
Succession of the Abandoned Rice Fields Restores the Riparian Forest

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
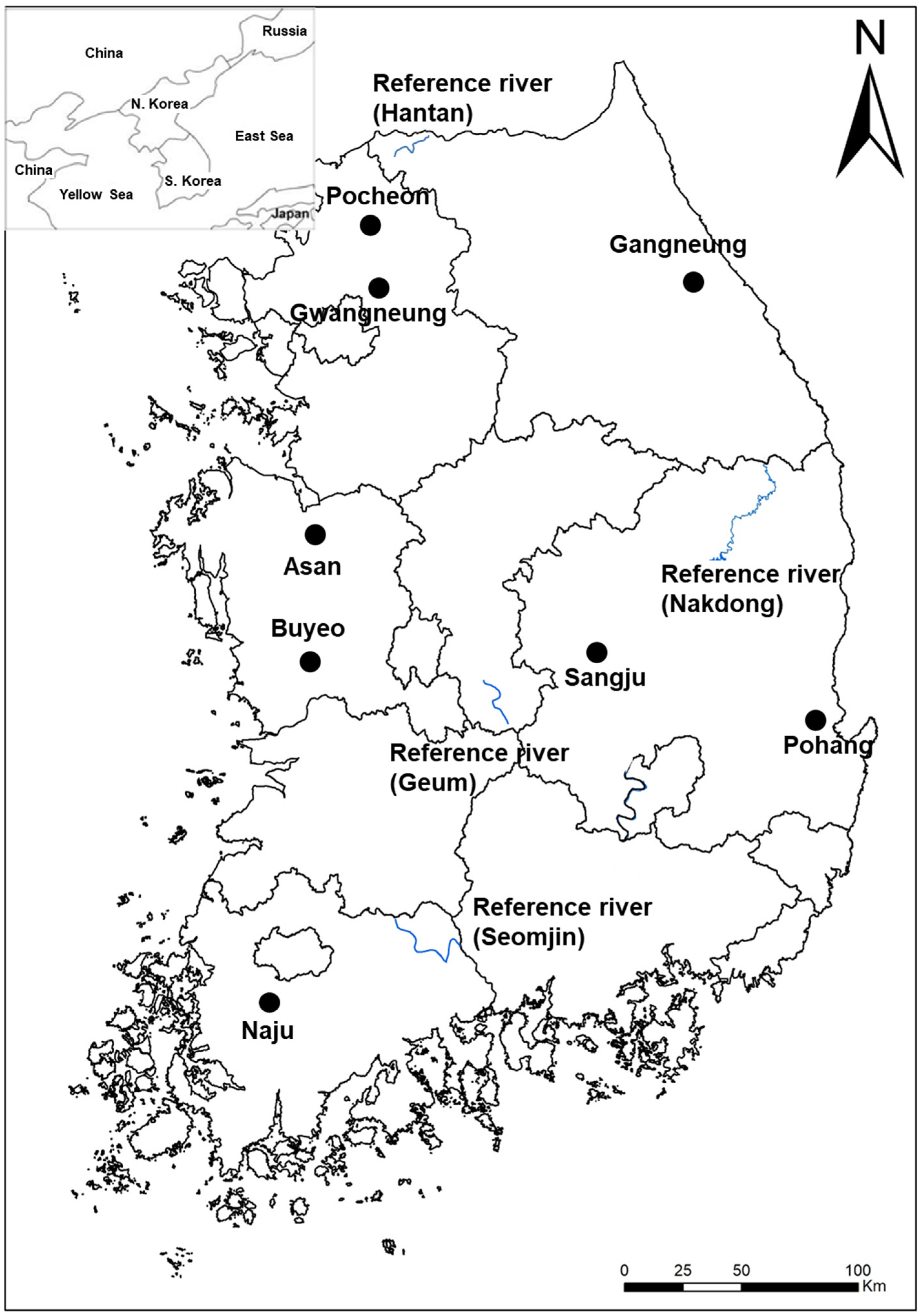
2.1. Site Description
2.2. Vegetation Survey
2.3. Systematization of Collected Vegetation Data into Reference Information
2.4. Statistical Analyses
3. Results
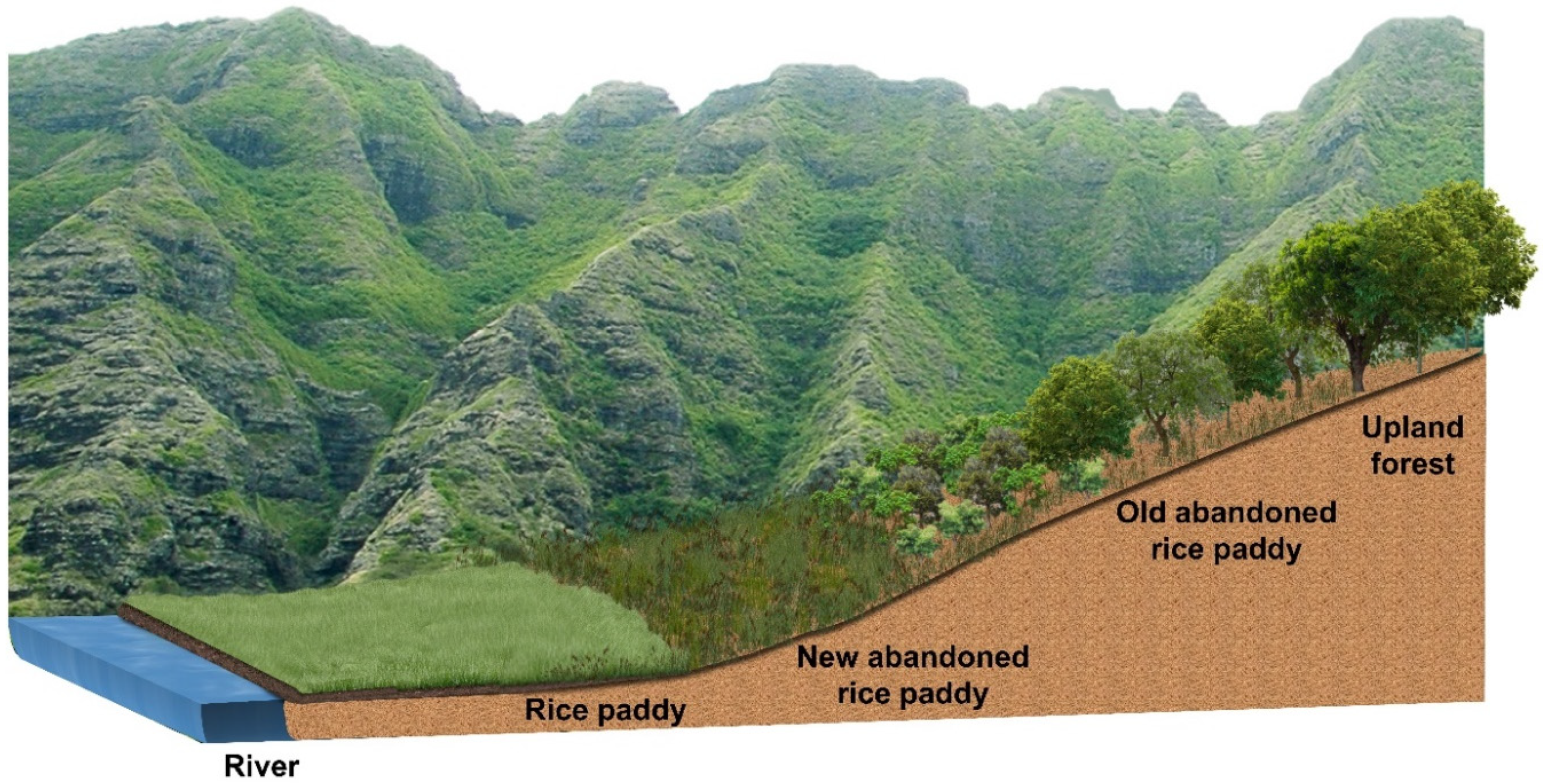
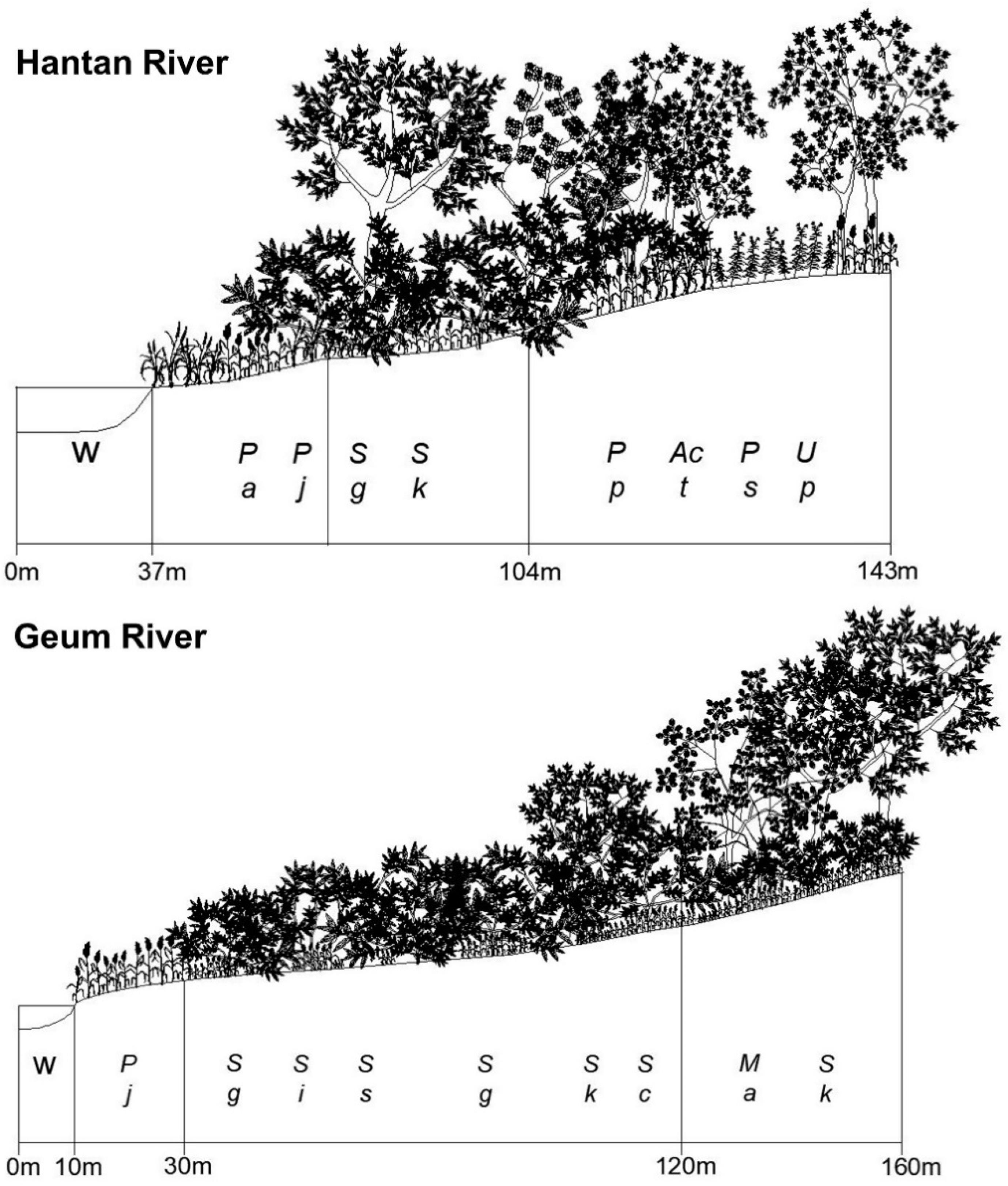
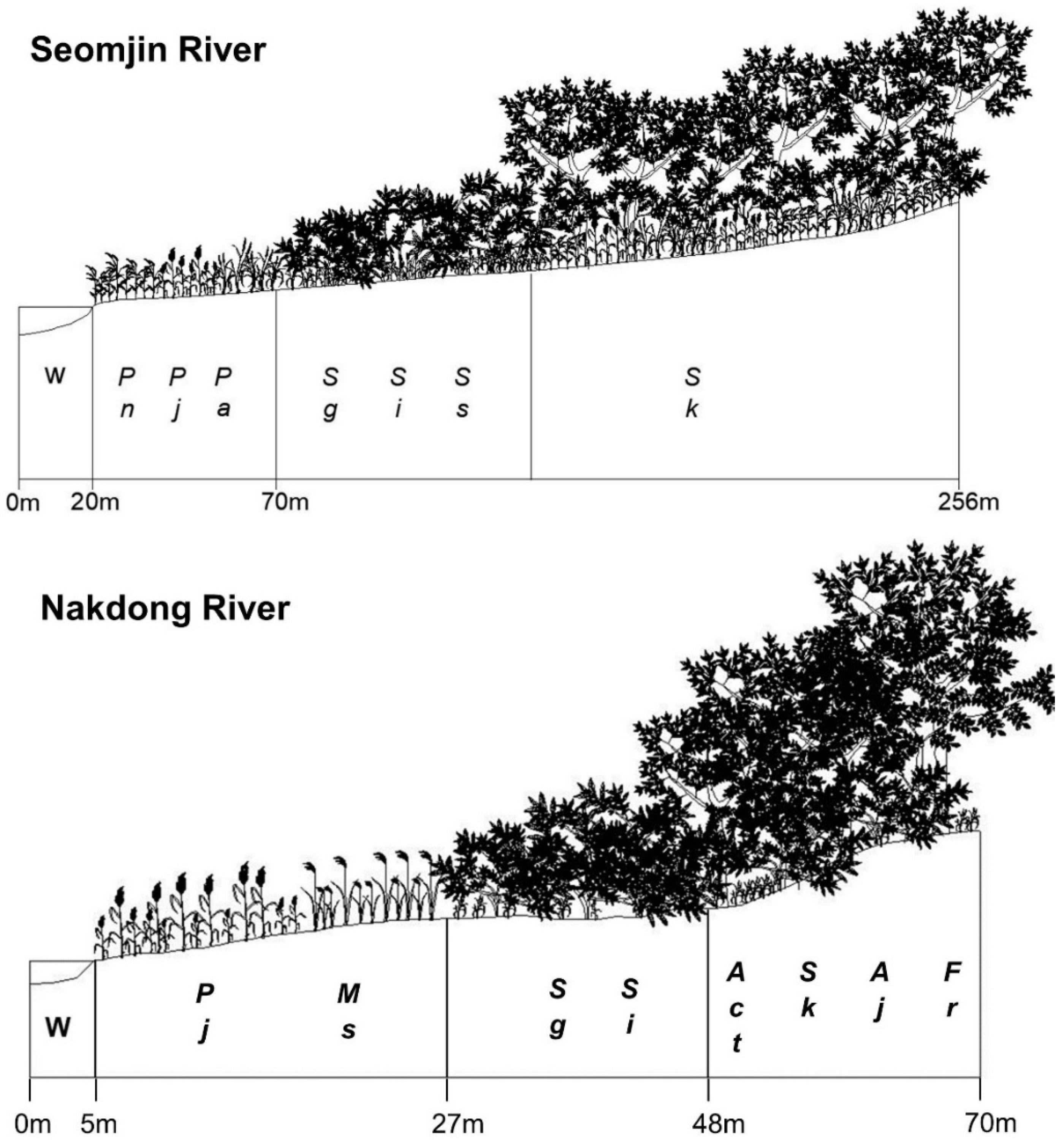
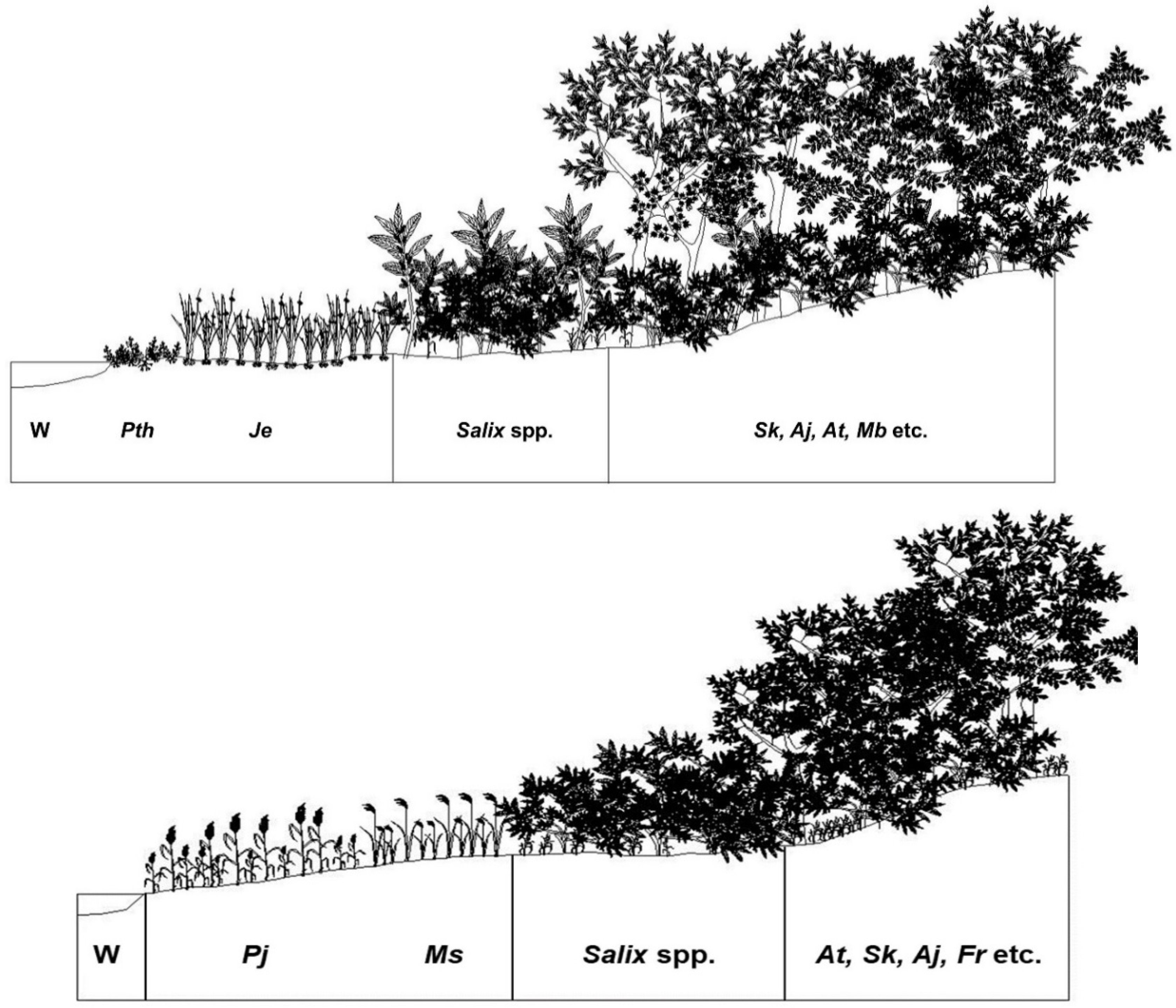
3.1. Successional Stages of the Abandoned Rice Fields Expressed as Stand Profile
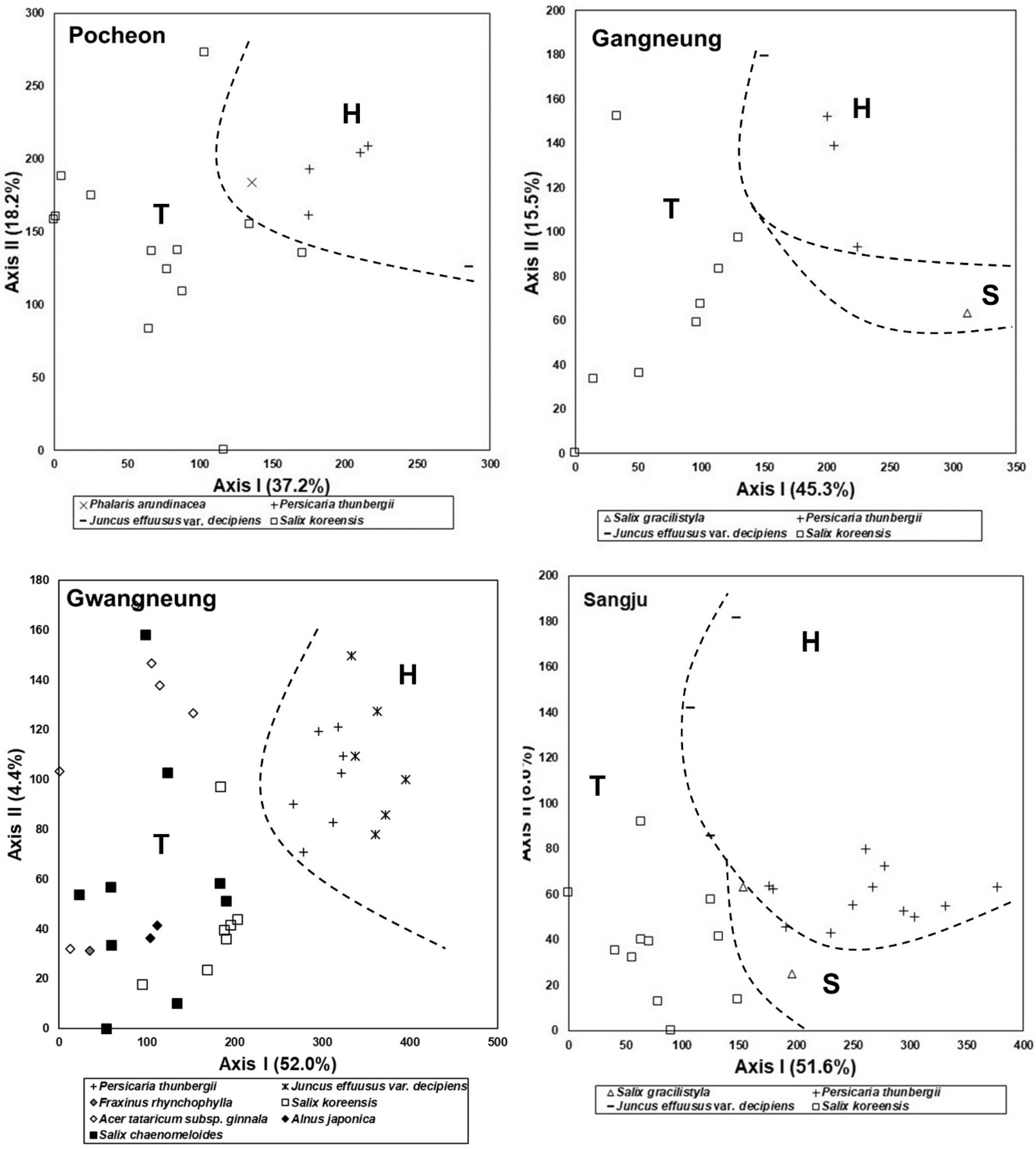
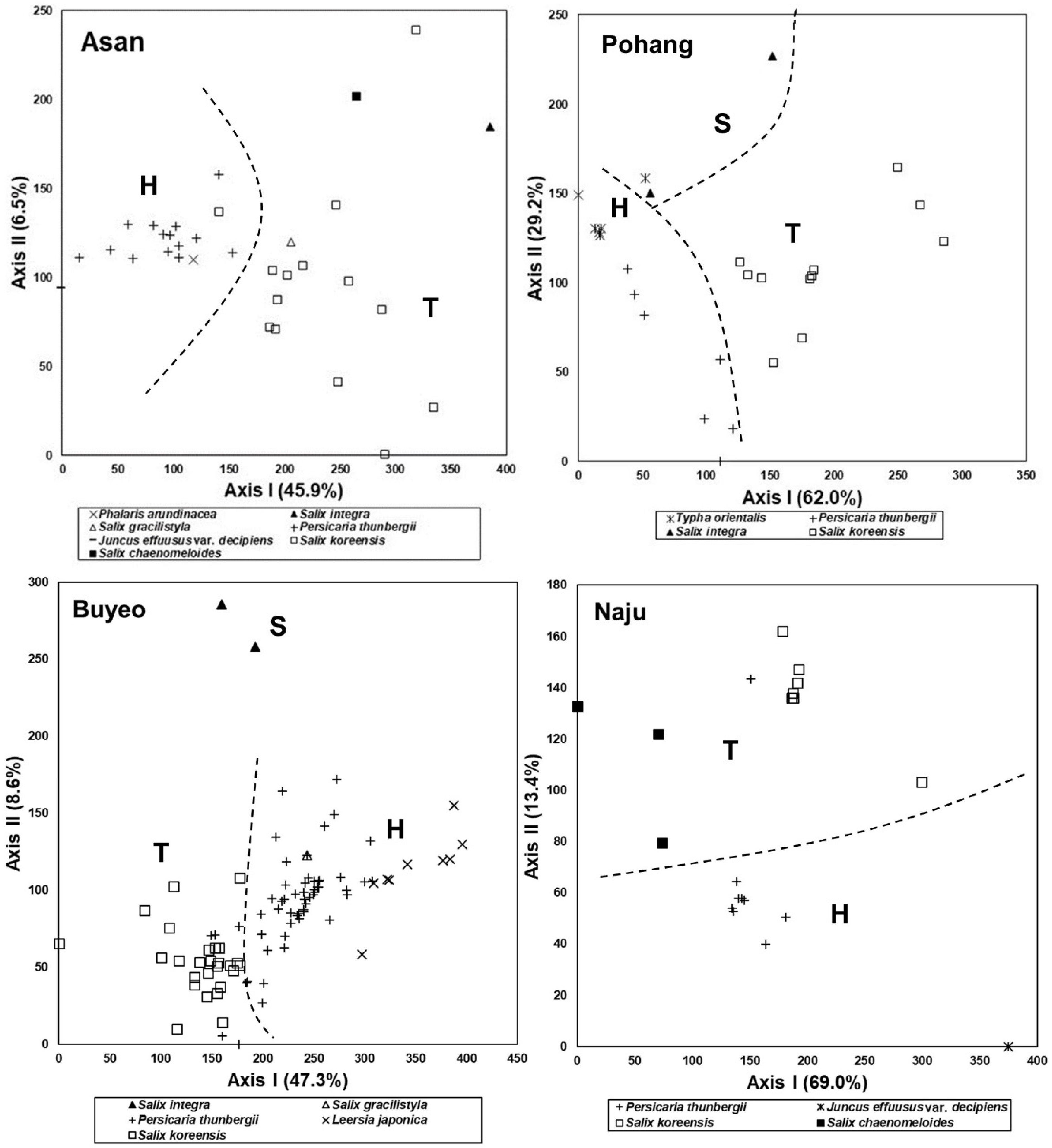
3.2. Changes in Species Composition Depending on Successional Stages
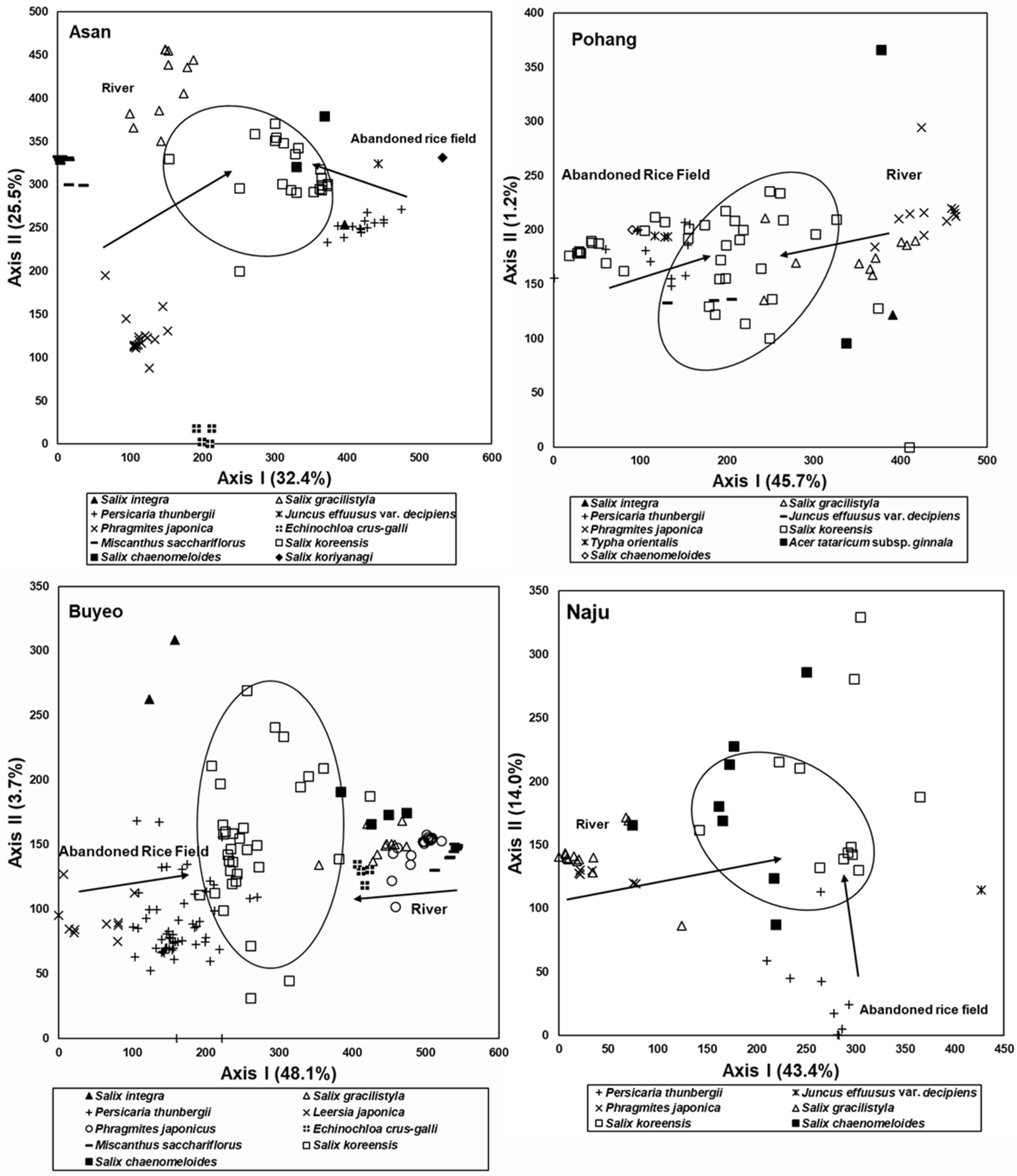
3.3. Spatial Distribution of the Riparian Vegetation Based on Stand Profile
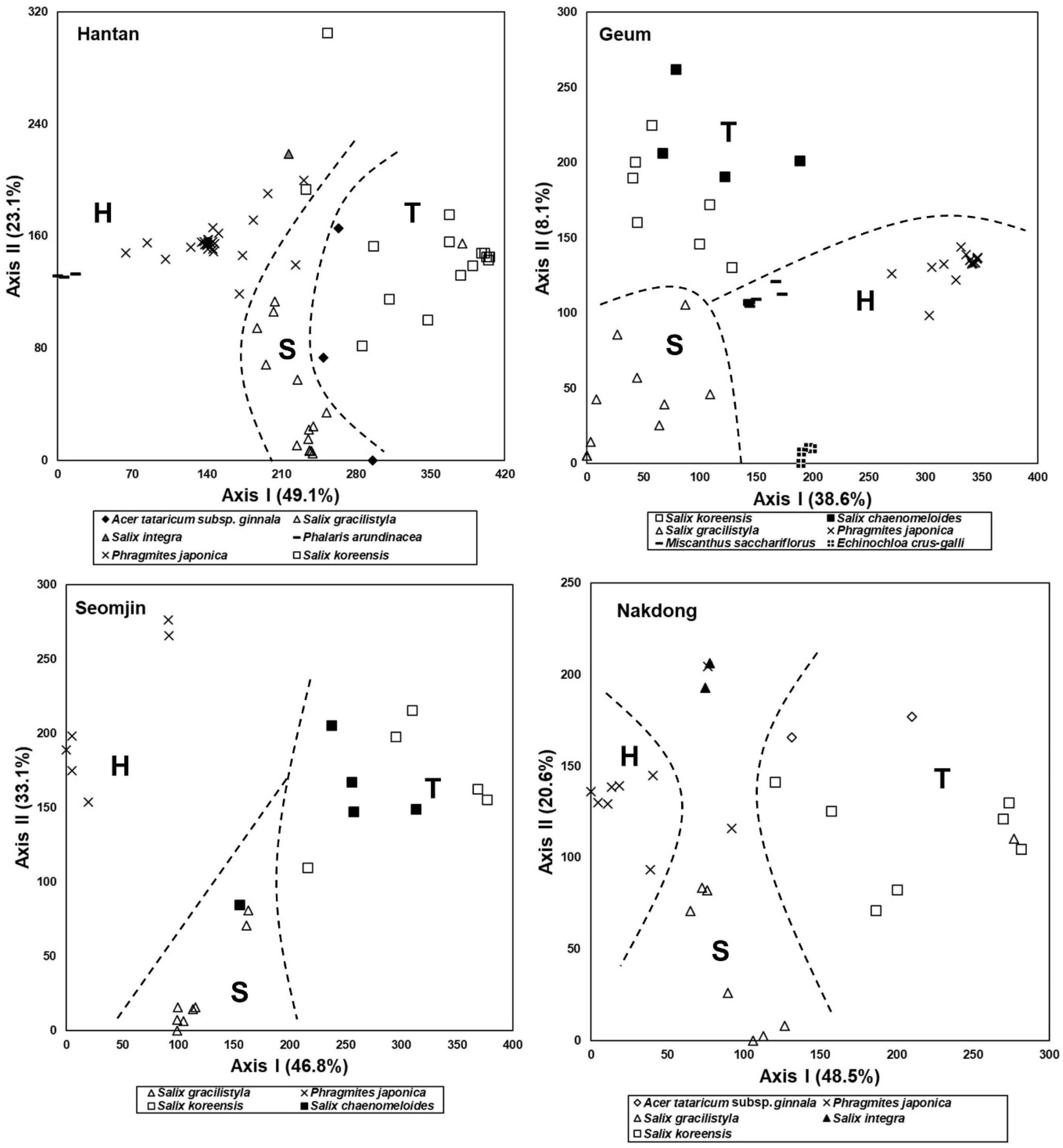
3.4. Species Composition of the Riparian Vegetation
3.5. Comparison of Species Composition between Vegetation in the Abandoned Rice Fields and the Riparian Vegetation
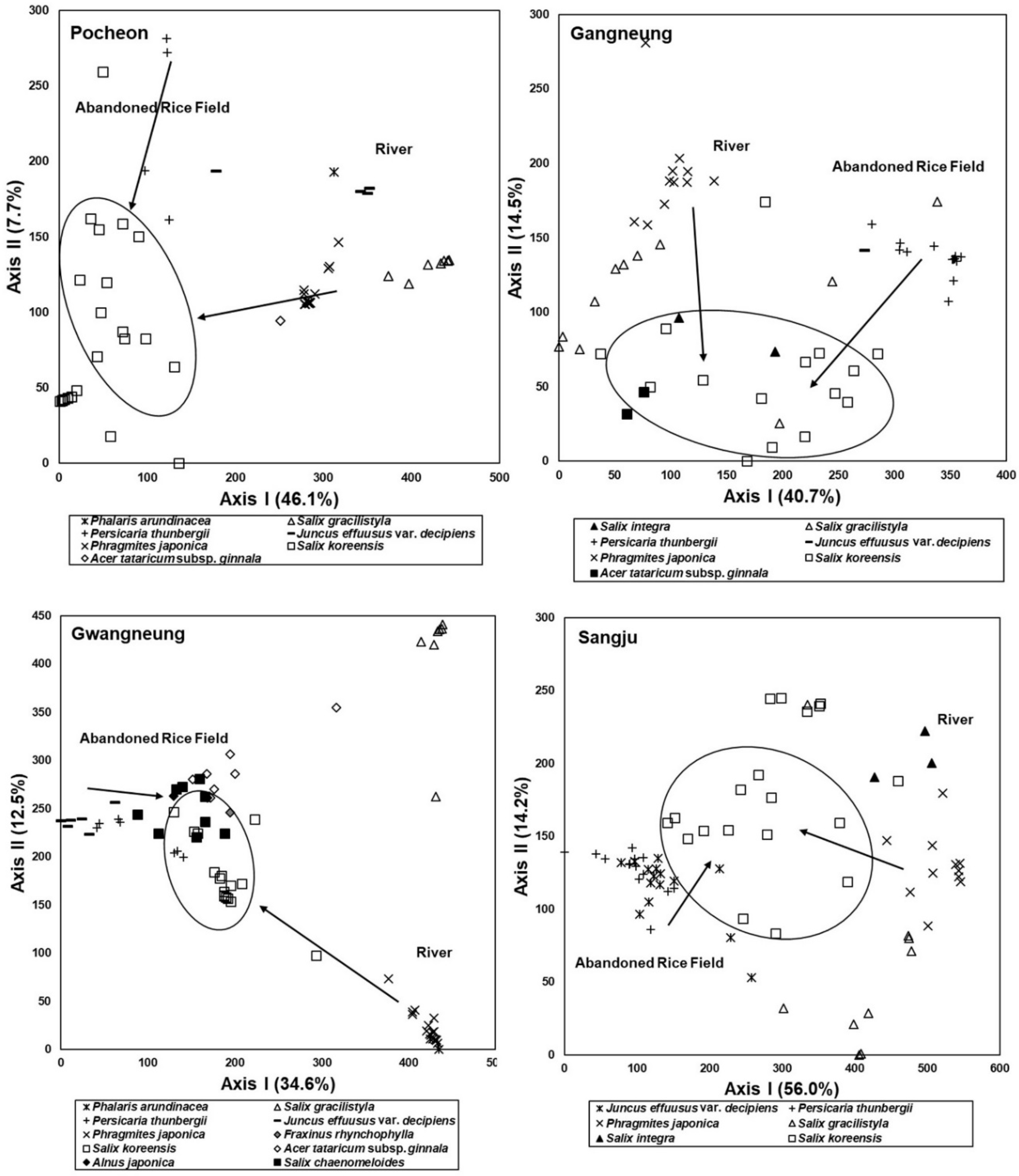
3.6. Feasibility of Vegetation Data Obtained from the Abandoned Rice Fields as Reference Information for Restoration of the Riparian Vegetation
4. Discussion
4.1. The Significance and Value of a Study on Succession
4.2. Relationship between Vegetation Sere in Abandoned Rice Fields and Spatial Distribution of Riparian Vegetation
4.3. Spatial Range of River as a Landscape
4.4. Land Use Pattern and Actual Status of Rivers in Korea
4.5. Necessity for Riparian Restoration
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Prach, K.; Walker, L.R. Four opportunities for studies of ecological succession. Trends Ecol. Evol. 2011, 26, 119–123. [Google Scholar] [CrossRef] [PubMed]
- Burton, P.J.; Jentsch, A.; Walker, L.R. The ecology of disturbance interactions. BioScience 2020, 70, 854–870. [Google Scholar] [CrossRef]
- Christensen, N.L.; Peet, R.K. Convergence during secondary forest succession. J. Ecol. 1984, 72, 25–36. [Google Scholar] [CrossRef]
- Huston, M.; Smith, T. Plant succession: Life history and competition. Am. Nat. 1987, 130, 168–198. [Google Scholar] [CrossRef]
- Pickett, S.T.A.; Collins, S.L.; Armesto, J.J. Models, mechanisms and pathways of succession. Bot. Rev. 1987, 53, 335–371. [Google Scholar] [CrossRef]
- Turner, M.G.; Baker, W.L.; Peterson, C.J.; Peet, R.K. Factors influencing succession: Lessons from large, infrequent natural disturbances. Ecosystems 1998, 1, 511–523. [Google Scholar] [CrossRef]
- Young, T.P.; Chase, J.M.; Huddleston, R.T. Community succession and assembly: Comparing, contrasting and combining paradigms in the context of ecological restoration. Ecol. Restor. 2001, 19, 5–18. [Google Scholar] [CrossRef]
- Chang, C.C.; Turner, B.L. Ecological succession in a changing world. J. Ecol. 2019, 107, 503–509. [Google Scholar] [CrossRef] [Green Version]
- Ellenberg, D.; Mueller-Dombois, D. Aims and Methods of Vegetation Ecology; Wiley: New York, USA, 1974. [Google Scholar]
- Rejmánek, M. Vegetation ecology matures in its homeland. Ecology 1990, 71, 2399–2400. [Google Scholar] [CrossRef]
- Glenn-Lewin, D.C.; Van Der Maarel, E. Patterns and processes of vegetation dynamics. In Plant Succession Theory Prediction; DC Glenn-Lewin, D.C., Peet, R.K., Veblen, T.T., Eds.; Chapman and Hall: London, UK, 1992; pp. 11–59. [Google Scholar]
- Rejmánek, M.; van Katwyk, K.P. Old Field Succession: A Bibliographic Review (1901–1991); University of California: Davis, CA, USA, 2005. [Google Scholar]
- Pickett, S.T.A. Population patterns through twenty years of oldfield succession. Vegetatio 1982, 49, 45–59. [Google Scholar] [CrossRef]
- Bazzaz, F.A.; Chiariello, N.R.; Coley, P.D.; Pitelka, L.F. Allocating resources to reproduction and defense. BioScience 1987, 37, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Goldberg, D.E. Neighborhood competition in an old-field plant community. Ecology 1987, 68, 1211–1223. [Google Scholar] [CrossRef]
- Inouye, R.S.; Tilman, D. Convergence and divergence of old-field plant communities along experimental nitrogen gradients. Ecology 1988, 69, 995–1004. [Google Scholar] [CrossRef]
- Schmidt, W. An experimental study of old-field succession in relation to different environmental factors. Vegetatio 1988, 77, 103–114. [Google Scholar] [CrossRef]
- Osbornová, J.; Kovárová, M.; Leps, J.; Prach, K. Succession in Abandoned Fields: Studies in Central Bohemia, Czechoslovakia; Springer Science & Business Media: Berlin, Germany, 1990; Volume 15. [Google Scholar]
- Holt, R.D.; Robinson, G.R.; Gaines, M.S. Vegetation dynamics in an experimentally fragmented landscape. Ecology 1995, 76, 1610–1624. [Google Scholar] [CrossRef]
- Cramer, V.A.; Hobbs, R.J.; Standish, R.J. What’s new about old fields? Land abandonment and ecosystem assembly. Trends Ecol. Evol. 2008, 23, 104–112. [Google Scholar] [CrossRef]
- Martínez-Duro, E.; Ferrandis, P.; Escudero, A.; Luzuriaga, A.L.; Herranz, J.M. Secondary old-field succession in an ecosystem with restrictive soils: Does time from abandonment matter? Appl. Veg. Sci. 2010, 13, 234–248. [Google Scholar] [CrossRef]
- Meiners, S.J.; Cadotte, M.W.; Fridley, J.D.; Pickett, S.T.A.; Walker, L.R. Is successional research nearing its climax? New approaches for understanding dynamic communities. Funct. Ecol. 2015, 29, 154–164. [Google Scholar] [CrossRef]
- Shimoda, M.; Suzuki, H. Vegetation of fallow rice fields in the Saijo Basin, Hiroshima Prefecture, Japan, Japan. Hikobia Suppl. 1981, 1, 321–339. [Google Scholar]
- Shimoda, M. Abandoned rice field vegetation and its evaluation, A case study of wet abandoned rice field vegetation in Hiroshima Prefecture. Veg. Sci. 1996, 13, 37–50. [Google Scholar]
- Kim, J.W.; Nam, H.K. Syntaxonomical and synecological characteristics of rice field vegetation. Korean J. Ecol. 1998, 21, 203–215. [Google Scholar]
- Cao, J.; Li, M.; Deo, R.C.; Adamowski, J.F.; Cerdà, A.; Feng, Q.; Liu, M.; Zhang, J.; Zhu, G.; Zhang, X.; et al. Comparison of social-ecological resilience between two grassland management patterns driven by grassland land contract policy in the Maqu, Qinghai-Tibetan Plateau. Land Use Policy 2018, 74, 88–96. [Google Scholar] [CrossRef] [Green Version]
- Hong, M.G.; Park, H.; Nam, B.E.; Kim, J.G. Vegetational characteristics of abandoned paddy terraces in comparison with natural and constructed wetlands. J. Wetl. Res. 2019, 21, 199–206. [Google Scholar]
- Haripal, K.; Sahoo, S. Vegetational dynamics in some tropical abandoned rice fields in the western part of Orissa, India. Afr. J. Environ. Sci. Technol. 2011, 5, 37–44. [Google Scholar]
- Lee, C.; You, Y.; Robinson, G.R. Secondary succession and natural habitat restoration in abandoned rice fields of central Korea. Restor. Ecol. 2002, 10, 306–314. [Google Scholar] [CrossRef]
- Yoon, J.; Nam, J.M.; Kim, H.; Bae, Y.J.; Kim, J.G. Nannophya pygmaea (odonata: Libellulidae), an endangered dragonfly in Korea, prefers abandoned paddy fields in the early seral stage. Environ. Entomol. 2010, 39, 278–285. [Google Scholar] [CrossRef]
- Shimoda, S.; Wagai, R. Ecosystem dynamics after abandonment of rice paddy fields: Does alien plant invasion enhance carbon storage? Ecosystems 2020, 23, 617–629. [Google Scholar] [CrossRef]
- Lee, C.S.; Cho, Y.C.; Shin, H.C.; Moon, J.S.; Lee, B.C.; Bae, Y.S.; Byun, H.G.; Yi, H.B. Ecological response of streams in Korea under different management regimes. Water Eng. Res. 2005, 6, 131–147. [Google Scholar]
- Lee, C.S.; Lee, H.; Kim, A.R.; Pi, J.H.; Bae, Y.J.; Choi, J.K.; Lee, W.S.; Moon, J.S. Ecological effects of daylighting and plant reintroduction to the Cheonggye stream in Seoul, Korea. Ecol. Eng. 2020, 152, 105879. [Google Scholar] [CrossRef]
- Lim, C.H.; Pi, J.H.; Kim, A.R.; Cho, H.J.; Lee, K.S.; You, Y.H.; Lee, K.H.; Kim, K.D.; Moon, J.S.; Lee, C.S. Diagnostic evaluation and preparation of the reference information for river restoration in South Korea. Int. J. Environ. Res. Public Health 2021, 18, 1724. [Google Scholar] [CrossRef]
- Lee, C.S.; Hong, S.K.; You, Y.H.; Kim, J.E.; Moon, J.S. Landscape structure in the green belt around the metropolitan area of Seoul, Korea. In Proceedings of the IAVS Symposium, Uppsala, Sweden, 23–28 July 2000; pp. 230–233. [Google Scholar]
- Lee, C.S.; Moon, J.S.; Kim, J.E. Landscape changes caused by forest fire and vegetation regeneration processes. In Proceedings of the IAVS Symposium, Uppsala, Sweden, 23–28 July 2000; pp. 99–103. [Google Scholar]
- Ok, Y.H. Changes of Soil Properties, Species Diversity and r-K Selection in Early Stage of Old-Field Succession; Seoul National University: Seoul, Korea, 1984. [Google Scholar]
- Ku, Y.B. Test of Connell and Slatyer’s Model by the Litter Treatment in Early Abandoned Field Succession; Seoul National University: Seoul, Korea, 1994. [Google Scholar]
- Lee, K.S.; Kim, J.H. Seral changes in environmental factors and recovery of soil fertility during abandoned field succession after shifting cultivation. Korean J. Ecol. 1995, 18, 243–253. [Google Scholar]
- Lee, K.S.; Kim, J.H. Seral changes in floristic composition during abandoned field succession after shifting cultivation. Korean J. Ecol. 1995, 18, 275–283. [Google Scholar]
- Lee, C.S.; Yon, Y.H. Ecological consideration for restoration of the degraded urban river. Korean J. Ecol. 2002, 25, 247–252. [Google Scholar] [CrossRef]
- Ferreira, L.V.; Stohlgren, T.J. Effects of river level fluctuation on plant species richness, diversity, and distribution in a floodplain forest in Central Amazonia. Oecologia 1999, 120, 582–587. [Google Scholar] [CrossRef]
- Casanova, M.T.; Brock, M.A. How do depth, duration and frequency of flooding influence the establishment of wetland plant communities? Plant Ecol. 2000, 147, 237–250. [Google Scholar] [CrossRef]
- Vandersande, M.W.; Glenn, E.P.; Walworth, J.L. Tolerance of five riparian plants from the lower Colorado River to salinity drought and inundation. J. Arid. Environ. 2001, 49, 147–159. [Google Scholar] [CrossRef]
- Parolin, P. Submergence tolerance vs. escape from submergence: Two strategies of seedling establishment in Amazonian floodplains. Environ. Exp. Bot. 2002, 48, 177–186. [Google Scholar] [CrossRef]
- Xiaoling, L.; Ning, L.; Jin, Y.; Fuzhou, Y.; Faju, C.; Fangqing, C. Morphological and photosynthetic responses of riparian plant Distylium chinense seedlings to simulated Autumn and Winter flooding in Three Gorges Reservoir Region of the Yangtze River, China. Acta Ecol. Sin. 2011, 31, 31–39. [Google Scholar] [CrossRef]
- Blom, C.W.P.M.; Voesenek, L.A.C.J. Flooding: The survival strategies of plants. Trends Ecol. Evol. 1996, 11, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Shafroth, P.B.; Stromberg, J.C.; Patten, D.T. Riparian vegetation response to altered disturbance and stress regimes. Ecol. Appl. 2002, 12, 107–123. [Google Scholar] [CrossRef]
- Auble, G.T.; Scott, M.L.; Friedman, J.M. Use of individualistic streamflow-vegetation relations along the Fremont River, Utah, USA to assess impacts of flow alteration on wetland and riparian areas. Wetlands 2005, 25, 143–154. [Google Scholar] [CrossRef]
- Capon, S.J. Flood variability and spatial variation in plant community composition and structure on a large arid floodplain. J. Arid. Environ. 2005, 60, 283–302. [Google Scholar] [CrossRef]
- Bornette, G.; Tabacchi, E.; Hupp, C.; Puijalon, S.; Rostan, J.-C. A model of plant strategies in fluvial hydrosystems. Freshw. Biol. 2008, 53, 1692–1705. [Google Scholar] [CrossRef] [Green Version]
- Hauer, F.R.; Dahm, C.N.; Lamberti, G.A.; Stanford, J.A. Landscapes and ecological variability of rivers in North America: Factors affecting restoration strategies. In Strategies for Restoring River Ecosystems: Sources of Variability and Uncertainty in Natural and Managed Systems; Wissmar, R.C., Bisson, P.A., Eds.; American Fisheries Society: Bethesda, MD, USA, 2003; pp. 81–105. [Google Scholar]
- Stanford, J.A.; Lorang, M.S.; Hauer, F.R. The shifting habitat mosaic of river ecosystems. SIL Proc. 2005, 29, 123–136. [Google Scholar] [CrossRef]
- Lorang, M.S.; Whited, D.C.; Hauer, F.R.; Kimball, J.S.; Stanford, J.A. Using airborne multispectral imagery to evaluate geomorphic work across floodplains of gravel-bed rivers. Ecol. Appl. 2005, 15, 1209–1222. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.B. Illustrated Flora of Korea; HyangMoonSa: Seoul, Korea, 1985. [Google Scholar]
- Park, S.-H. Colored Illustrations of Naturalized Plants of Korea; Ilchokak: Seoul, Korea, 1995; p. 371. [Google Scholar]
- Korea National Arboretum. Korean Plant Names Index. Available online: http://www.nature.go.kr/kbi/plant/pilbk/selectPlantPilbkGnrlList.do (accessed on 15 July 2022).
- Braun-Blanquet, J. Pflanzensoziologie: Grundzüge der Vegetationskunde; Springer: Berlin, Germany, 1964. [Google Scholar]
- Curtis, J.T.; McIntosh, R.P. An upland forest continuum in the prairie-forest border region of wisconsin. Ecology 1951, 32, 476–496. [Google Scholar] [CrossRef]
- Hill, M.O.; Gauch, H.G. Detrended Correspondence Analysis: An Improved Ordination Technique. In Classification and Ordination: Symposium on Advances in Vegetation Science, Nijmegen, The Netherlands, May 1979; van der Maarel, E., Ed.; Springer: Dordrecht, The Netherland, 1980; pp. 47–58. [Google Scholar]
- McCune, B.; Mefford, M.J. PC-ORD, Multivariate Analysis of Ecological Data; Version 4; MjM Software Design: Glenden Beach, OR, USA, 1999. [Google Scholar]
- Cowles, H.C. The ecological relations of the vegetation on the sand dunes of lake Michigan. Part I.-geographical relations of the dune floras. Bot. Gaz. 1899, 27, 95–117. [Google Scholar] [CrossRef] [Green Version]
- Margalef, R. Perspectives in Ecological Theory; University of Chicago Press: Chicago, IL, USA, 1968. [Google Scholar]
- Walker, L.R. Linking Restoration and Ecological Succession; Walker, L.R., Joe, W., Hobbs, R.J., Eds.; Springer: London, UK, 2007. [Google Scholar]
- Christensen, N.L. An historical perspective on forest succession and its relevance to ecosystem restoration and conservation practice in North America. For. Ecol. Manag. 2014, 330, 312–322. [Google Scholar] [CrossRef]
- Egerton, F.N. History of ecological sciences, part 54: Succession, community, and continuum. Bull. Ecol. Soc. Am. 2015, 96, 426–474. [Google Scholar] [CrossRef] [Green Version]
- Walker, L.R.; Wardle, D.A. Plant succession as an integrator of contrasting ecological time scales. Trends Ecol. Evol. 2014, 29, 504–510. [Google Scholar] [CrossRef]
- Chang, C.; HilleRisLambers, J. Integrating succession and community assembly perspectives. F1000Research 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Hernández, J.; Gavilán, R.G. Impacts of land-use changes on vegetation and ecosystem functioning: Old-field secondary succession. Plants 2021, 10, 990. [Google Scholar] [CrossRef]
- Pulsford, S.A.; Lindenmayer, D.B.; Driscoll, D.A. A succession of theories: Purging redundancy from disturbance theory. Biol. Rev. 2016, 91, 148–167. [Google Scholar] [CrossRef]
- Kim, A.R.; Lim, B.S.; Seol, J.; Lee, C.S. Principle of restoration ecology reflected in the process creating the national institute of ecology. J. Ecol. Environ. 2021, 45, 12. [Google Scholar] [CrossRef]
- Méndez-Toribio, M.; Martínez-Garza, C.; Ceccon, E. Challenges during the execution, results, and monitoring phases of ecological restoration: Learning from a country-wide assessment. PLoS ONE 2021, 16, e0249573. [Google Scholar] [CrossRef]
- Grime, J.P. Plant Strategies, Vegetation Processes; Wiley: New York, NY, USA, 1979. [Google Scholar]
- Connell, J.H.; Slatyer, R.O. Mechanisms of succession in natural communities and their role in community stability and organization. Am. Nat. 1977, 111, 1119–1144. [Google Scholar] [CrossRef]
- Wyse, S.V.; Wilmshurst, J.M.; Burns, B.R.; Perry, G.L.W. New Zealand forest dynamics a review of past and present vegetation responses to disturbance, and development of conceptual forest models. N. Z. J. Ecol. 2018, 42, 87–106. [Google Scholar] [CrossRef] [Green Version]
- White, P.S. Pattern, process, and natural disturbance in vegetation. Bot. Rev. 1979, 45, 229–299. [Google Scholar] [CrossRef]
- White, P.S. Natural disturbance and patch dynamics: An introduction. In The Ecology Natural Disturbance and Patch Dynamics; Pickett, S.T.A., White, P.S., Eds.; Academic Press: Orland, FL, USA, 1985; pp. 3–13. [Google Scholar]
- White, P.S.; Harrod, J.; Walker, J.L.; Jentsch, A. Disturbance, scale, and boundary in wilderness management. In Proceedings of the Wilderness Science in a Time of Change Conference, Missoula, MT, USA, 23–27 May 1999; pp. 27–42. [Google Scholar]
- Lugo, A.E. Effects of extreme disturbance events: From ecesis to social–Ecological–Technological systems. Ecosystems 2020, 23, 1726–1747. [Google Scholar] [CrossRef] [Green Version]
- Kramer, K.; Brinkman, B.; Kuiters, L.; Verdonschot, P.F.M. Is Ecological Succession Predictable?: Theory and Applications; Alterra: Wageningen, The Netherland, 2005. [Google Scholar]
- Naiman, R.J.; Décamps, H. The ecology of interfaces: Riparian zones. Annu. Rev. Ecol. Syst. 1997, 28, 621–658. [Google Scholar] [CrossRef] [Green Version]
- Van Looy, K.; Tormos, T.; Ferréol, M.; Villeneuve, B.; Valette, L.; Chandesris, A.; Bougon, N.; Oraison, F.; Souchon, Y. Benefits of riparian forest for the aquatic ecosystem assessed at a large geographic scale. Knowl. Managt. Aquat. Ecosyst. 2013, 408, 06. [Google Scholar] [CrossRef] [Green Version]
- LeRoy, N.; Allan, J.D.; Bain, M.B.; Karr, J.R.; Prestegaard, K.L.; Richter, B.D.; Sparks, R.E.; Stromberg, J.C. The natural flow regime. BioScience 1997, 47, 769–784. [Google Scholar]
- Bejarano, M.D.; González del Tánago, M.; de Jalón, D.G.; Marchamalo, M.; Sordo-Ward, Á.; Solana-Gutiérrez, J. Responses of riparian guilds to flow alterations in a Mediterranean stream. J. Veg. Sci. 2012, 23, 443–458. [Google Scholar] [CrossRef] [Green Version]
- Whited, D.C.; Lorang, M.S.; Harner, M.J.; Hauer, F.R.; Kimball, J.S.; Stanford, J.A. Climate, hydrologic disturbance, and succession: Drivers of floodplain pattern. Ecology 2007, 88, 940–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corenblit, D.; Tabacchi, E.; Steiger, J.; Gurnell, A.M. Reciprocal interactions and adjustments between fluvial landforms and vegetation dynamics in river corridors: A review of complementary approaches. Earth Sci. Rev. 2007, 84, 56–86. [Google Scholar] [CrossRef]
- Bendix, J.; Hupp, C.R. Hydrological and geomorphological impacts on riparian plant communities. Hydrol. Processes 2000, 14, 2977–2990. [Google Scholar] [CrossRef]
- Rivaes, R.; Pinheiro, A.N.; Egger, G.; Ferreira, T. The role of river Morphodynamic disturbance and groundwater hydrology as driving factors of riparian landscape patterns in Mediterranean rivers. Front. Plant Sci. 2017, 8, 1612. [Google Scholar] [CrossRef] [Green Version]
- Park, S.A.; Kim, G.S.; Pee, J.H.; Oh, W.S.; Kim, H.S.; Lee, C.S. Reference information for realizing ecological restoration of river: A case study in the Bongseonsa stream. J. Ecol. Environ. 2013, 36, 235–243. [Google Scholar] [CrossRef] [Green Version]
- Goodwin, C.N.; Hawkins, C.P.; Kershner, J.L. Riparian restoration in the western United States: Oveview and perspecive. Restor. Ecol. 1997, 5, 4–14. [Google Scholar] [CrossRef]
- van Rensburg, D. River Index of Habitat Integrity (IHI) and Riparian Vegetation Assessment for the Proposed Establishment of a Measuring Weir within the Caledon (Mohokare) River (Quaternary Drainage Region: D22D). Available online: https://sahris.sahra.org.za/sites/default/files/additionaldocs/Caledon%20IHI.pdf (accessed on 10 July 2022).
- An, J.H.; Lim, C.H.; Lim, Y.K.; Nam, K.B.; Lee, C.S. A review of restoration project evaluation and post management for ecological restoration of the river. J. Restor. Ecol. 2014, 4, 15–34. [Google Scholar]
- Ahn, S.M. The emergence of rice agriculture in Korea: Archaeobotanical perspectives. Archaeol. Anthropol. Sci. 2010, 2, 89–98. [Google Scholar] [CrossRef]
- Lee, C.S.; Cho, Y.C.; Shin, H.C.; Lee, S.M. Changes of landscape structure for the recent 20 years in the Wangsuk stream basin of the central Korea. J. Ecol. Environ. 2006, 29, 405–410. [Google Scholar] [CrossRef] [Green Version]
- Lim, C.H.; Jung, S.H.; Kim, A.R.; Kim, N.S.; Lee, C.S. Monitoring for changes in spring phenology at both temporal and spatial scales based on MODIS LST data in South Korea. Remote Sens. 2020, 12, 3282. [Google Scholar] [CrossRef]
- Jung, S.H.; Kim, A.r.; Seol, J.; Lim, B.S.; Lee, C.S. Characteristics and reference information of riparian vegetation for realizing ecological restoration classified by reach of the river in Korea. Korean Soc. Water Environ. 2018, 34, 447–461. [Google Scholar] [CrossRef]
- Kim, A.R.; Kim, D.U.; Lim, B.S.; Seol, J.; Lee, C.S. An evaluation on restoration effect in the restored Yangjae stream and the improvement plan based on the result. Korean J. Ecol. Environ. 2020, 53, 390–407. [Google Scholar] [CrossRef]
- Pi, J.H. The Evaluation of Impact of Dam Construction and Different Levels of Restoration on the Naturalness of Riparian Vegetation and Adaptive Management Plans for Improving the Naturalness of the Influenced Rivers of Restoration; Seoul Women’s University: Seoul, Korea, 2014. [Google Scholar]
- Pi, J.H. Construction of Vegetation Information of Reference River to Restore the Degraded River in Korean; Seoul Women’s University: Seoul, Korea, 2010. [Google Scholar]
- Tabari, H. Climate change impact on flood and extreme precipitation increases with water availability. Sci. Rep. 2020, 10, 13768. [Google Scholar] [CrossRef]
- Yamamoto, K.; Sayama, T.; Apip. Impact of climate change on flood inundation in a tropical river basin in Indonesia. Prog. Earth Planet. Sci. 2021, 8, 5. [Google Scholar] [CrossRef]
- Lee, C.S.; Jeong, Y.M.; Kang, H.S. Concept, direction, and task of ecological restoration. J. Restor. Ecol. 2011, 2, 59–71. [Google Scholar]
- Wohl, E.; Angermeier, P.L.; Bledsoe, B.; Kondolf, G.M.; MacDonnell, L.; Merritt, D.M.; Palmer, M.A.; Poff, N.L.; Tarboton, D. River restoration. Water Resour. Res. 2005, 41, 1–12. [Google Scholar] [CrossRef]
- Hesselink, A.W. History Makes a River. Man Induced Morphological Changes of the River Rhine and Its Implications for Future River Management in The Netherlands. In Proceedings of the 2nd International Workshop on River Environment, Ilsan, Korea, 26 June 2006; pp. 13–40. [Google Scholar]
- Fokkens, B. The Dutch Strategy for Safety and River Flood Prevention. In Extreme Hydrological Events: New Concepts for Security; Springer: Dordrecht, The Netherland; 2007, pp. 337–352.
- Seavy, N.E.; Gardali, T.; Golet, G.H.; Griggs, F.T.; Howell, C.A.; Kelsey, R.; Small, S.L.; Viers, J.H.; Weigand, J.F. Why climate change makes riparian restoration more important than ever: Recommendations for practice and research. Ecol. Restor. 2009, 27, 330–338. [Google Scholar] [CrossRef]
- Wohl, E.; Lane, S.N.; Wilcox, A.C. The science and practice of river restoration. Water Resour. Res. 2015, 51, 5974–5997. [Google Scholar] [CrossRef] [Green Version]
- Rijke, J.; van Herk, S.; Zevenbergen, C.; Ashley, R. Room for the river: Delivering integrated river basin management in the Netherlands. Int. J. River Basin Manag. 2012, 10, 369–382. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| River Name | Land Use Type (%) | ||||
|---|---|---|---|---|---|
| Urbanized Area | Agricultural Field | Forest | Road | Bare Ground | |
| Nakdong | 0.9 | 14.3 | 80.4 | 4.4 | 0.0 |
| Hantan | 1.7 | 47.8 | 49.9 | 0.2 | 0.4 |
| Geum | 8.9 | 26.7 | 64.3 | 0.1 | 0.0 |
| Seomjin | 8.7 | 27.3 | 63.9 | 0.1 | 0.0 |
| River Name | Site No. | Site Name | Latitude | Longitude |
|---|---|---|---|---|
| Nakdong | 1 | Songjeong | 37°03′54.10″ | 129°02′18.90″ |
| 2 | Gyeoldun bridge | 37°00′56.00″ | 129°04′36.70″ | |
| 3 | Seungbu station | 36°59′31.20″ | 129°05′02.10″ | |
| 4 | Wongok bridge | 36°57′42.32″ | 129°05′27.71″ | |
| 5 | Docheon | 36°51′51.05″ | 128°54′13.76″ | |
| 6 | Mt. Cheongnyang | 36°46′45.62″ | 128°53′12.61″ | |
| 7 | Andong Dam | 36°44′05.01″ | 128°52′39.92″ | |
| Hantan | 1 | Yangji | 38°15′02.3″ | 127°17′05.3″ |
| Geum | 1 | Simcheon | 36°12′17.4″ | 127°42′01.1″ |
| Seomjin | 1 | Gurye | 35°11′27.5″ | 127°32′47.3″ |
| Survey Areas for Abandoned Rice Paddies | Dominant Plant Community by Successional Stage | ||
|---|---|---|---|
| Early Stage | Mid-Stage | Late Stage | |
| Pocheon | Persicaria thunbergii community, Juncus effuusus var. decipiens community, and Phalaris arundinacea community | Young Salix koreensis community | Mature S. koreensis community |
| Gwangneung | P. thunbergii community and J. effuusus var. decipiens community | Not appeared | S. koreensis community, S. chaenomeloides community, Alnus japonica community, and Acer tataricum subsp. ginnala community |
| Asan | P. thunbergii community and P. arundinacea community | Salix gracilistyla community, Salix integra community, and young S. koreensis community | S. koreensis community and S. chaenomeloides community |
| Buyeo | Persicaria longiseta community, Digitaria ciliaris community, Echinochloa utilis community, Bidens frondosa community, P. thunbergii community, and Leersia japonica community | S. integra community, and young S. koreensis community | S. koreensis community |
| Naju | P. thunbergii community | S. gracilistyla community | S. koreensis community |
| Gangneung | P. thunbergii community and J. effuusus var. decipiens community | S. gracilistyla community and young S. koreensis community | S. koreensis community |
| Sangju | P. thunbergii community and J. effuusus var. decipiens community | S. gracilistyla community and young S. koreensis community | S. koreensis community |
| Pohang | P. thunbergii community and Typha orientalis community | S. integra community and young S. koreensis community | S. koreensis community |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, B.S.; Seol, J.; Kim, A.R.; An, J.H.; Lim, C.H.; Lee, C.S. Succession of the Abandoned Rice Fields Restores the Riparian Forest. Int. J. Environ. Res. Public Health 2022, 19, 10416. https://doi.org/10.3390/ijerph191610416
Lim BS, Seol J, Kim AR, An JH, Lim CH, Lee CS. Succession of the Abandoned Rice Fields Restores the Riparian Forest. International Journal of Environmental Research and Public Health. 2022; 19(16):10416. https://doi.org/10.3390/ijerph191610416
Chicago/Turabian StyleLim, Bong Soon, Jaewon Seol, A Reum Kim, Ji Hong An, Chi Hong Lim, and Chang Seok Lee. 2022. "Succession of the Abandoned Rice Fields Restores the Riparian Forest" International Journal of Environmental Research and Public Health 19, no. 16: 10416. https://doi.org/10.3390/ijerph191610416
APA StyleLim, B. S., Seol, J., Kim, A. R., An, J. H., Lim, C. H., & Lee, C. S. (2022). Succession of the Abandoned Rice Fields Restores the Riparian Forest. International Journal of Environmental Research and Public Health, 19(16), 10416. https://doi.org/10.3390/ijerph191610416