
The Impact of Training on the Loss of Cardiorespiratory Fitness in Aging Masters Endurance Athletes



Abstract
:1. Introduction
2. Materials and Methods
3. Results

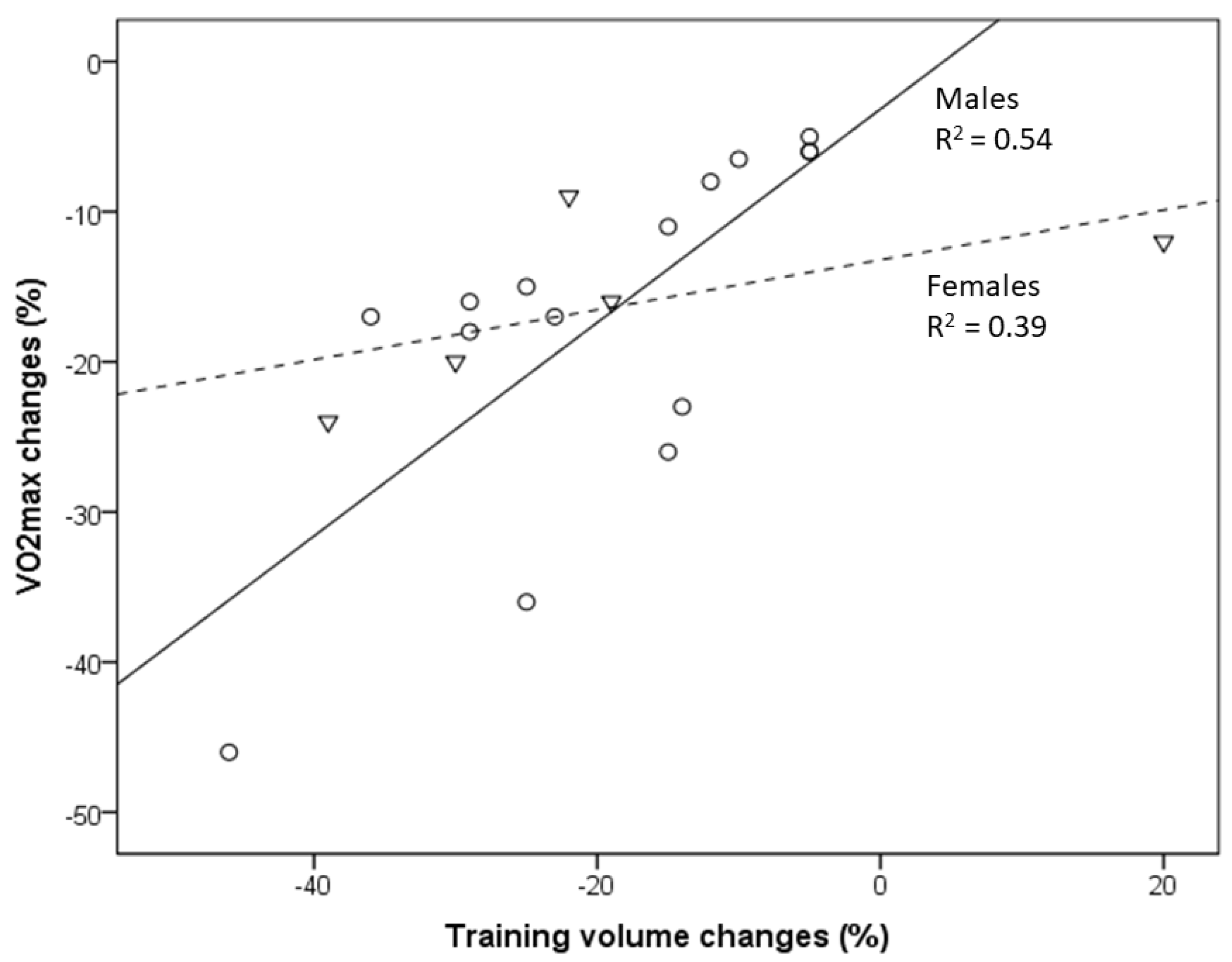{kind=link}
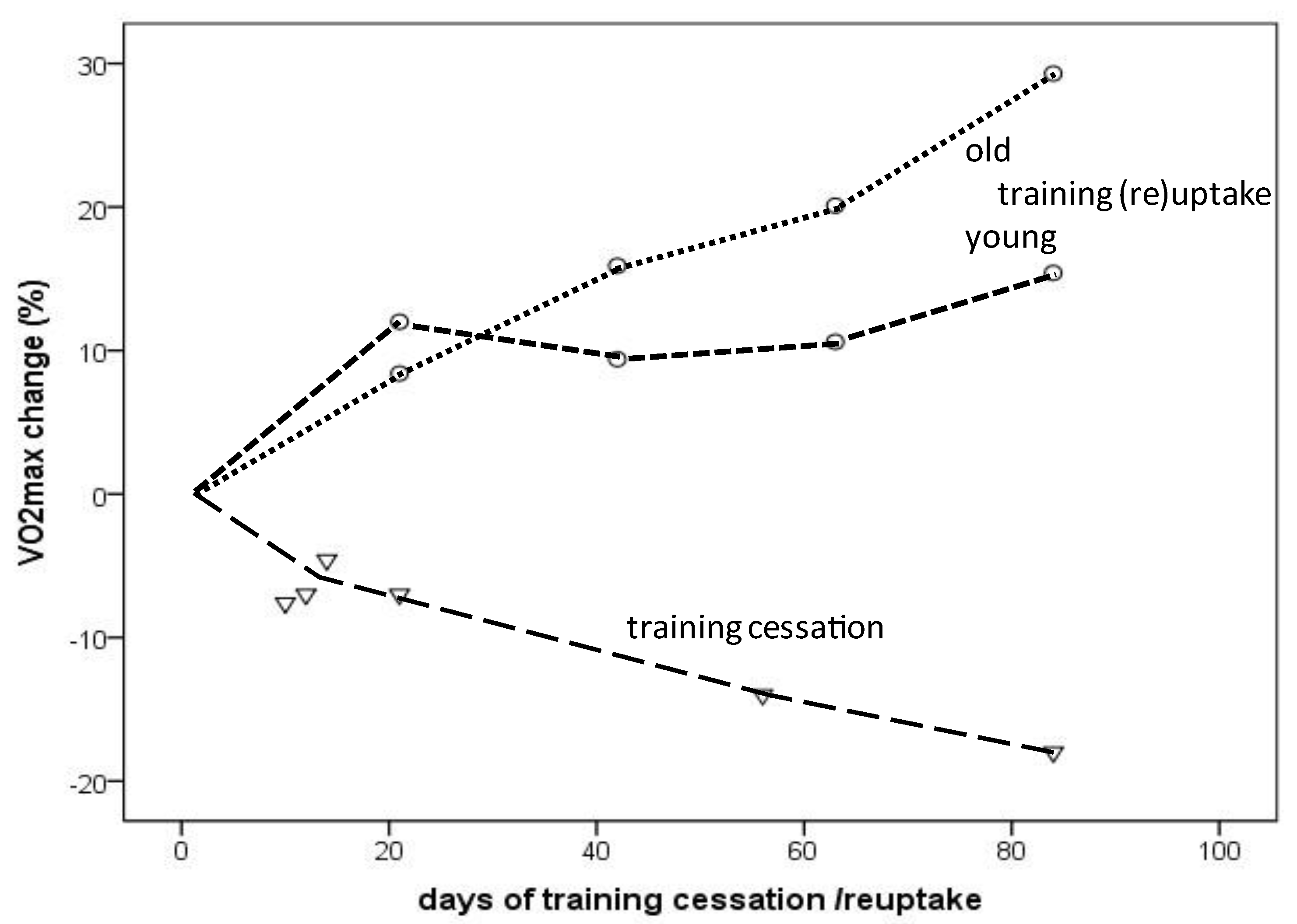{kind=link}
{kind=link}
| Reference | N m (Males) f (Females) | Observation Period (Years) | Age (Years) Post | VO2max (mL/min/kg) Pre vs. Post (% Change) Change Per Decade | Training V: Volume, km/Week I: Intensity, min/km Pre vs. Post (% Change) |
|---|---|---|---|---|---|
| Eskurza et al., 2002 [16] | 6 f | 6 | 61.0 | 45.2 ± 2.1 vs. 42.1 ± 2.1 (−7%) −12% | V: 38.0 vs. 45.8 (+20) I: 5.3 vs. 5.6 (+5%) |
| Eskuzra et al., 2002 [16] | 10 f | 6 | 56.0 | 50.0 ± 2.2 vs. 43.8 ± 2.2 (−12%) −20% | V: 62.0 vs. 42.0 (−32%) I: 5.3 vs. 5.6 (+5%) |
| Hawkins et al., 2001 [21] | 31 m | 9 | 53.5 | 58.7 ± 1.7 vs. 50.4 ± 1.5 (−14%) −16% | V: 61.8 vs. 43.6 (−29%) |
| Hawkins et al., 2001 [21] | 34 m | 8 | 62.2 | 53.4 ± 1.4 vs. 46.2 ± 1.4 (−13%) −17% | V: 56.2 vs. 43.3 (−23%) |
| Hawkins et al., 2001 [21] | 13 m | 9 | 71.1 | 46.2 ± 2.5 vs. 36.4 ± 2.6 (−21%) −23% | V: 43.8 vs. 37.5 (−14%) |
| Hawkins et al., 2001 [21] | 8 m | 7 | 82.8 | 41.5 ± 3.1 vs. 28.4 ± 2.7 (−32%) −46% | V: 49.4 vs. 26.7 (−46%) |
| Hawkins et al., 2001 [21] | 24 f | 8 | 51.2 | 48.7 ± 1.6 vs. 45.2 ± 1.2 (−7%) −9% | V: 55.1 vs. 37.7 (−22%) |
| Hawkins et al., 2001 [21] | 16 f | 8 | 58.3 | 46.7 ± 1.3 vs. 40.8 ± 1.8 (−13%) −16% | V: 39.4 vs. 31.8 (−19%) |
| Hawkins et al., 2001 [21] | 9 f | 8 | 73.2 | 39.4 ± 1.6 vs. 31.8 ± 2.8 (−19%) −24% | V: 43.6 vs. 26.6 (−39%) |
| Pollock et al. 1997 [19] | 9 m | 9.2 | 60.4 ± 8.5 | 55.4 ± 8.7 vs. 52.1 ± 6.8 (−6%) −6.5% | V: 61 vs. 55 (−10%) I: 4.9 vs. 5.2 (+6%) |
| Pollock et al. 1997 [19] | 9m | 10 | 70.4 ± 8.5 | 52.1 ± 6.8 vs. 43.2 ± 6.3 (−17%) −17% | V: 55 vs. 35 (−36%) I: 5.2 vs. 5.9 (+14%) |
| Pollock et al. 1997 [19] | 10 m | 10 | 59.5 ± 10.3 | 54.2 ± 7.7 vs. 50.0 ± 6.9 (−8%) −8% | V: 49 vs. 38 (−12%) I: 4.9 vs. 5.4 (+10%) |
| Pollock et al. 1997 [19] | 10 m | 10 | 69.8 ± 10.2 | 50.0 ± 6.9 vs. 40.8 ± 9.5 (−18%) −18% | V: 38 vs. 27 (−29%) I: 5.4 vs. 6.3 (+17%) |
| Katzel et al. 2001 [20] | 7 m | 8.7 | 70 | 51.3 ± 2.4 vs. 48.6 ± 1.8 (−5%) −6% | highly trained (no essential change) |
| Katzel et al. 2001 [20] | 21 m | 8.7 | 71 | 49.8 ± 1.1 vs. 38.2 ± 0.9 (−23%) −26% | moderately trained (volume and intensity reduction) |
| Katzel et al. 2001 [20] | 12 m | 8.7 | 74 | 49.4 ± 2.2 vs. 33.8 ± 1.8 (−32%) −36% | not trained (rather sedentary) |
| Rogers et al. 1990 [18] | 15 m | 8 | 62 | 54.0 ± 1.7 vs. 51.8 ± 1.8 (−4%) −5% | highly trained |
| Trappe et al. 1996 [17] | 10 m | 22 | 46.5 | 68.8 vs. 59.2 (−14%) −6% | highly trained |
| Trappe et al. 1996 [17] | 18 m | 22 | 46.5 | 64.1 vs. 48.9 (−24%) −11% | moderately trained |
| Trappe et al. 1996 [17] | 15 m | 22 | 46.5 | 70.7 vs. 46.7 (−34%) −15% | not trained |

4. Discussion
4.1. VO2max Decline in the Long Term: Effects of Training in Masters Athletes
4.2. Is There a Potential Role for Body Composition Explaining the VO2max Decline in Masters Athletes?
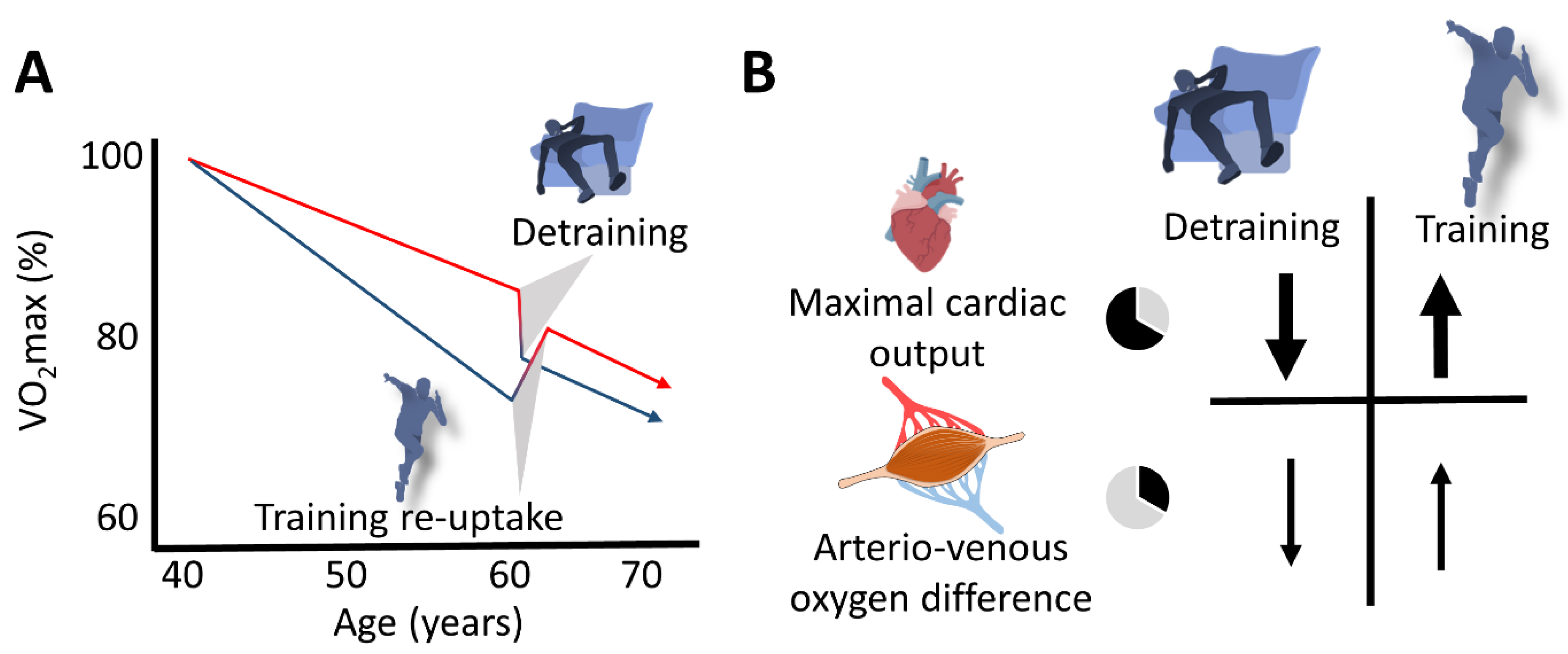
4.3. VO2max Decline in the Short Term: Effects of Training Cessation and Training (Re)Uptake
4.4. Aging-Related VO2max Loss despite Maintenance of High Training Volume and Intensity: The Slow Component
4.5. Aging-Related VO2max Decrease or Increase with Training Cessation or (Re)Uptake: The Fast Component
5. How to Support the Maintenance of Sufficient Training Stimuli in Aging Masters Athletes
6. Limitations
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Calbet, J.A.L. The biological and psychosocial aspects of successful aging in high functional elders: A longitudinal study. Scand. J. Med. Sci. Sports 2019, 29 (Suppl. S1), 5–6. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H. Aging of Competitive Athletes. Gerontology 2017, 63, 488–494. [Google Scholar] [CrossRef] [PubMed]
- Burtscher, J.; Ruedl, G.; Posch, M.; Greier, K.; Burtscher, M. The upper limit of cardiorespiratory fitness associated with longevity: An update. AIMS Public Health 2019, 6, 225–228. [Google Scholar] [CrossRef]
- Valenzuela, P.L.; Maffiuletti, N.A.; Joyner, M.J.; Lucia, A.; Lepers, R. Lifelong Endurance Exercise as a Countermeasure Against Age-Related V˙O2max Decline: Physiological Overview and Insights from Masters Athletes. Sports Med. 2020, 50, 703–716. [Google Scholar] [CrossRef]
- Burtscher, J.; Burtscher, M. Run for your life: Tweaking the weekly physical activity volume for longevity. Br. J. Sports Med. 2019, 54, 759–760. [Google Scholar] [CrossRef]
- Tanaka, H.; Seals, D.R. Endurance exercise performance in Masters athletes: Age-associated changes and underlying physiological mechanisms. J. Physiol. 2008, 586, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Clausen, J.S.; Marott, J.L.; Holtermann, A.; Gyntelberg, F.; Jensen, M.T. Midlife Cardiorespiratory Fitness and the Long-Term Risk of Mortality: 46 Years of Follow-Up. J. Am. Coll. Cardiol. 2018, 72, 987–995. [Google Scholar] [CrossRef]
- Strasser, B.; Burtscher, M. Survival of the fittest VO2 max, a key predictor of longevity. Front. Biosci. 2018, 23, 1505–1516. [Google Scholar] [CrossRef]
- Lazarus, N.R.; Harridge, S.D.R. Declining performance of master athletes: Silhouettes of the trajectory of healthy human ageing? J. Physiol. 2017, 595, 2941–2948. [Google Scholar] [CrossRef]
- Kondamudi, N.; Mehta, A.; Thangada, N.D.; Pandey, A. Physical Activity and Cardiorespiratory Fitness: Vital Signs for Cardiovascular Risk Assessment. Curr. Cardiol. Rep. 2021, 23, 172. [Google Scholar] [CrossRef]
- Saltin, B.; Blomqvist, G.; Mitchell, J.H.; Johnson, R.L.; Wildenthal, K.; Chapman, C.B. Response to exercise after bed rest and after training. Circulation 1968, 38, VII1–VII78. [Google Scholar] [PubMed]
- Nichols, J.F.; Robinson, D.; Douglass, D.; Anthony, J. Retraining of a competitive master athlete following traumatic injury: A case study. Med. Sci. Sports Exerc. 2000, 32, 1037–1042. [Google Scholar] [CrossRef] [PubMed]
- Fleg, J.L.; Morrell, C.H.; Bos, A.G.; Brant, L.J.; Talbot, L.A.; Wright, J.G.; Lakatta, E.G. Accelerated Longitudinal Decline of Aerobic Capacity in Healthy Older Adults. Circulation 2005, 112, 674–682. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, A.E.; Gentile, C.L.; Tanaka, H.; Seals, D.R.; Gates, P.E. Greater rate of decline in maximal aerobic capacity with age in endurance-trained than in sedentary men. J. Appl. Physiol. 2003, 94, 2406–2413. [Google Scholar] [CrossRef]
- Ferrari, R. Writing narrative style literature reviews. Med. Writ. 2015, 24, 230–235. [Google Scholar] [CrossRef]
- Eskurza, I.; Donato, A.J.; Moreau, K.L.; Seals, D.R.; Tanaka, H. Changes in maximal aerobic capacity with age in endurance-trained women: 7-yr follow-up. J. Appl. Physiol. 2002, 92, 2303–2308. [Google Scholar] [CrossRef] [PubMed]
- Trappe, S.W.; Costill, D.L.; Vukovich, M.; Jones, J.; Melham, T. Aging among elite distance runners: A 22-yr longitudinal study. J. Appl. Physiol. 1996, 80, 285–290. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.A.; Hagberg, J.M.; Martin, W.H.; Ehsani, A.A.; Holloszy, J.O. Decline in VO2max with aging in master athletes and sedentary men. J. Appl. Physiol. 1990, 68, 2195–2199. [Google Scholar] [CrossRef]
- Pollock, M.L.; Mengelkoch, L.J.; Graves, J.E.; Lowenthal, D.T.; Limacher, M.C.; Foster, C.; Wilmore, J.H. Twenty-year follow-up of aerobic power and body composition of older track athletes. J. Appl. Physiol. 1997, 82, 1508–1516. [Google Scholar] [CrossRef]
- Katzel, L.I.; Sorkin, J.D.; Fleg, J.L. A Comparison of Longitudinal Changes in Aerobic Fitness in Older Endurance Athletes and Sedentary Men. J. Am. Geriatr. Soc. 2001, 49, 1657–1664. [Google Scholar] [CrossRef]
- Hawkins, S.A.; Marcell, T.J.; Jaque, S.V.; Wiswell, R.A. A longitudinal assessment of change in &OV0312;O2max and maximal heart rate in master athletes. Med. Sci. Sports Exerc. 2001, 33, 1744–1750. [Google Scholar] [CrossRef] [PubMed]
- Coyle, E.F.; Martin, W.H.; Sinacore, D.R.; Joyner, M.J.; Hagberg, J.M.; Holloszy, J.O. Time course of loss of adaptations after stopping prolonged intense endurance training. J. Appl. Physiol. 1984, 57, 1857–1864. [Google Scholar] [CrossRef] [PubMed]
- Cullinane, E.M.; Sady, S.P.; Vadeboncoeur, L.; Burke, M.; Thompson, P.D. Cardiac size and VO2max do not decrease after short-term exercise cessation. Med. Sci. Sports Exerc. 1986, 18, 420–424. [Google Scholar] [CrossRef] [PubMed]
- Doherty, R.A.; Neary, J.P.; Bhambhani, Y.N.; Wenger, H.A. Fifteen-day cessation of training on selected physiological and performance variables in women runners. J. Strength Cond. Res. 2003, 17, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Drinkwater, B.L.; Horvath, S.M. Detraining effects on young women. Med. Sci. Sports 1972, 4, 91–95. [Google Scholar] [CrossRef]
- Giada, F.; Bertaglia, E.; De Piccoli, B.; Franceschi, M.; Sartori, F.; Raviele, A.; Pascotto, P. Cardiovascular adaptations to endurance training and detraining in young and older athletes. Int. J. Cardiol. 1998, 65, 149–155. [Google Scholar] [CrossRef]
- Heath, G.W.; Gavin, J.R.; Hinderliter, J.M.; Hagberg, J.M.; Bloomfield, S.A.; Holloszy, J.O. Effects of exercise and lack of exercise on glucose tolerance and insulin sensitivity. J. Appl. Physiol. 1983, 55, 512–517. [Google Scholar] [CrossRef]
- Houmard, J.A.; Hortobágyi, T.; Johns, R.A.; Bruno, N.J.; Nute, C.C.; Shinebarger, M.H.; Welborn, J.W. Effect of Short-Term Training Cessation on Performance Measures in Distance Runners. Int. J. Sports Med. 1992, 13, 572–576. [Google Scholar] [CrossRef]
- Katzel, L.I.; Busby-Whitehead, M.J.; Hagberg, J.M.; Fleg, J.L. Abnormal exercise electrocardiograms in master athletes after three months of deconditioning. J. Am. Geriatr. Soc. 1997, 45, 744–746. [Google Scholar] [CrossRef]
- Martin, W.H., 3rd; Coyle, E.F.; Bloomfield, S.A.; Ehsani, A.A. Effects of physical deconditioning after intense endurance training on left ventricular dimensions and stroke volume. J. Am. Coll. Cardiol. 1986, 7, 982–989. [Google Scholar] [CrossRef] [Green Version]
- Murias, J.M.; Kowalchuk, J.M.; Paterson, D.H. Time course and mechanisms of adaptations in cardiorespiratory fitness with endurance training in older and young men. J. Appl. Physiol. 2010, 108, 621–627. [Google Scholar] [CrossRef] [PubMed]
- Pavlik, G.; Bachl, N.; Wollein, W.; Lángfy, G.; Prokop, L. Resting Echocardiographic Parameters After Cessation of Regular Endurance Training. Int. J. Sports Med. 1986, 7, 226–231. [Google Scholar] [CrossRef] [PubMed]
- Prior, S.J.; Goldberg, A.P.; Ortmeyer, H.K.; Chin, E.R.; Chen, D.; Blumenthal, J.B.; Ryan, A.S. Increased Skeletal Muscle Capillarization Independently Enhances Insulin Sensitivity in Older Adults After Exercise Training and Detraining. Diabetes 2015, 64, 3386–3395. [Google Scholar] [CrossRef] [PubMed]
- Ready, A.E.; Quinney, H.A. Alterations in anaerobic threshold as the result of endurance training and detraining. Med. Sci. Sports Exerc. 1982, 14, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Schulman, S.P.; Fleg, J.L.; Goldberg, A.P.; Busby-Whitehead, J.; Hagberg, J.M.; O’Connor, F.C.; Gerstenblith, G.; Becker, L.C.; Katzel, L.I.; Lakatta, L.E.; et al. Continuum of Cardiovascular Performance Across a Broad Range of Fitness Levels in Healthy Older Men. Circulation 1996, 94, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Sinacore, D.R.; Coyle, E.F.; Hagberg, J.M.; Holloszy, J.O. Histochemical and Physiological Correlates of Training- and Detraining-Induced Changes in the Recovery From a Fatigue Test. Phys. Ther. 1993, 73, 661–667. [Google Scholar] [CrossRef]
- Smorawiński, J.; Nazar, K.; Kaciuba-Uscilko, H.; Kamińska, E.; Cybulski, G.; Kodrzycka, A.; Bicz, B.; Greenleaf, J.E. Effects of 3-day bed rest on physiological responses to graded exercise in athletes and sedentary men. J. Appl. Physiol. 2001, 91, 249–257. [Google Scholar] [CrossRef]
- Houston, M.E.; Bentzen, H.; Larsen, H. Interrelationships between skeletal muscle adaptations and performance as studied by detraining and retraining. Acta Physiol. Scand. 1979, 105, 163–170. [Google Scholar] [CrossRef]
- Ganse, B.; Kleerekoper, A.; Knobe, M.; Hildebrand, F.; Degens, H. Longitudinal trends in master track and field performance throughout the aging process: 83,209 results from Sweden in 16 athletics disciplines. GeroScience 2020, 42, 1609–1620. [Google Scholar] [CrossRef]
- Kusy, K.; Zieliński, J. Aerobic capacity in speed-power athletes aged 20-90 years vs endurance runners and untrained participants. Scand. J. Med. Sci. Sports 2014, 24, 68–79. [Google Scholar] [CrossRef]
- Bagley, L.; McPhee, J.S.; Ganse, B.; Müller, K.; Korhonen, M.T.; Rittweger, J.; Degens, H. Similar relative decline in aerobic and anaerobic power with age in endurance and power master athletes of both sexes. Scand. J. Med. Sci. Sport. 2019, 29, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Wiswell, R.A.; Hawkins, S.A.; Jaque, S.V.; Hyslop, D.; Constantino, N.; Tarpenning, K.; Marcell, T.; Schroeder, E.T. Relationship between physiological loss, performance decrement, and age in master athletes. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2001, 56, M618–M626. [Google Scholar] [CrossRef] [PubMed]
- Ganse, B.; Drey, M.; Hildebrand, F.; Knobe, M.; Degens, H. Performance Declines Are Accelerated in the Oldest-Old Track and Field Athletes 80 to 94 Years of Age. Rejuvenation Res. 2020, 24, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Sousa, C.V.; Da Silva Aguiar, S.; Rosemann, T.; Nikolaidis, P.T.; Knechtle, B. American Masters Road Running Records—The Performance Gap Between Female and Male Age Group Runners from 5 Km to 6 Days Running. Int. J. Environ. Res. Public Health 2019, 16, 2310. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Canestro, C.; Montero, D. Sex Dimorphism of VO2max Trainability: A Systematic Review and Meta-analysis. Sports Med. 2019, 49, 1949–1956. [Google Scholar] [CrossRef] [PubMed]
- Grimsmo, J.; Arnesen, H.; Maehlum, S.; Mæhlum, S. Changes in cardiorespiratory function in different groups of former and still active male cross-country skiers: A 28-30-year follow-up study. Scand. J. Med. Sci. Sports 2010, 20, e151–e161. [Google Scholar] [CrossRef]
- Reaburn, P.; Dascombe, B. Endurance performance in masters athletes. Eur. Rev. Aging Phys. Act. 2008, 5, 31–42. [Google Scholar] [CrossRef]
- Ogawa, T.; Spina, R.J.; Martin, W.H.; Kohrt, W.M.; Schechtman, K.B.; Holloszy, J.O.; Ehsani, A.A. Effects of aging, sex, and physical training on cardiovascular responses to exercise. Circulation 1992, 86, 494–503. [Google Scholar] [CrossRef]
- Burtscher, M. Exercise Limitations by the Oxygen Delivery and Utilization Systems in Aging and Disease: Coordinated Adaptation and Deadaptation of the Lung-Heart Muscle Axis—A Mini-Review. Gerontology 2013, 59, 289–296. [Google Scholar] [CrossRef]
- Kohrt, W.M.; Malley, M.T.; Coggan, A.R.; Spina, R.J.; Ogawa, T.; Ehsani, A.A.; Bourey, R.E.; Martin, W.H.; Holloszy, J.O. Effects of gender, age, and fitness level on response of VO2max to training in 60–71 yr olds. J. Appl. Physiol. 1991, 71, 2004–2011. [Google Scholar] [CrossRef]
- Tayrose, G.A.; Beutel, B.G.; Cardone, D.A.; Sherman, O.H. The Masters Athlete: A Review of Current Exercise and Treatment Recommendations. Sports Health 2015, 7, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Wilke, J.; Vogel, O.; Vogt, L. Why Are You Running and Does It Hurt? Pain, Motivations and Beliefs about Injury Prevention among Participants of a Large-Scale Public Running Event. Int. J. Environ. Res. Public Heath 2019, 16, 3766. [Google Scholar] [CrossRef] [PubMed]
- Malchrowicz-Mośko, E.; Gravelle, F.; Dąbrowska, A.; León-Guereño, P. Do Years of Running Experience Influence the Motivations of Amateur Marathon Athletes? Int. J. Environ. Res. Public Health 2020, 17, 585. [Google Scholar] [CrossRef] [PubMed]
- Burtscher, J.; Vanderriele, P.-E.; Legrand, M.; Predel, H.-G.; Niebauer, J.; O’Keefe, J.H.; Millet, G.P.; Burtscher, M. Could Repeated Cardio-Renal Injury Trigger Late Cardiovascular Sequelae in Extreme Endurance Athletes? Sports Med. 2022, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, S.A.; Wiswell, R.A. Rate and mechanism of maximal oxygen consumption decline with aging: Implications for exercise training. Sports Med. 2003, 33, 877–888. [Google Scholar] [CrossRef]
- Chambers, T.L.; Burnett, T.R.; Raue, U.; Lee, G.A.; Finch, W.H.; Graham, B.M.; Trappe, T.A.; Trappe, S. Skeletal muscle size, function, and adiposity with lifelong aerobic exercise. J. Appl. Physiol. 2020, 128, 368–378. [Google Scholar] [CrossRef]
- Pollock, R.D.; Carter, S.; Velloso, C.P.; Duggal, N.A.; Lord, J.M.; Lazarus, N.R.; Harridge, S.D.R. An investigation into the relationship between age and physiological function in highly active older adults. J. Physiol. 2015, 593, 657–680. [Google Scholar] [CrossRef]
- Alvero-Cruz, J.R.; Brikis, M.; Chilibeck, P.; Frings-Meuthen, P.; Guzmán, J.F.V.; Mittag, U.; Michely, S.; Mulder, E.; Tanaka, H.; Tank, J.; et al. Age-Related Decline in Vertical Jumping Performance in Masters Track and Field Athletes: Concomitant Influence of Body Composition. Front. Physiol. 2021, 12, 643649. [Google Scholar] [CrossRef]
- Campa, F.; Toselli, S.; Mazzilli, M.; Gobbo, L.A.; Coratella, G. Assessment of Body Composition in Athletes: A Narrative Review of Available Methods with Special Reference to Quantitative and Qualitative Bioimpedance Analysis. Nutrients 2021, 13, 1620. [Google Scholar] [CrossRef]
- Huang, G.; Gibson, C.A.; Tran, Z.V.; Osness, W.H. Controlled Endurance Exercise Training and VO2max Changes in Older Adults: A Meta-Analysis. Prev. Cardiol. 2005, 8, 217–225. [Google Scholar] [CrossRef]
- Støren, O.; Helgerud, J.; Sæbø, M.; Støa, E.M.; Bratland-Sanda, S.; Unhjem, R.J.; Hoff, J.; Wang, E. The Effect of Age on the VO2max Response to High-Intensity Interval Training. Med. Sci. Sports Exerc. 2017, 49, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Burtscher, J.; Millet, G.P.; Gatterer, H.; Vonbank, K.; Burtscher, M. Does Regular Physical Activity Mitigate the Age-Associated Decline in Pulmonary Function? Sports Med. 2022, 52, 963–970. [Google Scholar] [CrossRef]
- Rivera, A.M.; Pels, A.E.; Sady, S.P.; Sady, M.A.; Cullinane, E.M.; Thompson, P.D. Physiological factors associated with the lower maximal oxygen consumption of master runners. J. Appl. Physiol. 1989, 66, 949–954. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Monahan, K.D.; Seals, D.R. Age-predicted maximal heart rate revisited. J. Am. Coll. Cardiol. 2001, 37, 153–156. [Google Scholar] [CrossRef]
- Christou, D.D.; Seals, D.R. Decreased maximal heart rate with aging is related to reduced β-adrenergic responsiveness but is largely explained by a reduction in intrinsic heart rate. J. Appl. Physiol. 2008, 105, 24–29. [Google Scholar] [CrossRef]
- Schulman, S.P.; Lakatta, E.; Fleg, J.L.; Lakatta, L.; Becker, L.C.; Gerstenblith, G. Age-related decline in left ventricular filling at rest and exercise. Am. J. Physiol. Circ. Physiol. 1992, 263, H1932–H1938. [Google Scholar] [CrossRef]
- Pandey, A.; Kraus, W.E.; Brubaker, P.H.; Kitzman, D.W. Healthy Aging and Cardiovascular Function: Invasive Hemodynamics During Rest and Exercise in 104 Healthy Volunteers. JACC Heart Fail. 2019, 8, 111–121. [Google Scholar] [CrossRef]
- Steding-Ehrenborg, K.; Boushel, R.C.; Calbet, J.A.; Åkeson, P.; Mortensen, S.P. Left ventricular atrioventricular plane displacement is preserved with lifelong endurance training and is the main determinant of maximal cardiac output. J. Physiol. 2015, 593, 5157–5166. [Google Scholar] [CrossRef]
- Seals, D.R.; Moreau, K.L.; Gates, P.E.; Eskurza, I. Modulatory influences on ageing of the vasculature in healthy humans. Exp. Gerontol. 2006, 41, 501–507. [Google Scholar] [CrossRef]
- Nyberg, M.; Blackwell, J.R.; Damsgaard, R.; Jones, A.M.; Hellsten, Y.; Mortensen, S. Lifelong physical activity prevents an age-related reduction in arterial and skeletal muscle nitric oxide bioavailability in humans. J. Physiol. 2012, 590, 5361–5370. [Google Scholar] [CrossRef] [Green Version]
- Calbet, J.A.L. Ageing, exercise and cardiovascular health: Good and bad news. J. Physiol. 2012, 590, 5265–5266. [Google Scholar] [CrossRef] [PubMed]
- McKendry, J.; Joanisse, S.; Baig, S.; Liu, B.; Parise, G.; Greig, C.; Breen, L. Superior Aerobic Capacity and Indices of Skeletal Muscle Morphology in Chronically Trained Master Endurance Athletes Compared With Untrained Older Adults. J. Gerontol. Ser. A 2020, 75, 1079–1088. [Google Scholar] [CrossRef]
- Coyle, E.F.; Hemmert, M.K.; Coggan, A.R. Effects of detraining on cardiovascular responses to exercise: Role of blood volume. J. Appl. Physiol. 1986, 60, 95–99. [Google Scholar] [CrossRef]
- Petrella, R.J.; Cunningham, D.A.; Paterson, D.H. Effects of 5-Day Exercise Training in Elderly Subjects on Resting Left Ventricular Diastolic Function and VO2max. Can. J. Appl. Physiol. 1997, 22, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Convertino, V.A. Blood volume: Its adaptation to endurance training. Med. Sci. Sports Exerc. 1991, 23, 1338–1348. [Google Scholar] [CrossRef]
- Sawka, M.N.; Convertino, V.A.; Eichner, E.R.; Schnieder, S.M.; Young, A.J. Blood volume: Importance and adaptations to exercise training, environmental stresses, and trauma/sickness. Med. Sci. Sports Exerc. 2000, 32, 332–348. [Google Scholar] [CrossRef] [PubMed]
- Boushel, R.; Gnaiger, E.; Calbet, J.A.; Gonzalez-Alonso, J.; Wright-Paradis, C.; Sondergaard, H.; Ara, I.; Helge, J.W.; Saltin, B. Muscle mitochondrial capacity exceeds maximal oxygen delivery in humans. Mitochondrion 2011, 11, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Boushel, R.; Gnaiger, E.; Larsen, F.J.; Helge, J.W.; González-Alonso, J.; Ara, I.; Munch-Andersen, T.; van Hall, G.; Søndergaard, H.; Saltin, B.; et al. Maintained peak leg and pulmonary VO2 despite substantial reduction in muscle mitochondrial capacity. Scand. J. Med. Sci. Sports 2015, 25 (Suppl. S4), 135–143. [Google Scholar] [CrossRef] [PubMed]
- Hearon, C.M.; DiNenno, F.A. Regulation of skeletal muscle blood flow during exercise in ageing humans. J. Physiol. 2016, 594, 2261–2273. [Google Scholar] [CrossRef]
- Koch, D.W.; Newcomer, S.C.; Proctor, D.N. Blood Flow to Exercising Limbs Varies With Age, Gender, and Training Status. Can. J. Appl. Physiol. 2005, 30, 554–575. [Google Scholar] [CrossRef]
- Jendzjowsky, N.G.; DeLorey, D.S. Short-term exercise training enhances functional sympatholysis through a nitric oxide-dependent mechanism. J. Physiol. 2013, 591, 1535–1549. [Google Scholar] [CrossRef] [PubMed]
- DeLorey, D.S. Sympathetic vasoconstriction in skeletal muscle: Modulatory effects of aging, exercise training, and sex. Appl. Physiol. Nutr. Metab. 2021, 46, 1437–1447. [Google Scholar] [CrossRef] [PubMed]
- Hoier, B.; Hellsten, Y. Exercise-Induced Capillary Growth in Human Skeletal Muscle and the Dynamics of VEGF. Microcirculation 2014, 21, 301–314. [Google Scholar] [CrossRef] [PubMed]
- Degens, H. Age-Related Changes in the Microcirculation of Skeletal Muscle. Adv. Exp. Med. Biol. 1998, 454, 343–348. [Google Scholar] [CrossRef]
- DeLorey, D.S.; Paterson, D.H.; Kowalchuk, J.M. Effects of ageing on muscle O2 utilization and muscle oxygenation during the transition to moderate-intensity exercise. Appl. Physiol. Nutr. Metab. 2007, 32, 1251–1262. [Google Scholar] [CrossRef]
- Sun, N.; Youle, R.J.; Finkel, T. The Mitochondrial Basis of Aging. Mol. Cell 2016, 61, 654–666. [Google Scholar] [CrossRef]
- Chan, D.C. Mitochondria: Dynamic Organelles in Disease, Aging, and Development. Cell 2006, 125, 1241–1252. [Google Scholar] [CrossRef]
- Hood, D.A.; Memme, J.M.; Oliveira, A.N.; Triolo, M. Maintenance of Skeletal Muscle Mitochondria in Health, Exercise, and Aging. Annu. Rev. Physiol. 2019, 81, 19–41. [Google Scholar] [CrossRef]
- Huertas, J.R.; Casuso, R.A.; Agustín, P.H.; Cogliati, S. Stay Fit, Stay Young: Mitochondria in Movement: The Role of Exercise in the New Mitochondrial Paradigm. Oxidative Med. Cell. Longev. 2019, 2019, 7058350. [Google Scholar] [CrossRef]
- Granata, C.; Caruana, N.J.; Botella, J.; Jamnick, N.A.; Huynh, K.; Kuang, J.; Janssen, H.A.; Reljic, B.; Mellett, N.A.; Laskowski, A. Multi-omics reveal intricate network of mitochondrial adaptations to training in human skeletal muscle. bioRxiv 2021. [Google Scholar] [CrossRef]
- Bouzid, M.A.; Filaire, E.; Matran, R.; Robin, S.; Fabre, C. Lifelong Voluntary Exercise Modulates Age-Related Changes in Oxidative Stress. Inter. J. Sports Med. 2018, 39, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Betik, A.C.; Hepple, R.T. Determinants of VO2 max decline with aging: An integrated perspective. Appl. Physiol. Nutr. Metab. 2008, 33, 130–140. [Google Scholar] [CrossRef] [PubMed]
- Lepers, R.; Stapley, P.J. Master Athletes Are Extending the Limits of Human Endurance. Front. Physiol. 2016, 7, 613. [Google Scholar] [CrossRef]
- Mckean, K.A.; Manson, N.A.; Stanish, W.D. Musculoskeletal Injury in the Masters Runners. Clin. J. Sport Med. 2006, 16, 149–154. [Google Scholar] [CrossRef]
- Medic, N.; Starkes, J.L.; Young, B. Examining relative age effects on performance achievement and participation rates in Masters athletes. J. Sports Sci. 2007, 25, 1377–1384. [Google Scholar] [CrossRef] [PubMed]
- Pugliese, L.; Porcelli, S.; Vezzoli, A.; La Torre, A.; Serpiello, F.R.; Pavei, G.; Marzorati, M. Different Training Modalities Improve Energy Cost and Performance in Master Runners. Front. Physiol. 2018, 9, 21. [Google Scholar] [CrossRef] [PubMed]
- Stöggl, T.; Sperlich, B. Polarized training has greater impact on key endurance variables than threshold, high intensity, or high volume training. Front. Physiol. 2014, 5, 33. [Google Scholar] [CrossRef] [PubMed]
- Hawley, J.A.; Bishop, D.J. High-intensity exercise training—too much of a good thing? Nat. Rev. Endocrinol. 2021, 17, 385–386. [Google Scholar] [CrossRef]
- Daussin, F.N.; Zoll, J.; Dufour, S.P.; Ponsot, E.; Lonsdorfer-Wolf, E.; Doutreleau, S.; Mettauer, B.; Piquard, F.; Geny, B.; Richard, R. Effect of interval versus continuous training on cardiorespiratory and mitochondrial functions: Relationship to aerobic performance improvements in sedentary subjects. Am. J. Physiol. Integr. Comp. Physiol. 2008, 295, R264–R272. [Google Scholar] [CrossRef]
- Valenzuela, P.L.; Santos-Lozano, A.; Lucia, A. More on the Record-Breaking Performance in a 70-Year-Old Marathoner. N. Engl. J. Med. 2019, 381, 293–294. [Google Scholar] [CrossRef]
- Pantoja, P.D.; Morin, J.B.; Peyré-Tartaruga, L.A.; Brisswalter, J. Running Energy Cost and Spring-Mass Behavior in Young versus Older Trained Athletes. Med. Sci. Sports Exerc. 2016, 48, 1779–1786. [Google Scholar] [CrossRef] [PubMed]
- Murias, J.M.; Kowalchuk, J.M.; Ritchie, D.; Hepple, R.T.; Doherty, T.J.; Paterson, D.H. Adaptations in Capillarization and Citrate Synthase Activity in Response to Endurance Training in Older and Young Men. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2011, 66, 957–964. [Google Scholar] [CrossRef] [PubMed]
- Granata, C.; Jamnick, N.A.; Bishop, D.J. Principles of Exercise Prescription, and How They Influence Exercise-Induced Changes of Transcription Factors and Other Regulators of Mitochondrial Biogenesis. Sports Med. 2018, 48, 1541–1559. [Google Scholar] [CrossRef] [PubMed]
- Holloszy, J.O. Biochemical adaptations in muscle. Effects of exercise on mitochondrial oxygen uptake and respiratory enzyme activity in skeletal muscle. J. Biol. Chem. 1967, 242, 2278–2282. [Google Scholar] [CrossRef]
- MacInnis, M.J.; Gibala, M.J. Physiological adaptations to interval training and the role of exercise intensity. J. Physiol. 2017, 595, 2915–2930. [Google Scholar] [CrossRef]
- Granata, C.; Oliveira, R.S.F.; Little, J.P.; Renner, K.; Bishop, D.J. Mitochondrial adaptations to high-volume exercise training are rapidly reversed after a reduction in training volume in human skeletal muscle. FASEB J. 2016, 30, 3413–3423. [Google Scholar] [CrossRef]
- Granata, C.; Oliveira, R.S.F.; Little, J.P.; Bishop, D.J. Forty high-intensity interval training sessions blunt exercise-induced changes in the nuclear protein content of PGC-1α and p53 in human skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2020, 318, E224–E236. [Google Scholar] [CrossRef]
- Poole, D.C.; Jones, A.M. Oxygen Uptake Kinetics. Compr. Physiol. 2012, 2, 933–996. [Google Scholar] [CrossRef]
- Babcock, M.A.; Paterson, D.H.; Cunningham, D.A.; Dickinson, J.R. Exercise on-transient gas exchange kinetics are slowed as a function of age. Med. Sci. Sports Exerc. 1994, 26, 440–446. [Google Scholar] [CrossRef]
- Millet, G.P.; Candau, R.; Fattori, P.; Bignet, F.; Varray, A. VO2 responses to different intermittent runs at velocity associated with VO2max. Can. J. Appl. Physiol. 2003, 28, 410–423. [Google Scholar] [CrossRef]
- Billat, V.L.; Slawinksi, J.; Bocquet, V.; Chassaing, P.; Demarle, A.; Koralsztein, J.P. Very Short (15 s–15 s) Interval-Training Around the Critical Velocity Allows Middle-Aged Runners to Maintain VO2 max for 14 minutes. Int. J. Sports Med. 2001, 22, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Bowen, T.S.; Schuler, G.; Adams, V. Skeletal muscle wasting in cachexia and sarcopenia: Molecular pathophysiology and impact of exercise training. J. Cachexia Sarcopenia Muscle 2015, 6, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Borst, S.E. Interventions for sarcopenia and muscle weakness in older people. Age Ageing 2004, 33, 548–555. [Google Scholar] [CrossRef] [PubMed]
- Rynecki, N.D.; Siracuse, B.L.; Ippolito, J.A.; Beebe, K.S. Injuries sustained during high intensity interval training: Are modern fitness trends contributing to increased injury rates? J. Sports Med. Phys. Fit. 2019, 59, 1206–1212. [Google Scholar] [CrossRef] [PubMed]
- Mor, A.; Grijota, M.; Nørgaard, M.; Munthe, J.; Lind, M.; Déruaz, A.; Pedersen, A.B. Trends in arthroscopy-documented cartilage injuries of the knee and repair procedures among 15-60-year-old patients. Scand. J. Med. Sci. Sports 2015, 25, e400–e407. [Google Scholar] [CrossRef]
- Campos, Y.; Casado, A.; Vieira, J.G.; Guimarães, M.; Sant’Ana, L.; Leitão, L.; da Silva, S.F.; de Azevedo, P.H.S.M.; Vianna, J.; Domínguez, R. Training-intensity Distribution on Middle- and Long-distance Runners: A Systematic Review. Int. J. Sports Med. 2021, 43, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Bishop, D.J.; Botella, J.; Granata, C. CrossTalk opposing view: Exercise training volume is more important than training intensity to promote increases in mitochondrial content. J. Physiol. 2019, 597, 4115–4118. [Google Scholar] [CrossRef]
- MacInnis, M.J.; Skelly, L.E.; Gibala, M.J. CrossTalk proposal: Exercise training intensity is more important than volume to promote increases in human skeletal muscle mitochondrial content. J. Physiol. 2019, 597, 4111–4113. [Google Scholar] [CrossRef]
- Burtscher, J.; Mallet, R.T.; Pialoux, V.; Millet, G.P.; Burtscher, M. Adaptive Responses to Hypoxia and/or Hyperoxia in Humans. Antioxid. Redox Signal. 2022. [Google Scholar] [CrossRef]
- Menéndez, A.F.; Saudan, G.; Sperisen, L.; Hans, D.; Saubade, M.; Millet, G.P.; Malatesta, D. Effects of Short-Term Normobaric Hypoxic Walking Training on Energetics and Mechanics of Gait in Adults with Obesity. Obesity 2018, 26, 819–827. [Google Scholar] [CrossRef] [Green Version]
- Girard, O.; Malatesta, D.; Millet, G.P. Walking in Hypoxia: An Efficient Treatment to Lessen Mechanical Constraints and Improve Health in Obese Individuals? Front. Physiol. 2017, 8, 73. [Google Scholar] [CrossRef] [PubMed]
- Libardi, C.A.; Chacon-Mikahil, M.P.; Cavaglieri, C.R.; Tricoli, V.; Roschel, H.; Vechin, F.C.; Conceicao, M.S.; Ugrinowitsch, C. Effect of concurrent training with blood flow restriction in the elderly. Int. J. Sports Med. 2015, 36, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Millet, G.P.; Debevec, T.; Brocherie, F.; Malatesta, D.; Girard, O. Therapeutic Use of Exercising in Hypoxia: Promises and Limitations. Front. Physiol. 2016, 7, 224. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Kawahara, T.; Aono, H.; Yamada, S.; Ishizuka, S.; Takahashi, K.; Iijima, K. A comparison of sarcopenia prevalence between former Tokyo 1964 Olympic athletes and general community-dwelling older adults. J. Cachex. Sarcopenia Muscle 2021, 12, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Methenitis, S.; Mouratidis, A.; Manga, K.; Chalari, E.; Feidantsis, K.; Arnaoutis, G.; Arailoudi-Alexiadou, X.; Skepastianos, P.; Hatzitolios, A.; Mourouglakis, A.; et al. The importance of protein intake in master marathon runners. Nutrition 2021, 86, 111154. [Google Scholar] [CrossRef]
- Moore, D.R. Protein Requirements for Master Athletes: Just Older Versions of Their Younger Selves. Sports Med. 2021, 51, 13–30. [Google Scholar] [CrossRef]
- Lepers, R.; Bontemps, B.; Louis, J. Physiological Profile of a 59-Year-Old Male World Record Holder Marathoner. Med. Sci. Sports Exerc. 2020, 52, 623–626. [Google Scholar] [CrossRef]
- Robinson, A.T.; Watso, J.C.; Babcock, M.C.; Joyner, M.J.; Farquhar, W.B. Record-Breaking Performance in a 70-Year-Old Marathoner. N. Engl. J. Med. 2019, 380, 1485–1486. [Google Scholar] [CrossRef]


| Reference | N m (Males) f (Females) | Duration (Days) of Training Cessation (ce) Training (Re)Uptake (re) | Age (Years) | VO2max (mL/min/kg) Pre vs. Post (% Change) | Changes in Related Physiological Parameters (% Change) |
|---|---|---|---|---|---|
| Coyle et al., 1984 [22] | 6 m 1 f | 12 (ce) | 29.1 ± 3.2 | 62.1 ± 3.3 vs. 57.7 ± 2.6 (−7%) | heart rate (+4%) stroke volume (−10%) cardiac output (−7%) arteriovenous O2diff (+0.4%) oxygen pulse (−11%) citrate synthase (−17.1%) succinate dehydrogenase (−18.5%) |
| Coyle et al., 1984 [22] | 6 m 1 f | 21 (ce) | 29.1 ± 3.2 | 62.1 ± 3.3 vs. 59.7 ± 3.1 (−7%) | heart rate (+4%) stroke volume (−11%) cardiac output (−8%) arteriovenous O2diff (+2%) oxygen pulse (−11%) citrate synthase (−23.7%) succinate dehydrogenase (−23.9%) |
| Coyle et al., 1984 [22] | 6 m 1 f | 56 (ce) | 29.1 ± 3.2 | 62.1 ± 3.3 vs. 53.2 ± 2.1 (−14%) | heart rate (+6%) stroke volume (−14%) cardiac output (−9%) arteriovenous O2diff (−4%) oxygen pulse (−19%) citrate synthase (−40.6%) succinate dehydrogenase (−38.4%) |
| Coyle et al., 1984 [22] | 6 m 1 f | 84 (ce) | 29.1 ± 3.2 | 62.1 ± 3.3 vs. 50.8 ± 1.9 (−18%) | heart rate (+5%) stroke volume (−13%) cardiac output (−10%) arteriovenous O2diff (−7%) oxygen pulse (−20%) citrate synthase (−39.6%) succinate dehydrogenase (−32.5%) |
| Cullinane et al., 1986 [23] | 15 m | 10 (ce) | 28.2 ± 5.6 | 61.3 ± 6.2 vs. 61.2 ± 5.6 (−1.6%) | heart rate (+5%) ventilation (+1.5%) stroke volume (−2.6%) |
| Doherty et al., 2003 [24] | 7 f | 10 (ce) | 21.0 ± 2.6 | 49.8 ± 1.3 vs. 46.0 ± 1.3 (−7.6%) | heart rate (+1.5%) stroke volume (−1%) cardiac output (−0.5%) arteriovenous O2diff (−7%) |
| Drinkwater and Horwath, 1972 [25] | 7 f | 90 (ce) | 14-17 | 47.8 ± 1.8 vs. 40.4 ± 1.0 (−15.4%) | heart rate (+1.5%) ventilation (−10.3%) |
| Giada et al., 1998 [26] | 12 m | 60 (ce) | 55 ± 5 | 43 ± 7 vs. 36 ± 7 (−16.3%) | |
| Giada et al., 1998 | 12 m | 60 (ce) | 24 ± 6 | 59 ± 10 vs. 49 ± 9 (−16.9%) | |
| Heath et al., 1983 [27] | 6 m 2 f | 10 (ce) | 28 ± 3 | 58.6 ± 2.2 vs. 57.6 ± 2.1 (−1.7%) | |
| Houmard et al., 1992 [28] | 9 m 3 f | 14 (ce) | 20.1 ± 1.4 | 61.6 ± 2.2 vs. 58.7 ± 1.8 (−4.6%) | heart rate (+4.7%) plasma volume (−5.1%) citrate synthase (−25.3%) |
| Katzel et al., 1997 [29] | 10 m | 90 (ce) | 59 ± 8 | 50 ± 5 (−11 to −20%) | |
| Martin et al., 1986 [30] | 5 m 1 f | 42 (ce) | 26 ± 1 | 62.7 ± 4.0 (−6.5%), 21 days; (−20.3%), 56 days | stroke volume 21 days (−10%) 56 days (−17%) |
| Murias et al., 2010 [31] | 8 m | 21 (re) | 68.0 ± 7.0 | 28.3 ± 7.1 vs. 30.7 ± 6.0 (+8.4%) | heart rate (−3.5%) stroke volume (+6.8%) cardiac output (+7%) arteriovenous O2diff (+3.7%) |
| Murias et al., 2010 [31] | 8 m | 21 (re) | 23.0 ± 5.0 | 48.0 ± 6.1 vs. 53.8 ± 7.6 (+12%) | heart rate (−2.1%) stroke volume (+5.5%) cardiac output (+3.1%) arteriovenous O2diff (+7.5%) |
| Murias et al., 2010 [31] | 8 m | 42 (re) | 68.0 ± 7.0 | 28.3 ± 7.1 vs. 32.8 ± 7.6 (+15.9%) | heart rate (−2.1%) stroke volume (+9.1%) cardiac output (+11.3%) arteriovenous O2diff (+5.2%) |
| Murias et al., 2010 [31] | 8 m | 42 (re) | 23.0 ± 5.0 | 48.0 ± 6.1 vs. 52.5 ± 6.4% (+9.4%) | heart rate (−2.1%) stroke volume (+7.9%) cardiac output (+5.4%) arteriovenous O2diff (+4.8%) |
| Murias et al., 2010 [31] | 8 m | 63 (re) | 68.0 ± 7.0 | 28.3 ± 7.1 vs. 34.0 ± 5.8 (+20.1%) | heart rate (−1.4%) stroke volume (+15.2%) cardiac output (+17.9%) arteriovenous O2diff (+3.7%) |
| Murias et al., 2010 [31] | 8 m | 63 (re) | 23.0 ± 5.0 | 48.0 ± 6.1 vs. 53.1 ± 6.5% (+10.6%) | heart rate (−2.1%) stroke volume (+12.6%) cardiac output (+10.4%) arteriovenous O2diff (+0.7%) |
| Murias et al., 2010 [31] | 8 m | 84 (re) | 68.0 ± 7.0 | 28.3 ± 7.1 vs. 36.6 ± 6.5 (+29.3%) | heart rate (+0.7%) stroke volume (+14.8%) cardiac output (+20.8%) arteriovenous O2diff (+8.8%) |
| Murias et al., 2010 [31] | 8 m | 84 (re) | 23.0 ± 5.0 | 48.0 ± 6.1 vs. 55.4 ± 5.5% (+15.4%) | heart rate (−1.1%) stroke volume (+10.9%) cardiac output (+9.7%) arteriovenous O2diff (+6.8%) |
| Nichols et al., 2000 [12] | 1 f | 14 (re) | 49.4 | 42.0 vs. 48.1 (+14.5%) | heart rate (+1.6%) |
| Nichols et al., 2000 [12] | 1 f | 28 (re) | 49.4 | 42.0 vs. 55.1 (+31.2%) | heart rate (−2.7%) |
| Pavlik et al., 1986 [32] | 42 m | 60 (ce) | 22.9 ± 0.7 | 72.2 vs. 67.0 (30 days) vs. 62.5 (45 days) (−7% and −13%); no further decrease after 45 days | |
| Prior et al., 2015 [33] | 7 m 5 f | 14 (ce) | 65 ± 3 | 31.2 ± 2.3 vs. 29.3 ± 1.9 (−6%) | citrate synthase (−28.6%) |
| Ready and Quinney, 1982 [34] | 12 m | 63 (ce) | 25.0 ± 3.6 | 64.2 ± 9.5 vs. 59.3 ± 6.4, (3 weeks) 57.5 ± 6.4 (6 weeks) 57.3 ± 8.8 (9 weeks) (−10.7%) | |
| Schulman et al., 1996 [35] | 8 m | 84 (ce) | 59.6 ± 3 | 49.9 ± 1.9 vs. 42.0 ± 2.2 (−15.8%) | heart rate (+4.1%) cardiac index (−10.5%) stroke volume index (−14.2%) |
| Sinacore et al., 1993 [36] | 5 m 1 f | 84 (ce) | 29 ± 10 | 61.3 ± 7 vs. 50.8 ± 7 (−17.1%) | |
| Smorawinski et al., 2001 [37] | 10 m | 3 (ce, bedrest) | 20.3 ± 1.9 | 54.8 ± 2.1 (−16.5%) | blood lactate (−8% to −20%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Burtscher, J.; Strasser, B.; Burtscher, M.; Millet, G.P. The Impact of Training on the Loss of Cardiorespiratory Fitness in Aging Masters Endurance Athletes. Int. J. Environ. Res. Public Health 2022, 19, 11050. https://doi.org/10.3390/ijerph191711050
Burtscher J, Strasser B, Burtscher M, Millet GP. The Impact of Training on the Loss of Cardiorespiratory Fitness in Aging Masters Endurance Athletes. International Journal of Environmental Research and Public Health. 2022; 19(17):11050. https://doi.org/10.3390/ijerph191711050
Chicago/Turabian StyleBurtscher, Johannes, Barbara Strasser, Martin Burtscher, and Gregoire P. Millet. 2022. "The Impact of Training on the Loss of Cardiorespiratory Fitness in Aging Masters Endurance Athletes" International Journal of Environmental Research and Public Health 19, no. 17: 11050. https://doi.org/10.3390/ijerph191711050
APA StyleBurtscher, J., Strasser, B., Burtscher, M., & Millet, G. P. (2022). The Impact of Training on the Loss of Cardiorespiratory Fitness in Aging Masters Endurance Athletes. International Journal of Environmental Research and Public Health, 19(17), 11050. https://doi.org/10.3390/ijerph191711050