Fungicide-Tolerant Plant Growth-Promoting Rhizobacteria Mitigate Physiological Disruption of White Radish Caused by Fungicides Used in the Field Cultivation

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Toxicity of Fungicides to R. sativus In Vitro
Seed Germination and Plant Growth
2.2. Isolation of Rhizobacteria
2.3. Selection of Fungicide-Tolerant PGPR
2.4. Morphological, Biochemical and Molecular Identification of PGPR Isolates
2.5. Production of Plant Growth-Promoting Substances Under Fungicidal Stress
2.5.1. Indole-3-Acetic-Acid (IAA) and Siderophore Production
2.5.2. Phosphate Solubilization
2.5.3. Hydrogen Cyanide (HCN) and Ammonia Production
2.6. Assessment of Fungicidal Toxicity to PGPR Isolates
2.6.1. Morphological Distortion Induced by Fungicides Observed by Scanning Electron Microscope (SEM)
2.6.2. Assessment of Membrane Integrity by Confocal Laser Scanning Microscope (CLSM)
2.7. Crop-Based Experiments
2.7.1. Fungicide Application, Seed Treatment and Plant Culturing
2.7.2. Measurement of Growth Attributes
2.7.3. Effect of PGPR Inoculants on Photosynthetic Pigments
2.7.4. Determination of Total Soluble Protein
2.8. Assessment of Stressor Molecules and Antioxidant Enzymes in Radish
2.8.1. Proline Estimation
2.8.2. Lipid Peroxidation (MDA content)
2.8.3. Extraction and Determination of Antioxidant Enzymes
2.9. Statistical Analysis
3. Results and Discussion
3.1. Germination, Vigor Index and Biological Attributes of R. Sativus Under Fungicide-Stressed Conditions
3.2. Biochemical Characterization, Identification and Fungicide Tolerance
3.3. Plant Growth-Promoting Activities of PGPR Isolates under Fungicide-Stress
3.3.1. IAA and Siderophores
3.3.2. Phosphate Solubilization, Cyanogenic Compounds and Ammonia Production
3.4. Assessment of Cell Morphology and Permeability
3.5. Radish-Fungicide-PGPR Interactions
3.5.1. Performance of R. sativus under Fungicide-Stressed Conditions
Plant Growth and Elongation
Dry Biomass Accumulation
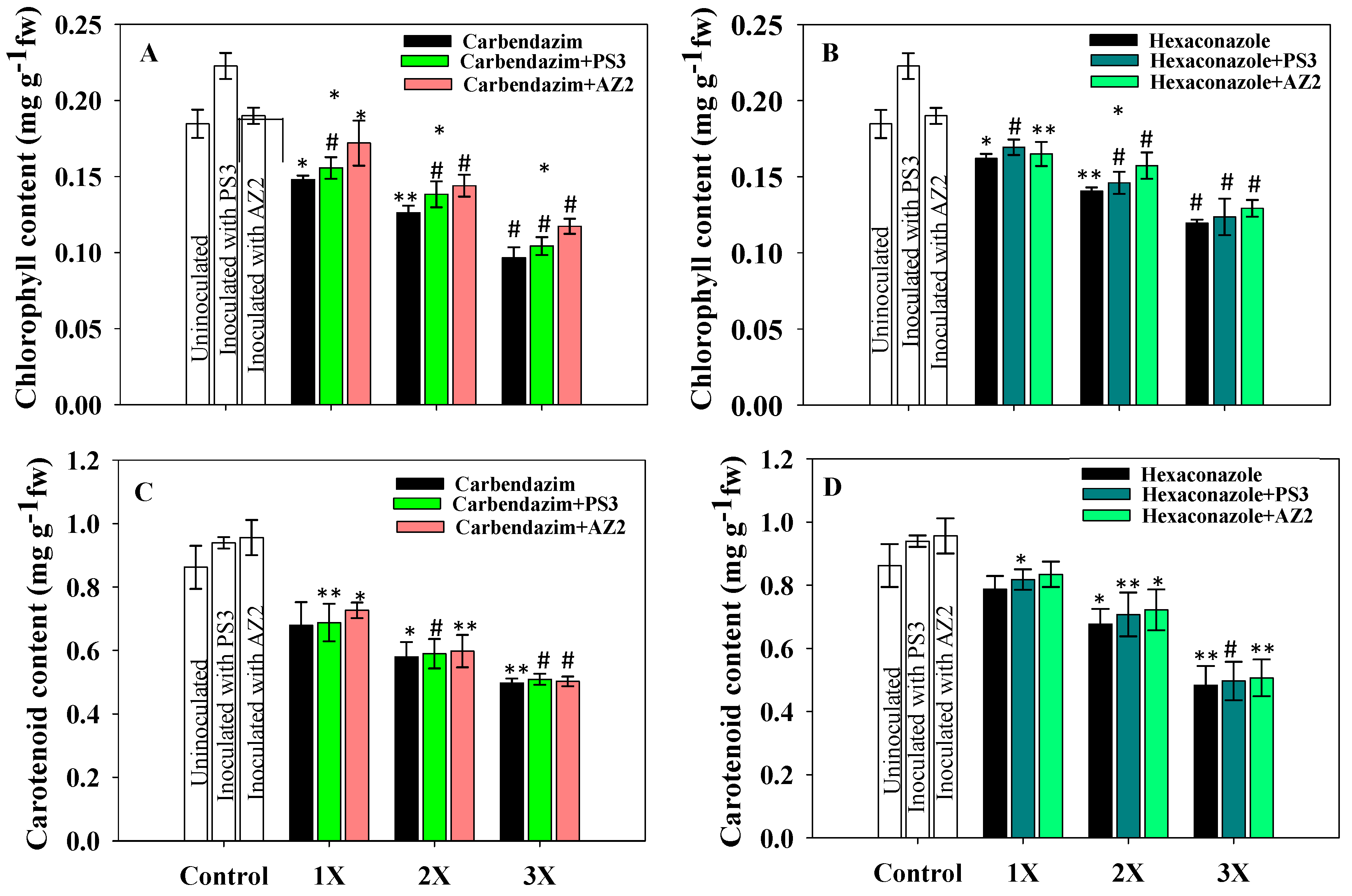
Total Chlorophyll and Carotenoid Content
3.6. Protein Content
3.7. Proline Accumulation
3.8. Lipid Peroxidation (MDA Content)
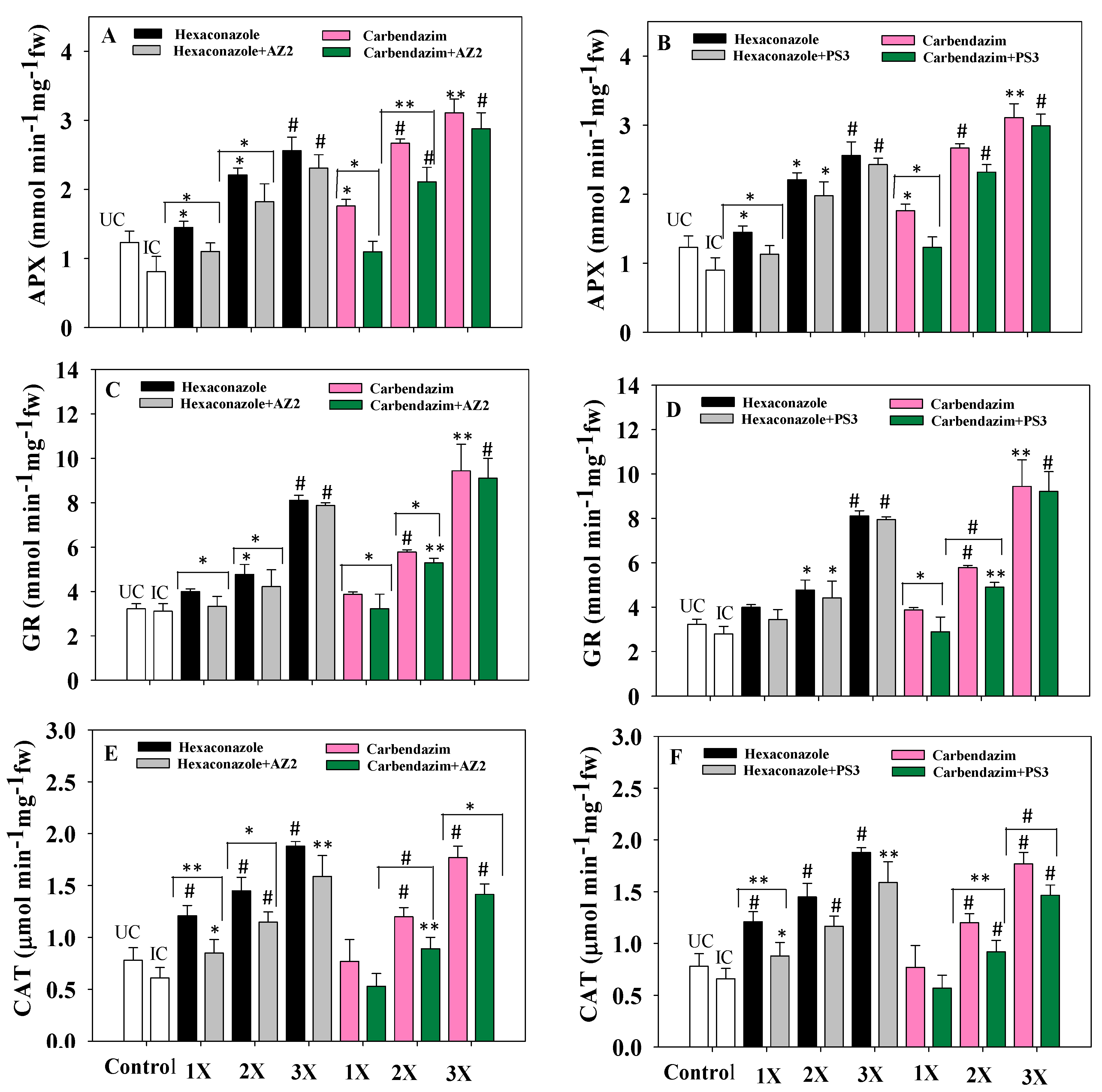
3.9. Antioxidant Enzymes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mallikarjunarao, K.; Singh, P.K.; Vaidya, A.; Pradhan, R.; Das, R.K. Genetic variability and selection parameters for different genotypes of radish (Raphanus sativus L.) under Kashmir valley. Ecol. Environ. Conserv. 2015, 21, 361–364. [Google Scholar]
- Jadoun, J.; Yazbak, A.; Rushrush, S.; Rudy, A.; Azaizeh, H. Identification of a new antibacterial sulfur compound from Raphanus sativus seeds. Evid. Based Complement. Altern. Med. 2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sridharan, R.; Raja, S.; Sakthivel, P. Effect of triazole compounds on induced changes in growth biomass and biochemical content of white radish (Raphanus sativus L.). J. Plant Stress Physiol. 2015, 1, 43–48. [Google Scholar] [CrossRef]
- Miao, Z.; He, S.; Zhang, X.; Wen, Z.; Bai, B.; Kong, X. Pathogens of root internal discoloration (black heart disease) in radish in Xining City of Qinghai Province, China. Mycosystema 2018, 37, 444–455. [Google Scholar]
- Lakra, B.S. Epiphytology and losses of downy mildew (Peronospora parasitica) of radish (Raphanus sativus) seed crop. Indian J. Agric. Sci. 2001, 71, 321–324. [Google Scholar]
- Gámiz, B.; Pignatello, J.J.; Cox, L.; Hermosín, M.C.; Celis, R. Environmental fate of the fungicide metalaxyl in soil amended with composted olive-mill waste and its biochar: An enantio selective study. Sci. Total Environ. 2017, 541, 776–783. [Google Scholar] [CrossRef]
- Mohamed, H.I.; Akladious, S.A. Changes in antioxidants potential, secondary metabolites and plant hormones induced by different fungicides treatment in cotton plants. Pestic. Biochem. Physiol. 2017, 142, 117–122. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, W.; Zhang, Y.; Teng, Y.; Xu, Z. Effects of fungicide iprodione and nitrification inhibitor 3,4-dimethylpyrazole phosphate on soil enzyme and bacterial properties. Sci. Total Environ. 2017, 599, 254–263. [Google Scholar] [CrossRef]
- Podile, A.R.; Kishore, G.K. Plant growth-promoting rhizobacteria. In Plant-Associated Bacteria; Springer: Dordrecht, The Netherlands, 2007; pp. 195–230. [Google Scholar]
- Rani, A.; Singh, R.; Kumar, P.; Shukla, G. Pros and cons of fungicides: An overview. Int. J. Eng. Sci. Res. Technol. 2017, 6, 112–117. [Google Scholar]
- Ju, C.; Xu, J.; Wu, X.; Dong, F.; Liu, X.; Tian, C.; Zheng, Y. Effects of hexaconazole application on soil microbe’s community and nitrogen transformations in paddy soils. Sci. Total Environ. 2017, 609, 655–663. [Google Scholar] [CrossRef]
- Singh, S.; Gupta, R.; Kumari, M.; Sharma, S. Nontarget effects of chemical pesticides and biological pesticide on rhizospheric microbial community structure and function in Vigna radiata. Environ. Sci. Pollut. Res. 2015, 22, 11290–11300. [Google Scholar] [CrossRef] [PubMed]
- Kengar, Y.D.; Patil, B.J. Ureide content of guar under influence of hexaconazole and triazophos. J. Fertil. Pestic. 2017, 8, 2. [Google Scholar] [CrossRef]
- Chiaia-Hernández, A.C.; Scheringer, M.; Müller, A.; Stieger, G.; Wächter, D.; Keller, A.; Pintado-Herrera, M.G.; Lara-Martin, P.A.; Bucheli, T.D.; Hollender, J. Target and suspect screening analysis reveals persistent emerging organic contaminants in soils and sediments. Sci. Total Environ. 2020, 740. [Google Scholar] [CrossRef] [PubMed]
- Narenderan, S.T.; Meyyanathan, S.N.; Babu, B. Review of pesticide residue analysis in fruits and vegetables. Pre-treatment, extraction and detection techniques. Food Res. Int. 2020. [Google Scholar] [CrossRef] [PubMed]
- Johri, R.; Dayal, V.; Johri, P.K. Finger printing technique for rapid monitoring of agrochemical residue in common vegetables and their inhibition arrest by phytoactivities of Moringa oleifera Lam. Biochem. Cell. Arch. 2011, 11, 19–27. [Google Scholar]
- Suryadi, Y.; Susilowati, D.N.; Fauziah, F. Management of Plant Diseases by PGPR-Mediated Induced Resistance with Special Reference to Tea and Rice Crops. In Plant Growth Promoting Rhizobacteria for Sustainable Stress Management; Springer: Singapore, 2019; pp. 65–110. [Google Scholar]
- Ziaf, K.; Latif, U.; Amjad, M.; Shabir, M.Z.; Asghar, W.; Ahmed, S.; Ahmad, I.; Jahangir, M.M.; Anwar, W. Combined use of microbial and synthetic amendments can improve radish (Raphanus sativus) yield. J. Environ. Agric. Sci. 2016, 6, 10–15. [Google Scholar]
- Lara, C.; Sanes, S.; Oviedo, L. Impact of native phosphate solubilizing bacteria on the growth and development of radish (Raphanus sativus) plants. Appl. Biotechnol. 2013, 30, 276–279. [Google Scholar]
- Khyade, V.B.; Dongare, S.K.; Shinde, M.R. Utilization of rhizobacteria, Pseudomonas trivialis (L) and Earthworm, Eudriluseugenae (L) for the qualitative and quantitative yield in the crop of radish (Raphanus sativus). Int. J. Agrochem. 2018, 4, 24–42. [Google Scholar]
- Mohamed, H.I.; Gomaa, E.Z. Effect of plant growth promoting Bacillus subtilis and Pseudomonas fluorescens on growth and pigment composition of radish plants (Raphanus sativus) under NaCl stress. Photosynthetica 2012, 50, 263–272. [Google Scholar] [CrossRef]
- Yildirim, E.; Turan, M.; Donmez, M.F. Mitigation of salt stress in radish (Raphanus sativus L.) by plant growth promoting rhizobacteria. Roum. Biotechnol. Lett. 2008, 13, 3933–3943. [Google Scholar]
- Ahmad, I.; Akhtar, M.J.; Mehmood, S.; Akhter, K.; Tahir, M.; Saeed, M.F.; Hussain, M.B.; Hussain, S. Combined application of compost and Bacillus sp. CIK-512 ameliorated the lead toxicity in radish by regulating the homeostasis of antioxidants and lead. Ecotoxicol. Environ. Saf. 2018, 148, 805–812. [Google Scholar] [CrossRef] [PubMed]
- Khatibi, R. Using sulfur oxidizing bacteria and P solubilizing for enhancing phosphorous availability to Raphanus sativus. Afr. J. Plant Sci. 2019, 5, 430–435. [Google Scholar]
- Gupta, S.; Kaushal, R.; Sood, G. Impact of plant growth–promoting rhizobacteria on vegetable crop production. Int. J. Veg. Sci. 2018. [Google Scholar] [CrossRef]
- Zaidi, A.; Khan, M.S.; Saif, S.; Rizvi, A.; Ahmed, B.; Shahid, M. Role of nitrogen-fixing plant growth- promoting rhizobacteria in sustainable production of vegetables: Current perspective. Microb. Strateg. Veg. Prod. 2017. [Google Scholar] [CrossRef]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant growth-promoting rhizobacteria: Context, mechanisms of action, and roadmap to commercialization of biostimulants for sustainable agriculture. Front. Plant Sci. 2018. [Google Scholar] [CrossRef] [Green Version]
- Rekha, K.; Ramasamy, M.; Usha, B. Root exudation of organic acids as affected by plant growth-promoting rhizobacteria Bacillus subtilis RR4 in rice. J. Crop Improv. 2020, 34, 571–586. [Google Scholar] [CrossRef]
- Abd El-Aziz, F.E.Z.A.; Bashandy, S.R. Dose-dependent effects of Pseudomonas trivialis rhizobacteria and synergistic growth stimulation effect with earthworms on the common radish. Rhizosphere 2019, 10. [Google Scholar] [CrossRef]
- Pal, A.K.; Sengupta, C. Isolation of cadmium and lead tolerant plant growth promoting rhizobacteria: Lysinibacillus varians and Pseudomonas putida from Indian Agricultural Soil. Soil Sediment Contam. 2019, 28, 601–629. [Google Scholar] [CrossRef]
- Akhtar, M.J.; Ullah, S.; Ahmad, I.; Rauf, A.; Nadeem, S.M.; Khan, M.Y.; Hussain, S.; Bulgariu, L. Nickel phytoextraction through bacterial inoculation in Raphanus sativus. Chemosphere 2018, 190, 234–242. [Google Scholar] [CrossRef]
- Hussein, K.A.; Joo, J.H. Plant growth-promoting rhizobacteria improved salinity tolerance of Lactuca sativa and Raphanus sativus. J. Microbiol. Biotechnol. 2018, 28, 938–945. [Google Scholar] [CrossRef] [Green Version]
- Shahid, M.; Zaidi, A.; Saghir, M. Modulations in growth, structure, cell viability and antioxidant enzyme of a nodule bacterium Mesorhizobium ciceri induced by pesticides. Environ. Dev. Sustain. 2020, 1–17. [Google Scholar] [CrossRef]
- Brick, J.M.; Bostock, R.M.; Silverstone, S.E. Rapid in situ assay for indole acetic acid production by bacteria immobilized on a nitrocellulose membrane. Appl. Environ. Microbiol. 1991, 57, 535–538. [Google Scholar] [CrossRef] [Green Version]
- Atkin, C.L.; Neilands, J.B.; Phaff, H.J. Rhodotorulic acid from species of Leucosporidium, Rhodosporidium, Rhodotorula, Sporidiobolus, and Sporobolomyces, and a new alanine-containing ferrichrome from Cryptococcus melibiosum. J. Bacteriol. 1970, 103, 722–733. [Google Scholar] [CrossRef] [Green Version]
- Shahid, M.; Khan, M.S. Cellular destruction, phytohormones and growth modulating enzymes production by Bacillus subtilis strain BC8 impacted by fungicides. Pestic. Biochem. Physiol. 2018, 149, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Jackson, M.L. Soil Chemical Analysis; Prentice Hall: New Delhi, India, 1976. [Google Scholar]
- Bakker, A.W.; Schippers, B. Microbial cyanide production in the rhizosphere in relation to potato yield reduction and Pseudomonas spp-mediated plant growth-stimulation. Soil Biol. Biochem. 1987, 19, 451–457. [Google Scholar] [CrossRef]
- Dye, D.W. The inadequacy of the usual determinative tests for the identification of Xanthomonas spp. N. Z. J. Sci. 1962, 5, 393–416. [Google Scholar]
- Shahid, M.; Zaidi, A.; Ehtram, A.; Khan, M.S. In vitro investigation to explore the toxicity of different groups of pesticides for an agronomically important rhizosphere isolate Azotobacter vinelandii. Pestic. Biochem. Physiol. 2019, 157, 33–44. [Google Scholar] [CrossRef]
- Shahid, M.; Khan, M.S. Glyphosate induced toxicity to chickpea plants and stress alleviation by herbicide tolerant phosphate solubilizing Burkholderia cepacia PSBB1 carrying multifarious plant growth promoting activities. 3 Biotech 2018, 8, 131. [Google Scholar] [CrossRef]
- Shahid, M.; Ahmed, B.; Khan, M.S. Evaluation of microbiological management strategy of herbicide toxicity to greengram plants. Biocatal. Agric. Biotechol. 2018, 14, 96–108. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Ahmed, B.; Dwivedi, S.; Abdin, M.Z.; Azam, A.; Al-Shaeri, M.; Khan, M.S.; Saquib, Q.; Al-Khedhairy, A.A.; Musarrat, J. Mitochondrial and chromosomal damage induced by oxidative stress in Zn2+ ions, ZnO-bulk and ZnO-NPs treated Allium cepa roots. Sci. Rep. 2017, 7, 40685. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.; Khan, M.S.; Kumar, M. Kitazin-pea interaction: Understanding the fungicide induced nodule alteration, cytotoxicity, oxidative damage and toxicity alleviation by Rhizobium leguminosarum. RSC Adv. 2019, 9, 16929–16947. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.; Kumar, A.; Singh, M.; Pandey, K.D. Isolation and characterization of plant growth promoting rhizobacteria from Momordica Charantia L. In PGPR Amelioration in Sustainable Agriculture; Woodhead Publishing: Sawaton, UK, 2019; pp. 217–238. [Google Scholar]
- Rajwar, A.; Sahgal, M. Phylogenetic relationships of fluorescent pseudomonads deduced from the sequence analysis of 16S rRNA, Pseudomonas-specific and rpoD genes. 3 Biotech 2016, 6, 80. [Google Scholar] [CrossRef] [Green Version]
- Alam, S.; Kumar, A.; Kumar, A.; Prasad, S.; Tiwari, A.; Srivastava, D.; Srivastava, S.; Tiwari, P.; Singh, J. Mathur, B. Isolation and characterization of pesticide tolerant bacteria from brinjal rhizosphere. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 4849–4859. [Google Scholar]
- Roy, T.; Das, N. Isolation, characterization, and identification of two methomyl-degrading bacteria from a pesticide-treated crop field in West Bengal. India Microbiol. 2017, 86, 753–764. [Google Scholar] [CrossRef]
- Anjum, R.; Krakat, N. Detection of multiple resistances, biofilm formation and conjugative transfer of Bacillus cereus from contaminated soils. Curr. Microbiol. 2016, 72, 321–328. [Google Scholar] [CrossRef]
- Ahemad, M.; Khan, M.S. Effect of fungicides on plant growth promoting activities of phosphate solubilizing Pseudomonas putida isolated from mustard (Brassica compestris) rhizosphere. Chemosphere 2012, 86, 945–950. [Google Scholar] [CrossRef]
- Ahemad, M.; Khan, M.S. Alleviation of fungicide-induced phytotoxicity in greengram [Vigna radiata (L.) Wilczek] using fungicide-tolerant and plant growth promoting Pseudomonas strain. Saudi J. Biol. Sci. 2012, 19, 451–459. [Google Scholar] [CrossRef] [Green Version]
- Bedini, A.; Mercy, L.; Schneider, C.; Franken, P.; Lucic-Mercy, E. Unraveling the initial plant hormone signalling, metabolic mechanisms and plant defence triggering the endomycorrhizal symbiosis behaviour. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Deinum, E.E.; Kohlen, W.; Geurts, R. Quantitative modelling of legume root nodule primordium induction by a diffusive signal of epidermal origin that inhibits auxin efflux. BMC Plant Biol. 2016, 16, 254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurth, C.; Kage, H.; Nett, M. Siderophores as molecular tools in medical and environmental applications. Org. Biomol. Chem. 2016, 14, 8212–8822. [Google Scholar] [CrossRef] [PubMed]
- Dorjey, S.; Dolkar, D.; Sharma, R. Plant growth promoting rhizobacteria Pseudomonas: A review. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 1335–1344. [Google Scholar] [CrossRef]
- Eshaghi, E.; Nosrati, R.; Owlia, P.; Malboobi, M.A.; Ghaseminejad, P.; Ganjali, M.R. Zinc solubilization characteristics of efficient siderophore-producing soil bacteria. Iran. J. Microbiol. 2019, 11, 418–430. [Google Scholar] [CrossRef]
- Karmakar, P.; Sharma, D.; Das, P.; Saha, A.K. Phosphate solubilizing capacity and siderophore production by arthroderma cuniculi dawson 1963 isolated from rhizospheric soil. Res. J. Life Sci. Bioinforma. Pharm. Chem. Sci. 2018, 4, 330. [Google Scholar]
- Saif, S.; Khan, M.S. Assessment of toxic impact of metals on proline, antioxidant enzymes, and biological characteristics of Pseudomonas aeruginosa inoculated Cicer arietinum grown in chromium and nickel-stressed sandy clay loam soils. Environ. Monit. Assess. 2018, 190, 290. [Google Scholar] [CrossRef] [PubMed]
- Rizvi, A.; Khan, M.S. Heavy metal induced oxidative damage and root morphology alterations of maize (Zea mays L.) plants and stress mitigation by metal tolerant nitrogen fixing Azotobacter chroococcum. Ecotoxicol. Environ. Saf. 2018, 157, 9–20. [Google Scholar] [CrossRef] [PubMed]
- Ditta, A.; Khalid, A. Bio-organo-phos: A sustainable approach for managing phosphorus deficiency in agricultural soils. In Organic Fertilizers-from Basic Concepts to Applied; InTechOpen: Rijeka, Croatia, 2016. [Google Scholar]
- Ramawat, K.G.; Goyal, S. Co-evolution of Secondary Metabolites During Biological Competition for Survival and Advantage: An Overview. In Co-Evolution of Secondary Metabolites; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar]
- Ansari, R.A.; Mahmood, I.; Rizvi, R.; Sumbul, A. Siderophores: Augmentation of soil health and crop productivity. In Probiotics in Agroecosystem; Springer: Singapore, 2017; pp. 291–312. [Google Scholar]
- Rath, M.; Mitchell, T.R.; Gold, S.E. Volatiles produced by Bacillus mojavensis RRC101 act as plant growth modulators and are strongly culture-dependent. Microbiol. Res. 2018, 208, 76–84. [Google Scholar] [CrossRef]
- Flury, P.; Vesga, P.; Péchy-Tarr, M.; Aellen, N.; Dennert, F.; Hofer, N.; Kupferschmied, K.P.; Kupferschmied, P.; Metla, Z.; Ma, Z.; et al. Antimicrobial and insecticidal: Cyclic lipopeptides and hydrogen cyanide produced by plant-beneficial Pseudomonas strains CHA0, CMR12a, and PCL1391 contribute to insect killing. Front. Microbiol. 2017, 8, 100. [Google Scholar] [CrossRef]
- Safni, I.; Antastia, W. In vitro antagonism of five rhizobacterial species against Athelia rolfsii collar rot disease in soybean. Open Agric. 2018, 3, 264–272. [Google Scholar] [CrossRef]
- Lahlali, R.; Aksissou, W.; Lyousfi, N.; Ezrari, S.; Blenzar, A.; Tahiri, A.; Ennahli, S.; Hrustić, J.; MacLean, D.; Amiri, S. Biocontrol activity and putative mechanism of Bacillus amyloliquefaciens (SF14 and SP10), Alcaligenes faecalis ACBC1, and Pantoea agglomerans ACBP1 against brown rot disease of fruit. Microb. Pathog. 2020, 139, 103914. [Google Scholar] [CrossRef] [PubMed]
- Chakravarty, R.; Banerjee, P.C. Morphological changes in an acidophilic bacterium induced by heavy metals. Extremophiles 2008, 12, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Jan, G.; Leverrier, P.; Pichereau, V.; Boyaval, P. Changes in protein synthesis and morphology during acid adaptation of Propionibacterium freudenreichii. Appl. Environ. Microbiol. 2001, 67, 2029–2036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahid, M.; Khan, M.S. Assessment of glyphosate and quizalofop mediated toxicity to greengram [Vigna radiata (L.) Wilczek], stress abatement and growth promotion by herbicide tolerant Bradyrhizobium and Pseudomonas species. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 3001–3016. [Google Scholar] [CrossRef]
- Nikaido, H. Molecular basis of bacterial outer membrane permeability revisited. Microbiol. Mol. Biol. Rev. 2003, 67, 593–656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, B.; Ameen, F.; Rizvi, A.; Ali, K.; Sonbol, H.; Zaidi, A.; Khan, M.S.; Musarrat, J. Destruction of cell topography, morphology, membrane, inhibition of respiration, biofilm formation, and bioactive molecule production by nanoparticles of Ag, ZnO, CuO, TiO2, and Al2O3 toward beneficial soil bacteria. ACS Omega 2020, 5, 7861–7876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef] [PubMed]
- Kaul, S.; Sharma, S.S.; Mehta, I.K. Free radical scavenging potential of l-proline: Evidence from in vitro assays. Amino Acids 2008, 34, 315–320. [Google Scholar] [CrossRef]
- Siddique, A.; Kandpal, G.; Kumar, P. Proline accumulation and its defensive role under diverse stress condition in plants: An overview. J. Pure Appl. Microbiol. 2018, 12, 1655–1659. [Google Scholar] [CrossRef]
- Ramakrishna, B.; Rao, S.S.R. 24-Epibrassinolide alleviated zinc-induced oxidative stress in radish (Raphanus sativus L.) seedlings by enhancing antioxidative system. Plant Growth Regul. 2012, 68, 249–259. [Google Scholar] [CrossRef]
- Shivakumar, S.; Bhaktavatchalu, S. Role of plant growth-promoting rhizobacteria (PGPR) in the improvement of vegetable crop production under stress conditions. In Microbial Strategies for Vegetable Production; Springer: Cham, Switzerland, 2017; pp. 81–97. [Google Scholar]
- Paul, S.; Upadhyay, S.K.; Lal, E.P. Accumulation of arsenic in radish (Raphanus sativus L.), and their effects on growth and antioxidant activities. Int. J. Pharm. Sci. Res. 2014, 5, 3536. [Google Scholar]
- Bhattacharjee, S. ROS and oxidative modification of cellular components. In Reactive Oxygen Species in Plant Biology; Springer: New Delhi, India, 2019; pp. 81–105. [Google Scholar]
- Christhuuthayam, M.; Vijayarengan, P.; Natarajan, A.; Maheswari, M. Changes in biochemical constitution of radish (Raphanus sativus L.) Under copper toxicity. J. Plant Stress Physiol. 2018, 4, 22–24. [Google Scholar]
- Curá, J.A.; Franz, D.R.; Filosofía, J.E.; Balestrasse, K.B. Inoculation with Azospirillum sp. and Herbaspirillum sp. bacteria increases the tolerance of maize to drought stress. Microorganisms 2017, 5, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khanna, K.; Jamwal, V.L.; Gandhi, S.G.; Ohri, P.; Bhardwaj, R. Metal resistant PGPR lowered Cd uptake and expression of metal transporter genes with improved growth and photosynthetic pigments in Lycopersicon esculentum under metal toxicity. Sci. Rep. 2019, 9, 5855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pravisya, P.; Jayaram, K.M.; Yusuf, A. Biotic priming with Pseudomonas fluorescens induces drought stress tolerance in Abelmoschus esculentus (L.) Moench (Okra). Physiol. Mol. Biol. Plants 2019, 25, 101–112. [Google Scholar] [CrossRef]
- Ge, H.; Zhang, F. Growth-Promoting Ability of Rhodopseudomonas palustris G5 and Its Effect on Induced Resistance in Cucumber Against Salt Stress. J. Plant Growth Regul. 2019, 38, 180–188. [Google Scholar] [CrossRef]
- Chew, O.; Whelan, J.; Millar, A.H. Molecular definition of the ascorbate-glutathione cycle in Arabidopsis mitochondria reveals dual targeting of antioxidant defenses in plants. J. Biol. Chem. 2003, 278, 46869–46877. [Google Scholar] [CrossRef] [Green Version]
- Giorgio, M.; Trinei, M.; Migliaccio, E.; Pelicci, P.G. Hydrogen peroxide: A metabolic by-product or a common mediator of ageing signals? Nat. Rev. Mol. Cell Biol. 2007, 8, 722–728. [Google Scholar] [CrossRef]
- Sivagamasundari, R. Anti-Hyperlipidemic Activity of Aerial Parts of Delonix Elata on High Cholesterol Diet Induced Hyperlipidemia in Rats. Ph.D. Thesis, Mohamed Sathak AJ College of Pharmacy, Chennai, India, 2017. [Google Scholar]
- Kumar, R.; Nongkhlaw, M.; Acharya, C.; Joshi, S.R. Growth media composition and heavy metal tolerance behaviour of bacteria characterized from the sub-surface soil of uranium rich ore bearing site of Domiasiat in Meghalaya. Indian J. Biotechnol. 2013, 12, 115–119. [Google Scholar]
- Deising, H.B.; Reimann, S.; Pascholati, S.F. Mechanisms and significance of fungicide resistance. Braz. J. Microbiol. 2008, 39, 286–295. [Google Scholar] [CrossRef]
- Kang, B.G.; Kim, W.T.; Yun, H.S.; Chang, S.C. Use of plant growth-promoting rhizobacteria to control stress responses of plant roots. Plant Biotechnol. Rep. 2010, 4, 179–183. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | Plant Growth-Promoting Rhizobacteria | |
|---|---|---|
| Isolate AZ2 | Isolate PS3 | |
| Colony morphology | Irregular margin, white and mucoid colony | Irregular margin, mucoid pale yellow colony |
| Gram reaction | Negative | Negative |
| Cell shape Pigmentation | Short rod Non- fluorescent | Short rod Green fluorescent |
| Citrate utilization | + | + |
| Indole production | + | ND |
| Methyl red test | + | + |
| Nitrate reduction | + | + |
| Oxidase activity | ND | + |
| Voges Proskauer test | + | ND |
| Dextrose utilization | ND | + |
| Lactose utilization | ND | + |
| Mannitol utilization | + | + |
| Sucrose utilization | + | + |
| Starch hydrolysis | + | + |
| Gelatin hydrolysis | + | + |
| Maximum tolerance dose exposed to carbendazim | 2400 µg mL−1 | 2400 µg mL−1 |
| Maximum tolerance dose exposed to hexaconazole | 1600 µg mL−1 | 3200 µg mL−1 |
| Bacterial Isolate | Treatment | Plant Growth-Promoting Activity | |||||
|---|---|---|---|---|---|---|---|
| Dose Rates (µg mL−1) | IAA (µg mL−1) | Phosphate-Solubilization (µg mL−1) | Siderophore Production | NH3 Production | HCN | ||
| Pseudomonas sp. PS3 | Control | 0 | 61.3 a ± 3.5 | 36.0 a ± 2.3 | ++ | +++ | ++ |
| Carbendazim | 500 | 49.4 c ± 2.3 | 28.8 b ± 3.0 | + | ++ | + | |
| 1000 | 39.8 d ± 2.8 | 21.0 c ± 1.3 | + | + | + | ||
| 1500 | 21.3 e ± 2.0 | 15.1 d ± 1.2 | ND | + | + | ||
| Hexaconazole | 500 | 56.3 b ± 3.2 | 28.9 b ± 3.2 | + | ++ | ++ | |
| 1000 | 40.0 d ± 3.0 | 21.3 c ± 1.7 | + | + | + | ||
| 1500 | 24.8 e ± 1.3 | 11.8 d ± 1.4 | ND | + | + | ||
| Mean | - | - | 41.7 | 20.5 | - | - | - |
| Pseudomonas sp. AZ2 | Control | 0 | 39.5 a ± 0 | 26.2 a ± 2.3 | +++ | +++ | ++ |
| Carbendazim | 500 | 33.9 b ± 1.3 | 25.8 b ± 3.0 | ++ | ++ | + | |
| 1000 | 26.4 cd ± 1.7 | 20.0 c ± 1.3 | + | + | + | ||
| 1500 | 18.9 e ± 2.3 | 14.4 d ± 1.2 | + | + | + | ||
| Hexaconazole | 500 | 29.5 c ± 3.3 | 23.1 b ± 3.2 | ++ | ++ | ++ | |
| 1000 | 23.0 d ± 1.0 | 19.3 c ± 1.7 | + | + | + | ||
| 1500 | 16.9 e ± 2.0 | 14.6 d ± 1.4 | + | + | + | ||
| Mean | - | - | 26.8 | 20.48 | - | - | - |
| LSD (p ≤ 0.05) | - | - | 92.46 | 45.4 | - | - | - |
| F value | - | - | 8.41 | 13.28 | - | - | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, S.; Shahid, M.; Khan, M.S.; Syed, A.; Bahkali, A.H.; Elgorban, A.M.; Pichtel, J. Fungicide-Tolerant Plant Growth-Promoting Rhizobacteria Mitigate Physiological Disruption of White Radish Caused by Fungicides Used in the Field Cultivation. Int. J. Environ. Res. Public Health 2020, 17, 7251. https://doi.org/10.3390/ijerph17197251
Khan S, Shahid M, Khan MS, Syed A, Bahkali AH, Elgorban AM, Pichtel J. Fungicide-Tolerant Plant Growth-Promoting Rhizobacteria Mitigate Physiological Disruption of White Radish Caused by Fungicides Used in the Field Cultivation. International Journal of Environmental Research and Public Health. 2020; 17(19):7251. https://doi.org/10.3390/ijerph17197251
Chicago/Turabian StyleKhan, Sadaf, Mohammad Shahid, Mohammad Saghir Khan, Asad Syed, Ali H. Bahkali, Abdallah M. Elgorban, and John Pichtel. 2020. "Fungicide-Tolerant Plant Growth-Promoting Rhizobacteria Mitigate Physiological Disruption of White Radish Caused by Fungicides Used in the Field Cultivation" International Journal of Environmental Research and Public Health 17, no. 19: 7251. https://doi.org/10.3390/ijerph17197251
APA StyleKhan, S., Shahid, M., Khan, M. S., Syed, A., Bahkali, A. H., Elgorban, A. M., & Pichtel, J. (2020). Fungicide-Tolerant Plant Growth-Promoting Rhizobacteria Mitigate Physiological Disruption of White Radish Caused by Fungicides Used in the Field Cultivation. International Journal of Environmental Research and Public Health, 17(19), 7251. https://doi.org/10.3390/ijerph17197251