

Titanium Dioxide Nanoparticles Altered the lncRNA Expression Profile in Human Lung Cells



Abstract
:1. Introduction
2. Materials and Methods
2.1. Physicochemical Properties of TiO2 NPs and Cell Culture
2.2. Cell Experimental Design
2.3. Construction of cDNA Libraries and RNA Sequencing
2.4. Analysis of lncRNA
2.5. Statistical Analysis
3. Results
3.1. Characterization of the TiO2 NPs
3.2. Predictions and Annotations of lncRNAs
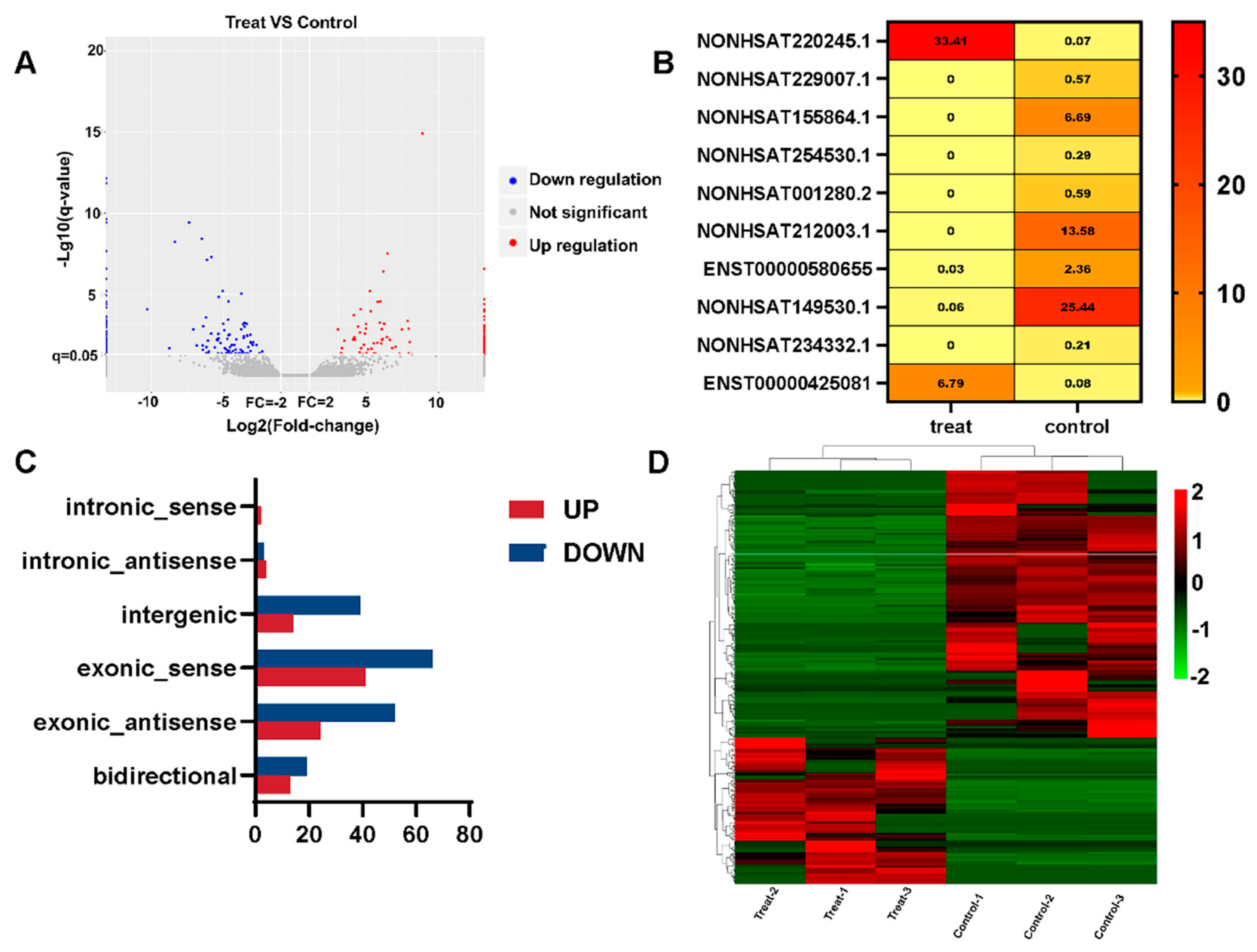
3.3. Differential Expression of lncRNAs
3.4. Enrichment Analysis of DE lncRNA Target Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhen, S.; Qian, Q.; Jia, G.; Zhang, J.; Chen, C.; Wei, Y. A panel study for cardiopulmonary effects produced by occupational exposure to inhalable titanium dioxide. J. Occup. Environ. Med. 2012, 54, 1389–1394. [Google Scholar] [CrossRef]
- Yamano, S.; Goto, Y.; Takeda, T.; Hirai, S.; Furukawa, Y.; Kikuchi, Y.; Kasai, T.; Misumi, K.; Suzuki, M.; Takanobu, K.; et al. Pulmonary dust foci as rat pneumoconiosis lesion induced by titanium dioxide nanoparticles in 13-week inhalation study. Part. Fibre Toxicol. 2022, 19, 58. [Google Scholar] [CrossRef]
- Gottschalk, F.; Nowack, B. The release of engineered nanomaterials to the environment. J. Environ. Monit. 2011, 13, 1145–1155. [Google Scholar] [CrossRef]
- Bressot, C.; Manier, N.; Pagnoux, C.; Aguerre-Chariol, O.; Morgeneyer, M. Environmental release of engineered nanomaterials from commercial tiles under standardized abrasion conditions. J. Hazard. Mater. 2017, 322, 276–283. [Google Scholar] [CrossRef]
- Al-Kattan, A.; Wichser, A.; Vonbank, R.; Brunner, S.; Ulrich, A.; Zuin, S.; Arroyo, Y.; Golanski, L.; Nowack, B. Characterization of materials released into water from paint containing nano-SiO2. Chemosphere 2015, 119, 1314–1321. [Google Scholar] [CrossRef] [PubMed]
- Shandilya, N.; Le Bihan, O.; Morgeneyer, M. Effect of the Normal Load on the Release of Aerosol Wear Particles During Abrasion. Tribology Letters 2014, 55, 227–234. [Google Scholar] [CrossRef]
- Kanduri, C. Long noncoding RNAs: Lessons from genomic imprinting. Biochim. Biophys. Acta 2016, 1859, 102–111. [Google Scholar] [CrossRef]
- Fatica, A.; Bozzoni, I. Long non-coding RNAs: New players in cell differentiation and development. Nat. Rev. Genet. 2014, 15, 7–21. [Google Scholar] [CrossRef]
- Moreira, L.; Costa, C.; Pires, J.; Teixeira, J.P.; Fraga, S. How can exposure to engineered nanomaterials influence our epigenetic code? A review of the mechanisms and molecular targets. Mutat. Res. Rev. Mutat. Res. 2021, 788, 108385. [Google Scholar] [CrossRef]
- Dempsey, J.L.; Cui, J.Y. Long Non-Coding RNAs: A Novel Paradigm for Toxicology. Toxicol. Sci. 2017, 155, 3–21. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Shi, J.; Han, S.; Zheng, P.; Chen, Z.; Jia, G. Titanium dioxide nanoparticles induced reactive oxygen species (ROS) related changes of metabolomics signatures in human normal bronchial epithelial (BEAS-2B) cells. Toxicol. Appl. Pharm. 2022, 444, 116020. [Google Scholar] [CrossRef]
- Jayaram, D.T.; Payne, C.K. Food-Grade TiO2 Particles Generate Intracellular Superoxide and Alter Epigenetic Modifiers in Human Lung Cells. Chem. Res. Toxicol. 2020, 33, 2872–2879. [Google Scholar] [CrossRef]
- Yamano, S.; Takeda, T.; Goto, Y.; Hirai, S.; Furukawa, Y.; Kikuchi, Y.; Kasai, T.; Misumi, K.; Suzuki, M.; Takanobu, K.; et al. No evidence for carcinogenicity of titanium dioxide nanoparticles in 26-week inhalation study in rasH2 mouse model. Sci. Rep. 2022, 12, 14969. [Google Scholar] [CrossRef]
- Leppänen, M.; Korpi, A.; Miettinen, M.; Leskinen, J.; Torvela, T.; Rossi, E.M.; Vanhala, E.; Wolff, H.; Alenius, H.; Kosma, V.M.; et al. Nanosized TiO₂ caused minor airflow limitation in the murine airways. Arch. Toxicol. 2011, 85, 827–839. [Google Scholar] [CrossRef]
- Chen, Z.; Shi, J.; Zhang, Y.; Han, S.; Zhang, J.; Jia, G. DNA Oxidative Damage as a Sensitive Genetic Endpoint to Detect the Genotoxicity Induced by Titanium Dioxide Nanoparticles. Nanomaterials 2022, 12, 2616. [Google Scholar] [CrossRef]
- Fresegna, A.M.; Ursini, C.L.; Ciervo, A.; Maiello, R.; Casciardi, S.; Iavicoli, S.; Cavallo, D. Assessment of the Influence of Crystalline Form on Cyto-Genotoxic and Inflammatory Effects Induced by TiO2 Nanoparticles on Human Bronchial and Alveolar Cells. Nanomaterials 2021, 11, 253. [Google Scholar] [CrossRef]
- Hou, J.; Wang, L.; Wang, C.; Zhang, S.; Liu, H.; Li, S.; Wang, X. Toxicity and mechanisms of action of titanium dioxide nanoparticles in living organisms. J. Environ. Sci. 2019, 75, 40–53. [Google Scholar] [CrossRef]
- Law, P.P.; Holland, M.L. DNA methylation at the crossroads of gene and environment interactions. Essays Biochem. 2019, 63, 717–726. [Google Scholar] [CrossRef]
- Bai, W.; Chen, Y.; Gao, A. Cross talk between poly(ADP-ribose) polymerase 1 methylation and oxidative stress involved in the toxic effect of anatase titanium dioxide nanoparticles. Int. J. Nanomed. 2015, 10, 5561–5569. [Google Scholar] [CrossRef] [Green Version]
- Biola-Clier, M.; Beal, D.; Caillat, S.; Libert, S.; Armand, L.; Herlin-Boime, N.; Sauvaigo, S.; Douki, T.; Carriere, M. Comparison of the DNA damage response in BEAS-2B and A549 cells exposed to titanium dioxide nanoparticles. Mutagenesis 2017, 32, 161–172. [Google Scholar] [CrossRef]
- Peschansky, V.J.; Wahlestedt, C. Non-coding RNAs as direct and indirect modulators of epigenetic regulation. Epigenetics 2014, 9, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Yang, P.K.; Kuroda, M.I. Noncoding RNAs and intranuclear positioning in monoallelic gene expression. Cell 2007, 128, 777–786. [Google Scholar] [CrossRef] [Green Version]
- Clemson, C.M.; McNeil, J.A.; Willard, H.F.; Lawrence, J.B. XIST RNA paints the inactive X chromosome at interphase: Evidence for a novel RNA involved in nuclear/chromosome structure. J. Cell Biol. 1996, 132, 259–275. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.; Cai, Y.; Kapranov, P.; Xu, D. Reverse-genetics studies of lncRNAs-what we have learnt and paths forward. Genome Biol. 2020, 21, 93. [Google Scholar] [CrossRef] [Green Version]
- Lv, J.; Qiu, M.; Xia, W.; Liu, C.; Xu, Y.; Wang, J.; Leng, X.; Huang, S.; Zhu, R.; Zhao, M.; et al. High expression of long non-coding RNA SBF2-AS1 promotes proliferation in non-small cell lung cancer. J. Exp. Clin. Cancer Res. 2016, 35, 75. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Sun, M.; Zang, C.; Ma, P.; He, J.; Zhang, M.; Huang, Z.; Ding, Y.; Shu, Y. Upregulated long non-coding RNA AGAP2-AS1 represses LATS2 and KLF2 expression through interacting with EZH2 and LSD1 in non-small-cell lung cancer cells. Cell Death Dis. 2016, 7, e2225. [Google Scholar] [CrossRef]
- Li, B.; Huang, N.; Wei, S.; Xv, J.; Meng, Q.; Aschner, M.; Li, X.; Chen, R. lncRNA TUG1 as a ceRNA promotes PM exposure-induced airway hyper-reactivity. J. Hazard. Mater. 2021, 416, 125878. [Google Scholar] [CrossRef]
- Konwar, C.; Asiimwe, R.; Inkster, A.M.; Merrill, S.M.; Negri, G.L.; Aristizabal, M.J.; Rider, C.F.; MacIsaac, J.L.; Carlsten, C.; Kobor, M.S. Risk-focused differences in molecular processes implicated in SARS-CoV-2 infection: Corollaries in DNA methylation and gene expression. Epigenetics Chromatin 2021, 14, 54. [Google Scholar] [CrossRef]
- Mao, W.; Wang, K.; Sun, S.; Wu, J.; Chen, M.; Geng, J.; Luo, M. ID2 Inhibits Bladder Cancer Progression and Metastasis via PI3K/AKT Signaling Pathway. Front. Cell Dev. Biol. 2021, 9, 738364. [Google Scholar] [CrossRef]
- Li, N.; Wu, L.; Zuo, X.; Luo, H.; Sheng, Y.; Yan, J. USP1 Promotes GC Metastasis via Stabilizing ID2. Dis. Markers 2021, 2021, 3771990. [Google Scholar] [CrossRef]
- Chen, J.T.; Hsu, Y.L.; Hsu, Y.C.; Tseng, Y.H.; Liu, M.H.; Weng, C.W.; Lin, C.H.; Pan, S.H.; Chen, J.J.W.; Wang, C.C. Id2 exerts tumor suppressor properties in lung cancer through its effects on cancer cell invasion and migration. Front. Oncol. 2022, 12, 801300. [Google Scholar] [CrossRef]
- Lim, J.Y.; Kim, S.W.; Kim, B.; Park, S.J. Knockdown of CARD14 Inhibits Cell Proliferation and Migration in Breast Cancer Cells. Anticancer Res. 2020, 40, 1953–1962. [Google Scholar] [CrossRef]
- Jiang, T.; Wang, H.; Liu, L.; Song, H.; Zhang, Y.; Wang, J.; Liu, L.; Xu, T.; Fan, R.; Xu, Y.; et al. CircIL4R activates the PI3K/AKT signaling pathway via the miR-761/TRIM29/PHLPP1 axis and promotes proliferation and metastasis in colorectal cancer. Mol. Cancer 2021, 20, 167. [Google Scholar] [CrossRef] [PubMed]
- Garavito, M.F.; Narváez-Ortiz, H.Y.; Zimmermann, B.H. Pyrimidine Metabolism: Dynamic and Versatile Pathways in Pathogens and Cellular Development. J. Genet. Genomics 2015, 42, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Poirier, S.; Samami, S.; Mamarbachi, M.; Demers, A.; Chang, T.Y.; Vance, D.E.; Hatch, G.M.; Mayer, G. The epigenetic drug 5-azacytidine interferes with cholesterol and lipid metabolism. J. Biol. Chem. 2014, 289, 18736–18751. [Google Scholar] [CrossRef] [Green Version]
- Lindahl, T. Instability and decay of the primary structure of DNA. Nature 1993, 362, 709–715. [Google Scholar] [CrossRef]
- Stasolla, C.; Loukanina, N.; Yeung, E.C.; Thorpe, T.A. Alterations in pyrimidine nucleotide metabolism as an early signal during the execution of programmed cell death in tobacco BY-2 cells. J. Exp. Bot. 2004, 55, 2513–2522. [Google Scholar] [CrossRef]
- Wang, K.; Wang, L.; Zhao, G.; Liu, Y.; Wang, F.; Song, H.; Sun, Y.; Zhou, Z.; Lu, X.; Hu, H.; et al. Mechanistic study of salidroside on ovalbumin-induced asthmatic model mice based on untargeted metabolomics analysis. Food Funct. 2023, 14, 413–426. [Google Scholar] [CrossRef]
- Peng, F.; Dai, J.; Qian, Q.; Cao, X.; Wang, L.; Zhu, M.; Han, S.; Liu, W.; Li, Y.; Xue, T.; et al. Serum metabolic profiling of coal worker’s pneumoconiosis using untargeted lipidomics. Environ. Sci. Pollut Res. Int. 2022, 29, 85444–85453. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Type | Primary Size (nm) | Hydrated Sizes in Ultrapure Water (nm) | Hydrated Sizes in DMEM (nm) | Zeta Potential in Ultrapure Water (mV) | Zeta Potential in DMEM (mV) |
|---|---|---|---|---|---|
| Anatase | 25.12 ± 5.64 | 609.43 ± 60.35 | 878.93 ± 105.75 | −8.33 ± 0.22 | −15.20 ± 0.92 |
| LncRNA_ID | Treat a | Control a | q-Value | Gene_ID | Gene_Name | Treat a | Control a | q-Value |
|---|---|---|---|---|---|---|---|---|
| NONHSAT248795.1 | 0.00 | 0.33 | 3.31 × 10−2 | ENSG00000218336 | TENM3 | 0.55 | 2.03 | 1.25 × 10−2 |
| NONHSAT226241.1 | 0.00 | 0.13 | 3.31 × 10−2 | ENSG00000187634 | SAMD11 | 0.03 | 1.05 | 1.59 × 10−5 |
| MSTRG.48669.1 | 0.37 | 1.81 | 3.31 × 10−2 | ENSG00000137573 | SULF1 | 0.19 | 0.98 | 2.29 × 10−3 |
| NONHSAT169524.1 | 2.89 | 0.04 | 1.45 × 10−3 | ENSG00000165555 | NOXRED1 | 0.07 | 0.22 | 3.31 × 10−2 |
| NONHSAT080219.2 | 0.04 | 2.50 | 8.61 × 10−3 | ENSG00000158445 | KCNB1 | 0.32 | 2.59 | 1.49 × 10−9 |
| NONHSAT193964.1 | 0.00 | 1.47 | 6.65 × 10−4 | ENSG00000241472 | PTPRG-AS1 | 0.11 | 0.30 | 1.08 × 10−5 |
| NONHSAT056352.2 | 0.35 | 0.04 | 4.25 × 10−2 | ENSG00000141527 | CARD14 | 0.02 | 0.05 | 1.18 × 10−4 |
| ENST00000315302 | 0.01 | 0.65 | 4.50 × 10−3 | ENSG00000218336 | TENM3 | 0.55 | 2.03 | 3.72 × 10−2 |
| ENST00000572730 | 0.00 | 1.07 | 1.11 × 10−6 | ENSG00000141527 | CARD14 | 0.02 | 0.05 | 5.62 × 10−20 |
| NONHSAT160825.1 | 0.13 | 1.90 | 2.26 × 10−2 | ENSG00000137699 | TRIM29 | 0.10 | 0.43 | 3.59 × 10−28 |
| NONHSAT226246.1 | 0.01 | 0.47 | 3.31 × 10−2 | ENSG00000187634 | SAMD11 | 0.03 | 1.05 | 1.43 × 10−15 |
| NONHSAT162857.1 | 0.00 | 0.22 | 5.65 × 10−3 | ENSG00000152137 | HSPB8 | 0.16 | 1.54 | 1.61 × 10−2 |
| NONHSAT257137.1 | 0.00 | 0.27 | 5.03 × 10−8 | ENSG00000204054 | LINC00963 | 0.68 | 1.93 | 4.77 × 10−22 |
| NONHSAT069007.2 | 0.13 | 0.00 | 1.00 × 10−2 | ENSG00000115738 | ID2 | 2.50 | 8.27 | 8.26 × 10−4 |
| NONHSAT248790.1 | 0.00 | 0.05 | 4.36 × 10−2 | ENSG00000218336 | TENM3 | 0.55 | 2.03 | 4.97 × 10−2 |
| MSTRG.48669.1 | 0.37 | 1.81 | 9.30 × 10−4 | ENSG00000006638 | TBXA2R | 0.19 | 0.06 | 5.62 × 10−20 |
| MSTRG.48669.1 | 0.37 | 1.81 | 2.52 × 10−4 | ENSG00000179841 | AKAP5 | 3.28 | 1.56 | 4.97 × 10−2 |
| MSTRG.48669.1 | 0.37 | 1.81 | 4.15 × 10−2 | ENSG00000169239 | CA5B | 1.76 | 0.85 | 4.50 × 10−10 |
| NONHSAT252820.1 | 0.77 | 0.10 | 6.29 × 10−6 | ENSG00000120278 | PLEKHG1 | 0.04 | 0.00 | 3.59 × 10−28 |
| NONHSAT176568.1 | 12.50 | 0.05 | 1.77 × 10−2 | ENSG00000177885 | GRB2 | 17.81 | 6.81 | 8.86 × 10−16 |
| NONHSAT209182.1 | 1.31 | 0.02 | 4.92 × 10−4 | ENSG00000244694 | PTCHD4 | 0.78 | 0.24 | 2.35 × 10−14 |
| ENST00000530595 | 0.00 | 0.26 | 7.37 × 10−3 | ENSG00000166793 | YPEL4 | 0.06 | 0.01 | 1.29 × 10−14 |
| NONHSAT213922.1 | 0.60 | 0.03 | 2.99 × 10−2 | ENSG00000234985 | AC074085.2 | 0.44 | 0.00 | 5.62 × 10−20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Y.; Shi, J.; Zhang, Y.; Chen, Z.; Jia, G. Titanium Dioxide Nanoparticles Altered the lncRNA Expression Profile in Human Lung Cells. Int. J. Environ. Res. Public Health 2023, 20, 1059. https://doi.org/10.3390/ijerph20021059
Ma Y, Shi J, Zhang Y, Chen Z, Jia G. Titanium Dioxide Nanoparticles Altered the lncRNA Expression Profile in Human Lung Cells. International Journal of Environmental Research and Public Health. 2023; 20(2):1059. https://doi.org/10.3390/ijerph20021059
Chicago/Turabian StyleMa, Ying, Jiaqi Shi, Yi Zhang, Zhangjian Chen, and Guang Jia. 2023. "Titanium Dioxide Nanoparticles Altered the lncRNA Expression Profile in Human Lung Cells" International Journal of Environmental Research and Public Health 20, no. 2: 1059. https://doi.org/10.3390/ijerph20021059
APA StyleMa, Y., Shi, J., Zhang, Y., Chen, Z., & Jia, G. (2023). Titanium Dioxide Nanoparticles Altered the lncRNA Expression Profile in Human Lung Cells. International Journal of Environmental Research and Public Health, 20(2), 1059. https://doi.org/10.3390/ijerph20021059