
Empty Zona Pellucida Only Case: A Critical Review of the Literature

 ,
, 
 , ,
, , 
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
Search Strategy
3. Results
3.1. Definitions
3.2. Effects of ZP Mutations and Polymorphisms
3.3. Defects in ZP Adversely Affects IVF Outcome
3.4. The Role of the Ovarian Stimulation Protocol
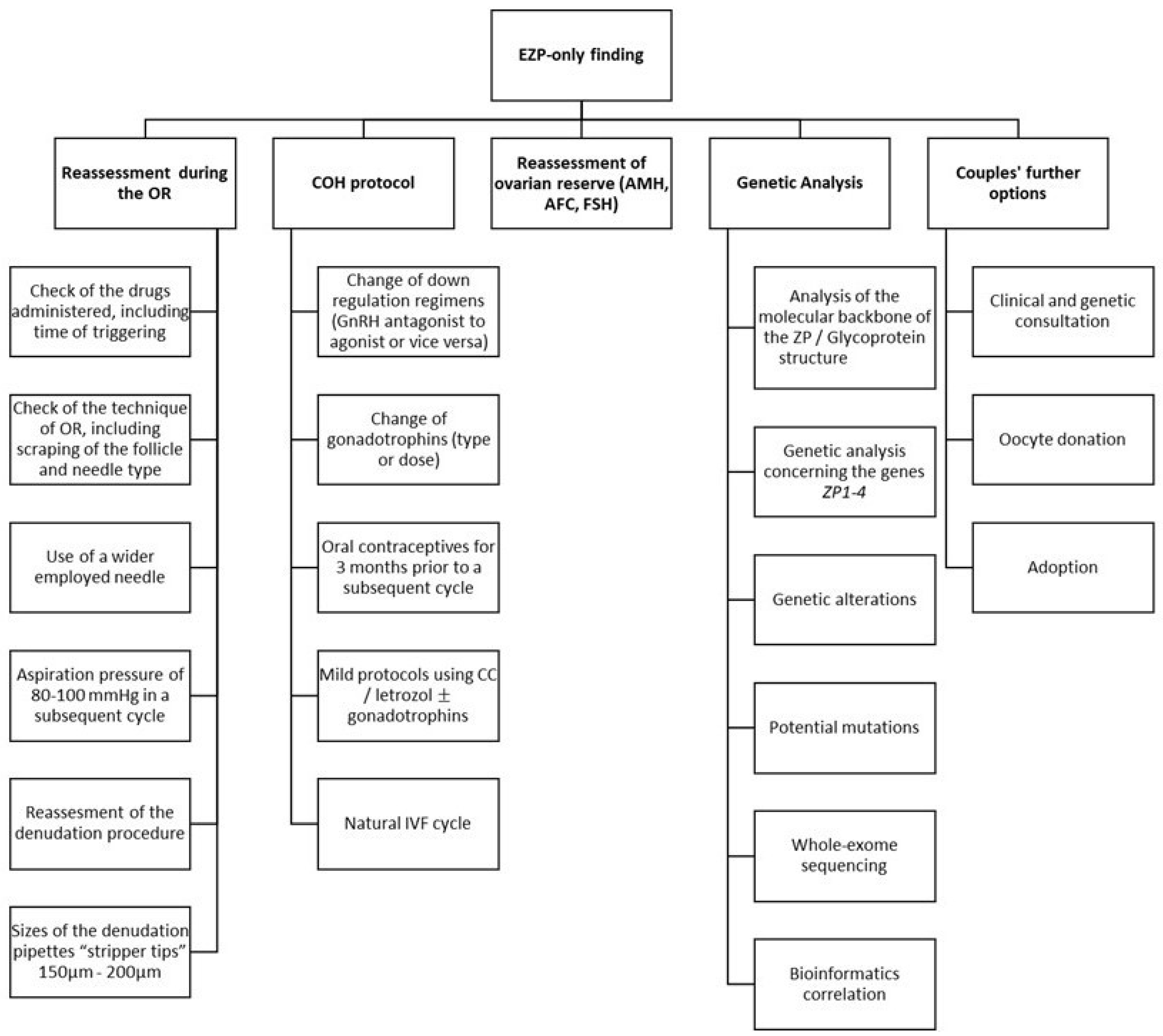
3.5. Management Approaches
3.6. Causative Factors and the Relevant Background
3.7. The Main Recommendation: Genetic Analysis and Bioinformatics
3.8. Further Recommendations
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Conti, M.; Franciosi, F. Acquisition of oocyte competence to develop as an embryo: Integrated nuclear and cytoplasmic events. Hum. Reprod. Update 2018, 24, 245–266. [Google Scholar] [CrossRef]
- Oride, A.; Kanasaki, H.; Hara, T.; Ohta, H.; Kyo, S. Characterization of oocyte retrieval cycles with empty zona pellucida. Reprod. Med. Biol. 2018, 17, 71–76. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Ni, C.; Wu, L.; Chen, B.; Xu, Y.; Zhang, Z.; Mu, J.; Li, B.; Yan, Z.; Fu, J. Novel mutations in ZP1, ZP2, and ZP3 cause female infertility due to abnormal zona pellucida formation. Hum. Genet. 2019, 138, 327–337. [Google Scholar] [CrossRef]
- Shen, Y.; Stalf, T.; Mehnert, C.; Eichenlaub-Ritter, U.; Tinneberg, H.-R. High magnitude of light retardation by the zona pellucida is associated with conception cycles. Hum. Reprod. 2005, 20, 1596–1606. [Google Scholar] [CrossRef]
- Buffone, M.G.; Hirohashi, N.; Gerton, G.L. Unresolved Questions Concerning Mammalian Sperm Acrosomal Exocytosis. Biol. Reprod. 2014, 90, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, M.; Fujiwara, E.; Kakiuchi, Y.; Okabe, M.; Satouh, Y.; Baba, S.A.; Chiba, K.; Hirohashi, N. Most fertilizing mouse spermatozoa begin their acrosome reaction before contact with the Zona Pellucida during in vitro fertilization. Proc. Natl. Acad. Sci. USA 2011, 108, 4892–4896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pökkylä, R.-M.; Lakkakorpi, J.T.; Nuojua-Huttunen, S.H.; Tapanainen, J.S. Sequence variations in human ZP genes as potential modifiers of zona pellucida architecture. Fertil. Steril. 2011, 95, 2669–2672. [Google Scholar] [CrossRef]
- Cao, Q.; Zhao, C.; Zhang, X.; Zhang, H.; Lu, Q.; Wang, C.; Hu, Y.; Ling, X.; Zhang, J.; Huo, R. Heterozygous mutations in ZP1 and ZP3 cause formation disorder of ZP and female infertility in human. J. Cell. Mol. Med. 2020, 24, 8557–8566. [Google Scholar] [CrossRef]
- Cinar, O.; Demir, B.; Dilbaz, S.; Saltek, S.; Aydin, S.; Goktolga, U. Does empty zona pellucida indicate poor ovarian response on intra cytoplasmic sperm injection cycles? Gynecol. Endocrinol. 2012, 28, 341–344. [Google Scholar] [CrossRef] [PubMed]
- Bols, P.; Van Soom, A.; Ysebaert, M.; Vandenheede, J.; de Kruif, A. Effects of aspiration vacuum and needle diameter on cumulus oocyte complex morphology and developmental capacity of bovine oocytes. Theriogenology 1996, 45, 1001–1014. [Google Scholar] [CrossRef]
- Atzmon, Y.; Michaeli, M.; Poltov, D.; Rotfarb, N.; Lebovitz, O.; Aslih, N.; Shalom-Paz, E. Degenerated oocyte in the cohort adversely affects IVF outcome. J. Ovarian Res. 2020, 13, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lazzaroni-Tealdi, E.; Barad, D.H.; Albertini, D.F.; Yu, Y.; Kushnir, V.A.; Russell, H.; Wu, Y.-G.; Gleicher, N. Oocyte scoring enhances embryo-scoring in predicting pregnancy chances with IVF where it counts most. PLoS ONE 2015, 10, e0143632. [Google Scholar] [CrossRef] [PubMed]
- Serhal, P.; Ranieri, D.; Kinis, A.; Marchant, S.; Davies, M.; Khadum, I. Oocyte morphology predicts outcome of intracytoplasmic sperm injection. Hum. Reprod. 1997, 12, 1267–1270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, P. Intracytoplasmic sperm injection: Correlation of oocyte grade based on polar body, perivitelline space and cytoplasmic inclusions with fertilization rate and embryo quality. Hum. Reprod. 1997, 12, 1750–1755. [Google Scholar] [CrossRef]
- Gupta, S.K. The human egg’s zona pellucida. Curr. Top. Dev. Biol. 2018, 130, 379–411. [Google Scholar]
- Lefièvre, L.; Conner, S.; Salpekar, A.; Olufowobi, O.; Ashton, P.; Pavlovic, B.; Lenton, W.; Afnan, M.; Brewis, I.A.; Monk, M. Four zona pellucida glycoproteins are expressed in the human. Hum. Reprod. 2004, 19, 1580–1586. [Google Scholar] [CrossRef] [Green Version]
- Ferré, M.; Amati-Bonneau, P.; Morinière, C.; Ferré-L’Hôtellier, V.; Lemerle, S.; Przyrowski, D.; Procaccio, V.; Descamps, P.; Reynier, P.; May-Panloup, P. Are zona pellucida genes involved in recurrent oocyte lysis observed during in vitro fertilization? J. Assist. Reprod. Genet. 2014, 31, 221–227. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Shen, Y.; Zhang, X.; Wang, X.; Li, D.; Wang, Y. Novel biallelic loss-of-function variants in ZP1 identified in an infertile female with empty follicle syndrome. J. Assist. Reprod. Genet. 2020, 37, 2151–2157. [Google Scholar] [CrossRef]
- Huang, H.-L.; Lv, C.; Zhao, Y.-C.; Li, W.; He, X.-M.; Li, P.; Sha, A.-G.; Tian, X.; Papasian, C.J.; Deng, H.-W. Mutant ZP1 in familial infertility. N. Engl. J. Med. 2014, 370, 1220–1226. [Google Scholar] [CrossRef] [Green Version]
- Wassarman, P.M.; Qi, H.; Litscher, E.S. Mutant female mice carrying a single mZP3 allele produce eggs with a thin Zona Pellucida, but reproduce normally. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1997, 264, 323–328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gook, D.A.; Edgar, D.; Borg, J.; Martic, M. Detection of zona pellucida proteins during human folliculogenesis. Hum. Reprod. 2008, 23, 394–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.; Bian, Y.; Liu, X.; Zhao, S.; Wu, K.; Yan, L.; Li, M.; Yang, Z.; Liu, H.; Zhao, H. A recurrent missense mutation in ZP3 causes empty follicle syndrome and female infertility. Am. J. Hum. Genet. 2017, 101, 459–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wassarman, P.M.; Jovine, L.; Litscher, E.S. A profile of fertilization in mammals. Nat. Cell Biol. 2001, 3, E59–E64. [Google Scholar] [CrossRef] [PubMed]
- Conner, S.; Lefièvre, L.; Hughes, D.; Barratt, C. Cracking the egg: Increased complexity in the zona pellucida. Hum. Reprod. 2005, 20, 1148–1152. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.K. Role of Zona Pellucida glycoproteins during fertilization in humans. J. Reprod. Immunol. 2015, 108, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.; Hu, L.; Gong, F.; Tan, Y.; Cai, S.; Zhang, S.; Dai, J.; Lu, C.; Chen, J.; Chen, Y. ZP2 pathogenic variants cause in vitro fertilization failure and female infertility. Genet. Med. 2019, 21, 431–440. [Google Scholar] [CrossRef]
- Zhang, D.; Zhu, L.; Liu, Z.; Ren, X.; Yang, X.; Li, D.; Luo, Y.; Peng, X.; Zhou, X.; Jia, W. A novel mutation in ZP3 causes empty follicle syndrome and abnormal Zona Pellucida formation. J. Assist. Reprod. Genet. 2021, 38, 251–259. [Google Scholar] [CrossRef]
- Luo, G.; Zhu, L.; Liu, Z.; Yang, X.; Xi, Q.; Li, Z.; Duan, J.; Jin, L.; Zhang, X. Novel mutations in ZP1 and ZP2 cause primary infertility due to empty follicle syndrome and abnormal zona pellucida. J. Assist. Reprod. Genet. 2020, 37, 2853–2860. [Google Scholar] [CrossRef]
- GeneCards: The Human Gene Database. Available online: https://www.genecards.org/ (accessed on 24 May 2021).
- Female Infertility Due to Zona Pellucida Defect. Available online: https://www.malacards.org/card/female_infertility_due_to_zona_pellucida_defect (accessed on 24 May 2021).
- Sousa, M.; da Silva, J.T.; Silva, J.; Cunha, M.; Viana, P.; Oliveira, E.; Sá, R.; Soares, C.; Oliveira, C.; Barros, A. Embryological, clinical and ultrastructural study of human oocytes presenting indented zona pellucida. Zygote 2015, 23, 145. [Google Scholar] [CrossRef]
- Monné, M.; Han, L.; Schwend, T.; Burendahl, S.; Jovine, L. Crystal structure of the ZP-N domain of ZP3 reveals the core fold of animal egg coats. Nature 2008, 456, 653–657. [Google Scholar] [CrossRef]
- Markstrom, E.; Svensson, E.; Shao, R.; Svanberg, B.; Billig, H. Survival factors regulating ovarian apoptosis--dependence on follicle differentiation. Reproduction 2002, 123, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Quirk, S.; Cowan, R.; Harman, R.; Hu, C.-L.; Porter, D. Ovarian follicular growth and atresia: The relationship between cell proliferation and survival. J. Anim. Sci. 2004, 82, E40–E52. [Google Scholar] [CrossRef]
- Høst, E.; Gabrielsen, A.; Lindenberg, S.; Smidt-Jensen, S. Apoptosis in human cumulus cells in relation to zona pellucida thickness variation, maturation stage, and cleavage of the corresponding oocyte after intracytoplasmic sperm injection. Fertil. Steril. 2002, 77, 511–515. [Google Scholar] [CrossRef]
- Stevenson, T.L.; Lashen, H. Empty follicle syndrome: The reality of a controversial syndrome, a systematic review. Fertil. Steril. 2008, 90, 691–698. [Google Scholar] [CrossRef]
- Vutyavanich, T.; Piromlertamorn, W.; Ellis, J. Immature Oocytes in “Apparent Empty Follicle Syndrome”: A Case Report. Case Rep. Med. 2010, 2010, 367505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jancar, N.; Virant-Klun, I.; Osredkar, J.; Bokal, E.V. Apoptosis, Reactive Oxygen Species and Follicular Anti-Müllerian Hormone in Natural versus Stimulated Cycles. Reprod. Biomed. Online 2008, 16, 640–648. [Google Scholar] [CrossRef]
- Shi, W.; Xu, B.; Wu, L.-M.; Jin, R.-T.; Luan, H.-B.; Luo, L.-H.; Zhu, Q.; Johansson, L.; Liu, Y.-S.; Tong, X.-H. Oocytes with a dark zona pellucida demonstrate lower fertilization, implantation and clinical pregnancy rates in IVF/ICSI cycles. PLoS ONE 2014, 9, e89409. [Google Scholar] [CrossRef] [Green Version]
- Detti, L.; Ambler, D.R.; Yelian, F.D.; Kruger, M.L.; Diamond, M.P.; Puscheck, E.E. Timing and duration of use of GnRH antagonist down-regulation for IVF/ICSI cycles have no impact on oocyte quality or pregnancy outcomes. J. Assist. Reprod. Genet. 2008, 25, 177–181. [Google Scholar] [CrossRef] [Green Version]
- Siriaroonrat, B.; Comizzoli, P.; Songsasen, N.; Monfort, S.; Wildt, D.; Pukazhenthi, B. Oocyte quality and estradiol supplementation affect in vitro maturation success in the white-tailed deer (Odocoileus virginianus). Theriogenology 2010, 73, 112–119. [Google Scholar] [CrossRef]
- Visser, J.A.; Durlinger, A.L.; Peters, I.J.; van den Heuvel, E.R.; Rose, U.M.; Kramer, P.; de Jong, F.H.; Themmen, A.P. Increased oocyte degeneration and follicular atresia during the estrous cycle in anti-Mullerian hormone null mice. Endocrinology 2007, 148, 2301–2308. [Google Scholar] [CrossRef] [Green Version]
- Horne, R.; Bishop, C.J.; Reeves, G.; Wood, C.; Kovacs, G.T. Aspiration of oocytes for in-vitro fertilization. Hum. Reprod. Update 1996, 2, 77–85. [Google Scholar] [CrossRef] [Green Version]
- Veeck, L.L. An Atlas of Human Gametes and Conceptuses: An Illustrated Reference for Assisted Reproductive Technology, 1st ed.; CC Press: Boca Raton, FL, USA, 1999. [Google Scholar]
- Normand, E.A.; Alaimo, J.T.; Van den Veyver, I.B. Exome and genome sequencing in reproductive medicine. Fertil. Steril. 2018, 109, 213–220. [Google Scholar] [CrossRef] [Green Version]
- Altmäe, S.; Martinez-Conejero, J.A.; Esteban, F.J.; Ruiz-Alonso, M.; Stavreus-Evers, A.; Horcajadas, J.A.; Salumets, A. MicroRNAs miR-30b, miR-30d, and miR-494 regulate human endometrial receptivity. Reprod. Sci. 2014, 20, 308–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siristatidis, C.; Stavros, S.; Drakeley, A.; Bettocchi, S.; Pouliakis, A.; Drakakis, P.; Papapanou, M.; Vlahos, N. Omics and Artificial Intelligence to Improve In Vitro Fertilization (IVF) Success: A Proposed Protocol. Diagnostics 2021, 11, 743. [Google Scholar] [CrossRef] [PubMed]
- Ouni, E.; Vertommen, D.; Amorim, C.A. The Human Ovary and Future of Fertility Assessment in the Post-Genome Era. Int. J. Mol. Sci. 2019, 20, 4209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Askoxylaki, M.; Siristatidis, C.; Chrelias, C.; Vogiatzi, P.; Creatsa, M.; Salamalekis, G.; Vrantza, T.; Vrachnis, N.; Kassanos, D. Reactive oxygen species in the follicular fluid of subfertile women undergoing in vitro fertilization: A short narrative review. J. Endocrinol. Investig. 2013, 36, 1117–1120. [Google Scholar]
- Siristatidis, C.; Askoxylaki, M.; Varounis, C.; Kassanos, D.; Chrelias, C. E-selectin, resistin and reactive oxygen species levels in GnRH-agonist and-antagonist protocols in IVF/ICSI: A prospective cohort study. J. Assist. Reprod. Genet. 2015, 32, 959–967. [Google Scholar] [CrossRef] [Green Version]
- Siristatidis, C.; Vogiatzi, P.; Varounis, C.; Askoxylaki, M.; Chrelias, C.; Papantoniou, N. The effect of reactive oxygen species on embryo quality in IVF. In Vivo 2016, 30, 149–153. [Google Scholar] [PubMed]
- Agarwal, A.; Gupta, S.; Sharma, R. Oxidative Stress and Its Implications in Female Infertility—A Clinician’s Perspective. Reprod. Biomed. Online 2005, 11, 641–650. [Google Scholar] [CrossRef]
- Lu, J.; Wang, Z.; Cao, J.; Chen, Y.; Dong, Y. A Novel and Compact Review on the Role of Oxidative Stress in Female Reproduction. Reprod. Biol. Endocrinol. 2018, 16, 1–18. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siristatidis, C.; Tzanakaki, D.; Simopoulou, M.; Vaitsopoulou, C.; Tsioulou, P.; Stavros, S.; Papapanou, M.; Drakakis, P.; Bakas, P.; Vlahos, N. Empty Zona Pellucida Only Case: A Critical Review of the Literature. Int. J. Environ. Res. Public Health 2021, 18, 9409. https://doi.org/10.3390/ijerph18179409
Siristatidis C, Tzanakaki D, Simopoulou M, Vaitsopoulou C, Tsioulou P, Stavros S, Papapanou M, Drakakis P, Bakas P, Vlahos N. Empty Zona Pellucida Only Case: A Critical Review of the Literature. International Journal of Environmental Research and Public Health. 2021; 18(17):9409. https://doi.org/10.3390/ijerph18179409
Chicago/Turabian StyleSiristatidis, Charalampos, Despoina Tzanakaki, Mara Simopoulou, Christina Vaitsopoulou, Petroula Tsioulou, Sofoklis Stavros, Michail Papapanou, Peter Drakakis, Panagiotis Bakas, and Nikolaos Vlahos. 2021. "Empty Zona Pellucida Only Case: A Critical Review of the Literature" International Journal of Environmental Research and Public Health 18, no. 17: 9409. https://doi.org/10.3390/ijerph18179409
APA StyleSiristatidis, C., Tzanakaki, D., Simopoulou, M., Vaitsopoulou, C., Tsioulou, P., Stavros, S., Papapanou, M., Drakakis, P., Bakas, P., & Vlahos, N. (2021). Empty Zona Pellucida Only Case: A Critical Review of the Literature. International Journal of Environmental Research and Public Health, 18(17), 9409. https://doi.org/10.3390/ijerph18179409