
Do Freshwater and Marine Bivalves Differ in Their Response to Wildfire Ash? Effects on the Antioxidant Defense System and Metal Body Burden

 ,
,  ,
,  ,
, 
 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Test Organisms
2.2. Preparation of Aqueous Ash Extracts (AEAs)
2.3. Ecotoxicological Tests
2.4. Biochemical Analyses
2.4.1. Catalase (CAT)
2.4.2. Glutathione S-Transferase (GST)
2.4.3. Glutathione Reductase (GR)
2.4.4. Glutathione Peroxidase (GPx)
2.4.5. Thiobarbituric Acid Reactive Substances (TBARS)
2.4.6. Total Protein Concentration
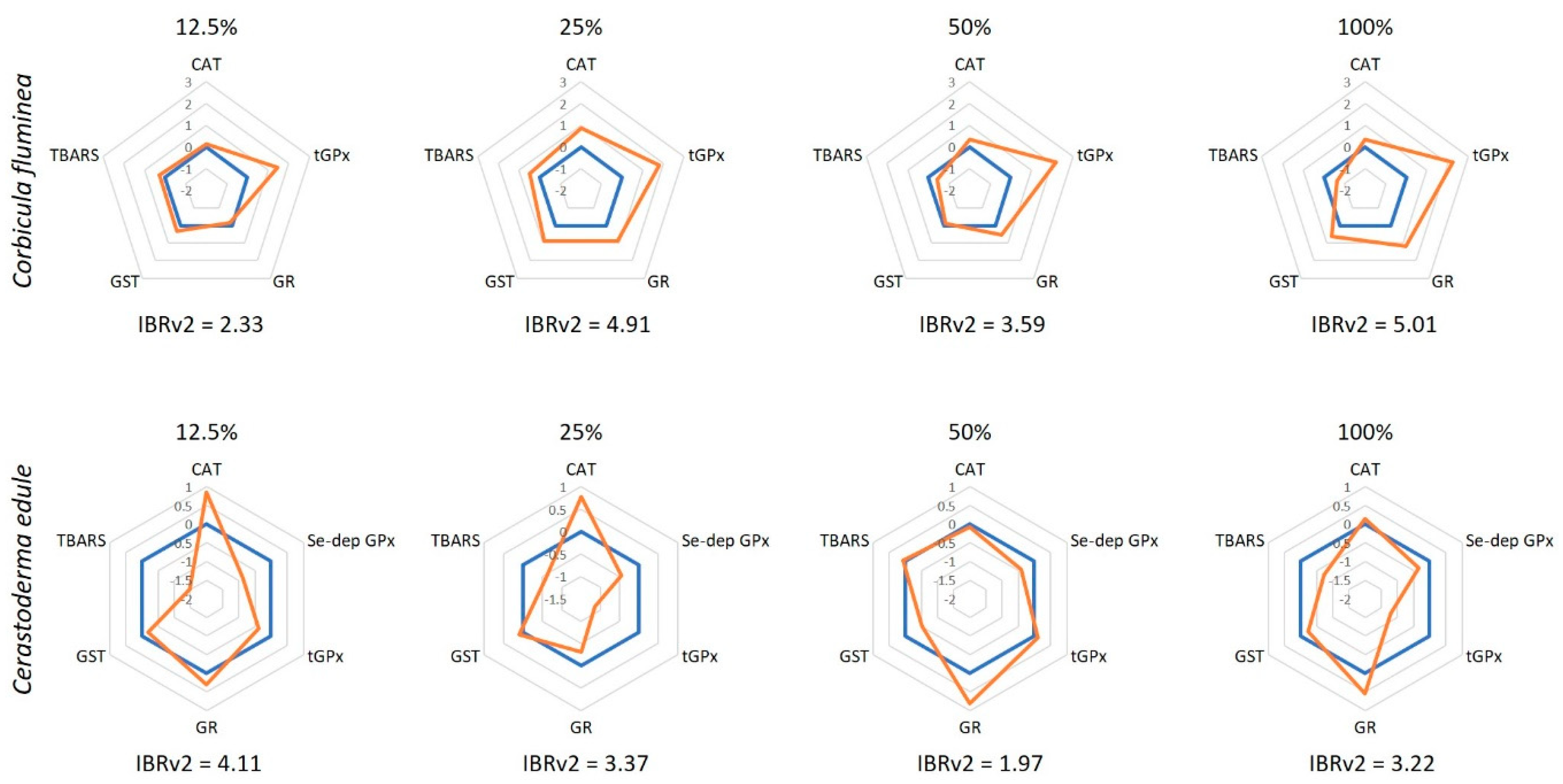
2.5. Determination of the IBRv2
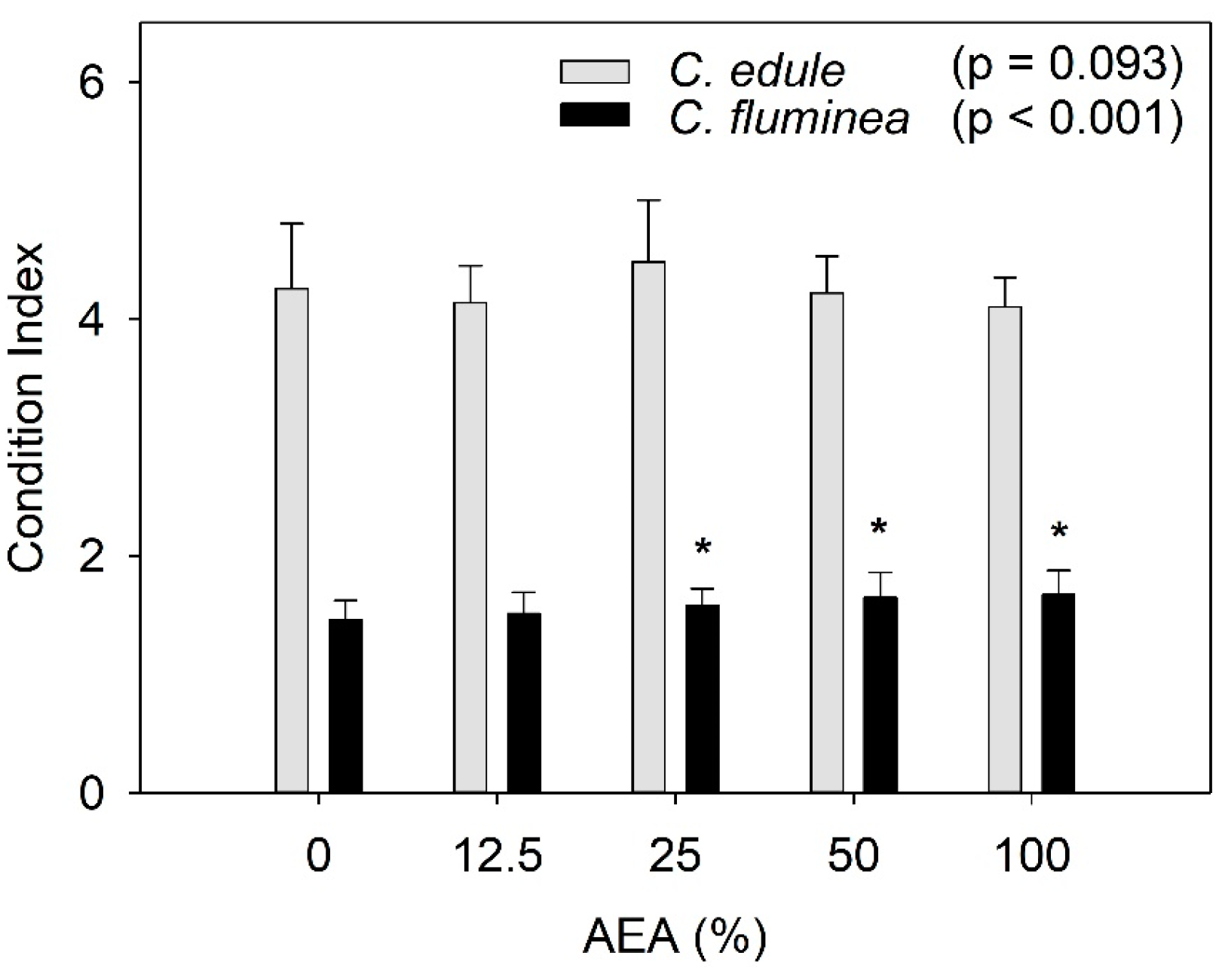
2.6. Condition Index
2.7. Chemical Analyses
2.8. Dietary Hazard Assessment
2.9. Data Treatment and Statistical Analyses
3. Results and Discussion
3.1. Chemical Analyses of the Exposure Media
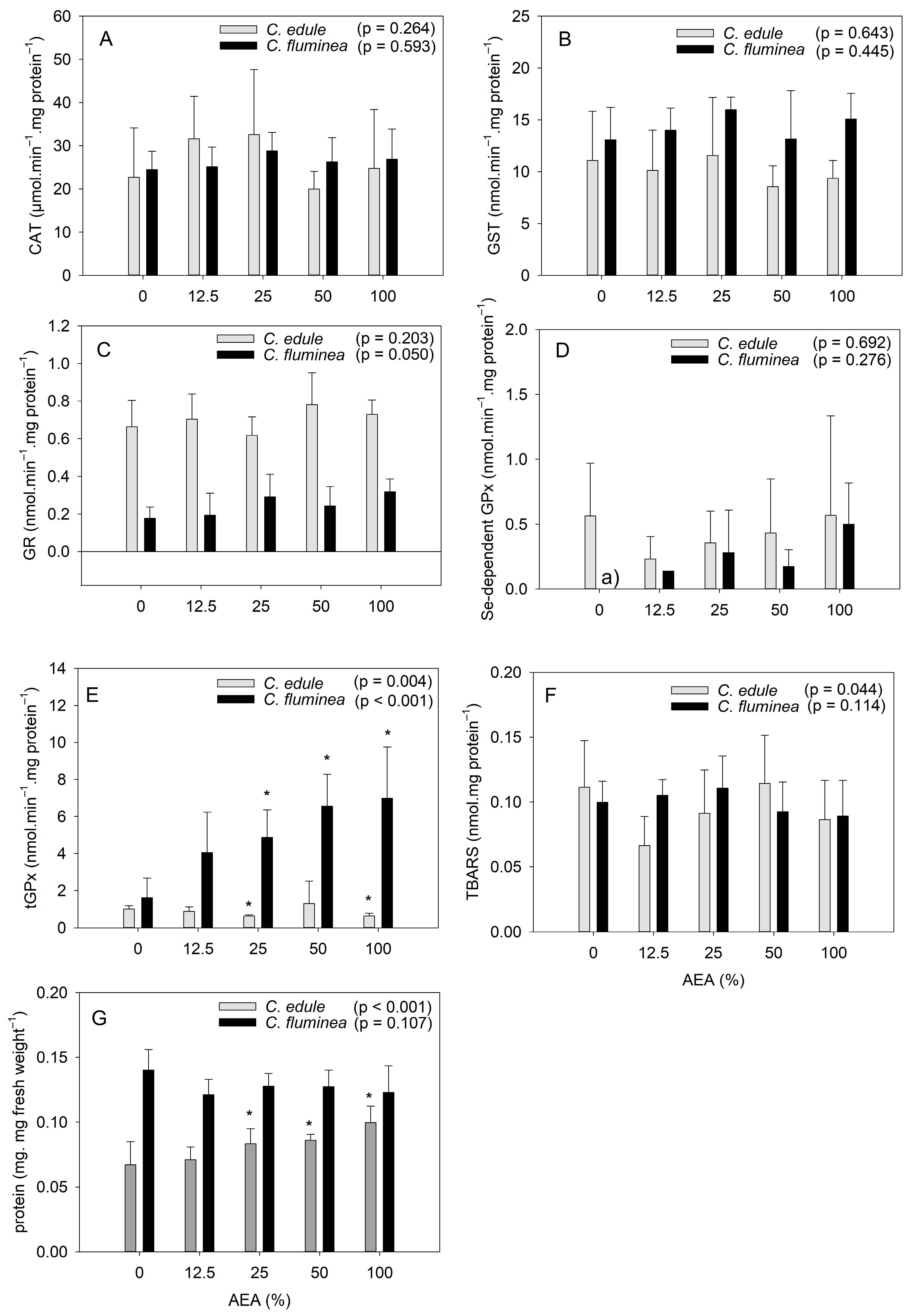
3.2. Biochemical Analyses
3.3. Condition Index (CI)
3.4. Metal Body Burden
3.5. Dietary Hazard Assessment
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Harper, A.R.; Santin, C.; Doerr, S.H.; Froyd, C.A.; Albini, D.; Otero, X.L.; Viñas, L.; Pérez-Fernández, B. Chemical composition of wildfire ash produced in contrasting ecosystems and its toxicity to Daphnia magna. Int. J. WildLand Fire 2019, 28, 726–737. [Google Scholar] [CrossRef] [Green Version]
- Isaza, D.F.G.; Cramp, R.L.; Franklin, C.E. Fire and rain: A systematic review of the impacts of wildfire and associated runoff on aquatic fauna. Glob. Chang. Biol. 2022, 28, 2578–2595. [Google Scholar] [CrossRef]
- Ré, A.; Campos, I.; Keizer, J.J.; Gonçalves, F.J.; Pereira, J.L.; Abrantes, N. Effects of post-fire contamination in sediment-dwelling species of riverine systems. Sci. Total Environ. 2021, 771, 144813. [Google Scholar] [CrossRef]
- Rinne, J.N. Management briefs: Short-term effects of wildfire on fishes and aquatic macroinvertebrates in the southwestern United States. N. Am. J. Fish. Manag. 1996, 16, 653–658. [Google Scholar] [CrossRef]
- Oliveira-Filho, E.C.; Brito, D.Q.; Dias, Z.M.; Guarieiro, M.S.; Carvalho, E.L.; Fascineli, M.L.; Niva, C.C.; Grisolia, C.K. Effects of ashes from a Brazilian savanna wildfire on water, soil and biota: An ecotoxicological approach. Sci. Total Environ. 2018, 618, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Ré, A.; Campos, I.; Saraiva, M.J.; Puga, J.; Keizer, J.J.; Gonçalves, F.J.; Pereira, J.L.; Abrantes, N. Wildfire effects on two freshwater producers: Combining in-situ and laboratory bioassays. Ecotox. Environ. Safe 2020, 194, 110361. [Google Scholar] [CrossRef]
- Campos, I.; Abrantes, N.; Vidal, T.; Bastos, A.C.; Gonçalves, F.J.M.; Keizer, J.J. Assessment of the toxicity of ash-loaded runoff from a recently burnt eucalypt plantation. Eur. J. For. Res. 2012, 131, 1889–1903. [Google Scholar] [CrossRef]
- Santos, R.; Fernandes, L.S.; Varandas, S.; Pereira, M.; Sousa, R.; Teixeira, A.; Lopes-Lima, M.; Cortes, R.; Pacheco, F. Impacts of climate change and land-use scenarios on Margaritifera margaritifera, an environmental indicator and endangered species. Sci. Total Environ. 2015, 511, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Brito, D.Q.; Passos, C.J.S.; Muniz, D.H.F.; Oliveira-Filho, E.C. Aquatic ecotoxicity of ashes from Brazilian savanna wildfires. Environ. Sci. Pollut. R. 2017, 24, 19671–19682. [Google Scholar] [CrossRef] [PubMed]
- Silva, V.; Abrantes, N.; Costa, R.; Keizer, J.J.; Gonçalves, F.; Pereira, J.L. Effects of ash-loaded post-fire runoff on the freshwater clam Corbicula fluminea. Ecol. Eng. 2016, 90, 180–189. [Google Scholar] [CrossRef]
- Ré, A.; Campos, I.; Puga, J.; Keizer, J.J.; Gonçalves, F.J.; Pereira, J.L.; Abrantes, N. Feeding inhibition following in-situ and laboratory exposure as an indicator of ecotoxic impacts of wildfires in affected waterbodies. Aquat. Toxicol. 2020, 227, 105587. [Google Scholar] [CrossRef] [PubMed]
- Bowen, L.; Miles, A.K.; Kolden, C.A.; Saarinen, J.A.; Bodkin, J.L.; Murray, M.J.; Tinker, M.T. Effects of wildfire on sea otter (Enhydra lutris) gene transcript profiles. Mar. Mammal. Sci. 2015, 31, 191–210. [Google Scholar] [CrossRef]
- Durán-Medraño, R.; Varela, E.; Garza-Gil, D.; Prada, A.; Vázquez, M.X.; Soliño, M. Valuation of terrestrial and marine biodiversity losses caused by forest wildfires. J. Behav. Exp. Econ. 2017, 71, 88–95. [Google Scholar] [CrossRef]
- Vernon, E.L.; Jha, A.N. Assessing relative sensitivity of marine and freshwater bivalves following exposure to copper: Application of classical and novel genotoxicological biomarkers. Mutat. Res./Genet. Toxicol. Environ. Mutagen. 2019, 842, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, A.; Carvalho, F.; Abrantes, N.; Campos, I.; Keizer, J.J.; Cássio, F.; Pascoal, C. Biochemical and functional responses of stream invertebrate shredders to post-wildfire contamination. Environ. Pollut. 2020, 267, 115433. [Google Scholar] [CrossRef] [PubMed]
- Plomp, R.D.; Klemish, J.L.; Pyle, G.G. The single and combined effects of wildfire runoff and sediment-bound copper on the freshwater amphipod Hyalella azteca. Environ. Toxicol. Chem. 2020, 39, 1988–1997. [Google Scholar] [CrossRef]
- Nunes, B.; Silva, V.; Campos, I.; Pereira, J.L.; Pereira, P.; Keizer, J.J.; Gonçalves, F.; Abrantes, N. Off-site impacts of wildfires on aquatic systems—Biomarker responses of the mosquitofish Gambusia holbrooki. Sci. Total Environ. 2017, 581, 305–313. [Google Scholar] [CrossRef]
- Ré, A.; Rocha, A.T.; Campos, I.; Marques, S.M.; Keizer, J.J.; Gonçalves, F.J.M.; Pereira, J.L.; Abrantes, N. Impacts of wildfires in aquatic organisms: Biomarker responses and erythrocyte nuclear abnormalities in Gambusia holbrooki exposed in situ. Environ. Sci. Pollut. R. 2021, 28, 51733–51744. [Google Scholar] [CrossRef]
- Santos, D.; Abrantes, N.; Campos, I.; Domingues, I.; Lopes, I. Effects of aqueous extracts of wildfire ashes on tadpoles of Pelophylax perezi: Influence of plant coverage. Sci. Total Environ. 2023, 854, 158746. [Google Scholar] [CrossRef]
- Li, D.; Wang, J.; Pi, J.; Yu, J.; Zhang, T. Biota-sediment metal accumulation and human health risk assessment of freshwater bivalve Corbicula fluminea in Dongting Lake, China. Environ. Sci. Pollut. Res. 2019, 26, 14951–14961. [Google Scholar] [CrossRef] [PubMed]
- Marques, A.; Piló, D.; Araújo, O.; Pereira, F.; Guilherme, S.; Carvalho, S.; Santos, M.A.; Pacheco, M.; Pereira, P. Propensity to metal accumulation and oxidative stress responses of two benthic species (Cerastoderma edule and Nephtys hombergii): Are tolerance processes limiting their responsiveness? Ecotoxicology 2016, 25, 664–676. [Google Scholar] [CrossRef] [Green Version]
- Rosa, I.C.; Costa, R.; Gonçalves, F.; Pereira, J. Bioremediation of metal-rich effluents: Could the invasive bivalve Corbicula fluminea work as a biofilter? J. Environ. Qual. 2014, 43, 1536–1545. [Google Scholar] [CrossRef]
- Rong, Y.; Tang, Y.; Ren, L.; Taylor, W.; Razlutskij, V.; Naselli-Flores, L.; Liu, Z.; Zhang, X. Effects of the filter-feeding benthic bivalve Corbicula fluminea on plankton community and water quality in aquatic ecosystems: A mesocosm study. Water 2021, 13, 1827. [Google Scholar] [CrossRef]
- Li, Z.; He, X.; Feng, C. A review of freshwater benthic clams (Corbicula fluminea): Accumulation capacity, underlying physiological mechanisms and environmental applications. Sci. Total Environ. 2023, 857, 159431. [Google Scholar] [CrossRef] [PubMed]
- Veiga, K.; Pedro, C.A.; Ferreira, S.M.F.; Gonçalves, S.C. Monitoring metal pollution on coastal lagoons using Cerastoderma edule—A report from a moderately impacted system in Western Portugal (Óbidos Lagoon). Environ. Sci. Pollut. Res. 2019, 26, 2710–2721. [Google Scholar] [CrossRef]
- Figueira, E.; Lima, A.; Branco, D.; Quintino, V.; Rodrigues, A.M.; Freitas, R. Health concerns of consuming cockles (Cerastoderma edule L.) from a low contaminated coastal system. Environ. Int. 2011, 37, 965–972. [Google Scholar] [PubMed]
- López-Pedrouso, M.; Lorenzo, J.M.; Varela, Z.; Fernández, J.Á.; Franco, D. Finding biomarkers in antioxidant molecular mechanisms for ensuring food safety of bivalves threatened by marine pollution. Antioxidants 2022, 11, 369. [Google Scholar] [CrossRef]
- Dupuy, J.-L.; Fargeon, H.; Martin-StPaul, N.; Pimont, F.; Ruffault, J.; Guijarro, M.; Hernando, C.; Madrigal, J.; Fernandes, P. Climate change impact on future wildfire danger and activity in southern Europe: A review. Ann. For. Sci. 2020, 77, 35. [Google Scholar] [CrossRef]
- Barros, T.L.; Bracewell, S.; Mayer-Pinto, M.; Dafforn, K.; Simpson, S.; Farrell, M.; Johnston, E. Wildfires cause rapid changes to estuarine benthic habitat. Environ. Pollut. 2022, 308, 119571. [Google Scholar] [CrossRef]
- Machado, A.; Serpa, D.; Santos, A.K.; Gomes, A.P.; Keizer, J.J.; Oliveira, B.R. Effects of different amendments on the quality of burnt eucalypt forest soils—A strategy for ecosystem rehabilitation. J. Environ. Manag. 2022, 320, 115766. [Google Scholar] [CrossRef]
- González, A.-B.M.; Campos, I.; Re, A.; Martínez-Guitarte, J.-L.; Abrantes, N. Effects of wildfire ashes on aquatic invertebrates: First molecular approach on Chironomus riparius Larvae. Sci. Total Environ. 2023, 858, 159899. [Google Scholar] [CrossRef]
- Domingues, A.; Rosa, I.C.; da Costa, J.P.; Rocha-Santos, T.A.; Gonçalves, F.J.; Pereira, R.; Pereira, J.L. Potential of the bivalve Corbicula fluminea for the remediation of olive oil wastewaters. J. Clean. Prod. 2020, 252, 119773. [Google Scholar] [CrossRef]
- Mesquita, A.F.; Marques, S.M.; Marques, J.C.; Gonçalves, F.J.M.; Gonçalves, A.M.M. Copper sulphate impact on the antioxidant defence system of the marine bivalves Cerastoderma edule and Scrobicularia plana. Sci. Rep. 2019, 9, 16458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, X.; Feng, C. Biological toxicity response of Asian Clam (Corbicula fluminea) to pollutants in surface water and sediment. Sci. Total Environ. 2018, 631–632, 56–70. [Google Scholar] [CrossRef] [PubMed]
- Claiborne, A. Catalase activity. In Handbook of Methods in Oxygen Radical Research; Ra, G., Ed.; CRC Press Inc: Boca Raton, FL, USA, 1985; pp. 283–284. [Google Scholar]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases: The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Carlberg, I.; Mannervik, B. [59] Glutathione reductase. In Methods in Enzymology; Academic Press Inc.: London, UK, 1985; pp. 484–490. [Google Scholar]
- Flohé, L.; Günzler, W.A. [12] Assays of glutathione peroxidase. In Methods in Enzymology; Academic Press Inc.: London, UK, 1984; pp. 114–120. [Google Scholar]
- Buege, J.A.; Aust, S.D. [30] Microsomal lipid peroxidation. In Methods in Enzymology; Academic Press Inc.: London, UK, 1978; pp. 302–310. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Sanchez, W.; Burgeot, T.; Porcher, J. A novel “Integrated Biomarker Response” calculation based on reference deviation concept. Environ. Sci. Pollut. R. 2013, 20, 2721–2725. [Google Scholar] [CrossRef]
- Cataldo, D.H.; Boltovskoy, D.; Stripeikis, J.; Pose, M. Condition index and growth rates of field caged Corbicula fluminea (Bivalvia) as biomarkers of pollution gradients in the Paraná river delta (Argentina). Aquat. Ecosyst. Health Manag. Sci. 2001, 4, 187–201. [Google Scholar] [CrossRef]
- Kagley, A.N.; Snider, R.G.; Krishnakumar, P.K.; Casillas, E. Assessment of seasonal variability of cytochemical responses to contaminant exposure in the blue mussel Mytilus edulis (Complex). Arch. Environ. Contam. Toxicol. 2003, 44, 0043–0052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rak, A.E.; Nasir, S.N.A.M.; Nor, M.M.; Han, D.K.; Appalasamy, S.; Abdullah, F.; Ghazi, R.M. Proximate analysis and fatty acid of Corbicula fluminea (C. fluminea) tissue in Kelantan, Malaysia. Environ. Sci. Pollut. Res. 2020, 27, 24772–24785. [Google Scholar] [CrossRef]
- JECFA; Food, Agriculture Organization; World Health Organization. Safety evaluation of certain food additives and contaminants/prepared by the 61st meeting of the Joint FAO/WHO Expert Committee on Food Additives (JEFCA). In WHO Food Additives Series Geneva: International Programme on Chemical Safety; World Health Organization: Geneva, Switzerland, 2004. [Google Scholar]
- Coelho, L.; Afonso, M.; Jesus, F.; Campos, I.; Abrantes, N.; Gonçalves, F.J.M.; Serpa, D.; Marques, S.M. Effects of Eucalypt ashes from moderate and high severity wildfires on the skin microbiome of the Iberian frog (Rana iberica). Environ. Pollut. 2022, 313, 120065. [Google Scholar] [CrossRef] [PubMed]
- Afonso, M.; Coelho, L.; Jesus, F.; Campos, I.; Abrantes, N.; Gonçalves, F.J.; Marques, S.; Serpa, D. Effects of Pine and Eucalypt ashes on bacterial isolates from the skin microbiome of the fire salamander (Salamandra salamandra). Sci. Total Environ. 2022, 841, 156677. [Google Scholar] [CrossRef] [PubMed]
- Silva, V.; Pereira, J.L.; Campos, I.; Keizer, J.J.; Gonçalves, F.; Abrantes, N. Toxicity assessment of aqueous extracts of ash from forest fires. Catena 2015, 135, 401–408. [Google Scholar] [CrossRef]
- Ré, A.; Rocha, A.T.; Campos, I.; Keizer, J.J.; Gonçalves, F.J.M.; Oliveira, H.; Pereira, J.L.; Abrantes, N. Cytotoxic effects of wildfire ashes: In-vitro responses of skin cells. Environ. Pollut. 2021, 285, 117279. [Google Scholar] [CrossRef]
- Vranković, J. Environmental impact on the antioxidant responses in Corbicula fluminea (Bivalvia: Veneroida: Corbiculidae) from the Danube River. Ital. J. Zool. 2015, 82, 378–386. [Google Scholar] [CrossRef]
- Bonnail, E.; Cunha Lima, R.; Bautista-Chamizo, E.; Salamanca, M.J.; Cruz-Hernández, P. Biomarker responses of the freshwater clam Corbicula fluminea in acid mine drainage polluted systems. Environ. Pollut. 2018, 242, 1659–1668. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, L.F.; Silva, S.M.C.P.; Martinez, C.B.R. Assessment of domestic landfill leachate toxicity to the Asian clam Corbicula fluminea via biomarkers. Ecotox. Environ. Safe 2014, 103, 17–23. [Google Scholar] [CrossRef]
- Freitas, R.; Costa, E.; Velez, C.; Santos, J.; Lima, A.; Oliveira, C.; Maria Rodrigues, A.; Quintino, V.; Figueira, E. Looking for suitable biomarkers in benthic macroinvertebrates inhabiting coastal areas with low metal contamination: Comparison between the bivalve Cerastoderma edule and the Polychaete Diopatra neapolitana. Ecotox. Environ. Safe 2012, 75, 109–118. [Google Scholar] [CrossRef]
- Regoli, F.; Giuliani, M.E. Oxidative pathways of chemical toxicity and oxidative stress biomarkers in marine organisms. Mar. Environ. Res. 2014, 93, 106–117. [Google Scholar] [CrossRef]
- Dallarés, S.; Carrasco, N.; Álvarez-Muñoz, D.; Rambla-Alegre, M.; Solé, M. Multibiomarker biomonitoring approach using three bivalve species in the Ebro Delta (Catalonia, Spain). Environ. Sci. Pollut. R. 2018, 25, 36745–36758. [Google Scholar] [CrossRef]
- Wheeler, J.R.; Leung, K.M.Y.; Morritt, D.; Sorokin, N.; Rogers, H.; Toy, R.; Holt, M.; Whitehouse, P.; Crane, M. Freshwater to saltwater toxicity extrapolation using species sensitivity distributions. Environ. Toxicol. Chem. 2002, 21, 2459–2467. [Google Scholar] [CrossRef]
- Markich, S.J. Sensitivity of the glochidia (larvae) of freshwater mussels (Bivalvia: Unionida: Hyriidae) to cadmium, cobalt, copper, lead, nickel and zinc: Differences between metals, species and exposure time. Sci. Total Environ. 2017, 601–602, 1427–1436. [Google Scholar] [CrossRef]
- Liu, S.; Zhao, H.; Zheng, M.; Wang, H.; Jing, C.; Zhang, W.; Hu, F. The physiological, biochemical and transcriptional responses to sulfamethoxazole in the Asian clam, Corbicula fluminea (O. F. Müller, 1774). Comparat. Biochem. Physiol. Part C: Toxicol. Pharmacol. 2022, 260, 109406. [Google Scholar]
- Bonnail, E.; Riba, I.; de Seabra, A.A.; DelValls, T.Á. Sediment quality assessment in the Guadalquivir River (SW, Spain) using caged Asian clams: A biomarker field approach. Sci. Total Environ. 2019, 650, 1996–2003. [Google Scholar] [CrossRef] [PubMed]
- Paskerová, H.; Hilscherová, K.; Bláha, L. Oxidative stress and detoxification biomarker responses in aquatic freshwater vertebrates exposed to microcystins and cyanobacterial biomass. Environ. Sci. Pollut. R. 2012, 19, 2024–2037. [Google Scholar] [CrossRef] [PubMed]
- Peltier, G.L.; Wright, M.S.; Hopkins, W.A.; Meyer, J.L. Accumulation of trace elements and growth responses in Corbicula fluminea downstream of a coal-fired power plant. Ecotox. Environ. Safe 2009, 72, 1384–1391. [Google Scholar] [CrossRef] [PubMed]
- Kesler, D.H. Influence of a lentic area on condition indices of Corbicula fluminea in the Wolf River, Tennessee. J. FreshWater Ecol. 2004, 19, 445–453. [Google Scholar] [CrossRef]
- Bodí, M.B.; Martin, D.A.; Balfour, V.N.; Santín, C.; Doerr, S.H.; Pereira, P.; Cerdà, A.; Mataix-Solera, J. Wildland fire ash: Production, composition and eco-hydro-geomorphic effects. Earth-Sci. Rev. 2014, 130, 103–127. [Google Scholar] [CrossRef]
- Lobo, J.; Costa, P.M.; Caeiro, S.; Martins, M.; Ferreira, A.M.; Caetano, M.; Cesário, R.; Vale, C.; Costa, M.H. Evaluation of the potential of the common cockle (Cerastoderma edule L.) for the ecological risk assessment of estuarine sediments: Bioaccumulation and biomarkers. Ecotoxicology 2010, 19, 1496–1512. [Google Scholar]
- Freitas, R.; Pires, A.; Quintino, V.; Rodrigues, A.M.; Figueira, E. Subcellular partitioning of elements and availability for trophic transfer: Comparison between the Bivalve Cerastoderma edule and the Polychaete Diopatra neapolitana. Estuar. Coast. Shelf Sci. 2012, 99, 21–30. [Google Scholar] [CrossRef]
- Saavedra, Y.; González, A.; Fernández, A.; Blanco, J. Interspecific variation of metal concentrations in three bivalve Mollusks from Galicia. Arch. Environ. Contam. Toxicol. 2004, 47, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Anajjar, E.M.; Chiffoleau, J.-F.; Bergayou, H.; Moukrim, A.; Burgeot, T.; Cheggour, M. Monitoring of trace metal contamination in the Souss Estuary (South Morocco) using the clams Cerastoderma edule and Scrobicularia plana. B. Environ. Contam. Toxicol. 2008, 80, 283–288. [Google Scholar] [CrossRef]
- Commission Regulation (EC) No. 1881/2006 of 19 December 2006; Setting Maximum Levels for Certain Contaminants in Foodstuffs (Text with EEA Relevance). Official Journal of the European Union: Luxembourg, 2006; pp. L364/5–L364/24.
- Vannoort, R.; Thomson, B. 2003/04 New Zealand Total Diet Survey. Agricultural Compound Residues, Selected Contaminants and Nutrients; New Zealand Food Safety Authority: Wellington, New Zealand, 2005. [Google Scholar]
- Jia, Y.; Wang, L.; Qu, Z.; Yang, Z. Distribution, contamination and accumulation of heavy metals in water, sediments, and freshwater shellfish from Liuyang River, Southern China. Environ. Sci. Pollut. R. 2018, 25, 7012–7020. [Google Scholar] [CrossRef]
- Shoults-Wilson, W.A.; Peterson, J.T.; Unrine, J.M.; Rickard, J.; Black, M.C. The Asian clam Corbicula fluminea as a biomonitor of trace element contamination: Accounting for different sources of variation using an hierarchical linear model. Environ. Toxicol. Chem. Int. J. 2009, 28, 2224–2232. [Google Scholar] [CrossRef]
- Gauthier, P.T.; Norwood, W.P.; Prepas, E.E.; Pyle, G.G. Metal–PAH mixtures in the aquatic environment: A review of co-toxic mechanisms leading to more-than-additive outcomes. Aquat. Toxicol. 2014, 154, 253–269. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.; Stelzenmueller, V.; Zauke, G.-P. Spatial distribution of heavy metal concentrations and biomass indices in Cerastoderma edule Linnaeus (1758) from the German Wadden Sea: An integrated biomonitoring approach. J. Exp. Mar. Biol. Ecol. 2006, 338, 81–95. [Google Scholar] [CrossRef]
- Baudrimont, M.; Schäfer, J.; Marie, V.; Maury-Brachet, R.; Bossy, C.; Boudou, A.; Blanc, G. Geochemical survey and metal bioaccumulation of three bivalve species (Crassostrea gigas, Cerastoderma edule and Ruditapes philippinarum) in the Nord Médoc salt marshes (Gironde estuary, France). Sci. Total Environ. 2005, 337, 265–280. [Google Scholar] [CrossRef]
- Bonnail, E.; Macías, F.; Osta, V. Ecological improvement assessment of a passive remediation technology for acid mine drainage: Water quality biomonitoring using bivalves. Chemosphere 2019, 219, 695–703. [Google Scholar] [CrossRef]
- Cheggour, M.; Chafik, A.; Langston, W.J.; Burt, G.R.; Benbrahim, S.; Texier, H. Metals in sediments and the edible cockle Cerastoderma edule from two Moroccan Atlantic lagoons: Moulay Bou Selham and Sidi Moussa. Environ. Pollut. 2001, 115, 149–7525. [Google Scholar] [CrossRef]
- Di Bella, G.; Turco, V.L.; Potortì, A.G.; Rando, R.; Licata, P.; Dugo, G. Statistical analysis of heavy metals in Cerastoderma edule glaucum and Venerupis aurea laeta from Ganzirri Lake, Messina (Italy). Environ. Monit. Assess. 2013, 185, 7517–7525. [Google Scholar] [CrossRef]
- Reis, P.A.; Guilhermino, L.; Antunes, C.; Sousa, R.G. Assessment of the ecological quality of the Minho estuary (Northwest Iberian Peninsula) based on metal concentrations in sediments and in Corbicula fluminea. Limnetica 2014, 33, 161–174. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| V | Cr | Mn | Fe | Co | Ni | Cu | Zn | As | Cd | Pb | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AEA in freshwater (%) | new medium | QL (µg L−1) | 1 | 1 | 0.3 | 10 | 0.1 | 1 | 2 | 2 | 2 | 0.1 | 0.1 |
| 0% | BQL | BQL | BQL | BQL | BQL | BQL | BQL | 3.03 (0.59) | BQL | BQL | BQL | ||
| 12.5% | 1.17 (0.058) | 1.2 (0.10) | 0.33 | BQL | 0.10 | BQL | 4.5 | 5.4 (4.67) | BQL | BQL | BQL | ||
| 25% | 1.37 (0.058) | 1.3 (0.00) | BQL | BQL | 0.14 (0.006) | BQL | 2.3 | 2.4 | BQL | BQL | BQL | ||
| 50% | 2.03 (0.55) | 1.83 (0.38) | 0.43 | BQL | 0.22 (0.042) | 2.15 (1.34) | 3.7 (0.62) | 2.5 | BQL | BQL | BQL | ||
| 100% | 2.93 (0.67) | 2.27 (0.64) | 2.93 (2.51) | 10.5 | 0.33 (0.046) | 2.67 (1.85) | 4.87 (0.76) | BQL | BQL | BQL | BQL | ||
| 48 h old medium | 0% | BQL | BQL | BQL | BQL | 0.17 (0.03) | BQL | BQL | 13.7 (0.58) | BQL | BQL | BQL | |
| 12.5% | BQL | BQL | BQL | BQL | 0.18 (0.055) | BQL | BQL | 9.6 (3.81) | BQL | BQL | BQL | ||
| 25% | BQL | BQL | BQL | BQL | 0.18 (0.035) | BQL | BQL | 5.93 (2.56) | BQL | BQL | BQL | ||
| 50% | 1.3 (0.14) | 2.00 (0.61) | BQL | BQL | 0.31 (0.067) | BQL | 2.73 (0.15) | 39.5 (9.19) | BQL | BQL | BQL | ||
| 100% | 2.55 (0.21) | 2.87 (1.45) | 0.33 | BQL | 0.49 (0.25) | BQL | 7.23 (1.14) | 79.5 (19.09) | BQL | BQL | BQL | ||
| AEA in brackish water (%) | new medium | QL (µg L−1) | 40 | 20 | 4 | 150 | 2 | 50 | 40 | 40 | 50 | 2 | 2 |
| 0% | BQL | BQL | BQL | BQL | BQL | BQL | BQL | BQL | BQL | BQL | BQL | ||
| 12.5% | BQL | BQL | 6.0 (2.65) | BQL | BQL | BQL | BQL | BQL | BQL | BQL | BQL | ||
| 25% | BQL | BQL | 7.0 (1.0) | BQL | BQL | BQL | BQL | BQL | BQL | BQL | BQL | ||
| 50% | BQL | BQL | 5.5 (0.71) | BQL | 2.0 | BQL | BQL | BQL | BQL | BQL | BQL | ||
| 100% | BQL | BQL | 12.7 (4.16) | BQL | 2.0 (0.00) | BQL | BQL | BQL | BQL | BQL | BQL | ||
| 48 h old medium | 0% | BQL | BQL | BQL | BQL | 2.0 | BQL | BQL | BQL | BQL | BQL | BQL | |
| 12.5% | BQL | BQL | BQL | BQL | BQL | BQL | BQL | BQL | BQL | BQL | BQL | ||
| 25% | BQL | BQL | BQL | BQL | BQL | BQL | BQL | BQL | BQL | BQL | BQL | ||
| 50% | BQL | BQL | BQL | BQL | BQL | BQL | BQL | BQL | BQL | BQL | BQL | ||
| 100% | BQL | BQL | BQL | BQL | BQL | BQL | BQL | BQL | BQL | BQL | BQL |
| V | Cr | Mn | Fe | Co | Ni | Cu | Zn | As | Cd | Pb | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Corbicula fluminea | field | 5.08 (1.41) | 132.05 (18.88) | 0.13 (0.02) | 12.07 (1.35) | 37.75 (14.74) | <0.03 a | <0.03 a | |||||
| t = 0 h | 3.43 (2.32) | 51.19 (11.69) | 0.11 (0.03) | 9.44 (1.78) | 14.53 (1.37) | 0.04 (0.01) | 0.13 (0.16) | ||||||
| t = 96 h | 0% | 0.67 (0.03) | 2.31 (0.54) | 49.46 (3.62) | 0.08 (0.008) | 7.28 (1.08) | 14.4 (0.34) | <0.27 a | <0.27 a | ||||
| 12.5% | 0.75 (0.017) | 2.26 (0.60) | 42.66 (3.86) | 0.07 (0.018) | 5.22 (0.29) * | 12.12 (1.14) | 0.04 (0.009) | 0.03 (0.001) | |||||
| 25% | 2.3 (0.56) | 40.33 (2.78) * | 0.1 (0.005) | 5.48 (0.63) * | 12.57 (1.57) | 0.04 (0.004) | 0.04 (0.008) | ||||||
| 50% | 1.59 (0.35) | 38.67 (6.52) * | 0.1 (0.027) | 5.27 (0.75) * | 12.71 (0.98) | 0.05 (0.02) | 0.03 (0.004) | ||||||
| 100% | 1.36 (0.23) * | 33.98 (1.72) * | 0.09 (0.020) | 5.12 (0.86) * | 10.81 (1.86) * | 0.03 (0.005) | 0.04 (0.025) | ||||||
| Cerastoderma edule | field | 0.13 (0.02) | 0.08 (0.02) | 0.74 (0.17) | 50.95 (19.71) | 0.11 (0.02) | 1.65 (0.07) | 0.49 (0.09) | 5.74 (0.51) | 1.51 (0.17) | 0.03 (0.02) | 0.06 (0.03) | |
| t = 0 h | < 0.08 a | 0.61 (0.16) | 35.08 (3.91) | 0.14 (0.05) | 1.93 (0.8) | 0.56 (0.05) | 7.36 (0.89) | 1.76 (0.08) | 0.03 (0.01) | 0.04 (0.01) | |||
| t = 96 h | 0% | < 0.07 a | 0.46 (0.15) | 35.54 (4.16) | 0.16 (0.022) | 2.00 (0.062) | 0.53 (0.037) | 8.12 (0.99) | 1.71 (0.07) | 0.04 (0.007) | 0.03 (0.006) | ||
| 12.5% | < 0.07 a | 0.56 (0.11) | 35.76 (6.17) | 0.13 (0.022) | 2.16 (0.43) | 0.56 (0.053) | 8.09 (0.99) | 1.63 (0.04) | 0.04 (0.015) | 0.04 (0.012) | |||
| 25% | 0.07 (0.01) | 0.52 (0.10) | 34.75 (4.23) | 0.14 (0.026) | 1.83 (0.46) | 0.55 (0.042) | 7.04 (0.89) | 1.70 (0.12) | 0.04 (0.013) | 0.02 (0.005) | |||
| 50% | 0.08 (0.01) | 0.59 (0.10) | 37.16 (4.75) | 0.12 (0.035) | 2.03 (0.44) | 0.61 (0.072) | 7.29 (0.23) | 1.82 (0.088) | 0.03 (0.007) | 0.03 (0.005) | |||
| 100% | 0.08 (0.01) | 0.72 (0.15) | 39.22 (9.38) | 0.16 (0.073) | 2.63 (0.80) | 0.58 (0.061) | 7.61 (0.94) | 1.71 (0.098) | 0.04 (0.01) | 0.03 (0.005) | |||
| ML | EFSA | - | - | - | - | - | - | - | - | - | 1.0 | 1.5 | |
| FSANZ | - | - | - | - | - | - | - | - | 1 | 2 | 2 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jesus, F.; Mesquita, F.; Virumbrales Aldama, E.; Marques, A.; Gonçalves, A.M.M.; Magalhães, L.; Nogueira, A.J.A.; Ré, A.; Campos, I.; Pereira, J.L.; et al. Do Freshwater and Marine Bivalves Differ in Their Response to Wildfire Ash? Effects on the Antioxidant Defense System and Metal Body Burden. Int. J. Environ. Res. Public Health 2023, 20, 1326. https://doi.org/10.3390/ijerph20021326
Jesus F, Mesquita F, Virumbrales Aldama E, Marques A, Gonçalves AMM, Magalhães L, Nogueira AJA, Ré A, Campos I, Pereira JL, et al. Do Freshwater and Marine Bivalves Differ in Their Response to Wildfire Ash? Effects on the Antioxidant Defense System and Metal Body Burden. International Journal of Environmental Research and Public Health. 2023; 20(2):1326. https://doi.org/10.3390/ijerph20021326
Chicago/Turabian StyleJesus, Fátima, Filipa Mesquita, Elisa Virumbrales Aldama, Ana Marques, Ana M. M. Gonçalves, Luísa Magalhães, António J. A. Nogueira, Ana Ré, Isabel Campos, Joana Luísa Pereira, and et al. 2023. "Do Freshwater and Marine Bivalves Differ in Their Response to Wildfire Ash? Effects on the Antioxidant Defense System and Metal Body Burden" International Journal of Environmental Research and Public Health 20, no. 2: 1326. https://doi.org/10.3390/ijerph20021326
APA StyleJesus, F., Mesquita, F., Virumbrales Aldama, E., Marques, A., Gonçalves, A. M. M., Magalhães, L., Nogueira, A. J. A., Ré, A., Campos, I., Pereira, J. L., Gonçalves, F. J. M., Abrantes, N., & Serpa, D. (2023). Do Freshwater and Marine Bivalves Differ in Their Response to Wildfire Ash? Effects on the Antioxidant Defense System and Metal Body Burden. International Journal of Environmental Research and Public Health, 20(2), 1326. https://doi.org/10.3390/ijerph20021326