
The Positive Effects of Poria cocos Extract on Quality of Sleep in Insomnia Rat Models

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Drugs Used and Laboratory Animals
2.1.1. Poria cocos Extract
2.1.2. Drugs Used in the Experiment
2.1.3. Laboratory Animals
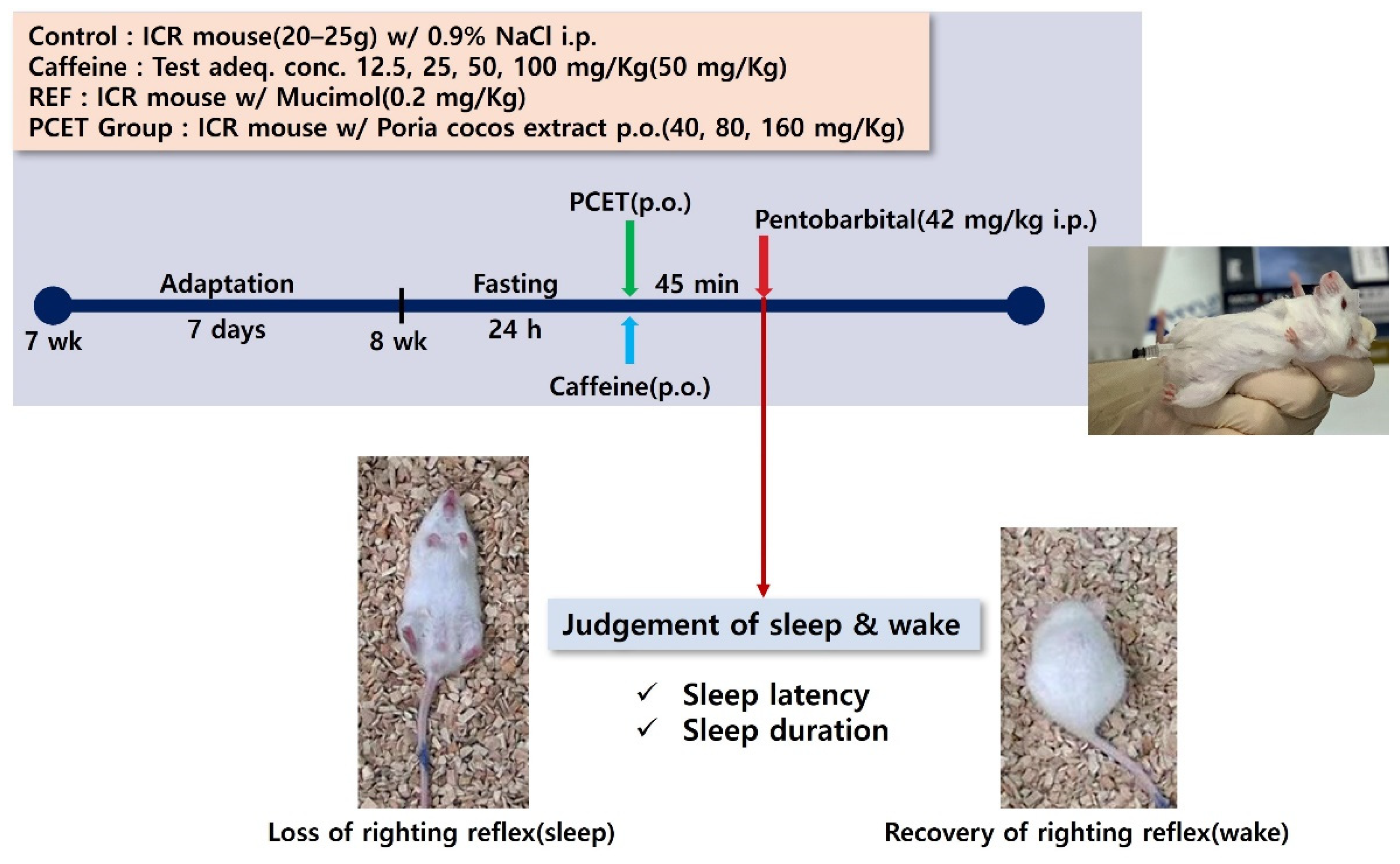
2.1.4. Sleep Induction Test and Sleep Disorder Animal Model Development
2.2. Separation of the Superior Cervical Ganglion
2.3. Cl− Influx Test and GABA-Induced Cl− Current Measurement
2.4. Statistical Analysis
3. Results
3.1. Strengthening of GABA Receptor Actions in the Superior Cervical Ganglion of Poria cocos Extract
3.1.1. Increased Cl− Influx Induced by Poria cocos Extract in Superior Cervical Ganglion Cells
3.1.2. Enhancement of GABA-Induced Cl− Current by Poria cocos Extract in Superior Cervical Ganglion Cells
3.2. Evaluation of Sleep-Promoting Efficacy of Poria cocos Extract
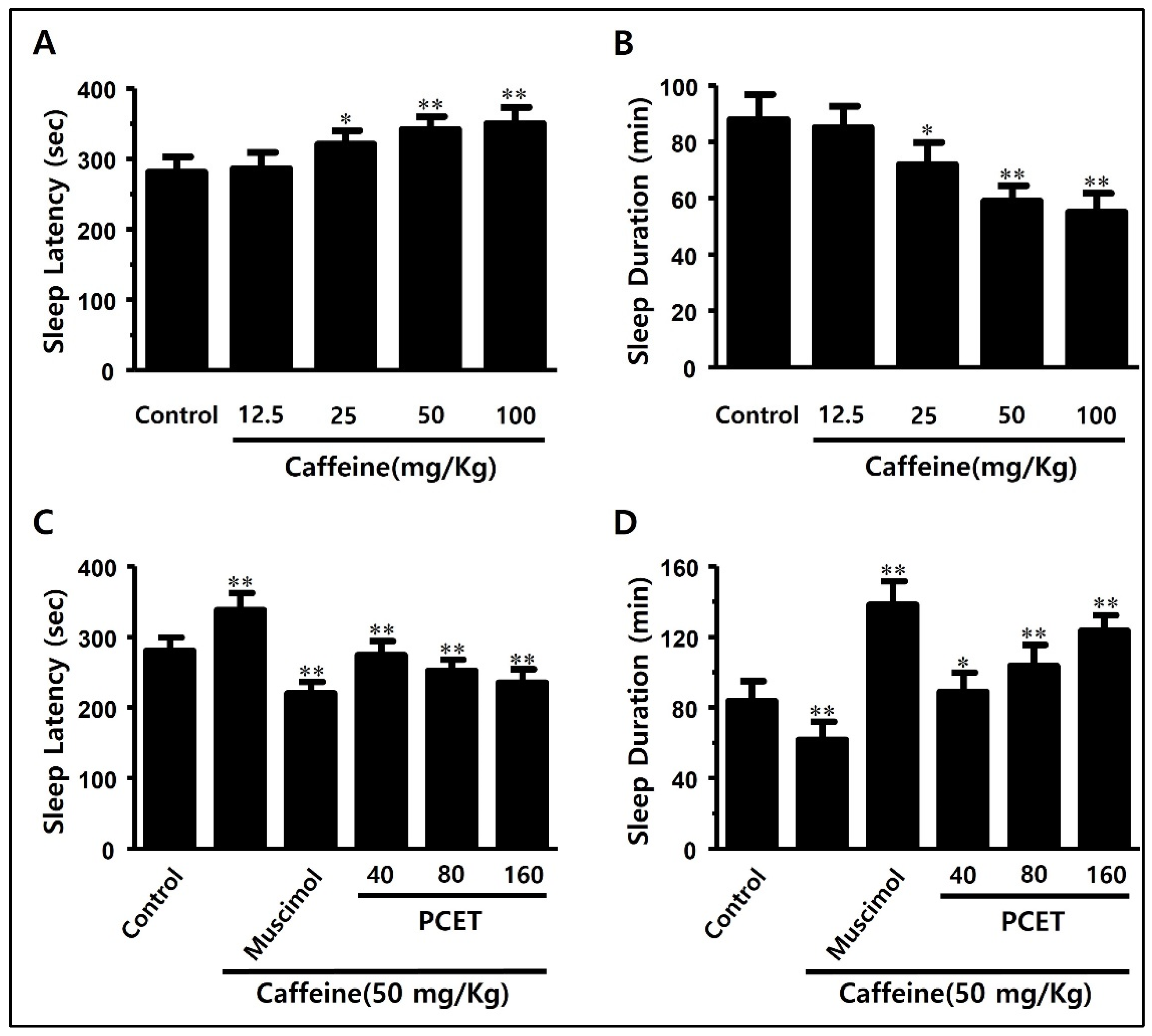
3.2.1. Evaluation of Sleep-Promoting Efficacy of Poria cocos Extract in Normal Mice
3.2.2. Evaluation of Arousal Inhibitory Effect of Poria cocos Extract in Caffeine-Induced Arousal-Stimulating Mice
3.3. Evaluation of PCET Sleep Improvement Effect on ACTH-Induced Sleep Disorder Model
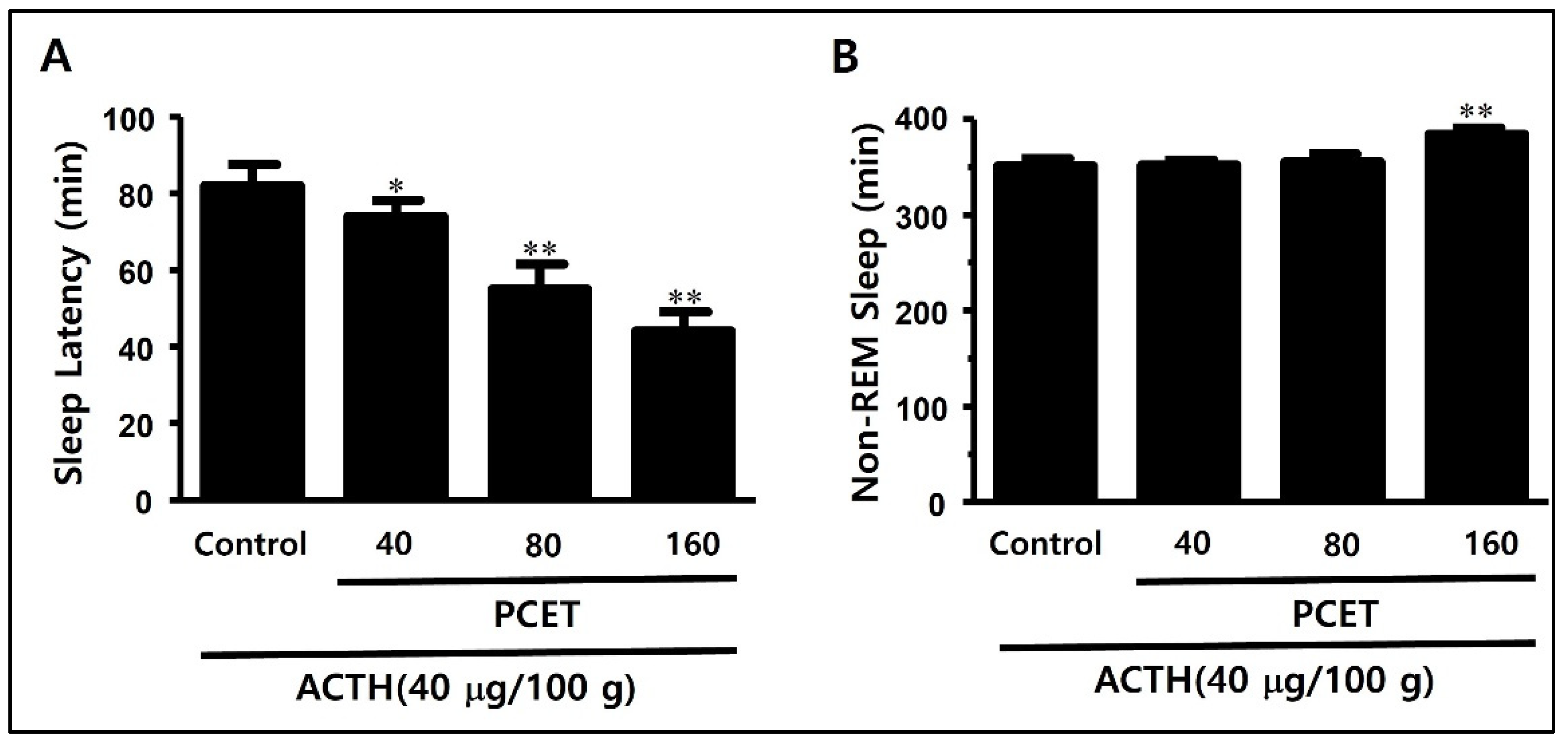
3.3.1. Inducing Sleep Disorder Using Subcutaneous ACTH Injection
3.3.2. Sleep Structure Improvement Effect of PCET in the ACTH-Induced Sleep Disorder Model
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jahrami, H.; BaHammam, A.S.; Bragazzi, N.L.; Saif, Z.; Faris, M.; Vitiello, M.V. Sleep problems during the COVID-19 pandemic by population: A systemic review and meta-analysis. J. Clin. Sleep Med. 2021, 17, 299–313. [Google Scholar] [CrossRef] [PubMed]
- Becker, P.M. Overview of sleep management during COVID-19. Sleep Med. 2022, 91, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Harris, C.D. Neurophysiology of sleep and wakefulness. Respir. Care Clin. N. Am. 2005, 11, 567–586. [Google Scholar] [PubMed]
- Zammit, G.K.; Weiner, J.; Damato, N.; Sillup, G.P.; McMillan, C.A. Quality of life in people with insomnia. Sleep 1999, 22 (Suppl. S2), S379–S385. [Google Scholar]
- Bibek, K.; Vikas, G. Sleep Disorder. In StatPearls [Internet]; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Proserpio, P.; Terzaghi, M.; Manni, R.; Nobili, L. Drugs used in parasomnia. Sleep Med. Clin. 2020, 15, 289–300. [Google Scholar] [CrossRef]
- Van Sweden, B. Sleep and the temporal lobe. Acta Neurol. Belg. 1996, 96, 19–30. [Google Scholar]
- Malhotra, R.K. Neurodegenerative disorders and sleep. Sleep Med. Clin. 2018, 13, 63–70. [Google Scholar] [CrossRef]
- Gao, Q.; Kou, T.; Zhuang, B.; Ren, Y.; Dong, X.; Wang, Q. The association between Vitamin D deficiency and sleep disorders: A systematic review and meta-analysis. Nutrients 2018, 10, 1395. [Google Scholar] [CrossRef] [Green Version]
- Berk, M. Sleep and depression—Theory and practice. Aust. Fam. Physician 2009, 38, 302–304. [Google Scholar]
- Rosenberg, R.P. Sleep maintenance insomnia: Strengths and weaknesses of current pharmacologic therapies. Ann. Clin. Psychiatry 2006, 18, 49–56. [Google Scholar] [CrossRef]
- Wafford, K.A.; Ebert, B. Emerging anti-insomnia drugs: Tackling sleeplessness and the quality of wake time. Nat. Rev. Drug Discov. 2008, 7, 530–540. [Google Scholar] [CrossRef] [PubMed]
- Barnard, E.A.; Skolnick, P.; Olsen, R.W.; Mohler, H.; Sieghart, W.; Biggio, G.; Braestrup, C.; Bateson, A.N.; Langer, S.Z. International Union of Pharmacology. XV. Subtypes of gamma-aminobutyric acidA receptors: Classification on the basis of subunit structure and receptor function. Pharmacol. Rev. 1998, 50, 291–313. [Google Scholar] [PubMed]
- Lu, J.; Karadsheh, M.; Delpire, E. Developmental regulation of the neuronal-specific isoform of K-Cl cotransporter KCC2 in postnatal rat brains. J. Neurobiol. 1999, 39, 558–568. [Google Scholar] [CrossRef]
- Sieghart, W.; Sperk, G. Subunit composition, distribution and function of GABA(A) receptor subtypes. Curr. Top. Med. Chem. 2002, 2, 795–816. [Google Scholar] [CrossRef] [PubMed]
- Shah, V.K.; Choi, J.J.; Han, J.Y.; Lee, M.K.; Hong, J.T.; Oh, K.W. Pachymic acid enhances pentobarbital-induced sleeping behaviors via GABAA-ergic systems in mice. Biomol. Ther. 2014, 22, 314–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spelman, K.; Burns, J.; Nichols, D.; Winters, N.; Ottersberg, S.; Tenborg, M. Modulation of cytokine expression by traditional medicines: A review of herbal immunomodulators. Altern. Med. Rev. 2006, 11, 128–150. [Google Scholar]
- Kaminaga, T.; Yasukawa, K.; Kanno, H.; Tai, T.; Nunoura, Y.; Takido, M. Inhibitory effects of lanostane-type triterpene acids, the components of Poria cocos, on tumor promotion by 12-O-tetradecanoylphorbol-13-acetate in two-stage carcinogenesis in mouse skin. Oncology 1996, 53, 382–385. [Google Scholar] [CrossRef]
- Tai, T.; Akita, Y.; Kinoshita, K.; Koyama, K.; Takahashi, K.; Watanabe, K. Anti-emetic principles of Poria cocos. Planta Med. 1995, 61, 527–530. [Google Scholar] [CrossRef]
- Cuélla, M.J.; Giner, R.M.; Recio, M.C.; Just, M.J.; Máñez, S.; Ríos, J.L. Two fungal lanostane derivatives as phospholipase A2 inhibitors. J. Nat. Prod. 1996, 59, 977–979. [Google Scholar] [CrossRef]
- Shah, V.K.; Na, S.S.; Chong, M.S.; Woo, J.H.; Kwon, Y.O.; Lee, M.K.; Oh, K.W. Poria cocos ethanol extract and its active constituent, pachymic acid, modulate sleep architectures via activation of GABAA-ergic transmission in rats. J. Biomed. Res. 2015, 16, 84–92. [Google Scholar] [CrossRef]
- Zhu, Y.; Zboran, E.L.; Ikeda, S.R. Phenotype-specific expression of T-type calcium channels in neurons of the major pelvic ganglion of the adult male rat. J. Physiol. 1995, 489 Pt 2, 363–375. [Google Scholar] [CrossRef]
- West, M.R.; Molloy, C.R. A microplate assay measuring chloride ion channel activity. Anal. Biochem. 1996, 241, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Hamill, O.P.; Marty, A.; Neher, E.; Sakmann, B.; Sigworth, F.J. Improved patch-clamp techniques for high-resolution current recording from cells and cell-free membrane patches. Pflugers Arch. 1981, 391, 85–100. [Google Scholar] [CrossRef] [PubMed]
- Farkas, Z.; Kása, P.; Balcar, V.J.; Joó, F.; Wolff, J.R. Type A and B gaba receptors mediate inhibition of acetylcholine release from cholinergic nerve terminals in the superior cervical ganglion of rat. Neurochem. Int. 1986, 8, 565–572. [Google Scholar] [CrossRef]
- Revel, F.G.; Gottowik, J.; Gatti, S.; Wettstein, J.G.; Moreau, J.L. Rodent models of insomnia: A review of experimental procedures that induce sleep disturbances. Neurosci. Biobehav. Rev. 2009, 33, 874–899. [Google Scholar] [CrossRef] [PubMed]
- Wheatley, D. Medicinal plants for insomnia: A review of their pharmacology, efficacy and tolerability. J. Psychopharmacol. 2005, 19, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.M.; Zhao, Z.; Stock, H.S.; Mehl, K.A.; Buggy, J.; Hand, G.A. Central nervous system effects of caffeine and adenosine on fatigue. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2003, 284, R399–R404. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.L.; Qu, W.M.; Eguchi, N.; Chen, J.F.; Schwarzschild, M.A.; Fredholm, B.B.; Urade, Y.; Hayaishi, O. Adenosine A2A, but not A1, receptors mediate the arousal effect of caffeine. Nat. Neurosci. 2005, 8, 858–859. [Google Scholar] [CrossRef]
- Strassnig, M.; Brar, J.S.; Ganguli, R. Increased caffeine and nicotine consumption in community-dwelling patients with schizophrenia. Schizophr. Res. 2006, 86, 269–275. [Google Scholar] [CrossRef]
- Noachtar, S.; Rémi, J. The role of EEG in epilepsy: A critical review. Epilepsy Behav. 2009, 15, 22–33. [Google Scholar] [CrossRef]
- Appavu, B.; Fox, J.; Little, R.; Kerrigan, J.F.; Obeid, M. Seizures induced by exiting water: A unique form of reflex epilepsy. Neurologist 2017, 22, 196–198. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, Y.; Araki, H.; Gomita, Y. Influence of ACTH on the effects of imipramine, desipramine and lithium on duration of immobility of rats in the forced swim test. Pharmacol. Biochem. Behav. 2002, 71, 63–69. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.; Park, I.; Park, K.; Park, S.; Kim, Y.I.; Park, B.-G. The Positive Effects of Poria cocos Extract on Quality of Sleep in Insomnia Rat Models. Int. J. Environ. Res. Public Health 2022, 19, 6629. https://doi.org/10.3390/ijerph19116629
Kim H, Park I, Park K, Park S, Kim YI, Park B-G. The Positive Effects of Poria cocos Extract on Quality of Sleep in Insomnia Rat Models. International Journal of Environmental Research and Public Health. 2022; 19(11):6629. https://doi.org/10.3390/ijerph19116629
Chicago/Turabian StyleKim, Hyeyun, Injune Park, Kyunyong Park, Seohyun Park, Yeong In Kim, and Byong-Gon Park. 2022. "The Positive Effects of Poria cocos Extract on Quality of Sleep in Insomnia Rat Models" International Journal of Environmental Research and Public Health 19, no. 11: 6629. https://doi.org/10.3390/ijerph19116629
APA StyleKim, H., Park, I., Park, K., Park, S., Kim, Y. I., & Park, B. -G. (2022). The Positive Effects of Poria cocos Extract on Quality of Sleep in Insomnia Rat Models. International Journal of Environmental Research and Public Health, 19(11), 6629. https://doi.org/10.3390/ijerph19116629