
Enterobacteria Survival, Percolation, and Leaching on Soil Fertilized with Swine Manure

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Soil and Swine Manure Characterization
2.2. Preparation of the Bacterial Inoculum
2.3. Microbial Survival Assay
2.4. Microbial Percolation Assay
2.5. Microbial Leaching after Rain
2.6. Inactivation Kinetics
2.7. Statistical Analysis
3. Results and Discussion
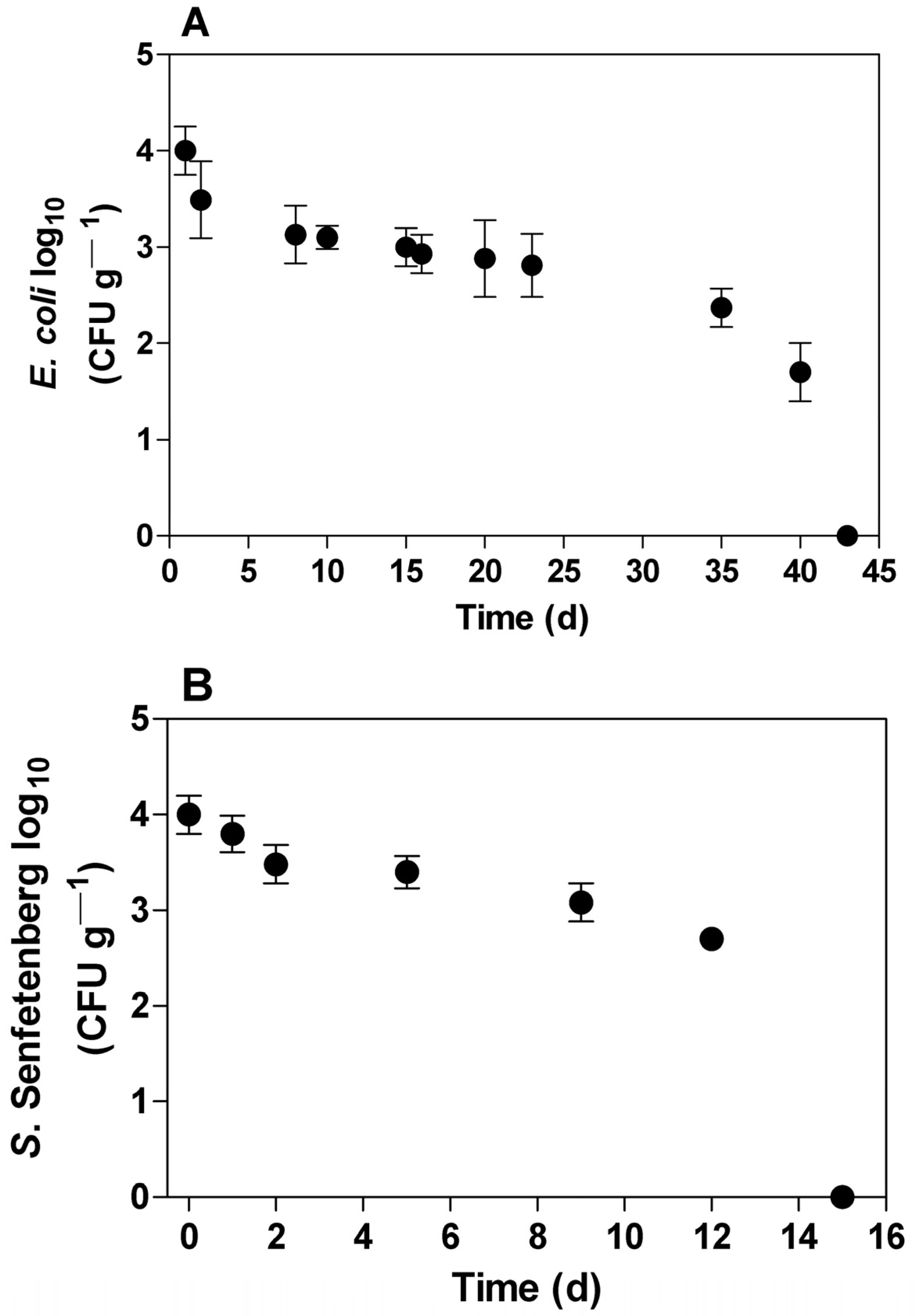
3.1. Enterobacteriaceae Decay Profile in Soil
3.2. Decay Kinetics of Enterobacteriaceae in Soil
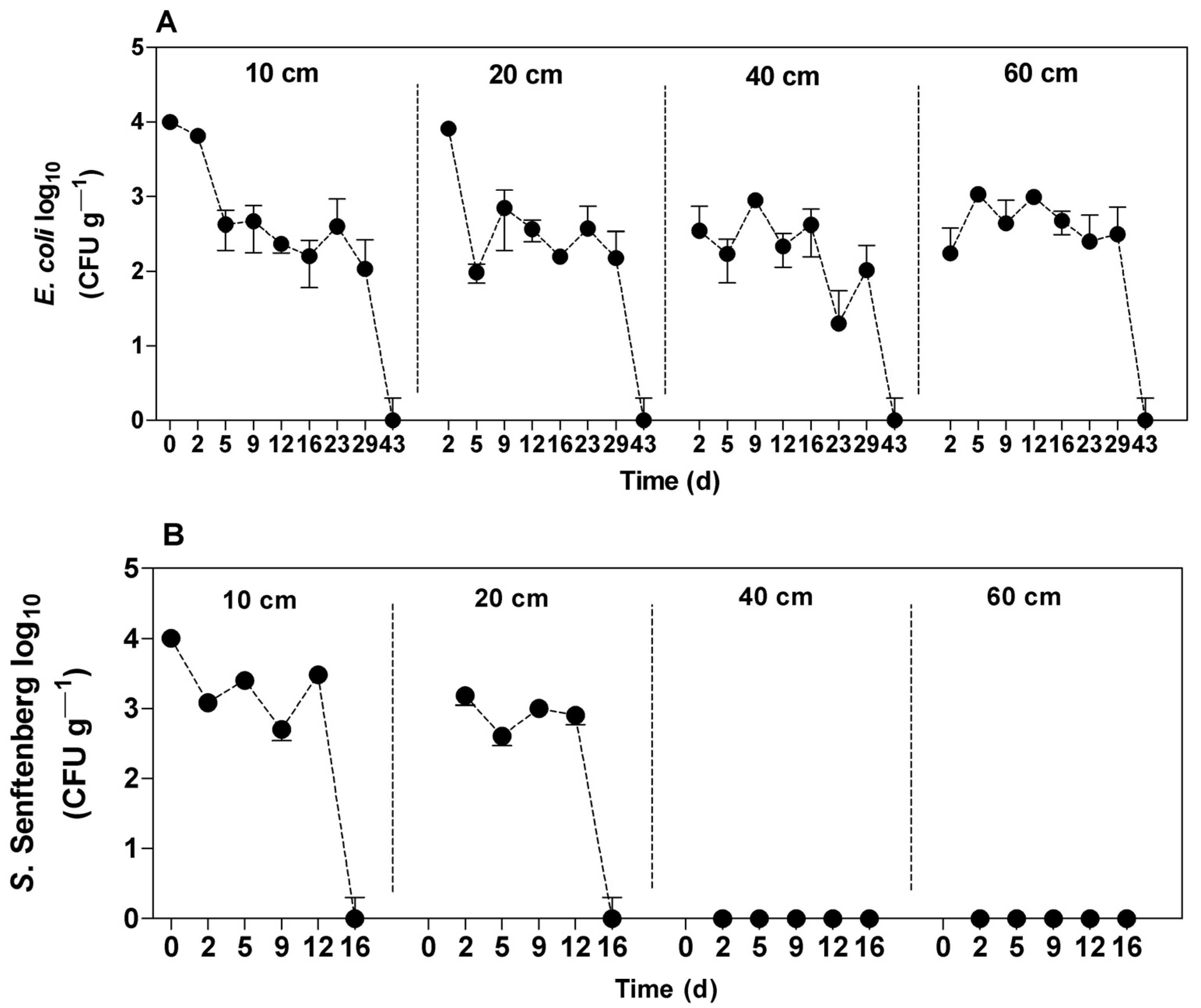
3.3. Percolation of Enterobacteriaceae in Soil
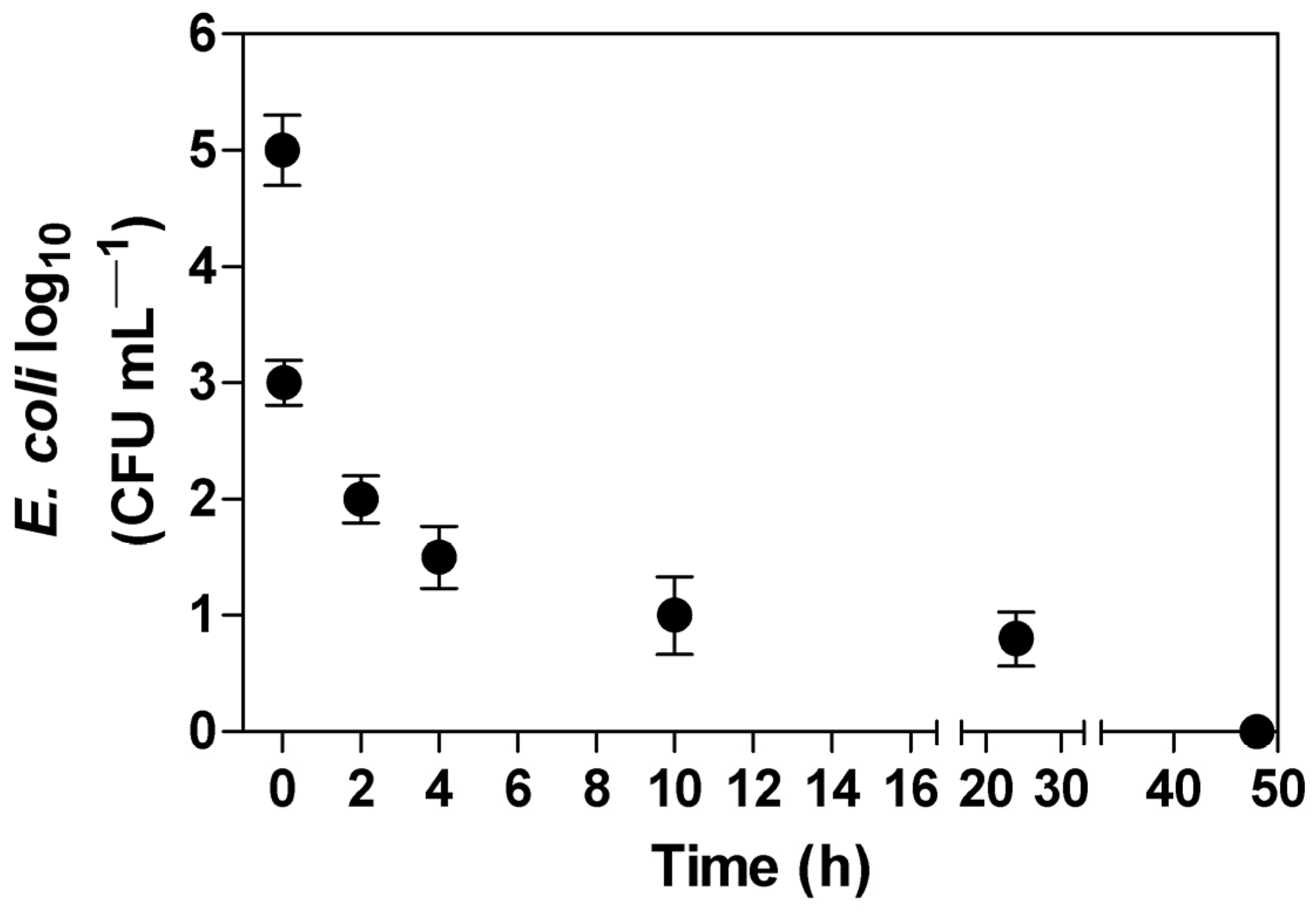
3.4. Leaching of E. coli in Soil
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lima, T.; Domingues, S.; Da Silva, G.J. Manure as a Potential Hotspot for Antibiotic Resistance Dissemination by Horizontal Gene Transfer Events. Vet. Sci. 2020, 7, 110. [Google Scholar] [CrossRef] [PubMed]
- Dadrasnia, A.; de Bona Muñoz, I.; Yáñez, E.H.; Lamkaddam, I.U.; Mora, M.; Ponsá, S.; Ahmed, M.; Argelaguet, L.L.; Williams, P.M.; Oatley-Radcliffe, D.L. Sustainable Nutrient Recovery from Animal Manure: A Review of Current Best Practice Technology and the Potential for Freeze Concentration. J. Clean. Prod. 2021, 315, 128106. [Google Scholar] [CrossRef]
- Cotta, M.A.; Whitehead, T.R.; Zeltwanger, R.L. Isolation, Characterization and Comparison of Bacteria from Swine Faeces and Manure Storage Pits. Environ. Microbiol. 2003, 5, 737–745. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Castillo, F.; Loera-Muro, A.; Jacques, M.; Garneau, P.; Avelar-González, F.; Harel, J.; Guerrero-Barrera, A. Waterborne Pathogens: Detection Methods and Challenges. Pathogens 2015, 4, 307–334. [Google Scholar] [CrossRef] [PubMed]
- Rossi, E.; Cimdins, A.; Lüthje, P.; Brauner, A.; Sjöling, Å.; Landini, P.; Römling, U. “It’s a Gut Feeling”—Escherichia coli Biofilm Formation in the Gastrointestinal Tract Environment. Crit. Rev. Microbiol. 2018, 44, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Havelaar, A.H.; Kirk, M.D.; Torgerson, P.R.; Gibb, H.J.; Hald, T.; Lake, R.J.; Praet, N.; Bellinger, D.C.; de Silva, N.R.; Gargouri, N.; et al. World Health Organization Global Estimates and Regional Comparisons of the Burden of Foodborne Disease in 2010. PLoS Med. 2015, 12, e1001923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bass, D.; Stentiford, G.D.; Wang, H.-C.; Koskella, B.; Tyler, C.R. The Pathobiome in Animal and Plant Diseases. Trends Ecol. Evol. 2019, 34, 996–1008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffith, R.W.; Carlson, S.A.; Krull, A.C. Salmonellosis. In Diseases of Swine; Wiley: Hoboken, NJ, USA, 2019; pp. 912–925. [Google Scholar]
- Santos, S.; Martins, C.; Pereira, C.; Silvestre, A.; Rocha, S. Current Challenges and Perspectives for the Use of Aqueous Plant Extracts in the Management of Bacterial Infections: The Case-Study of Salmonella enterica Serovars. Int. J. Mol. Sci. 2019, 20, 940. [Google Scholar] [CrossRef] [Green Version]
- Ezugworie, F.N.; Igbokwe, V.C.; Onwosi, C.O. Proliferation of Antibiotic-Resistant Microorganisms and Associated Genes during Composting: An Overview of the Potential Impacts on Public Health, Management and Future. Sci. Total Environ. 2021, 784, 147191. [Google Scholar] [CrossRef]
- Franz, E.; Visser, A.; Vandiepeningen, A.; Klerks, M.; Termorshuizen, A.; Vanbruggen, A. Quantification of Contamination of Lettuce by GFP-Expressing Escherichia coli O157:H7 and Salmonella enterica Serovar Typhimurium. Food Microbiol. 2007, 24, 106–112. [Google Scholar] [CrossRef]
- Abu-Ashour, J.; Joy, D.M.; Lee, H.; Whiteley, H.R.; Zelin, S. Transport of Microorganisms through Soil. Water Air Soil Pollut. 1994, 75, 141–158. [Google Scholar] [CrossRef]
- Mantha, S.; Anderson, A.; Acharya, S.P.; Harwood, V.J.; Weidhaas, J. Transport and Attenuation of Salmonella Enterica, Fecal Indicator Bacteria and a Poultry Litter Marker Gene Are Correlated in Soil Columns. Sci. Total Environ. 2017, 598, 204–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villholth, K.G.; Jensen, K.H.; Fredericia, J. Flow and Transport Processes in a Macroporous Subsurface-Drained Glacial till Soil I: Field Investigations. J. Hydrol. 1998, 207, 98–120. [Google Scholar] [CrossRef]
- Horswell, J.; Hewitt, J.; Prosser, J.; Van Schaik, A.; Croucher, D.; Macdonald, C.; Burford, P.; Susarla, P.; Bickers, P.; Speir, T. Mobility and Survival of Salmonella Typhimurium and Human Adenovirus from Spiked Sewage Sludge Applied to Soil Columns. J. Appl. Microbiol. 2010, 108, 104–114. [Google Scholar] [CrossRef] [Green Version]
- Moradi, A.; Mosaddeghi, M.R.; Chavoshi, E.; Safadoust, A.; Soleimani, M. Effect of Crude Oil-Induced Water Repellency on Transport of Escherichia coli and Bromide through Repacked and Physically-Weathered Soil Columns. Environ. Pollut. 2019, 255, 113230. [Google Scholar] [CrossRef] [PubMed]
- Sepehrnia, N.; Bachmann, J.; Hajabbasi, M.A.; Rezanezhad, F.; Lichner, L.; Hallett, P.D.; Coyne, M. Transport, Retention, and Release of Escherichia coli and Rhodococcus Erythropolis through Dry Natural Soils as Affected by Water Repellency. Sci. Total Environ. 2019, 694, 133666. [Google Scholar] [CrossRef]
- Tate, R.L. Cultural and Environmental Factors Affecting the Longevity of Escherichia coli in Histosols. Appl. Environ. Microbiol. 1978, 35, 925–929. [Google Scholar] [CrossRef] [Green Version]
- Moynihan, E.L.; Richards, K.G.; Brennan, F.P.; Tyrrel, S.F.; Ritz, K. Enteropathogen Survival in Soil from Different Land-Uses Is Predominantly Regulated by Microbial Community Composition. Appl. Soil Ecol. 2015, 89, 76–84. [Google Scholar] [CrossRef] [Green Version]
- Brennan, F.P.; Moynihan, E.; Griffiths, B.S.; Hillier, S.; Owen, J.; Pendlowski, H.; Avery, L.M. Clay Mineral Type Effect on Bacterial Enteropathogen Survival in Soil. Sci. Total Environ. 2014, 468–469, 302–305. [Google Scholar] [CrossRef]
- Fenlon, D.R.; Ogden, I.D.; Vinten, A.; Svoboda, I. The Fate of Escherichia coli and E. coli O157 in Cattle Slurry after Application to Land. J. Appl. Microbiol. 2000, 88, 149S–156S. [Google Scholar] [CrossRef]
- Young, I.; Ritz, K. Tillage, Habitat Space and Function of Soil Microbes. Soil Tillage Res. 2000, 53, 201–213. [Google Scholar] [CrossRef]
- Shipitalo, M.J.; Gibbs, F. Potential of Earthworm Burrows to Transmit Injected Animal Wastes to Tile Drains. Soil Sci. Soc. Am. J. 2000, 64, 2103–2109. [Google Scholar] [CrossRef]
- Brennan, F.P.; O’Flaherty, V.; Kramers, G.; Grant, J.; Richards, K.G. Long-Term Persistence and Leaching of Escherichia coli in Temperate Maritime Soils. Appl. Environ. Microbiol. 2010, 76, 1449–1455. [Google Scholar] [CrossRef] [Green Version]
- ABPA. ABPA—Associação Brasileira de Proteína Animal—Relatório Anual. São Paulo. 2021. Available online: Https://Abpa-Br.Org/Relatorios/ (accessed on 23 November 2022).
- Oliveira, P.A.V. Manual de Manejo e Utilizacão Dos Dejetos de Suínos; EMBRAPA-CNPSA: Concórdia, Brazil, 1993; ISBN 0101-6245. [Google Scholar]
- da Rosa, G.A.; Broetto, L.F.; Demczuk, T.; Viancelli, A.; Michelon, W. Water Footprint and Productivity in Broilers and Swine Production in Brazil from 2008 to 2018. Environ. Sci. Pollut. Res. 2022, 29, 73020–73028. [Google Scholar] [CrossRef] [PubMed]
- ABNT. Associação Brasileira de Normas Técnicas NBR 6457; Amostra de Solo—Preparação Para Ensaios de Compactação e Ensaios de Caracterização. ABNT: Rio de Janeiro, Brazil, 2016.
- ABNT. Associação Brasileira de Normas Técnicas (ABNT). NBR 7181; Solo—Análise Granulométrica. ABNT: Rio de Janeiro, Brazil, 2018.
- ABNT. Associação Brasileira de Normas Técnicas (ABNT). NBR 7180; Solo—Determinação Do Limite de Plasticidade. ABNT: Rio de Janeiro, Brazil, 2016.
- ABNT. Associação Brasileira de Normas Técnicas (ABNT). NBR 6459; Solo—Determinação Do Limite de Liquidez. ABNT: Rio de Janeiro, Brazil, 2017.
- DNER. Departamento Nacional de Estradas de Rodagem (DNER). ME 093; Solos—Determinação Da Densidade Real. ABNT: Rio de Janeiro, Brazil, 1994.
- APHA. Standard Methods for the Examination for Water and Wastewater, 22nd ed.; American Water Works Association: Washington, DC, USA, 2012. [Google Scholar]
- Fongaro, G.; García-González, M.C.; Hernández, M.; Kunz, A.; Barardi, C.R.M.; Rodríguez-Lázaro, D. Different Behavior of Enteric Bacteria and Viruses in Clay and Sandy Soils after Biofertilization with Swine Digestate. Front. Microbiol. 2017, 8, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finney, M.; Smullen, J.; Foster, H.A.; Brokx, S.; Storey, D.M. Evaluation of Chromocult Coliform Agar for the Detection and Enumeration of Enterobacteriaceae from Faecal Samples from Healthy Subjects. J. Microbiol. Methods 2003, 54, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Magri, M.E.; Philippi, L.S.; Vinnerås, B. Inactivation of Pathogens in Feces by Desiccation and Urea Treatment for Application in Urine-Diverting Dry Toilets. Appl. Environ. Microbiol. 2013, 79, 2156–2163. [Google Scholar] [CrossRef] [Green Version]
- Ottoson, J.R.; Schnürer, A.; Vinnerås, B. In Situ Ammonia Production as a Sanitation Agent during Anaerobic Digestion at Mesophilic Temperature. Lett. Appl. Microbiol. 2008, 46, 325–330. [Google Scholar] [CrossRef]
- Moretti, S.M.L.; Bertoncini, E.I.; Abreu-Junior, C.H. Characterization of Raw Swine Waste and Effluents Treated Anaerobically: Parameters for Brazilian Environmental Regulation Construction Aiming Agricultural Use. J. Mater. Cycles Waste Manag. 2021, 23, 165–176. [Google Scholar] [CrossRef]
- Blumenthal, U.J.; Duncan Mara, D.; Peasey, A.; Ruiz-Palacios, G.; Stott, R. Guidelines for the Microbiological Quality of Treated Wastewater Used in Agriculture: Recommendations for Revising WHO Guidelines. Bull. World Health Organ. Suppl. 2000, 78, 1104–1116. [Google Scholar]
- Viancelli, A.; Kunz, A.; Steinmetz, R.L.R.; Kich, J.D.; Souza, C.K.; Canal, C.W.; Coldebella, A.; Esteves, P.A.; Barardi, C.R.M. Performance of Two Swine Manure Treatment Systems on Chemical Composition and on the Reduction of Pathogens. Chemosphere 2013, 90, 1539–1544. [Google Scholar] [CrossRef] [Green Version]
- Sánchez, G.; Bosch, A. Survival of Enteric Viruses in the Environment and Food. In Viruses in Foods; Springer International Publishing: Cham, Switzerland, 2016; pp. 367–392. [Google Scholar]
- Alegbeleye, O.O.; Singleton, I.; Sant’Ana, A.S. Sources and Contamination Routes of Microbial Pathogens to Fresh Produce during Field Cultivation: A Review. Food Microbiol. 2018, 73, 177–208. [Google Scholar] [CrossRef]
- Mawdsley, J.L.; Bardgett, R.D.; Merry, R.J.; Pain, B.F.; Theodorou, M.K. Pathogens in Livestock Waste, Their Potential for Movement through Soil and Environmental Pollution. Appl. Soil Ecol. 1995, 2, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Maule, A. Survival of Verocytotoxigenic Escherichia coli O157 in Soil, Water and on Surfaces. J. Appl. Microbiol. 2000, 88, 71S–78S. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, F.A.; Groves, S.J.; Chambers, B.J. Pathogen Survival during Livestock Manure Storage and Following Land Application. Bioresour. Technol. 2005, 96, 135–143. [Google Scholar] [CrossRef]
- Estrada, I.B.; Aller, A.; Aller, F.; Gómez, X.; Morán, A. The Survival of Escherichia coli, Faecal Coliforms and Enterobacteriaceae in General in Soil Treated with Sludge from Wastewater Treatment Plants. Bioresour. Technol. 2004, 93, 191–198. [Google Scholar] [CrossRef]
- Côté, C.; Quessy, S. Persistence of Escherichia coli and Salmonella in Surface Soil Following Application of Liquid Hog Manure for Production of Pickling Cucumbers. J. Food Prot. 2005, 68, 900–905. [Google Scholar] [CrossRef]
- Liu, X.; Gao, C.; Ji, D.; Walker, S.L.; Huang, Q.; Cai, P. Survival of Escherichia coli O157:H7 in Various Soil Particles: Importance of the Attached Bacterial Phenotype. Biol. Fertil. Soils 2017, 53, 209–219. [Google Scholar] [CrossRef]
- Cai, P.; Huang, Q.; Walker, S.L. Deposition and Survival of Escherichia coli O157:H7 on Clay Minerals in a Parallel Plate Flow System. Environ. Sci. Technol. 2013, 47, 1896–1903. [Google Scholar] [CrossRef]
- Huang, Q.; Wu, H.; Cai, P.; Fein, J.B.; Chen, W. Atomic Force Microscopy Measurements of Bacterial Adhesion and Biofilm Formation onto Clay-Sized Particles. Sci. Rep. 2015, 5, 16857. [Google Scholar] [CrossRef] [Green Version]
- Busscher, H.J.; van der Mei, H.C. How Do Bacteria Know They Are on a Surface and Regulate Their Response to an Adhering State? PLoS Pathog. 2012, 8, e1002440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donlan, R.M.; Costerton, J.W. Biofilms: Survival Mechanisms of Clinically Relevant Microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef] [Green Version]
- Drummond, J.D.; Davies-Colley, R.J.; Stott, R.; Sukias, J.P.; Nagels, J.W.; Sharp, A.; Packman, A.I. Microbial Transport, Retention, and Inactivation in Streams: A Combined Experimental and Stochastic Modeling Approach. Environ. Sci. Technol. 2015, 49, 7825–7833. [Google Scholar] [CrossRef] [PubMed]
- Tyrrel, S.F.; Quinton, J.N. Overland Flow Transport of Pathogens from Agricultural Land Receiving Faecal Wastes. J. Appl. Microbiol. 2003, 94, 87–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krog, J.S.; Forslund, A.; Larsen, L.E.; Dalsgaard, A.; Kjaer, J.; Olsen, P.; Schultz, A.C. Leaching of Viruses and Other Microorganisms Naturally Occurring in Pig Slurry to Tile Drains on a Well-Structured Loamy Field in Denmark. Hydrogeol. J. 2017, 25, 1045–1062. [Google Scholar] [CrossRef] [Green Version]
- Bernal, M.P.; Sommer, S.G.; Chadwick, D.; Qing, C.; Guoxue, L.; Michel, F.C. Current Approaches and Future Trends in Compost Quality Criteria for Agronomic, Environmental, and Human Health Benefits. In Advances in Agronomy; Elsevier Academic Press Inc: San Diego, CA, USA, 2017; pp. 143–233. [Google Scholar]
- Ethelberg, S.; Mølbak, K.; Josefsen, M.H. Bacteria: Salmonella Non-Typhi. In Encyclopedia of Food Safety; Elsevier: Amsterdam, The Netherlands, 2014; pp. 501–514. [Google Scholar]
- El-Hajj, Z.W.; Newman, E.B. How Much Territory Can a Single E. coli Cell Control? Front. Microbiol. 2015, 6, 309. [Google Scholar] [CrossRef]
- Smith, M.S.; Thomas, G.W.; White, R.E.; Ritonga, D. Transport of Escherichia coli Through Intact and Disturbed Soil Columns. J. Environ. Qual. 1985, 14, 87–91. [Google Scholar] [CrossRef]
- Jacobsen, C.S.; Bech, T.B. Soil Survival of Salmonella and Transfer to Freshwater and Fresh Produce. Food Res. Int. 2012, 45, 557–566. [Google Scholar] [CrossRef]
- Gannon, J.T.; Mingelgrin, U.; Alexander, M.; Wagenet, R.J. Bacterial Transport through Homogeneous Soil. Soil Biol. Biochem. 1991, 23, 1155–1160. [Google Scholar] [CrossRef]
- Wang, Y.; Bradford, S.A.; Šimůnek, J. Estimation and Upscaling of Dual-Permeability Model Parameters for the Transport of E. coli D21g in Soils with Preferential Flow. J. Contam. Hydrol. 2014, 159, 57–66. [Google Scholar] [CrossRef] [Green Version]
- Stenström, T.A. Bacterial Hydrophobicity, an Overall Parameter for the Measurement of Adhesion Potential to Soil Particles. Appl. Environ. Microbiol. 1989, 55, 142–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huysman, F.; Verstraete, W. Water-Facilitated Transport of Bacteria in Unsaturated Soil Columns: Influence of Cell Surface Hydrophobicity and Soil Properties. Soil Biol. Biochem. 1993, 25, 83–90. [Google Scholar] [CrossRef]
- Saini, R.; Halverson, L.J.; Lorimor, J.C. Rainfall Timing and Frequency Influence on Leaching of Escherichia coli RS2G through Soil Following Manure Application. J. Environ. Qual. 2003, 32, 1865–1872. [Google Scholar] [CrossRef] [Green Version]
- Sepehrnia, N.; Memarianfard, L.; Moosavi, A.A.; Bachmann, J.; Guggenberger, G.; Rezanezhad, F. Bacterial Mobilization and Transport through Manure Enriched Soils: Experiment and Modeling. J. Environ. Manage. 2017, 201, 388–396. [Google Scholar] [CrossRef]
- Unc, A.; Goss, M.J. Transport of Bacteria from Manure and Protection of Water Resources. Appl. Soil Ecol. 2004, 25, 1–18. [Google Scholar] [CrossRef]
- Blaustein, R.A.; Pachepsky, Y.A.; Shelton, D.R.; Hill, R.L. Release and Removal of Microorganisms from Land-Deposited Animal Waste and Animal Manures: A Review of Data and Models. J. Environ. Qual. 2015, 44, 1338–1354. [Google Scholar] [CrossRef]
- Fangueiro, D.; Surgy, S.; Napier, V.; Menaia, J.; Vasconcelos, E.; Coutinho, J. Impact of Slurry Management Strategies on Potential Leaching of Nutrients and Pathogens in a Sandy Soil Amended with Cattle Slurry. J. Environ. Manage. 2014, 146, 198–205. [Google Scholar] [CrossRef]
- Stocker, M.D.; Pachepsky, Y.A.; Hill, R.L.; Shelton, D.R. Depth-Dependent Survival of Escherichia coli and Enterococci in Soil after Manure Application and Simulated Rainfall. Appl. Environ. Microbiol. 2015, 81, 4801–4808. [Google Scholar] [CrossRef] [Green Version]
- Fierer, N.; Schimel, J.P.; Holden, P.A. Variations in Microbial Community Composition through Two Soil Depth Profiles. Soil Biol. Biochem. 2003, 35, 167–176. [Google Scholar] [CrossRef]
- Alegbeleye, O.O.; Sant’Ana, A.S. Manure-Borne Pathogens as an Important Source of Water Contamination: An Update on the Dynamics of Pathogen Survival/Transport as Well as Practical Risk Mitigation Strategies. Int. J. Hyg. Environ. Health 2020, 227, 113524. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Soil particle size | Clay–Ø < 0.002 mm (%) | 72.0 |
| Silt–0.002 mm < Ø < 0.06 mm (%) | 22.5 | |
| Thin sand–0.06 mm < Ø < 0.2 mm (%) | 1.8 | |
| Medium sand–0.2 mm < Ø < 0.6 mm (%) | 2.2 | |
| Coarse sand–0.6 mm < Ø < 2.0 mm (%) | 1.5 | |
| Boulder–2.0 mm < Ø < 60 mm (%) | 0.0 | |
| Geotechnical characteristics | Liquid limit (%) | 62 |
| Plastic limit (%) | 37 | |
| Plastic index (%) | 25 | |
| Specific weight of grains (g cm−3) | 2.85 | |
| Textural classification | Clay | |
| H.R.B *-A.A.S.H.T.O. ** | A7-5 |
| Characteristic | Value |
|---|---|
| Total solids (g L−1) | 3–8 |
| Total organic carbon (g L−1) | 1.5–6.5 |
| BOD5 (g L−1) | 2.5–4.5 |
| Alkalinity as CaCO3 (g L−1) | 5–8 |
| Phosphate (g L−1) | 0.048–0.07 |
| Total nitrogen (g L−1) | 1.5–2 |
| Ammoniacal nitrogen (g L−1) | 0.9–1.5 |
| pH | 7.9 |
| Microorganism | k (d−1) | T90 (d) | r2 |
|---|---|---|---|
| E. coli | 0.1029 | 9.71 | 0.85 |
| S. senftenberg | 0.096 | 10.42 | 0.94 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Michelon, W.; Peter, N.R.W.; Schneider, T.M.; Segalla, D.C.; Viancelli, A. Enterobacteria Survival, Percolation, and Leaching on Soil Fertilized with Swine Manure. Int. J. Environ. Res. Public Health 2023, 20, 5283. https://doi.org/10.3390/ijerph20075283
Michelon W, Peter NRW, Schneider TM, Segalla DC, Viancelli A. Enterobacteria Survival, Percolation, and Leaching on Soil Fertilized with Swine Manure. International Journal of Environmental Research and Public Health. 2023; 20(7):5283. https://doi.org/10.3390/ijerph20075283
Chicago/Turabian StyleMichelon, William, Nivia Rosana Weber Peter, Tamili Martins Schneider, Dara Cristina Segalla, and Aline Viancelli. 2023. "Enterobacteria Survival, Percolation, and Leaching on Soil Fertilized with Swine Manure" International Journal of Environmental Research and Public Health 20, no. 7: 5283. https://doi.org/10.3390/ijerph20075283
APA StyleMichelon, W., Peter, N. R. W., Schneider, T. M., Segalla, D. C., & Viancelli, A. (2023). Enterobacteria Survival, Percolation, and Leaching on Soil Fertilized with Swine Manure. International Journal of Environmental Research and Public Health, 20(7), 5283. https://doi.org/10.3390/ijerph20075283