rapid identification and prevalence. Environ. Microbiol. 2011, 13, 2468–2477. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.R.; Stell, A.L. Extended virulence genotypes of Escherichia coli strains from patients with urosepsis in relation to phylogeny and host compromise. J. Infect. Dis. 2000, 181, 261–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.R.; O’Bryan, T.T. Detection of the Escherichia coli group 2 polysaccharide capsule synthesis gene kpsM by a rapid and specific PCR-based assay. J. Clin. Microbiol. 2004, 42, 1773–1776. [Google Scholar] [CrossRef] [Green Version]
- Mellata, M.; Ameiss, K.; Mo, H.; Curtiss, R. Characterization of the contribution to virulence of three large plasmids of avian pathogenic Escherichia coli chi7122 (O78:K80:H9). Infect. Immun. 2010, 78, 1528–1541. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, S.; Terai, A.; Yuri, K.; Kurazono, H.; Takeda, Y.; Yoshida, O. Detection of urovirulence factors in Escherichia coli by multiplex polymerase chain reaction. FEMS Immunol. Med. Microbiol. 1995, 12, 85–90. [Google Scholar] [CrossRef]
- Restieri, C.; Garriss, G.; Locas, M.C.; Dozois, C.M. Autotransporter-encoding sequences are phylogenetically distributed among Escherichia coli clinical isolates and reference strains. Appl. Environ. Microbiol. 2007, 73, 1553–1562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blumer, C.; Kleefeld, A.; Lehnen, D.; Heintz, M.; Dobrindt, U.; Nagy, G.; Michaelis, K.; Emödy, L.; Polen, T.; Rachel, R.; et al. Regulation of type 1 fimbriae synthesis and biofilm formation by the transcriptional regulator LrhA of Escherichia coli. Microbiology 2005, 151, 3287–3298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwan, W.R.; Lee, J.L.; Lenard, F.A.; Matthews, B.T.; Beck, M.T. Osmolarity and pH growth conditions regulate fim gene transcription and type 1 pilus expression in uropathogenic Escherichia coli. Infect. Immun. 2002, 70, 1391–1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewers, C.; Li, G.; Wilking, H.; Kiessling, S.; Alt, K.; Antáo, E.M.; Laturnus, C.; Diehl, I.; Glodde, S.; Homeier, T.; et al. Avian pathogenic, uropathogenic, and newborn meningitis-causing Escherichia coli: How closely related are they? Int. J. Med. Microbiol. 2007, 297, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Chapman, T.A.; Wu, X.Y.; Barchia, I.; Bettelheim, K.A.; Driesen, S.; Trott, D.; Wilson, M.; Chin, J.J. Comparison of virulence gene profiles of Escherichia coli strains isolated from healthy and diarrheic swine. Appl. Environ. Microbiol. 2006, 72, 4782–4795. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Statist. Soc. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Lindstrom, M.J.; Bates, D.M. Nonlinear mixed effects models for repeated measures data. Biometrics 1990, 46, 673–687. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing. 2019. Available online: https://repo.bppt.go.id/cran/web/packages/dplR/vignettes/intro-dplR.pdf (accessed on 31 October 2019).
- Spencer, B.T.; Howell, P.G. Some husbandry factors influencing weaning stresses in piglets. J. S. Afr. Vet. Assoc. 1989, 60, 62–64. [Google Scholar]
- Bibbal, D.; Dupouy, V.; Prère, M.F.; Toutain, P.L.; Bousquet-Mélou, A. Relatedness of Escherichia coli strains with different susceptibility phenotypes isolated from swine feces during ampicillin treatment. Appl. Environ. Microbiol. 2009, 75, 2999–3006. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Zheng, H.; Yang, M.; Xu, Z.; Wang, X.; Wei, L.; Tang, B.; Liu, F.; Zhang, Y.; Ding, Y.; et al. Genome analysis and in vivo virulence of porcine extraintestinal pathogenic Escherichia coli strain PCN033. BMC Genom. 2016, 16, 717. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Tang, X.; Lu, P.; Wu, B.; Xu, Z.; Liu, W.; Zhang, R.; Bei, W.; Chen, H.; Tan, C. Clonal analysis and virulent traits of pathogenic extraintestinal Escherichia coli isolates from swine in China. BMC Vet. Res. 2012, 8, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dezfulian, H.; Batisson, I.; Fairbrother, J.M.; Lau, P.C.; Nassar, A.; Szatmari, G.; Harel, J. Presence and characterization of extraintestinal pathogenic Escherichia coli virulence genes in F165-positive E. coli strains isolated from diseased calves and pigs. J. Clin. Microbiol. 2003, 41, 1375–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maynard, C.; Bekal, S.; Sanschagrin, F.; Levesque, R.C.; Brousseau, R.; Masson, L.; Lariviere, S.; Harel, J. Heterogeneity among virulence and antimicrobial resistance gene profiles of extraintestinal Escherichia coli isolates of animal and human origin. J. Clin. Microbiol. 2004, 42, 5444–5452. [Google Scholar] [CrossRef] [Green Version]
- Katouli, M.; Lund, A.; Wallgren, P.; Kuhn, I.; Soderlind, O.; Mollby, R. Phenotypic characterization of intestinal Escherichia coli of pigs during suckling, postweaning, and fattening periods. Appl. Environ. Microbiol. 1995, 61, 778–783. [Google Scholar]
- Ahmed, S.; Olsen, J.E.; Herrero-Fresno, A. The genetic diversity of commensal Escherichia coli strains isolated from non-antimicrobial treated pigs varies according to age group. PLoS ONE 2017, 12, e0178623. [Google Scholar] [CrossRef] [PubMed]
- Nowrouzian, F.L.; Adlerberth, I.; Wold, A.E. Enhanced persistence in the colonic microbiota of Escherichia coli strains belonging to phylogenetic group B2: Role of virulence factors and adherence to colonic cells. Microbes Infect. 2006, 8, 834–840. [Google Scholar] [CrossRef]
- Johnson, J.R.; Clabots, C.; Kuskowski, M.A. Multiple-host sharing, long-term persistence, and virulence of Escherichia coli clones from human and animal household members. J. Clin. Microbiol. 2008, 46, 4078–4082. [Google Scholar] [CrossRef] [Green Version]
- Schierack, P.; Walk, N.; Ewers, C.; Wilking, H.; Steinrück, H.; Filter, M.; Wieler, L.H. ExPEC-typical virulence-associated genes correlate with successful colonization by intestinal E. coli in a small piglet group. Environ. Microbiol. 2008, 10, 1742–1751. [Google Scholar] [CrossRef]
- Lemaître, C.; Mahjoub-Messai, F.; Dupont, D.; Caro, V.; Diancourt, L.; Bingen, E.; Bidet, P.; Bonacorsi, S. A conserved virulence plasmidic region contributes to the virulence of the multiresistant Escherichia coli meningitis strain S286 belonging to phylogenetic group C. PLoS ONE 2013, 8, e74423. [Google Scholar] [CrossRef]
- Lüthje, P.; Brauner, A. Virulence factors of uropathogenic E. coli and their interaction with the host. Adv. Microb. Physiol. 2014, 65, 337–372. [Google Scholar]
- Pusz, P.; Bok, E.; Mazurek, J.; Stosik, M.; Baldy-Chudzik, K. Type 1 fimbriae in commensal Escherichia coli derived from healthy humans. Acta Biochim. Pol. 2014, 61, 389–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, M.C.; Mobley, H.L.T. Role of P-fimbrial-mediated adherence in pyelonephritis and persistence of uropathogenic Escherichia coli (UPEC) in the mammalian kidney. Kidney Int. 2007, 72, 19–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Functional Category VG | Number (%) of E. coli Isolates with VGs | p-Value | |
|---|---|---|---|
| Piglets | Sows | ||
| N = 110 | N = 164 | ||
| Adhesins | |||
| fimH | 107 (97.3) | 142 (86.6) | 0.0065 * |
| papAH | 13 (11.8) | 8 (4.9) | 0.0449 * |
| sfaS | 2 (1.8) | 0 | - |
| Iron acquisition | |||
| fyuA | 29 (26.4) | 23 (14.0) | 0.0931 |
| iutA | 56 (50.9) | 33 (20.1) | <0.0001 * |
| iroN | 43 (39.1) | 29 (17.7) | 0.0012 * |
| ireA | 7 (6.4) | 5 (3.0) | 0.2313 |
| Protectins | |||
| kpsMTII: | 12 (10.9) | 10 (6.1) | 0.2867 |
| -K1 | 4 (3.6) | 9 (5.5) | 0.5026 |
| -K2 | 8 (7.3) | 9 (5.5) | 0.7091 |
| -K5 | 7 (6.4) | 10 (6.1) | 0.9264 |
| kpsMTIII | 1 (0.9) | 2 (1.2) | 0.9971 |
| ompT | 36 (32.7) | 28 (17.1) | 0.0175 * |
| traT | 94 (85.5) | 90 (54.9) | <0.0001 * |
| iss | 35 (31.8) | 25 (15.2) | 0.0143 * |
| Toxins | |||
| cnf1 | 5 (4.5) | 0 | - |
| hlyA | 2 (1.8) | 0 | - |
| Biofilm formation | |||
| agn43: | 59 (53.6) | 87 (53.0) | 0.9275 |
| -a | 5 (4.5) | 3 (1.8) | 0.4890 |
| -b | 2 (1.8) | 2 (1.2) | 0.6874 |
| VG | Number (%) of E. coli Isolates with VGs within Phylogenetic Groups | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Piglets | Sows | |||||||||||||||
| A | B1 | B2 | C | D | E | F | NT | A | B1 | B2 | D | E | F | Clade I | NT | |
| n = 24 | n = 53 | n = 5 | n = 6 | n = 3 | n = 12 | n = 3 | n = 4 | n = 79 | n = 38 | n = 4 | n = 8 | n = 3 | n = 20 | n = 4 | n = 8 | |
| Adhesins | ||||||||||||||||
| fimH | 23 (95.8) | 51 (96.2) | 5 (100) | 6 (100) | 3 (100) | 12 (100) | 3 (100) | 4 (100) | 77 (97.5) | 38 (100) | 2 (50.0) | 5 (62.5) | 3 (100) | 5 (25.0) | 4 (100) | 8 (100) |
| papA | 0 | 9 (17.0) | 4 (80.0) | 0 | 0 | 0 | 0 | 0 | 0 | 4 (10.5) | 1 (25.0) | 1 (12.5) | 0 | 2 (10.0) | 0 | 0 |
| sfaS | 1 (4.2) | 0 | 1 (20.0) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Iron acquisition | ||||||||||||||||
| fyuA | 9 (37.5) | 8 (15.1) | 5 (100) | 6 (100) | 0 | 0 | 1 (33.3) | 0 | 11 (13.9) | 6 (15.8) | 2 (50.0) | 0 | 1 (33.3) | 0 | 0 | 3 (37.5) |
| iutA | 13 (54.2) | 23 (43.4) | 5 (100) | 6 (100) | 2 (66.7) | 4 (33.3) | 3 (100) | 0 | 8 (10.1) | 9 (23.7) | 4 (100) | 2 (25.0) | 0 | 7 (35.0) | 0 | 3 (37.5) |
| iroN | 9 (37.5) | 18 (34.0) | 5 (100) | 6 (100) | 1 (33.3) | 0 | 3 (100) | 1 (25.0) | 7 (8.9) | 13 (34.2) | 1 (25.0) | 0 | 0 | 5 (25.0) | 0 | 3 (37.5) |
| ireA | 3 (12.5) | 2 (3.8) | 1 (20.0) | 1 (16.7) | 0 | 0 | 0 | 0 | 2 (2.5) | 1 (2.6) | 0 | 0 | 0 | 1 | 0 | 1 (12.5) |
| Protectins | ||||||||||||||||
| kpsMTII | 2 (8.3) | 1 (1.9) | 4 (80.0) | 0 | 1 (33.3) | 1 (8.3) | 2 (66.7) | 1 (25.0) | 2 (2.5) | 3 (7.9) | 0 | 1 (12.5) | 0 | 1 (5.0) | 0 | 3 (37.5) |
| kpsMTIII | 0 | 1 (1.9) | 0 | 0 | 0 | 0 | 0 | 0 | 2 (2.5) | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| ompT | 6 (25.0) | 14 (26.4) | 5 (100) | 6 (100) | 2 (66.7) | 0 | 3 (100) | 0 | 7 (8.9) | 11 (28.9) | 0 | 0 | 0 | 7 (35.0) | 0 | 3 (37.5) |
| traT | 19 (79.2) | 49 (92.5) | 5 (100) | 5 (83.3) | 2 (66.7) | 9 (75.0) | 3 (100) | 2 (50.0) | 36 (45.6) | 25 (65.8) | 2 (50.0) | 5 (62.5) | 2 (66.7) | 13 (65.0) | 1 (25.0) | 6 (75.0) |
| iss | 6 (25.0) | 14 (26.4) | 5 (100) | 6 (100) | 1 (33.3) | 0 | 3 (100) | 0 | 7 (8.9) | 9 (23.7) | 0 | 0 | 0 | 6 (30.0) | 0 | 3 (37.5) |
| Toxins | ||||||||||||||||
| cnf1 | 0 | 1 (1.9) | 3 (60.0) | 1 (16.7) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| hlyA | 0 | 1 (1.9) | 1 (20.0) | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Biofilm formation | ||||||||||||||||
| agn43 | 20 (83.3) | 19 (35.8) | 4 (80.0) | 5 (83.3) | 2 (66.7) | 5 (41.7) | 2 (66.7) | 2 (50.0) | 41 (51.9) | 19 (50.0) | 3 (75.0) | 3 (37.5) | 2 (66.7) | 10 (50.0) | 2 (50.0) | 7 (87.5) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bok, E.; Kożańska, A.; Mazurek-Popczyk, J.; Wojciech, M.; Baldy-Chudzik, K. Extended Phylogeny and Extraintestinal Virulence Potential of Commensal Escherichia coli from Piglets and Sows. Int. J. Environ. Res. Public Health 2020, 17, 366. https://doi.org/10.3390/ijerph17010366
Bok E, Kożańska A, Mazurek-Popczyk J, Wojciech M, Baldy-Chudzik K. Extended Phylogeny and Extraintestinal Virulence Potential of Commensal Escherichia coli from Piglets and Sows. International Journal of Environmental Research and Public Health. 2020; 17(1):366. https://doi.org/10.3390/ijerph17010366
Chicago/Turabian StyleBok, Ewa, Aleksandra Kożańska, Justyna Mazurek-Popczyk, Magdalena Wojciech, and Katarzyna Baldy-Chudzik. 2020. "Extended Phylogeny and Extraintestinal Virulence Potential of Commensal Escherichia coli from Piglets and Sows" International Journal of Environmental Research and Public Health 17, no. 1: 366. https://doi.org/10.3390/ijerph17010366
APA StyleBok, E., Kożańska, A., Mazurek-Popczyk, J., Wojciech, M., & Baldy-Chudzik, K. (2020). Extended Phylogeny and Extraintestinal Virulence Potential of Commensal Escherichia coli from Piglets and Sows. International Journal of Environmental Research and Public Health, 17(1), 366. https://doi.org/10.3390/ijerph17010366