

The Impact of Harsh Stratospheric Conditions on Survival and Antibiotic Resistance Profile of Non-Spore Forming Multidrug Resistant Human Pathogenic Bacteria Causing Hospital-Associated Infections

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Cultures
2.2. Sample Preparation and Pre-Flight Analyses
- Experimental Design
- Bacterial Culture
- Viable Count Assay
- Antibiotic Resistance Determination
2.3. Post–Flight Analyses
- Bacterial Culture
- Viable Count Assay
- Antibiotic Resistance Determination
- PCR Analyses
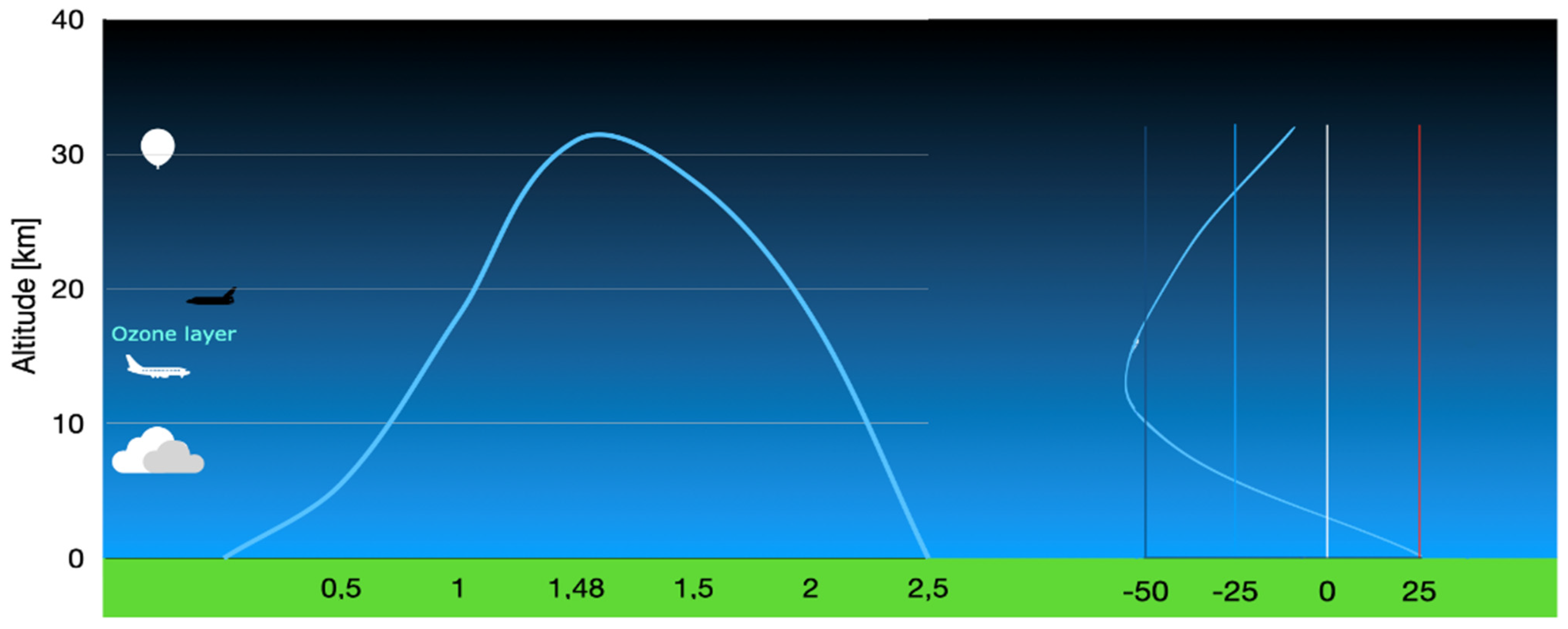
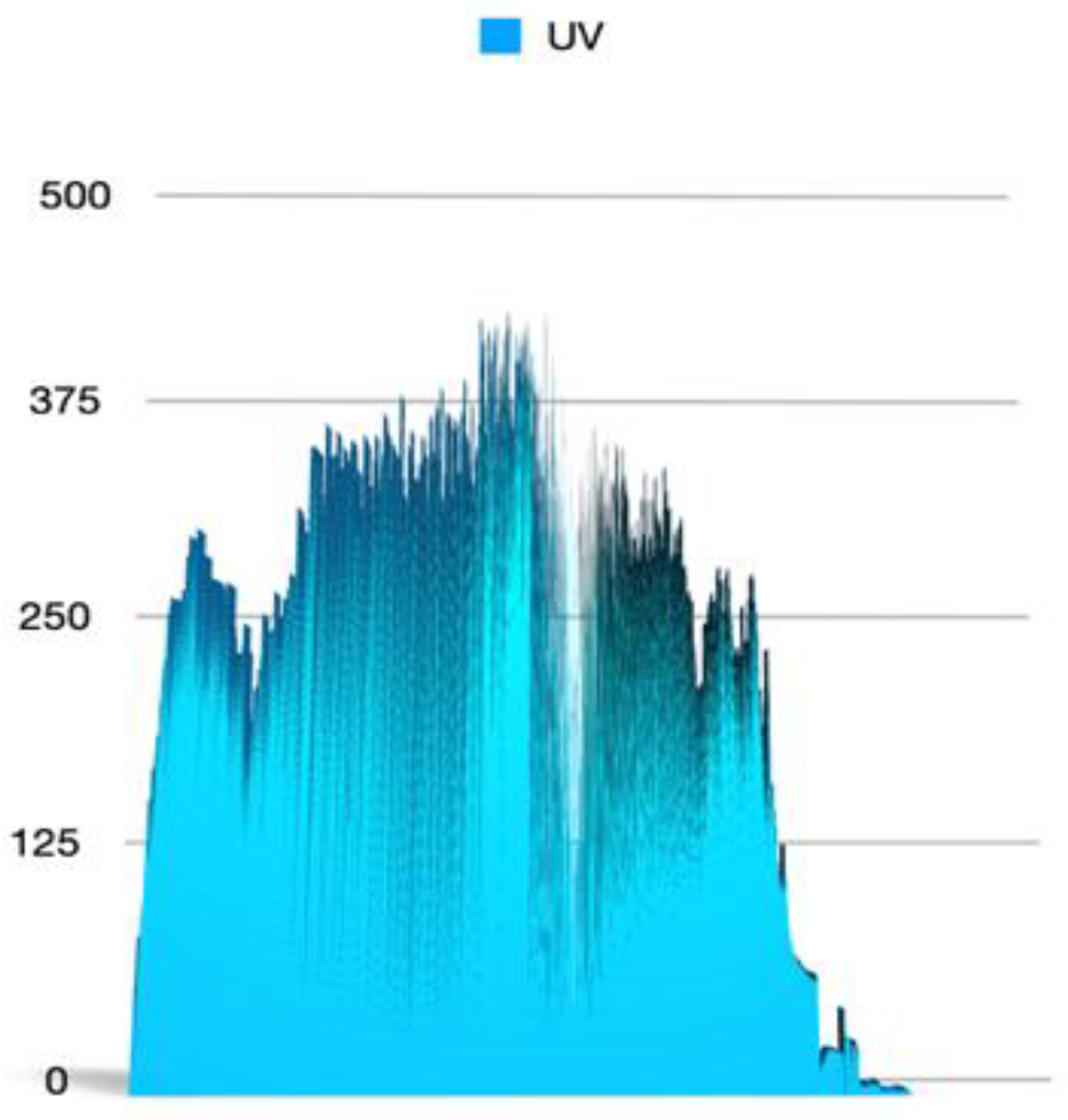
2.4. Stratospheric Flight (Technical Details)
3. Results
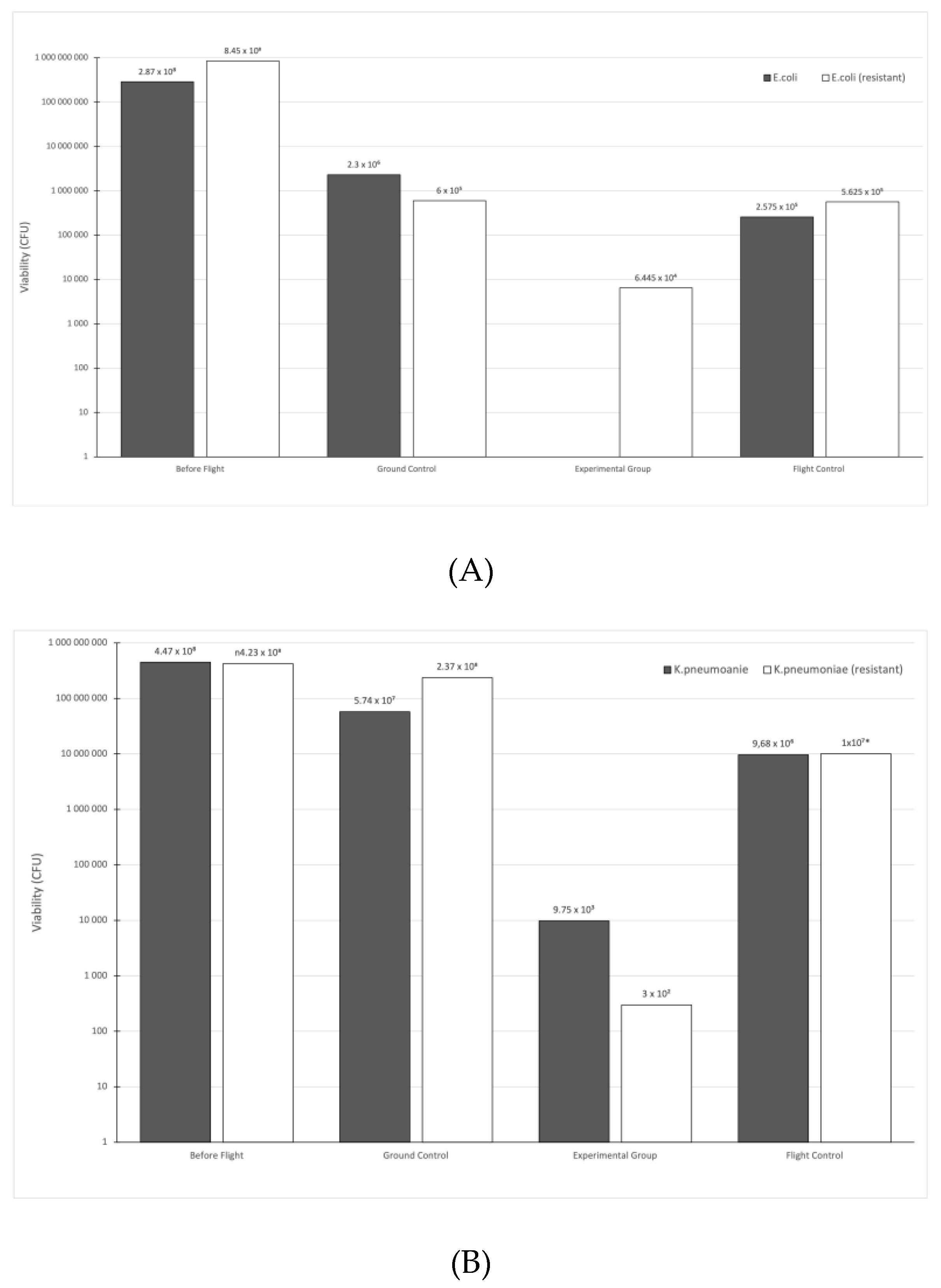
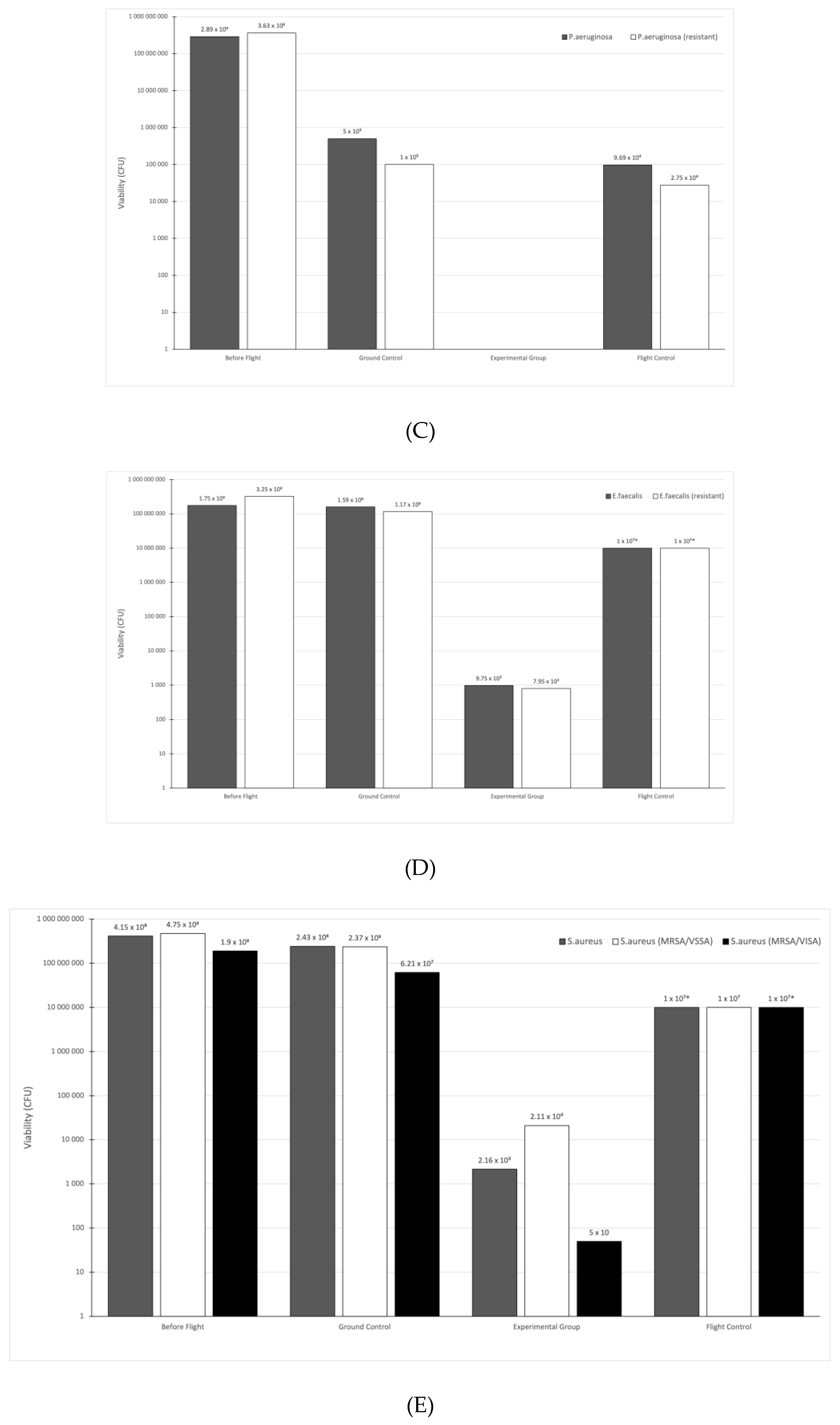
3.1. Viable Count Assay
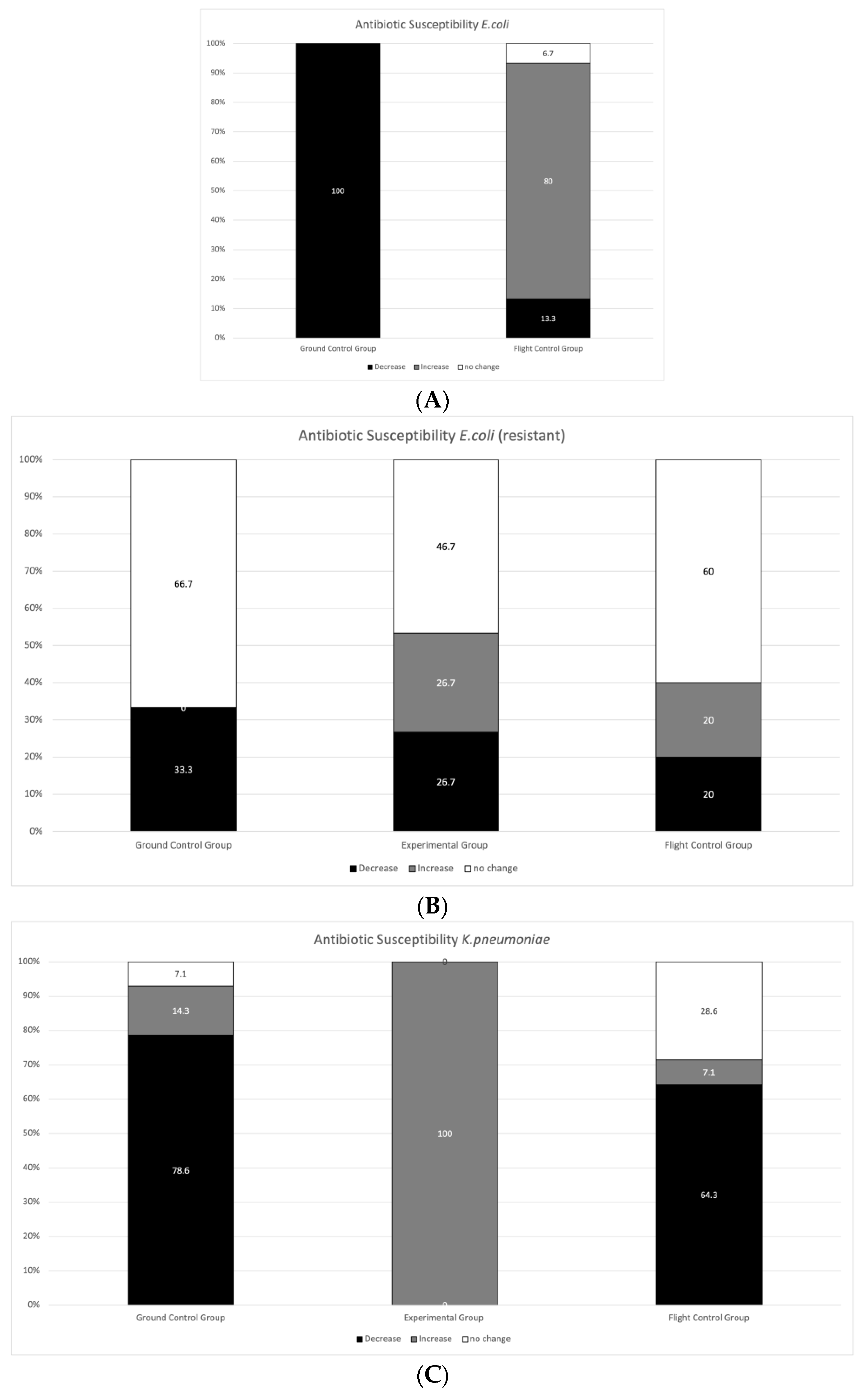
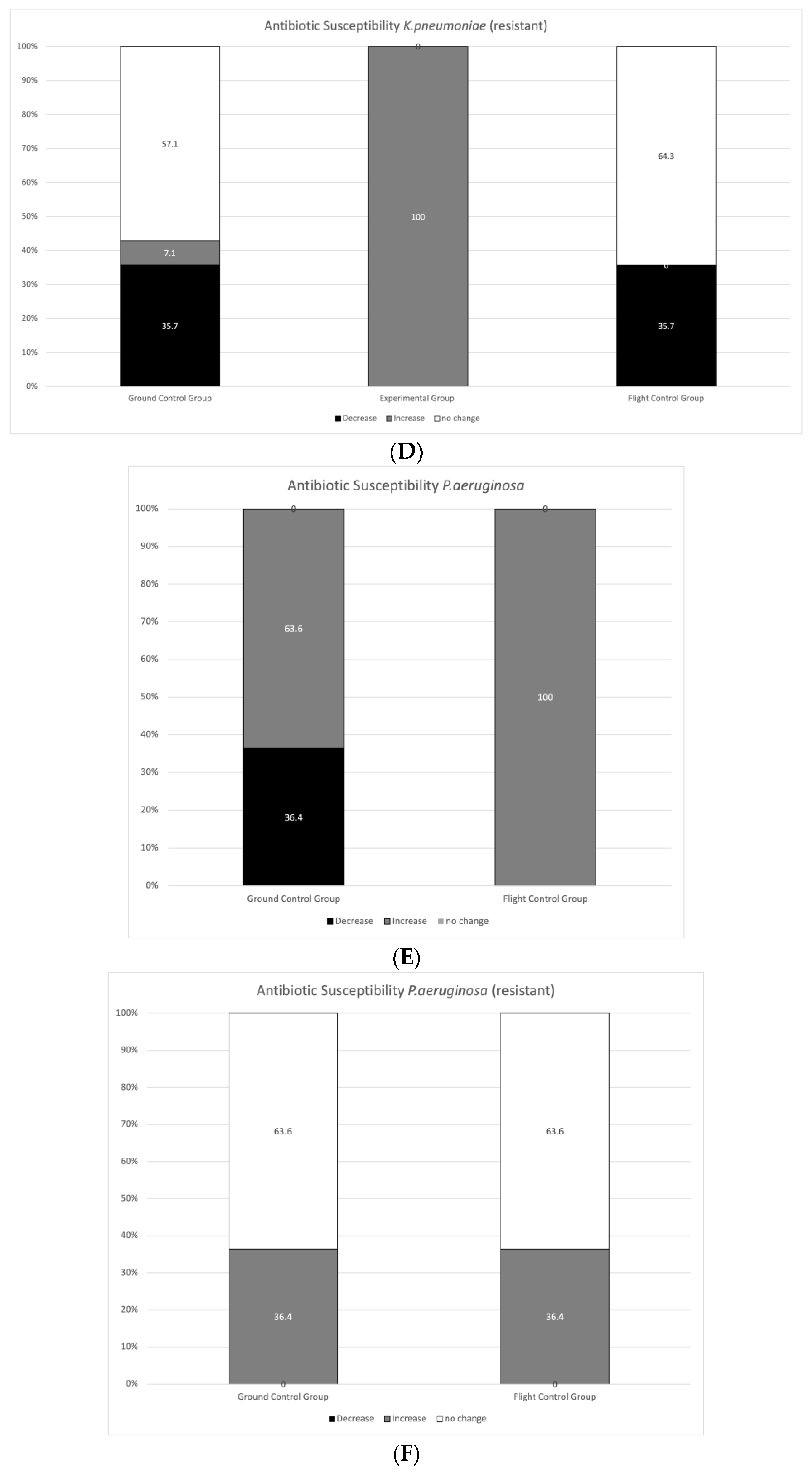
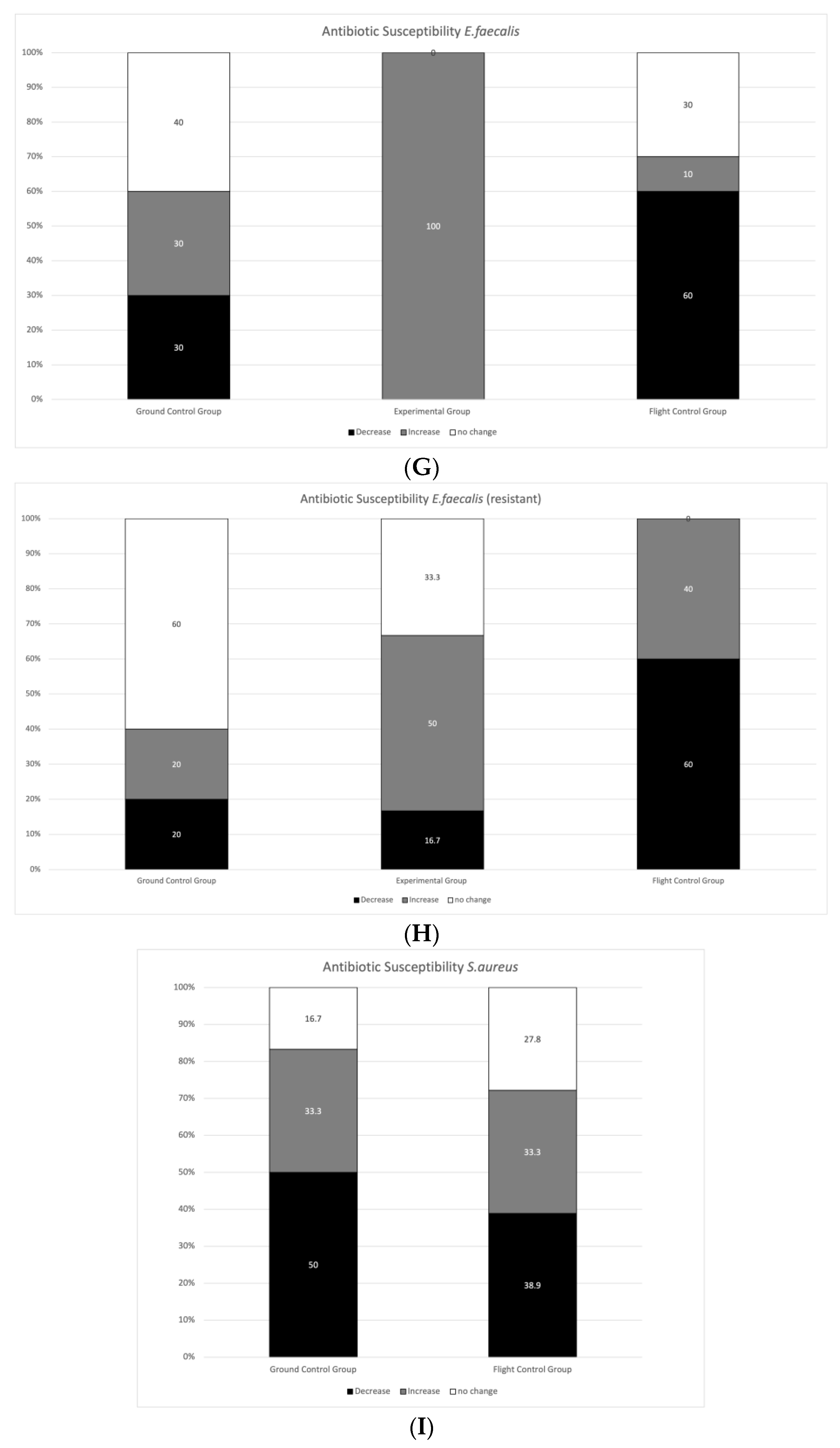
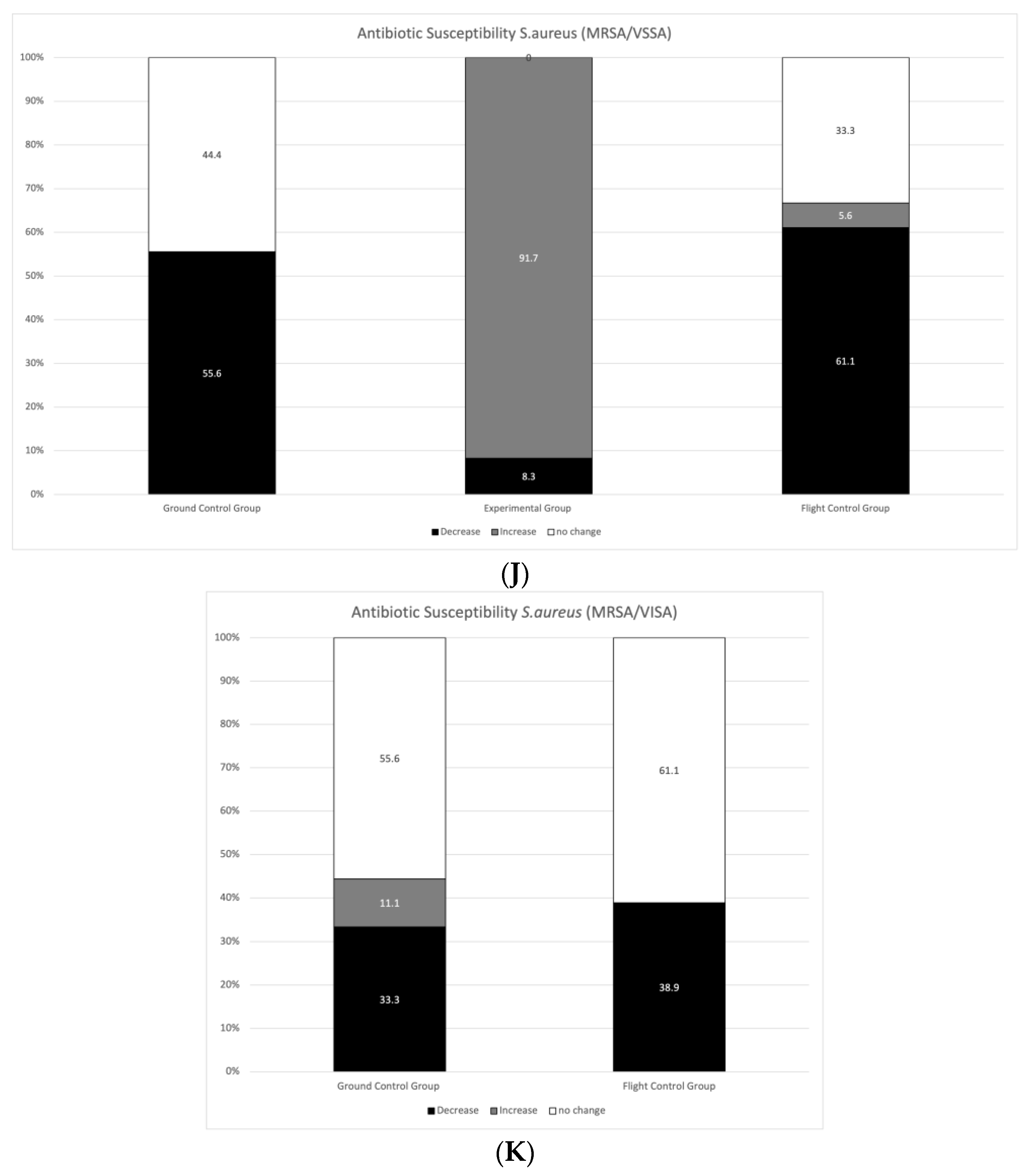
3.2. Antibiotic Susceptibility
3.3. Genes Mediating Antibiotic Resistance
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hidron, A.I.; Edwards, J.R.; Patel, J.; Horan, T.C.; Sievert, D.M.; Pollock, D.A.; Fridkin, S.K. National healthcare safety network team and participating national healthcare safety network facilities. NHSN annual update: Antimicrobial-resistant pathogens associated with healthcare-associated infections: Annual summary of data reported to the national healthcare safety network at the centers for disease control and prevention, 2006–2007. Infect. Control Hosp. Epidemiol. 2008, 29, 996–1011. [Google Scholar] [CrossRef] [PubMed]
- Allegranzi, B.; Nejad, S.B.; Combescure, C.; Graafmans, W.; Attar, H.; Donaldson, L.; Pittet, D. Burden of endemic health-care-associated infection in developing countries: Systematic review and meta-analysis. Lancet 2020, 377, 228–241. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Gao, J.; Tang, W. Nosocomial infection and its molecular mechanisms of antibiotic resistance. Biosci. Trends 2016, 10, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Tenover, F.C. Mechanisms of antimicrobial resistance in bacteria. Am. J. Med. 2006, 119, S3–S10. [Google Scholar] [CrossRef]
- Munita, J.M.; Arias, C.A. Mechanisms of antibiotic resistance. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef]
- Magill, S.S.; Edwards, J.R.; Bamberg, W.; Beldavs, Z.G.; Dumyati, G.; Kainer, M.A.; Lynfield, R.; Maloney, M.; McAllister-Hollod, L.; Nadle, J.; et al. Multistate point-prevalence survey of healthcare-associated infections. N. Engl. J. Med. 2014, 370, 1198–1208. [Google Scholar] [CrossRef] [PubMed]
- Song, M.D.; Wachi, M.; Doi, M.; Ishino, F.; Matsuhaschi, M. Evolution of an inducible penicillin-target protein in methicillin-resistant Staphylococcus aureus by gene fusion. FEBS Lett. 1987, 221, 167–171. [Google Scholar] [CrossRef]
- Baba, T.; Kuwahara-Arai, K.; Uchiyama, I.; Takeuchi, F.; Ito, T.; Hiramatsu, K. Complete genome sequence of Macrococcus caseolyticus strain JCSCS5402, [corrected] reflecting the ancestral genome of the human-pathogenic staphylococci. J. Bacteriol. 2009, 191, 1180–1190. [Google Scholar] [CrossRef]
- García-Álvarez, L.; Holden, M.T.; Lindsay, H.; Webb, C.R.; Brown, D.F.J.; Curran, M.D.; Walpole, E.; Brooks, K.; Pickard, D.J.; Teale, C.; et al. Methicillin-resistant Staphylococcus aureus with a novel mecA homologue in human and bovine populations in the UK and Denmark: A descriptive study. Lancet Infect. Dis. 2011, 11, 595–603. [Google Scholar] [CrossRef]
- Gould, I.M.; David, M.Z.; Esposito, S.; Garau, J.; Lina, G.; Mazzei, T.; Peters, G. New insights into methicillin-resistant Staphylococcus aureus (MRSA) pathogenesis, treatment and resistance. Int. J. Antimicrob. Agents 2012, 39, 96–104. [Google Scholar] [CrossRef]
- Turner, N.A.; Sharma-Kuinkel, B.K.; Maskarinec, S.A.; Eichenberger, E.M.; Shah, P.P.; Carugati, M.; Holland, T.L. Methicillin-resistant Staphylococcus aureus: An overview of basic and clinical research. Nat. Rev. Microbiol. 2019, 17, 203–218. [Google Scholar] [CrossRef] [PubMed]
- McGuinness, W.A.; Malachowa, N.; DeLeo, F.R. Vancomycin resistance in Staphylococcus aureus. Yale J. Biol. Med. 2017, 90, 269–281. [Google Scholar] [PubMed]
- Partridge, S.R. Resistance mechanisms in Enterobacteriaceae. Pathology 2015, 47, 276–284. [Google Scholar] [CrossRef]
- Martin, R.M.; Bachman, M.A. Colonization, infection, and the accessory genome of Klebsiella pneumoniae. Front. Cell. Infect. Microbiol. 2018, 8, 4. [Google Scholar] [CrossRef]
- Miller, W.R.; Munita, J.M.; Arias, C.A. Mechanisms of antibiotic resistance in enterococci. Expert Rev. Anti-Infect. Ther. 2014, 12, 1221–1236. [Google Scholar] [CrossRef]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.J.; Cheng, Z. Antibiotic resistance in Pseudomonas aeruginosa: Mechanisms and alternative therapeutic strategies. Biotechnol. Adv. 2019, 37, 177–192. [Google Scholar] [CrossRef]
- DasSarma, P.; DasSarma, S. Survival of microbes in Earth’s stratosphere. Curr. Opin. Microbiol. 2018, 43, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.J. Microbes in the upper atmosphere and unique opportunities for astrobiology research. Astrobiology. 2013, 13, 981–990. [Google Scholar] [CrossRef]
- Van Leuken, J.; Swart, A.; Havelaar, A.; Pul, A.V.; Van der Hoek, W.; Heederik, D. Atmospheric dispersion modelling of bioaerosols that are pathogenic to humans and livestock—A review to inform risk assessment studies. Microb. Risk Anal. 2016, 1, 19–39. [Google Scholar] [CrossRef]
- Przystupski, D.; Gorska, A.; Rozborska, P.; Bartosik, W.; Michel, O.; Rossowska, J.; Szewczyk, A.; Drag-Zalesinska, M.; Kasperkiewicz, P.; Gorski, J.; et al. The cytoprotective role of antioxidants in mammalian cells under rapidly varying UV conditions during stratospheric balloon campaign. Front. Pharmacol. 2019, 10, 851. [Google Scholar] [CrossRef] [Green Version]
- Chudobova, D.; Cihalova, K.; Jelinkov, P.; Zitka, J.; Nejdl, L.; Guran, R.; Klimanek, M.; Adam, V.; Kizek, R. Effects of stratospheric conditions on the viability, metabolism and proteome of prokaryotic cells. Atmosphere 2015, 6, 1290–1306. [Google Scholar] [CrossRef]
- Khodadad, C.L.; Wong, G.M.; James, L.M.; Thakrar, P.J.; Lane, M.A.; Catechis, J.A.; Smith, D.J. Stratosphere conditions inactivate bacterial endospores from a Mars spacecraft assembly facility. Astrobiology 2017, 17, 337–350. [Google Scholar] [CrossRef] [PubMed]
- DasSarma, P.; Laye, V.; Harvey, J.; Reid, C.; Shultz, J.; Yarborough, A.; Lamb, A.; Koske-Phillips, A.; Herbst, A.; Molina, F.; et al. Survival of halophilic Archaea in Earth’s cold stratosphere. Int. J. Astrobiol. 2017, 16, 321–327. [Google Scholar] [CrossRef]
- Pulschen, A.A.; de Araujo, G.G.; de Carvalho, A.C.S.R.; Cerini, M.F.; de Mendonca Fonseca, L.; Galante, D.; Rodriguez, F. Survival of extremophilic yeasts in the stratospheric environment during balloon flights and in laboratory simulations. Appl. Environ. Microbiol. 2018, 84, e01942-18. [Google Scholar] [CrossRef]
- De Goffau, M.C.; Yang, X.; Van Dijl, J.M.; Harmsen, H.J.M. Bacterial pleomorphism and competition in a relative humidity gradient. Environ. Microbiol. 2009, 11, 809–822. [Google Scholar] [CrossRef]
- Smith, D.J.; Sowa, M.B. Ballooning for biologists: Mission essentials for flying life science experiments to near space on NASA large scientific balloons. Gravit. Space Res. 2017, 5, 52–73. [Google Scholar] [CrossRef]
- Victorin, K. Review of the genotoxicity of ozone. Mutat. Res. Rev. Genet. Toxicol. 1992, 277, 221–238. [Google Scholar] [CrossRef] [PubMed]
- Thanomsub, B.; Anupunpisit, V.; Chanphetch, S.; Watcharachaipong, T.; Poonkhum, R.; Srisukonth, C. Effects of ozone treatment on cell growth and ultrastructural changes in bacteria. J. Gen. Appl. Microbiol. 2002, 48, 193–199. [Google Scholar] [CrossRef]
- Pokorny, N.; Boulter-Bitzer, J.; Hart, M.; Storey, L.; Lee, H.; Trevors, J.T. Hypobaric bacteriology: Growth, cytoplasmic membrane polarization and total cellular fatty acids in Escherichia coli and Bacillus subtilis. Int. J. Astrobiol. 2005, 4, 187. [Google Scholar] [CrossRef]
- Schuerger, A.C.; Nicholson, W.L. Twenty species of hypobarophilic bacteria recovered from diverse soils exhibit growth under simulated Martian conditions at 0.7 kPa. Astrobiology 2016, 16, 964–976. [Google Scholar] [CrossRef]
- Santos, A.L.; Oliveira, V.; Baptista, I.; Gomes, N.C.M.; Almeida, A.; Correia, A.; Cunha, A. Wavelength dependence of biological damage induced by UV radiation on bacteria. Arch. Microbiol. 2013, 195, 63–74. [Google Scholar] [CrossRef]
- Baumstark-Khan, C.; Facius, R. Life under conditions of ionizing radiation. In Astrobiology; Horneck, G., Baumstark-Khan, C., Eds.; Springer: Berlin/Heidelberg, Germany, 2002; pp. 261–284. [Google Scholar] [CrossRef]
- Confalonieri, F.; Sommer, S. Bacterial and archaeal resistance to ionizing radiation. J. Phys. Conf. Ser. 2011, 261, 012005. [Google Scholar] [CrossRef]
- Shivaji, S.; Prakash, J.S. How do bacteria sense and respond to low temperature? Arch. Microbiol. 2010, 192, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Zhang, Z.; Hu, S.; Yu, J. On the molecular mechanism of GC content variation among eubacterial genomes. Biol. Direct 2012, 7, 2. [Google Scholar] [CrossRef]
- Matallana-Surget, S.; Meador, J.A.; Joux, F.; Douki, T. Effect of the GC content of DNA on the distribution of UVB-induced bipyrimidine photoproducts. Photochem. Photobiol. Sci. 2008, 7, 794–801. [Google Scholar] [CrossRef]
- Daly, M.J. A new perspective on radiation resistance based on Deinococcus radiodurans. Nat. Rev. Microbiol. 2009, 7, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Ramos, J.L.; Gallegos, M.T.; Marqués, S.; Ramos-Gonzales, M.I.; Espinosa-Urgel, M.; Segura, A. Responses of Gram-negative bacteria to certain environmental stressors. Curr. Opin. Microbiol. 2001, 4, 166–171. [Google Scholar] [CrossRef]
- Jordan, S.; Hutchings, M.I.; Mascher, T. Cell envelope stress response in Gram-positive bacteria. FEMS Microbiol. Rev. 2008, 32, 107–146. [Google Scholar] [CrossRef]
- Limoli, D.H.; Jones, C.J.; Wozniak, D.J. Bacterial extracellular polysaccharides in biofilm formation and function. Microbiol. Spectr. 2015, 3, 1–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paczosa, M.K.; Mecsas, J. Klebsiella pneumoniae: Going on the offense with a strong defense. Microbiol. Mol. Biol. Rev. 2016, 80, 629–661. [Google Scholar] [CrossRef] [PubMed]
- Durão, P.; Balbontín, R.; Gordo, I. Evolutionary mechanisms shaping the maintenance of antibiotic resistance. Trends Microbiol. 2018, 26, 677–691. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Savalia, C.V. Effect of different stress conditions on antibiotic susceptibility of coagulase positive thermo tolerant Staphylococcus aureus. Octa J. Biosci. 2020, 8, 62–72. [Google Scholar]
- Partridge, S.R.; Kwong, S.M.; Firth, N.; Jensen, S.O. Mobile genetic elements associated with antimicrobial resistance. Clin Microbiol. Rev. 2018, 31, e00088-17. [Google Scholar] [CrossRef]
- Poirel, L.; Walsh, T.R.; Cuvillier, V.; Nordmann, P. Multiplex PCR for detection of acquired carbapenemase genes. Diagn. Microbiol. Infect. Dis. 2011, 70, 119–123. [Google Scholar] [CrossRef]
- Mulvey, M.R.; Grant, J.M.; Plewes, K.; Roscoe, D.; Boyd, D.A. New Delhi metallo-β-lactamase in Klebsiella pneumoniae and Escherichia coli, Canada. Emerg. Infect. Dis. 2011, 17, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Dutka-Malen, S.; Evers, S.; Courvalin, P. Detection of glycopeptide resistance genotypes and identification to the species level of clinically relevant enterococci by PCR. J. Clin. Microbiol. 1995, 33, 1434. [Google Scholar] [CrossRef] [PubMed]
- Francois, P.; Huyghe, A.; Charbonnier, Y.; Bento, M.; Herzig, S.; Topolski, I.; Fleury, B.; Lew, D.; Vaudaux, P.; Harbarth, S.; et al. Use of an automated multiple-locus, variable-number tandem repeat-based method for rapid and high-throughput genotyping of Staphylococcus aureus isolates. J. Clin. Microbiol. 2005, 43, 3346–3355. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time [min] | Temp [°C] 1 | Temp [°C] 2 | Temp [°C] 3 | Temp [°C] 4 |
|---|---|---|---|---|
| 0 | 31.5 | 23.06 | 26.50 | 28.37 |
| 41 | −45.94 | −56.69 | −54.88 | −49.88 |
| 105 | 14.25 | −15.88 | −8.81 | −7.69 |
| 133 | −32.06 | −51.81 | −48.81 | −46.06 |
| 156 | 25.56 | 24.25 | 24.50 | 24.75 |
| 206 | 24.94 | 22.69 | 23.75 | 25.37 |
| Strain | Viability (%) | ||
|---|---|---|---|
| Ground Control a | Experimental Group b | Flight Control c | |
| E. coli | 8 × 10 | 0 | 8.97 × 10−2 |
| E. coli (resistant) | 7.1 × 10−2 | 7.6 × 10−4 | 6.7 × 10−2 |
| K. pneumoniae | 1.284 × 10 | 2.2 × 10−3 | 2.17 |
| K. pneumoniae (resistant) | 5.603 × 10 | 7.1 × 10−5 | 2.36 * |
| P. aeruginosa | 1.7 × 10−1 | 0 | 3.4 × 10−2 |
| P. aeruginosa (resistant) | 2.8 × 10−2 | 0 | 7.6 × 10−3 |
| E. faecalis | 9.086 × 10 | 5.6 × 10−4 | 5.71 * |
| E. faecalis (resistant) | 3.6 × 10 | 2.5 × 10−4 | 3.08 * |
| S. aureus | 5.855 × 10 | 5.2 × 10−4 | 2.41 |
| S. aureus (MRSA/VSSA) | 4.989 × 10 | 4.4 × 10−3 | 2.11 * |
| S. aureus (MRSA/VISA) | 3.268 × 10 | 2.6 × 10−5 | 5.26 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Górecki, I.; Kołodziejczyk, A.; Harasymczuk, M.; Młynarczyk, G.; Szymanek-Majchrzak, K. The Impact of Harsh Stratospheric Conditions on Survival and Antibiotic Resistance Profile of Non-Spore Forming Multidrug Resistant Human Pathogenic Bacteria Causing Hospital-Associated Infections. Int. J. Environ. Res. Public Health 2023, 20, 2787. https://doi.org/10.3390/ijerph20042787
Górecki I, Kołodziejczyk A, Harasymczuk M, Młynarczyk G, Szymanek-Majchrzak K. The Impact of Harsh Stratospheric Conditions on Survival and Antibiotic Resistance Profile of Non-Spore Forming Multidrug Resistant Human Pathogenic Bacteria Causing Hospital-Associated Infections. International Journal of Environmental Research and Public Health. 2023; 20(4):2787. https://doi.org/10.3390/ijerph20042787
Chicago/Turabian StyleGórecki, Ignacy, Agata Kołodziejczyk, Matt Harasymczuk, Grażyna Młynarczyk, and Ksenia Szymanek-Majchrzak. 2023. "The Impact of Harsh Stratospheric Conditions on Survival and Antibiotic Resistance Profile of Non-Spore Forming Multidrug Resistant Human Pathogenic Bacteria Causing Hospital-Associated Infections" International Journal of Environmental Research and Public Health 20, no. 4: 2787. https://doi.org/10.3390/ijerph20042787
APA StyleGórecki, I., Kołodziejczyk, A., Harasymczuk, M., Młynarczyk, G., & Szymanek-Majchrzak, K. (2023). The Impact of Harsh Stratospheric Conditions on Survival and Antibiotic Resistance Profile of Non-Spore Forming Multidrug Resistant Human Pathogenic Bacteria Causing Hospital-Associated Infections. International Journal of Environmental Research and Public Health, 20(4), 2787. https://doi.org/10.3390/ijerph20042787