Interaction of the CMTM7 rs347134 Polymorphism with Dietary Patterns and the Risk of Obesity in Han Chinese Male Children


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Anthropometric Assessments
2.3. Dietary and Physical Activity Assessments
2.4. Laboratory Assays
2.5. Genotyping
2.6. Statistical Analyses
3. Results
3.1. Association between CMTM7 rs347134 Polymorphism and Obesity-Related Indices
3.2. Association between Dietary Patterns and Obesity-Related Indices in Male Children
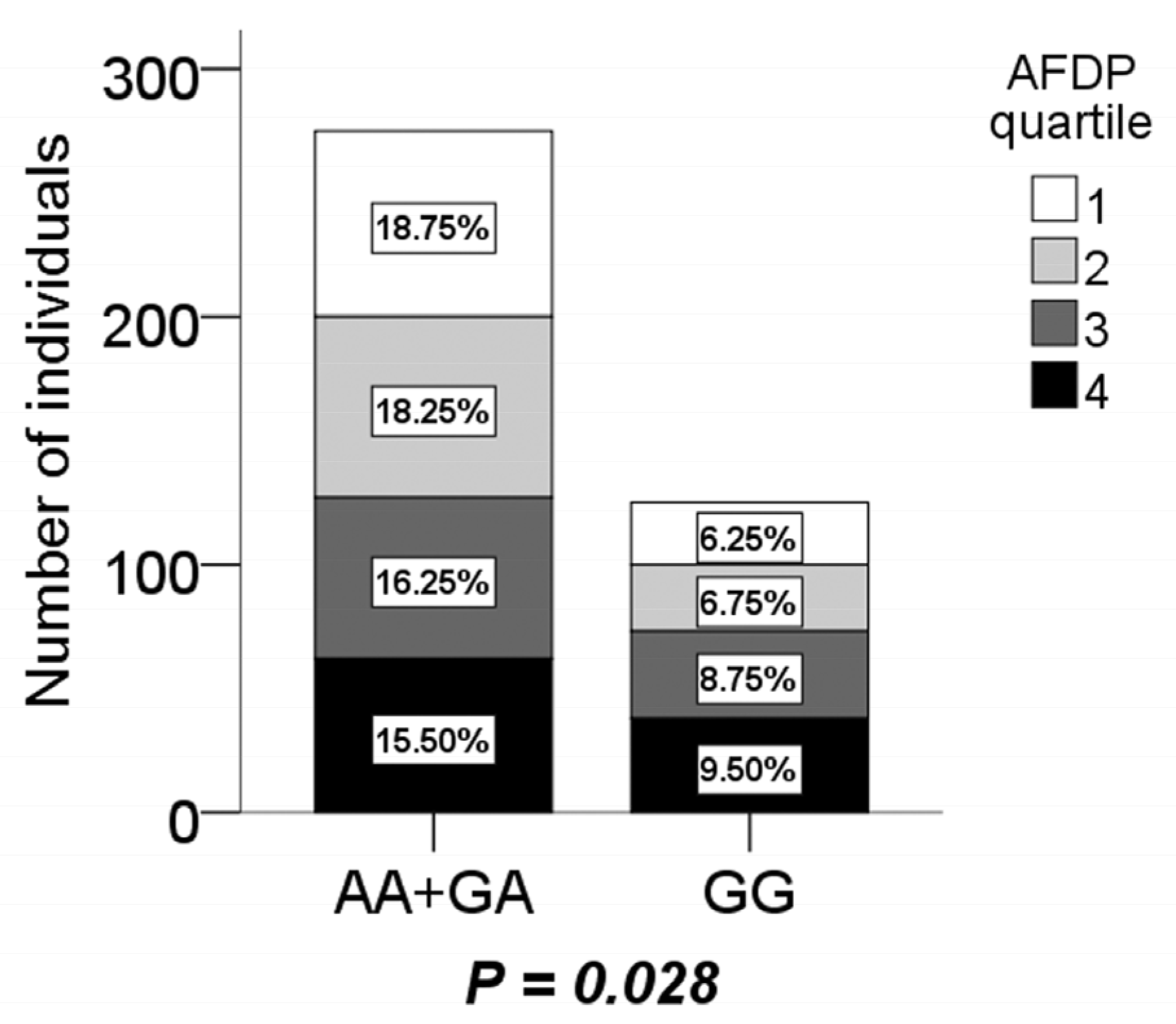
3.3. Association of CMTM7 rs347134 Polymorphism with Energy Intake, Activity and Dietary Pattern in Boys
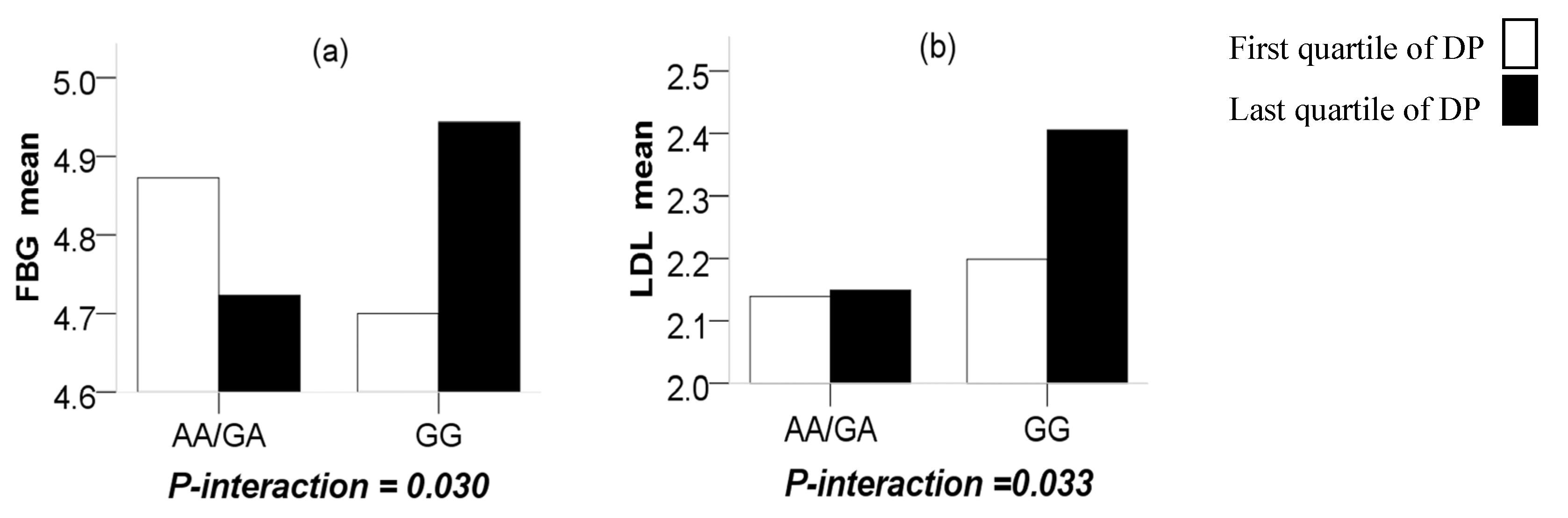
3.4. Interaction of CMTM7 rs347134 Polymorphism with Energy Intake, Activity and Dietary Pattern in Boys
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ng, M.; Fleming, T.; Robinson, M.; Thomson, B.; Graetz, N.; Margono, C.; Mullany, E.C.; Biryukov, S.; Abbafati, C.; Abera, S.F.; et al. Global, Regional, and National Prevalence of Overweight and Obesity in Children and Adults during 1980–2013: A Systematic Analysis for the Global Burden of Disease Study 2013. Lancet 2014, 384, 766–781. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Dong, Y.H.; Wang, Z.H.; Zou, Z.Y.; Ma, J. Trends in overweight and obesity among Chinese children of 7–18 years old during 1985–2014. Chin. J. Prev. Med. 2017, 51, 300–305. [Google Scholar]
- Song, Y.; Wang, H.J.; Dong, B.; Ma, J.; Wang, Z.Q.; Agardh, A. 25-year trends in gender disparity for obesity and overweight by using WHO and IOTF definitions among Chinese school-aged children: A multiple cross-sectional study. BMJ Open 2016, 6, e011904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.; Ma, J.; Wang, H.J.; Wang, Z.Q.; Zhang, B.; Agard, A. Secular trends of obesity prevalence in Chinese children from 1985 to 2010: Urban-rural disparity. Obesity 2015, 23, 448–453. [Google Scholar] [CrossRef] [PubMed]
- Sabin, M.A.; Shield, J.P. Childhood obesity. Front. Horm. Res. 2008, 36, 85–96. [Google Scholar] [PubMed]
- Adair, L.S.; Gordon-Larsen, P.; Du, S.F.; Zhang, B.; Popkin, B.M. The emergence of cardiometabolic disease risk in Chinese children and adults: Consequences of changes in diet, physical activity, and obesity. Obes. Rev. 2014, 15 (Suppl. S1), 49–59. [Google Scholar] [CrossRef]
- Chan, R.S.M.; Woo, J. Prevention of Overweight and Obesity: How Effective is the Current Public Health Approach. Int. J. Environ. Res. Public Health 2010, 7, 765–783. [Google Scholar] [CrossRef] [Green Version]
- Castillo, J.J.; Orlando, R.A.; Garver, W.S. Gene-nutrient interactions and susceptibility to human obesity. Genes Nutr. 2017, 12, 29. [Google Scholar] [CrossRef] [Green Version]
- Elks, C.E.; den Hoed, M.; Zhao, J.H.; Sharp, S.J.; Wareham, N.J.; Loos, R.J.F.; Ong, K.K. Variability in the Heritability of Body Mass Index: A Systematic Review and Meta-Regression. Front. Endocrinol. 2012, 3, 29. [Google Scholar] [CrossRef] [Green Version]
- Clement, K.; Garner, C.; Hager, J.; Philippi, A.; LeDuc, C.; Carey, A.; Harris, T.J.; Jury, C.; Cardon, L.R.; Basdevant, A.; et al. Indication for linkage of the human OB gene region with extreme obesity. Diabetes 1996, 45, 687–690. [Google Scholar] [CrossRef] [Green Version]
- Loos, R.J.F.; Yeo, G.S.H. The Bigger Picture of FTO: The First GWAS-identified Obesity Gene. Nat. Rev. Endocrinol. 2014, 10, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Young, E.H.; Wareham, N.J.; Farooqi, S.; Hinney, A.; Hebebrand, J.; Scherag, A.; O’rahilly, S.; Barroso, I.; Sandhu, M.S. The V103I polymorphism of the MC4R gene and obesity: Population based studies and meta-analysis of 29 563 individuals. Int. J. Obes. 2007, 31, 1437–1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodarzi, M.O. Genetics of Obesity: What Genetic Association Studies Have Taught Us about the Biology of Obesity and Its Complications. Lancet Diabetes Endocrinol. 2018, 6, 223–236. [Google Scholar] [CrossRef]
- Peng, S.; Zhu, Y.M.; Xu, F.Y.; Ren, X.B.; Li, X.B.; Lai, M.D. FTO Gene Polymorphisms and Obesity Risk: A Meta-Analysis. BMC Med. 2011, 9, 71. [Google Scholar] [CrossRef] [Green Version]
- Speakman, J.R.; Rance, K.A.; Johnstone, A.M. Polymorphisms of the FTO Gene Are Associated with Variation in Energy Intake, but Not Energy Expenditure. Obesity 2008, 16, 1961–1965. [Google Scholar] [CrossRef]
- Prats-Puig, A.; Grau-Cabrera, P.; Riera-Pérez, E.; Cortés-Marina, R.; Fortea, E.; Soriano-Rodríguez, P.; de Zegher, F.; Ibánez, L.; Bassols, J.; López-Bermejo, A.; et al. Variations in the Obesity Genes FTO, TMEM18 and NRXN3 Influence the Vulnerability of Children to Weight Gain Induced by Short Sleep Duration. Int. J. Obes. 2013, 37, 182–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xi, B.; Chandak, G.R.; Shen, Y.; Wang, Q.J.; Zhou, D.H. Association between Common Polymorphism near the MC4R Gene and Obesity Risk: A Systematic Review and Meta-Analysis. PLoS ONE 2012, 7, e45731. [Google Scholar] [CrossRef] [PubMed]
- Stutzmann, F.; Cauchi, S.; Durand, E.; Calvacanti-Proença, C.; Pigeyre, M.; Hartikainen, A.-L.; Sovio, U.; Tichet, J.; Marre, M.; Weill, J.; et al. Common Genetic Variation near MC4R Is Associated with Eating Behaviour Patterns in European Populations. Int. J. Obes. 2009, 33, 373–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, D.; Zhang, D.D.; Wang, H.; Zhang, Y.; Liang, L.; Fu, J.F.; Xiong, F.; Liu, G.L.; Gong, C.X.; Luo, F.H.; et al. Genetic variations in SEC16B, MC4R, MAP2K5 and KCTD15 were associated with childhood obesity and interacted with dietary behaviors in Chinese school-age population. Gene 2015, 560, 149–155. [Google Scholar] [CrossRef]
- Li, H.X.; Wu, Y.; Loos, R.J.F.; Hu, F.B.; Liu, Y.; Wang, J.; Yu, Z.J.; Lin, X. Variants in the Fat Mass- And Obesity-Associated (FTO) Gene Are Not Associated with Obesity in a Chinese Han Population. Diabetes 2008, 57, 264–268. [Google Scholar] [CrossRef] [Green Version]
- Hao, Y.C. Genome-Wide Association Study between Height and Body Mass Index in Chinese Han Population. Ph.D. Thesis, Peking Union Medical College, Beijing, China, 2013. [Google Scholar]
- Han, W.L.; Lou, Y.; Tang, J.; Zhang, Y.; Chen, Y.; Li, Y.; Gu, W.; Huang, J.; Gui, L.; Tang, Y.; et al. Molacteecular cloning and characterization of chemokine-like factor1 (CKLF1), a novel human cytokine with unique structure and potential chemotactic activity. Biochem. J. 2001, 357 Pt 1, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Han, W.L.; Ding, P.G.; Xu, M.X.; Wang, L.; Rui, M.; Shi, S.; Liu, Y.A.; Zheng, Y.; Chen, Y.Y.; Yang, T.; et al. Identification of eight genes encoding chemokine-like factor superfamily members 1–8 (CKLFSF1–8) by in silico cloning and experimental validation. Genomics 2003, 81, 609–617. [Google Scholar] [CrossRef]
- Working Group on Obesity in China. Classification criteria for overweight and obesity screening of school-age children and adolescents in China. Chin. J. Epidemiol. 2004, 25, 97–102. [Google Scholar]
- Zhou, D.; Yang, M.; Yuan, Z.P.; Lv, D.; Liu, Y.; Zhu, Y.M. Waist-to-height ratio: A simple, effective, and practical predictor of obesity and metabolic syndrome in children and adolescents. In Proceedings of the Annual Conference of Clinical Epidemiology and Evidence-Based Medicine, Hangzhou, Zhejiang, China, 5 September 2014. [Google Scholar]
- Ainsworth, B.E.; Haskell, W.L.; Whitt, M.C.; Irwin, M.L.; Swartz, A.M.; Strath, S.J.; O’Brien, W.L.; Bassett, D.R., Jr.; Schmitz, K.H.; Emplaincourt, P.O.; et al. Compendium of Physical Activities: An update of activity codes and MET intensities. Med. Sci. Sports Exerc. 2000, 32, S498–S504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, L.G.; Sun, L.L.; Wu, S.W.; Yang, Y.D.; Li, X.H.; Wang, Z.H.; Wu, L.; Wang, F.Z.; Ma, J. The Influence of Secular Trends in Body Height and Weight on the Prevalence of Overweight and Obesity among Chinese Children and Adolescents. Biomed. Environ. Sci. 2016, 29, 849–857. [Google Scholar] [PubMed]
- Kuba, V.M.; Leone, C.; Damiani, D. Is waist-to-height ratio a useful indicator of cardio-metabolic risk in 6–10-year-old children? BMC Pediatr. 2013, 13, 91. [Google Scholar] [CrossRef] [Green Version]
- Sung, R.Y.T.; Yu, C.C.W.; Choi, K.C.; McManus, A.; Li, A.M.C.; Xu, S.L.Y.; Chan, D.; Lo, A.F.C.; Chan, J.C.N.; Fok, T.F. Waist circumference and body mass index in Chinese children: Cutoff values for predicting cardiovascular risk factors. Int. J. Obes. 2007, 31, 550–558. [Google Scholar] [CrossRef] [Green Version]
- Moreno, L.A.; Pineda, I.; Rodriguez, G.; Fleta, J.; Sarria, A.; Bueno, M. Waist circumference for the screening of the metabolic syndrome in children. Acta Paediatr. 2002, 91, 1307–1312. [Google Scholar] [CrossRef]
- Li, H.; Li, J.; Su, Y.; Fan, Y.; Guo, X.; Li, L.; Su, X.; Rong, R.; Ying, J.; Mo, X.; et al. A novel 3p22.3 gene CMTM7 represses oncogenic EGFR signaling and inhibits cancer cell growth. Oncogene 2014, 33, 3109–3118. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Su, Y.; Li, T.; Yuan, W.Q.; Mo, X.N.; Li, H.; He, Q.H.; Ma, D.L.; Han, W.L. CMTM7 knockdown increases tumorigenicity of human non-small cell lung cancer cells and EGFR-AKT signaling by reducing Rab5 activation. Oncotarget 2015, 6, 41092–41107. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, A.; Yogosawa, S.; Murakami, A.; Kitamura, D. Identification of CMTM7 as a transmembrane linker of BLNK and the B-cell receptor. PLoS ONE 2012, 7, e31829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.F.; Wang, J.Y.; Han, W.L. A role for CMTM7 in BCR expression and survival in B-1a but not B-2 cells. Int. Immunol. 2014, 26, 47–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrison, A.C.; Felix, J.F.; Cupples, L.A.; Glazer, N.L.; Loehr, L.R.; Dehghan, A.; Demissie, S.; Bis, J.C.; Rosamond, W.D.; Aulchenko, Y.S.; et al. Genomic variation associated with mortality among adults of European and African ancestry with heart failure: The cohorts for heart and aging research in genomic epidemiology consortium. Circ. Cardiovasc. Genet. 2010, 3, 248–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kenchaiah, S.; Evans, J.C.; Levy, D.; Wilson, P.W.F.; Benjamin, E.J.; Larson, M.G.; Kannel, W.B.; Vasan, R.S. Obesity and the risk of heart failure. N. Engl. J. Med. 2002, 347, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Pourshahidi, L.K.; Kerr, M.A.; McCaffrey, T.A.; Livingstone, M.B.E. Influencing and Modifying Children’s Energy Intake: The Role of Portion Size and Energy Density. Proc. Nutr. Soc. 2014, 73, 397–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernandez-Rodas, M.C.; Valenzuela, R.; Videla, L.A. Relevant Aspects of Nutritional and Dietary Interventions in Non-Alcoholic Fatty Liver Disease. Int. J. Mol. Sci. 2015, 16, 25168–25198. [Google Scholar] [CrossRef] [Green Version]
- Figueiredo, P.S.; Inada, A.C.; Marcelino, G.; Cardozo, C.M.L.; Freitas, K.C.; Guimaraes, R.C.A.; Castro, A.P.; Nascimento, V.A.; Hiane, P.A. Fatty Acids Consumption: The Role Metabolic Aspects Involved in Obesity and Its Associated Disorders. Nutrients 2017, 9, 1158. [Google Scholar] [CrossRef] [Green Version]
- Tappy, L. Fructose-containing Caloric Sweeteners as a Cause of Obesity and Metabolic Disorders. J. Exp. Biol. 2018, 221 (Suppl. S1). [Google Scholar] [CrossRef] [Green Version]
- Anderson, J.M.; Baird, P.B.; Davis, R.H., Jr.; Ferreri, S.; Knudtson, M.; Koraym, A.; Waters, V.; Williams, C.L. Health Benefits of Dietary Fiber. Nutr. Rev. 2009, 67, 188–205. [Google Scholar] [CrossRef]
- Hosseini, B.; Saedisomeolia, A.; Allman-Farinelli, M. Association between Antioxidant Intake/Status and Obesity: A Systematic Review of Observational. Biol. Trace Elem. Res. 2017, 175, 287–297. [Google Scholar] [CrossRef]
- Mello, A.H.; Uberti, M.F.; Farias, B.X.; Souza, N.A.R.; Rezin, G.T. n-3 PUFA and Obesity: From Peripheral Tissues to the Central Nervous System. Br. J. Nutr. 2018, 119, 1312–1323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, L.; Toumpakari, Z.; Papadaki, A. Social gradients and physical activity trends in an obesogenic dietary pattern: Crosssectional analysis of the UK National Diet and Nutrition Survey 2008–2014. Nutrients 2018, 10, 388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- So, E.; Kim, J.; Joo, S.; Lee, J.S.; Joung, H. Association of dietary patterns with overweight risk and all-cause mortality in children with cancer. Nutr. Res. Pract. 2017, 11, 492–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalenda, A.; Landgraf, K.; Löffler, D.; Kovacs, P.; Kiess, W.; Körner, W. The BDNF Val66Met polymorphism is associated with lower BMI, lower postprandial glucose levels and elevated carbohydrate intake in children and adolescents. Pediatr. Obes. 2017, 13, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Mollahosseini, M.; Rahimi, M.H.; Yekaninejad, M.S.; Maghbooli, Z.; Mirzaei, K. Dietary Patterns Interact With Chromosome 9p21 rs1333048 Polymorphism on the Risk of Obesity and Cardiovascular Risk Factors in Apparently Healthy Tehrani Adults. Eur. J. Nutr. 2020, 59, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Qi, Q.B.; Kilpeläinen, T.O.; Downer, M.K.; Tanaka, T.; Smith, G.E.; Sluijs, I.; Sonestedt, E.; Chu, A.Y.; Renstrom, F.; Lin, X.C.; et al. FTO genetic variants, dietary intake and body mass index: Insights from 177330 individuals. Hum. Mol. Genet. 2014, 23, 6961–6972. [Google Scholar] [CrossRef]
- Astrup, A.; Raben, A.; Geiker, N. The Role of Higher Protein Diets in Weight Control and Obesity-Related Comorbidities. Int. J. Obes. 2015, 39, 721–726. [Google Scholar] [CrossRef] [Green Version]
- Hall, K.D.; Chung, S.T. Low-carbohydrate Diets for the Treatment of Obesity and Type 2 Diabetes. Curr. Opin. Clin. Nutr. Metab. Care 2018, 21, 308–312. [Google Scholar] [CrossRef]
- Burgeiro, A.; Cerqueira, M.G.; Varela-Rodríguez, B.M.; Nunes, S.; Neto, P.; Pereira, F.C.; Reis, F.; Carvalho, E. Glucose and Lipid Dysmetabolism in a Rat Model of Prediabetes Induced by a High-Sucrose Diet. Nutrients 2017, 9, 638. [Google Scholar] [CrossRef]
- Van Rompay, M.I.; McKeown, N.M.; Goodman, E.; Eliaziw, M.; Chomitz, V.R.; Gordon, C.M.; Economos, C.D.; Sacheck, J.M. Sugar-Sweetened Beverage Intake Is Positively Associated with Baseline Triglyceride Concentrations, and Changes in Intake Are Inversely Associated with Changes in HDL Cholesterol over 12 Months in a Multi-Ethnic Sample of Children. J. Nutr. 2015, 145, 2389–2395. [Google Scholar] [CrossRef] [Green Version]
- Shang, X.W.; Liu, A.L.; Zhang, Q.; Hu, X.Q.; Du, S.M.; Ma, J.; Xu, G.F.; Li, Y.; Guo, H.W.; Du, L.; et al. Report on Childhood Obesity in China (9): Sugar-sweetened Beverages Consumption and Obesity. Biomed. Environ. Sci. 2012, 25, 125–132. [Google Scholar] [PubMed]
- Gui, Z.H.; Zhu, Y.N.; Cai, L.; Sun, F.H.; Ma, Y.H.; Jing, J.; Chen, Y.J. Sugar-Sweetened Beverage Consumption and Risks of Obesity and Hypertension in Chinese Children and Adolescents: A National Cross-Sectional Analysis. Nutrients 2017, 9, 1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bitok, E.; Sabaté, J. Nuts and Cardiovascular Disease. Prog. Cardiovasc. Dis. 2018, 61, 33–37. [Google Scholar] [CrossRef] [PubMed]
- Mohammadifard, N.; Haghighatdoost, F.; Mansourian, M.; Hassannejhad, R.; Sadeghi, M.; Roohafza, H.; Sajjadi, F.; Maghroun, M.; Alikhasi, H.; Sarrafzadegan, N. Long-term association of nut consumption and cardiometabolic risk factors. Nutr. Metab. Cardiovasc. Dis. 2019, 29, 972–982. [Google Scholar] [CrossRef] [PubMed]
- Bergeron, N.; Chiu, S.; Williams, P.T.; King, S.M.; Krauss, R.M. Effects of Red Meat, White Meat, and Nonmeat Protein Sources on Atherogenic Lipoprotein Measures in the Context of Low Compared with High Saturated Fat Intake: A Randomized Controlled Trial. Am. J. Clin. Nutr. 2019, 110, 24–33. [Google Scholar] [CrossRef]
- Rouhani, M.H.; Salehi-Abargouei, A.; Surkan, P.J.; Azadbakht, L. Is there a relationship between red or processed meat intake and obesity? A systematic review and meta-analysis of observational studies. Obes. Rev. 2014, 15, 740–748. [Google Scholar] [CrossRef]
- Nour, M.; Lutze, S.A.; Grech, A.; Allman-Farinelli, M. The Relationship between Vegetable Intake and Weight Outcomes: A Systematic Review of Cohort Studies. Nutrients 2018, 10, 1626. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.; Wang, T.; Heianza, Y.; Zheng, Y.; Sun, D.; Kang, J.H.; Pasquale, L.R.; Rimm, E.B.; Manson, J.E.; Hu, F.B.; et al. Habitual Consumption of Long-Chain n-3 PUFAs and Fish Attenuates Genetically Associated Long-Term Weight Gain. Am. J. Clin. Nutr. 2019, 109, 665–673. [Google Scholar] [CrossRef]
- Meng, L.P.; Liu, A.L.; Hu, X.Q.; Zhang, Q.; Du, S.M.; Fang, H.Y.; Ma, J.; Xu, G.F.; Li, Y.; Guo, H.W.; et al. Report on Childhood Obesity in China (10): Association of Sleep Duration with Obesity. Biomed. Environ. Sci. 2012, 25, 133–140. [Google Scholar]
- Krishnan, M.; Shelling, A.N.; Wall, C.R.; Mitchell, E.A.; Murphy, R.; McCowan, L.M.E.; Thompson, J.M.D.; on behalf of the Children of SCOPE Study Group. Gene-by-environment interactions of the CLOCK, PEMT, and GHRELIN loci with average sleep duration in relation to obesity traits using a cohort of 643 New Zealand European children. Sleep Med. 2017, 37, 19–26. [Google Scholar] [CrossRef]
- Vanderwall, C.; Clark, R.R.; Eickhoff, J.; Carrel, A.L. BMI is a poor predictor of adiposity in young overweight and obese children. BMC Pediatr. 2017, 17, 135–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Variables | Normal (n = 1041) | General Overweight/Obesity (n = 251) | p-Value 1 | Normal (n = 1104) | Central Obesity (n = 188) | p-Value 1 |
|---|---|---|---|---|---|---|
| SP (mmHg) | 97.87 ± 9.13 | 104.14 ± 9.95 | <0.001 | 98.16 ± 9.20 | 104.52 ± 10.21 | <0.001 |
| DP (mmHg) | 60.83 ± 6.55 | 63.78 ± 6.86 | <0.001 | 60.98 ± 6.58 | 63.89 ± 6.97 | <0.001 |
| TG (mmol/L) | 0.81 ± 0.48 | 0.92 ± 0.58 | 0.002 | 0.81 ± 0.49 | 0.97 ± 0.51 | <0.001 |
| TCH (mmol/L) | 3.85 ± 0.82 | 3.96 ± 0.68 | 0.063 | 3.86 ± 0.81 | 3.97 ± 0.73 | 0.077 |
| HDL (mmol/L) | 1.43 ± 0.27 | 1.34 ± 0.24 | <0.001 | 1.43 ± 0.27 | 1.31 ± 0.24 | <0.001 |
| LDL (mmol/L) | 2.27 ± 0.64 | 2.41 ± 0.60 | 0.001 | 2.26 ± 0.64 | 2.49 ± 0.58 | <0.001 |
| FBG (mmol/L) | 4.63 ± 0.45 | 4.72 ± 0.42 | 0.005 | 4.64 ± 0.45 | 4.69 ± 0.42 | 0.138 |
| Variables | Male (n = 658) | Female (n = 634) | p-Value |
|---|---|---|---|
| General overweight/obesity rate | 24.17% | 14.51% | <0.001 a |
| Central obesity rate | 16.72% | 12.30% | 0.024 a |
| BMI | 17.08 ± 2.85 | 16.19 ± 2.41 | <0.001 b |
| Waist(cm) | 57.24 ± 8.03 | 54.05 ± 6.60 | <0.001 b |
| WHtR | 0.43 ± 0.05 | 0.41 ± 0.04 | <0.001 b |
| SP (mmHg) | 100.05 ±9.72 | 98.09 ± 9.41 | <0.001 b |
| DP (mmHg) | 61.70 ± 6.76 | 61.09 ± 6.65 | 0.098 b |
| TCH (mmol/L) | 3.85 ± 0.77 | 3.89 ± 0.82 | 0.351 b |
| TG (mmol/L) | 0.82 ± 0.53 | 0.85 ± 0.47 | 0.215 b |
| LDL (mmol/L) | 2.26 ± 0.61 | 2.33 ± 0.66 | 0.039 b |
| HDL (mmol/L) | 1.44 ± 0.28 | 1.39 ± 0.26 | 0.002 b |
| FBG (mmol/L) | 4.72 ± 0.46 | 4.58 ± 0.42 | <0.001 b |
| Variables | AA + GA (n = 465) | GG (n = 193) | P1 | P-Ancova 2 |
|---|---|---|---|---|
| Weight (kg) | 32.04 ± 7.59 | 34.09 ± 8.45 | 0.016 | 0.074 |
| BMI | 17.20 ± 2.77 | 17.98 ± 3.30 | 0.015 | 0.039 |
| Waist (cm) | 58.04 ± 7.77 | 60.12 ± 8.96 | 0.019 | 0.046 |
| WHtR | 0.43 ± 0.05 | 0.44 ± 0.06 | 0.039 | 0.047 |
| SP (mmHg) | 99.45 ± 9.61 | 101.81 ± 9.77 | 0.024 | 0.049 |
| DP (mmHg) | 61.72 ± 6.39 | 62.49 ± 6.20 | 0.259 | 0.347 |
| TG (mmol/L) | 0.81 ± 0.52 | 0.87 ± 0.68 | 0.362 | 0.448 |
| TCH (mmol/L) | 3.87 ± 0.71 | 3.75 ± 0.90 | 0.179 | 0.162 |
| HDL (mmol/L) | 1.45 ± 0.27 | 1.42 ± 0.29 | 0.331 | 0.343 |
| LDL (mmol/L) | 2.26 ± 0.59 | 2.24 ± 0.64 | 0.792 | 0.734 |
| FBG (mmol/L) | 4.75 ± 0.43 | 4.71 ± 0.46 | 0.466 | 0.286 |
| Food Groups | Dietary Patterns | |||
|---|---|---|---|---|
| HBDP | NSDP | AFDP | WDDP | |
| Vegetables | 0.632 | 0.412 | ||
| Eggs | 0.609 | |||
| Tubers | 0.593 | |||
| Dairy | 0.334 | 0.388 | ||
| Nuts | 0.713 | |||
| Pastries | 0.704 | |||
| Sweets | 0.457 | |||
| Pork | 0.703 | |||
| Fish | 0.696 | |||
| Poultry | 0.376 | |||
| Wheaten | 0.765 | |||
| Rice | −0.618 | |||
| Eigenvalues | 1.673 | 1.513 | 1.462 | 1.429 |
| Variance (%) | 8.0 | 7.2 | 7.0 | 6.8 |
| Total variance (%) = 32.4 | ||||
| Quartile 1 | Quartile 2 | Quartile 3 | Quartile 4 | P3 | P-Ancova 4 | |
|---|---|---|---|---|---|---|
| Overweight/obesity incidence rate (%) 1 | ||||||
| HBDP | 29.1 | 27.2 | 21.4 | 22.3 | 0.51 | |
| NSDP | 27.2 | 23.3 | 20.4 | 29.1 | 0.47 | |
| AFDP | 22.3 | 27.2 | 26.2 | 24.3 | 0.86 | |
| WDDP | 26.2 | 25.2 | 23.3 | 25.2 | 0.97 | |
| Central obesity incidence rate (%) 1 | ||||||
| HBDP | 25.3 | 33.3 | 21.3 | 20.0 | 0.26 | |
| NSDP | 28.0 | 29.3 | 20.0 | 22.7 | 0.54 | |
| AFDP | 21.3 | 28.0 | 29.3 | 21.3 | 0.57 | |
| WDDP | 24.0 | 29.3 | 20.0 | 26.7 | 0.63 | |
| Weight (kg) 2 | ||||||
| HBDP | 32.83 ± 7.57 | 32.85 ± 8.08 | 32.37 ± 8.24 | 32.68 ± 7.87 | 0.97 | 0.97 |
| NSDP | 34.31 ± 8.50 | 31.92 ± 7.64 | 32.09 ± 7.15 | 32.40 ± 8.19 | 0.12 | 0.68 |
| AFDP | 31.27 ± 7.01 | 32.80 ± 8.09 | 33.84 ± 8.76 | 32.82 ±7.61 | 0.15 | 0.26 |
| WDDP | 32.61 ± 7.92 | 32.87 ± 8.00 | 32.13 ± 8.03 | 33.12 ± 7.80 | 0.84 | 0.80 |
| BMI 2 | ||||||
| HBDP | 17.52 ± 2.78 | 17.52 ± 3.22 | 17.29 ± 3.03 | 17.45 ± 2.85 | 0.94 | 0.94 |
| NSDP | 17.77 ± 3.15 | 17.34 ± 3.01 | 17.16 ± 2.66 | 17.51 ± 3.03 | 0.52 | 0.69 |
| AFDP | 16.87 ± 2.48 | 17.64 ± 3.33 | 17.79 ± 3.22 | 17.48 ±2.72 | 0.14 | 0.19 |
| WDDP | 17.50 ± 3.05 | 17.52 ± 3.09 | 17.22 ± 2.81 | 17.55 ±2.93 | 0.85 | 0.86 |
| Waist (cm) 2 | ||||||
| HBDP | 59.12 ± 7.66 | 59.02 ± 8.83 | 58.22 ± 8.73 | 58.42 ± 7.62 | 0.83 | 0.85 |
| NSDP | 59.88 ± 8.66 | 58.13 ± 7.98 | 58.15 ± 7.78 | 58.61 ± 8.38 | 0.39 | 0.77 |
| AFDP | 57.22 ± 7.03 | 58.92 ± 8.72 | 60.03 ± 9.20 | 58.61 ± 7.57 | 0.11 | 0.19 |
| WDDP | 58.76 ± 8.24 | 59.36 ± 8.66 | 57.87 ± 7.87 | 58.79 ± 8.09 | 0.64 | 0.55 |
| WHtR 2 | ||||||
| HBDP | 0.43 ± 0.05 | 0.43 ± 0.06 | 0.43 ± 0.05 | 0.43 ± 0.05 | 0.81 | 0.83 |
| NSDP | 0.43 ± 0.05 | 0.43 ± 0.05 | 0.43 ± 0.05 | 0.43 ± 0.05 | 0.81 | 0.79 |
| AFDP | 0.42 ± 0.04 | 0.43 ± 0.06 | 0.44 ± 0.06 | 0.43 ± 0.05 | 0.20 | 0.22 |
| WDDP | 0.43 ± 0.05 | 0.44 ± 0.05 | 0.43 ± 0.05 | 0.43 ± 0.05 | 0.58 | 0.59 |
| SBP (mmHg) 2 | ||||||
| HBDP | 100.07 ± 8.99 | 100.88 ± 10.96 | 98.03 ± 8.58 | 101.76 ± 9.88 | 0.14 | 0.14 |
| NSDP | 100.49 ± 9.55 | 99.34 ± 9.70 | 99.96 ± 9.41 | 100.95 ± 10.22 | 0.68 | 0.72 |
| AFDP | 99.61 ± 9.12 | 98.45 ± 9.34 | 101.39 ± 10.16 | 101.29 ± 10.01 | 0.19 | 0.17 |
| WDDP | 100.87 ± 10.10 | 101.02 ± 10.26 | 98.49 ± 9.36 | 100.36 ± 8.99 | 0.23 | 0.37 |
| DBP (mmHg) 2 | ||||||
| HBDP | 61.64 ± 6.18 | 62.76 ± 6.82 | 61.00 ± 6.23) | 62.43 ± 6.01 | 0.20 | 0.21 |
| NSDP | 62.16 ± 5.79 | 61.65 ± 6.65 | 61.18 ± 6.48) | 62.84 ± 6.35 | 0.29 | 0.32 |
| AFDP | 62.19 ± 6.19 | 61.17 ± 6.74 | 62.23 ± 6.24) | 62.24 ± 6.18 | 0.56 | 0.67 |
| WDDP | 62.26 ± 6.30 | 62.38 ± 5.76 | 60.88 ± 6.55) | 62.31 ± 6.66 | 0.28 | 0.29 |
| TG (mmol/L) 2 | ||||||
| HBDP | 0.77 ± 0.44 | 0.80 ± 0.52 | 0.80 ± 0.51 | 0.95 ± 0.76 | 0.12 | 0.68 |
| NSDP | 0.78 ± 0.45 | 0.78 ± 0.54 | 0.80 ± 0.52 | 0.95 ± 0.73 | 0.11 | 0.18 |
| AFDP | 0.86 ± 0.51 | 0.71 ± 0.45 | 0.86 ± 0.66 | 0.88 ± 0.64 | 0.11 | 0.37 |
| WDDP | 0.83 ± 0.68 | 0.90 ± 0.59 | 0.72 ± 0.45 | 0.86 ± 0.54 | 0.12 | 0.13 |
| TCH (mmol/L) 2 | ||||||
| HBDP | 3.87 ± 0.70 | 3.76 ± 0.71 | 3.91 ± 0.81 | 3.78 ± 0.87 | 0.48 | 0.44 |
| NSDP | 3.86 ± 0.78 | 3.89 ± 0.61 | 3.85 ± 0.81 | 3.72 ± 0.88 | 0.43 | 0.91 |
| AFDP | 3.76 ± 0.76 | 3.82 ± 0.71 | 3.92 ± 0.84 | 3.82 ± 0.79 | 0.57 | 0.37 |
| WDDP | 3.89 ± 0.69 | 3.81 ± 0.82 | 3.91 ± 0.78 | 3.71 ± 0.81 | 0.25 | 0.58 |
| HDL (mmol/L) 2 | ||||||
| HBDP | 1.40 ± 0.25 | 1.44 ± 0.27 | 1.45 ± 0.28 | 1.46 ± 0.30 | 0.47 | 0.29 |
| NSDP | 1.41 ± 0.28 | 1.44 ± 0.29 | 1.45 ± 0.26 | 1.46 ± 0.27 | 0.69 | 0.55 |
| AFDP | 1.45 ± 0.26 | 1.42 ± 0.27 | 1.45 ± 0.29 | 1.44 ± 0.29 | 0.79 | 0.63 |
| WDDP | 1.44 ± 0.27 | 1.41 ± 0.25 | 1.47 ± 0.30 | 1.43 ± 0.27 | 0.50 | 0.47 |
| LDL (mmol/L) 2 | ||||||
| HBDP | 2.26 ± 0.57 | 2.20 ± 0.59 | 2.31 ± 0.66 | 2.23 ± 0.60 | 0.62 | 0.61 |
| NSDP | 2.28 ± 0.67 | 2.23 ± 0.53 | 2.28 ± 0.63 | 2.22 ± 0.59 | 0.85 | 0.92 |
| AFDP | 2.18 ± 0.51 | 2.16 ± 0.61 | 2.36 ± 0.65 | 2.37 ± 0.63 | 0.05 | 0.04 |
| WDDP | 2.26 ± 0.66 | 2.34 ± 0.57 | 2.22 ± 0.64 | 2.19 ± 0.55 | 0.32 | 0.34 |
| FBG (mmol/L) 2 | ||||||
| HBDP | 4.80 ± 0.41 | 4.71 ± 0.44 | 4.73 ± 0.42 | 4.71 ± 0.48 | 0.47 | 0.48 |
| NSDP | 4.61 ± 0.43 | 4.68 ± 0.46 | 4.69 ± 0.43 | 4.77 ± 0.43 | 0.03 | 0.04 |
| AFDP | 4.73 ± 0.42 | 4.73 ± 0.41 | 4.72 ± 0.47 | 4.78 ± 0.45 | 0.80 | 0.70 |
| WDDP | 4.73 ± 0.41 | 4.70 ± 0.47 | 4.76 ± 0.41 | 4.66 ± 0.44 | 0.13 | 0.11 |
| Variable | AA + GA (n = 465) | GG (n = 193) | p-Value 1 |
|---|---|---|---|
| Total energy intake (kcal/d) | 2074.47 ± 474.65 | 2091.57 ± 559.29 | 0.856 |
| Energy from protein (%) | 22.03 ± 5.40 | 24.05 ± 5.45 | 0.043 |
| Energy from fat (%) | 29.09 ± 9.43 | 29.95 ± 9.37 | 0.428 |
| Energy from carbohydrate (%) | 54.08 ± 10.64 | 51.16 ± 10.91 | 0.046 |
| Physical activity time (h/d) | 2.52 ± 1.39 | 2.57 ± 1.42 | 0.766 |
| Sedentary time (h/d) | 4.25 ± 1.28 | 4.21 ± 1.37 | 0.843 |
| Sleep duration (h/d) | 8.85 ± 1.08 | 8.53 ± 1.09 | 0.031 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Q.; Xue, K.; Guo, H.W.; Deng, F.F.; Yang, Y.H. Interaction of the CMTM7 rs347134 Polymorphism with Dietary Patterns and the Risk of Obesity in Han Chinese Male Children. Int. J. Environ. Res. Public Health 2020, 17, 1515. https://doi.org/10.3390/ijerph17051515
Zhu Q, Xue K, Guo HW, Deng FF, Yang YH. Interaction of the CMTM7 rs347134 Polymorphism with Dietary Patterns and the Risk of Obesity in Han Chinese Male Children. International Journal of Environmental Research and Public Health. 2020; 17(5):1515. https://doi.org/10.3390/ijerph17051515
Chicago/Turabian StyleZhu, Qi, Kun Xue, Hong Wei Guo, Fei Fei Deng, and Yu Huan Yang. 2020. "Interaction of the CMTM7 rs347134 Polymorphism with Dietary Patterns and the Risk of Obesity in Han Chinese Male Children" International Journal of Environmental Research and Public Health 17, no. 5: 1515. https://doi.org/10.3390/ijerph17051515
APA StyleZhu, Q., Xue, K., Guo, H. W., Deng, F. F., & Yang, Y. H. (2020). Interaction of the CMTM7 rs347134 Polymorphism with Dietary Patterns and the Risk of Obesity in Han Chinese Male Children. International Journal of Environmental Research and Public Health, 17(5), 1515. https://doi.org/10.3390/ijerph17051515