Fatty Acids, Hydrocarbons and Terpenes of Nannochloropsis and Nannochloris Isolates with Potential for Biofuel Production

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation and Cultivation of Microalgae
2.2. Obtaining Axenic Cultures
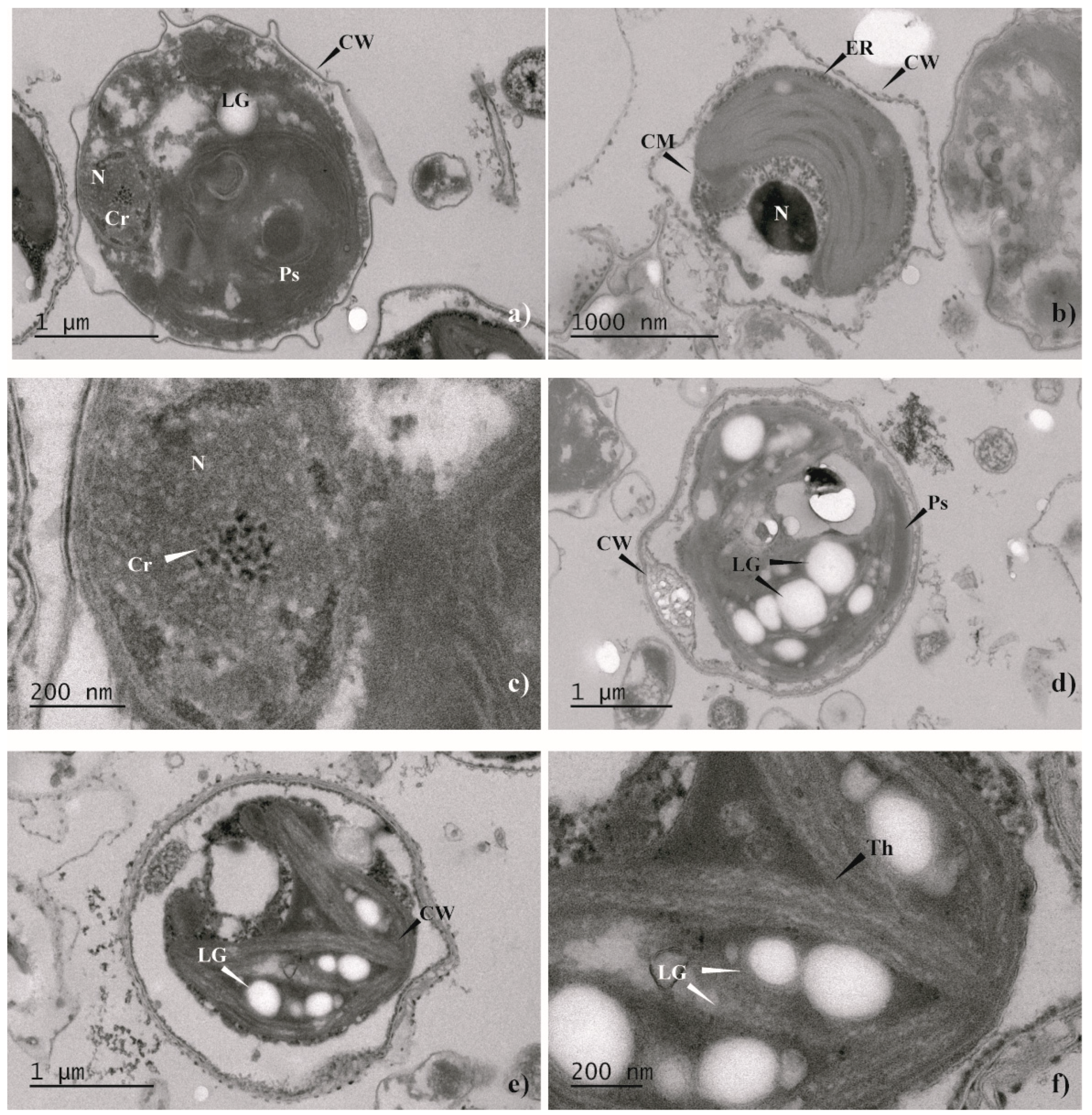
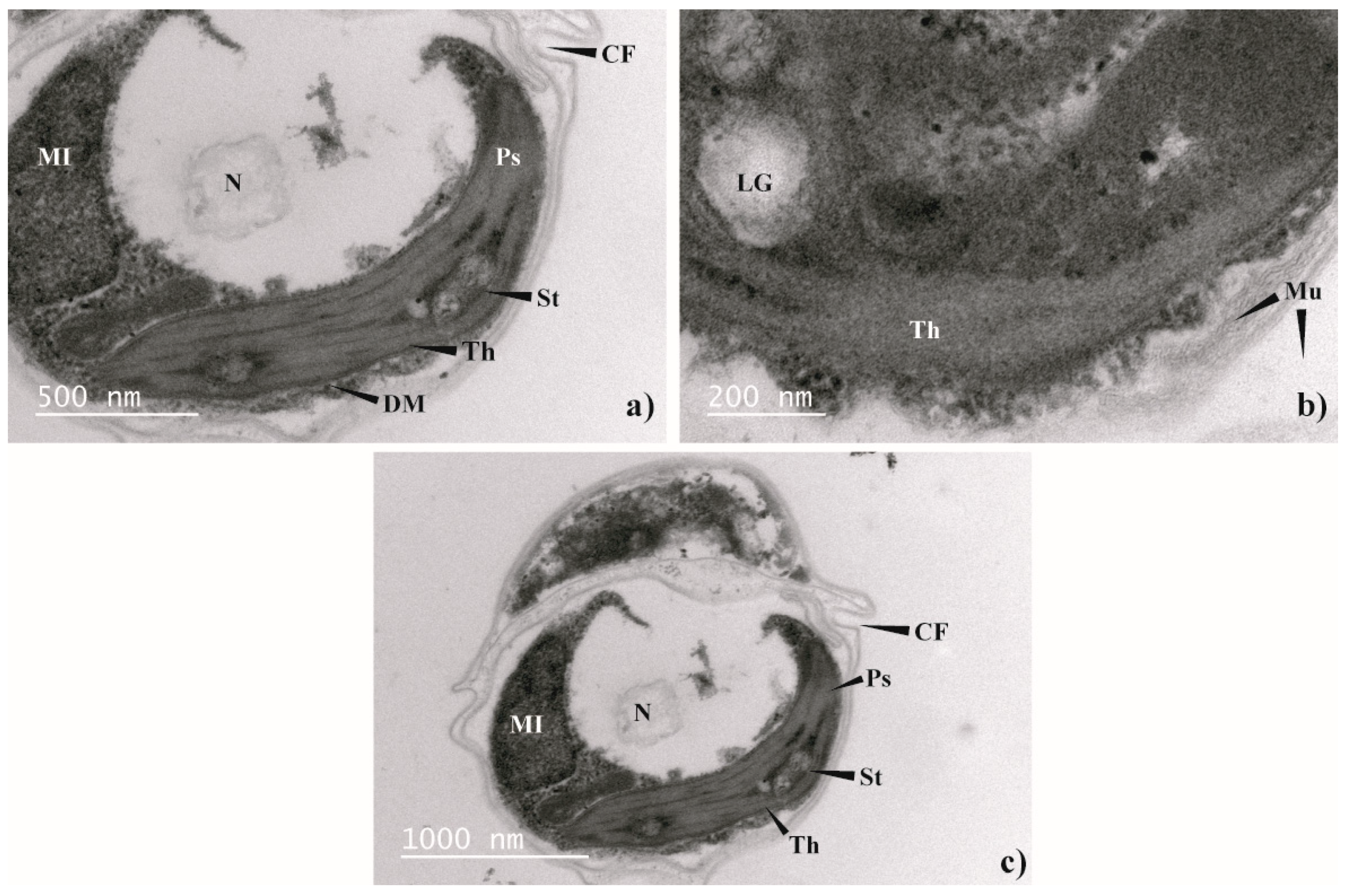
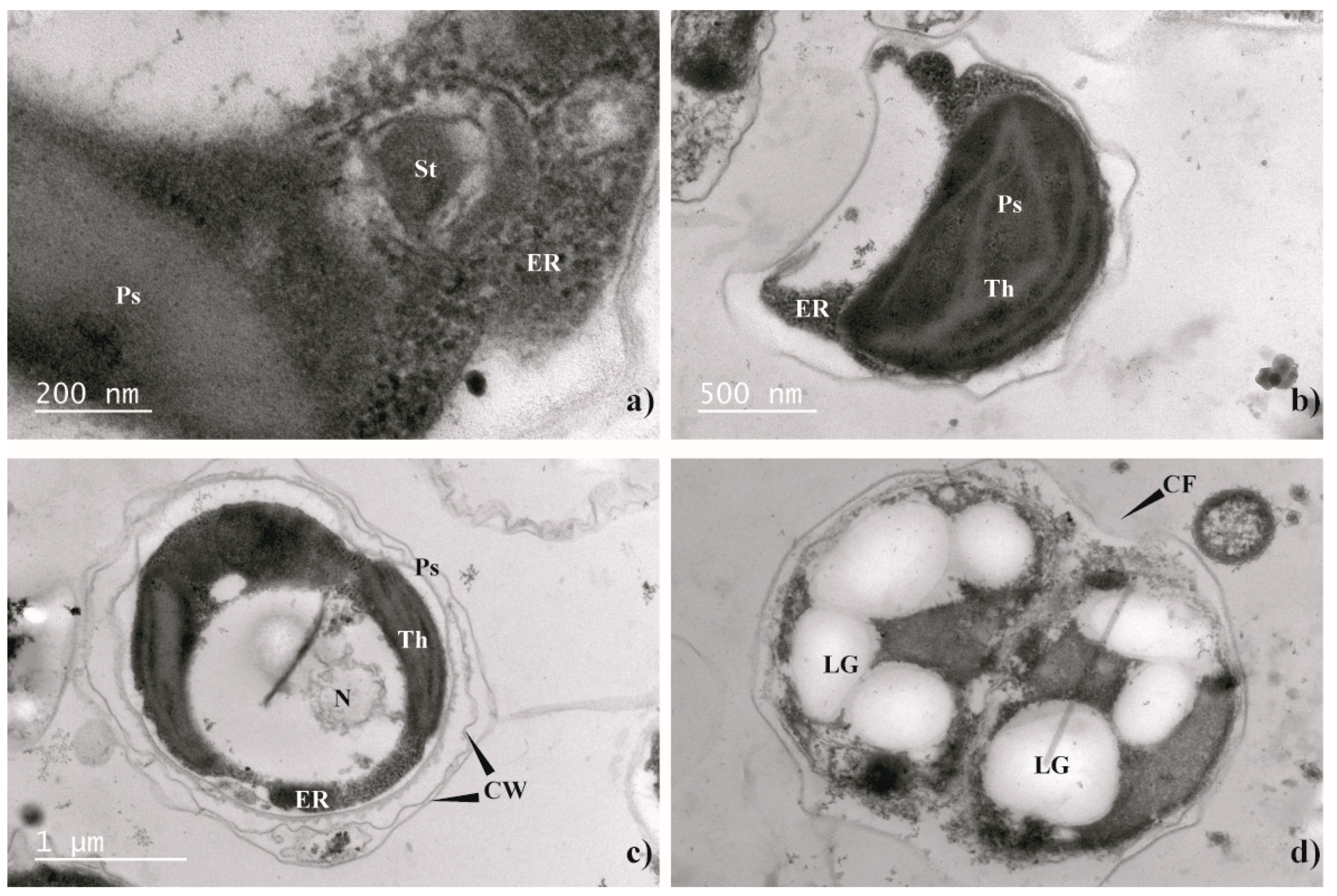
2.3. Morphological Analysis. Light Microscopy and Transmission Electron Microscopy (TEM)
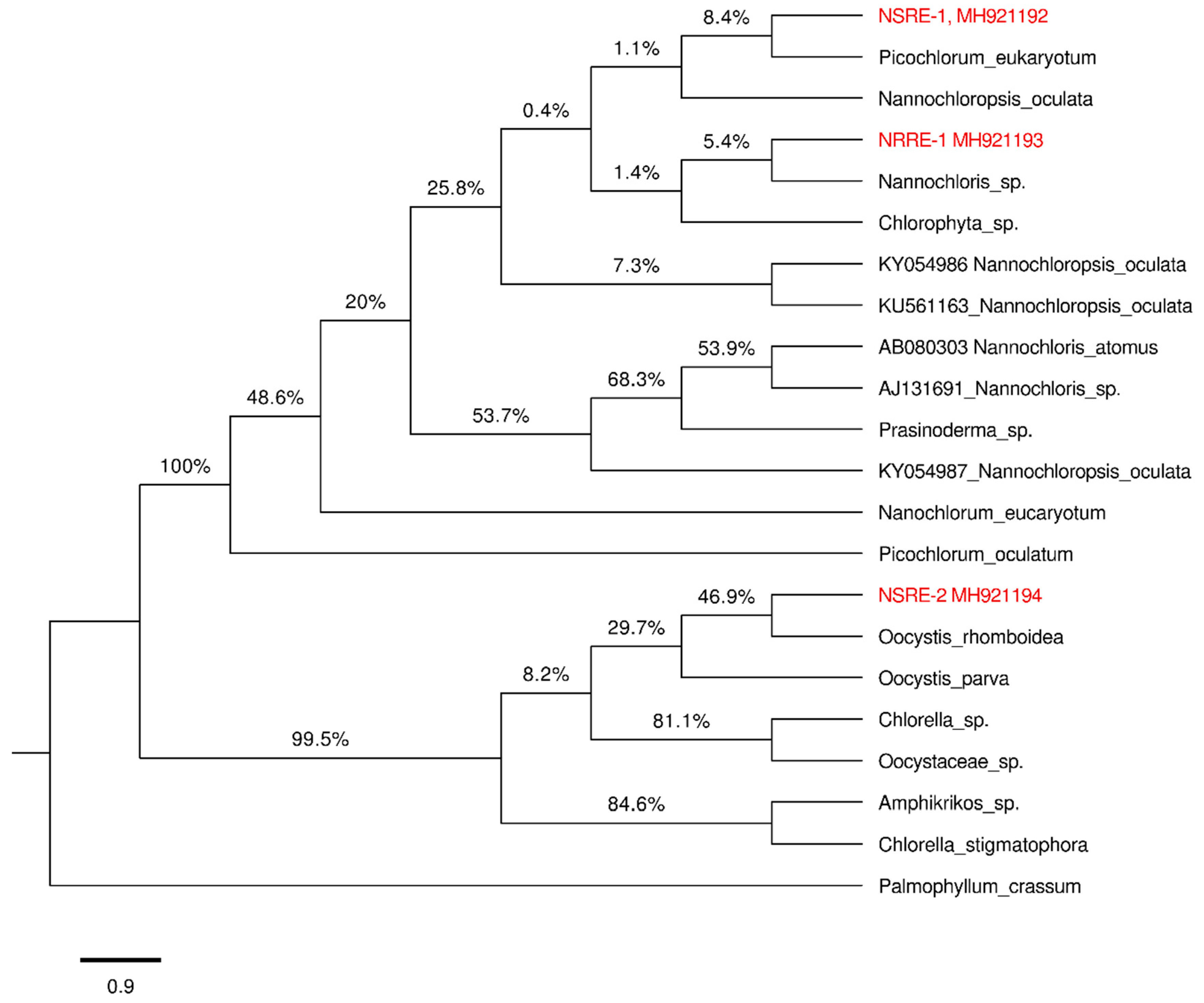
2.4. DNA Extraction and 18S rRNA Gene Amplification
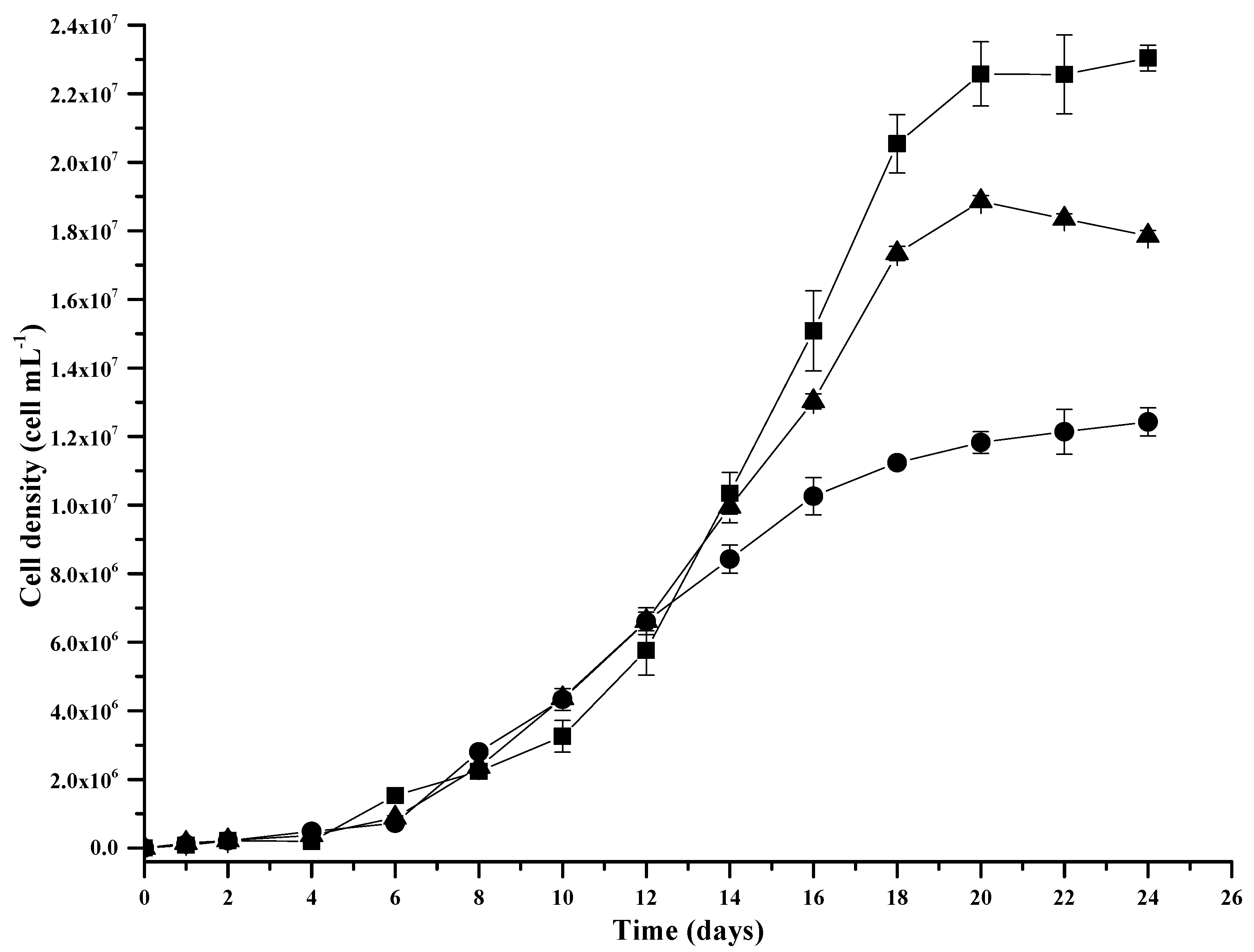
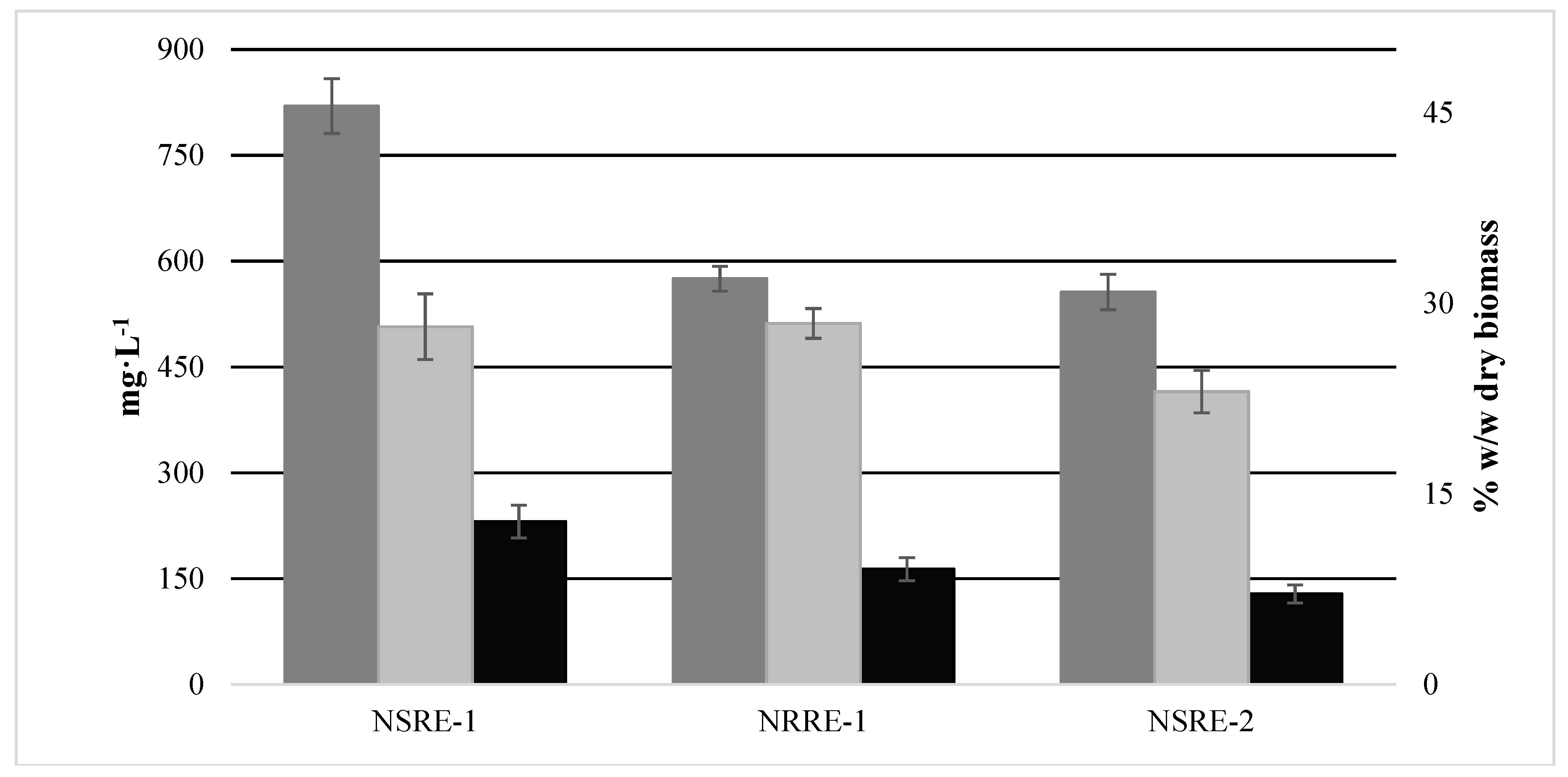
2.5. Biomass Production
2.6. Total Lipid Extraction
2.7. Thin Layer Chromatography
2.8. Column Chromatography
2.9. Transesterification
2.10. Lipid Profile Analysis (GC-MS)
2.11. Catalytic Hydrodeoxygenation of Oxygenates from Strain NSRE-2
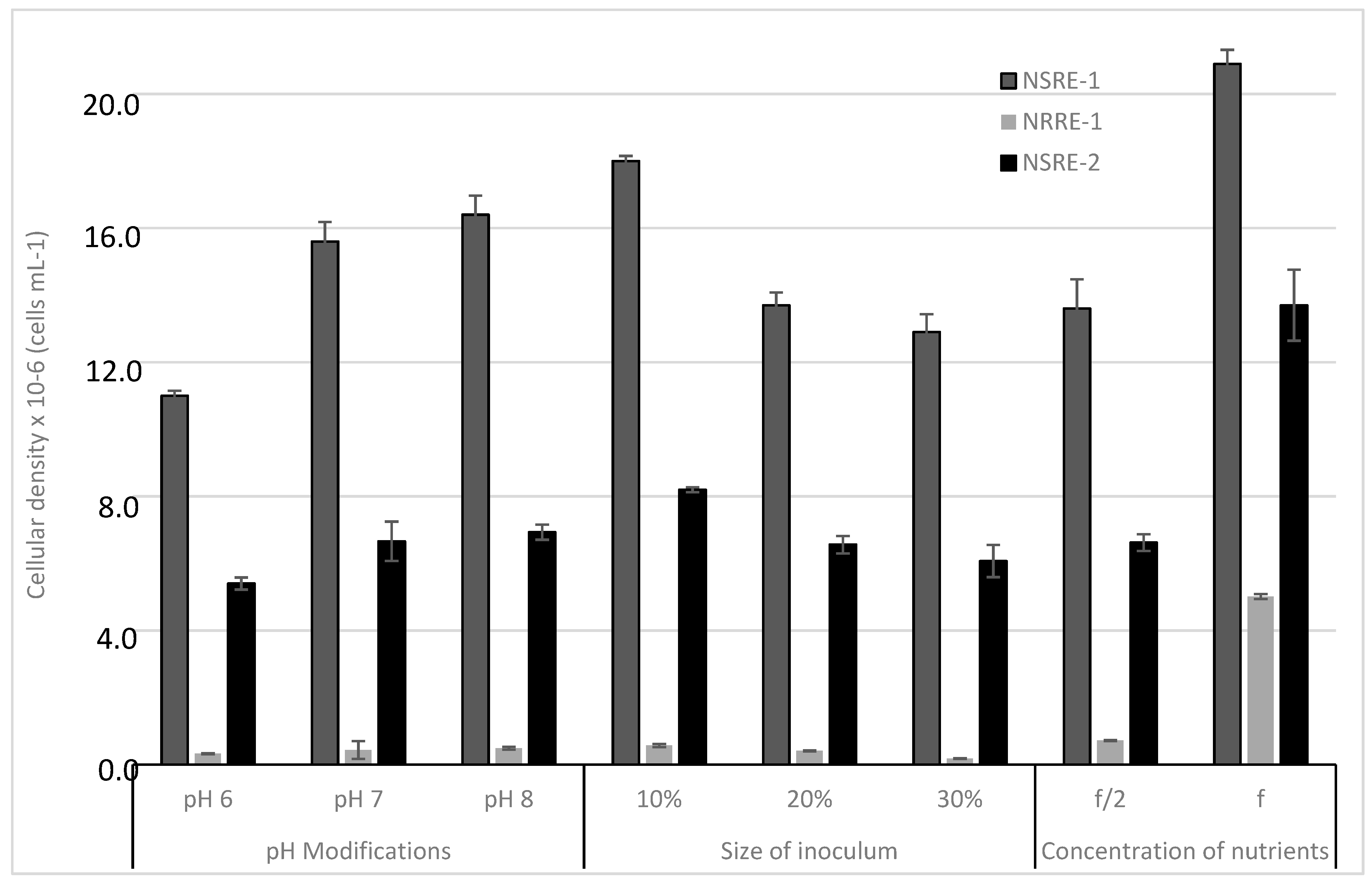
2.12. Statistical Analysis
3. Results and Discussion
3.1. Obtaining Axenic Cultures
3.2. Morphological Analysis (Light Microscopy and TEM)
3.3. 18S rRNA Analysis
3.4. Biomass Concentration, Lipid Extraction and Analysis
3.5. Lipid Profile Analysis (GC-MS)
3.5.1. Hydrocarbons
3.5.2. FAMEs
3.5.3. Organic acids
3.5.4. Terpenes
3.5.5. Sterols
3.5.6. Amines
3.6. Potential of Lipids as Feedstock for Biofuel Production
- i)
- The equation reported by [63] was used in this work for the calculation of the cold filter plugging point (CFPP):where SATC16–C24 is the content (wt %) of saturated FAMEs having chains with 16 to 24 C atoms.CFPP (°C) = −26 + 〖5.76(〖SAT〗_(C16–C24))〗^0.5
- ii)
- Density at 15 °C, kinematic viscosity, higher heating value and oxidative stability of methyl oleate were taken from reference [64].
3.6.1. Analysis of the Hydrocarbon, FAME and Oxygenated Fractions as Fuel Precursor
3.6.2. Catalytic Hydrodeoxygenation of Oxygenates from Strain NSRE-2
3.6.3. Estimation of Biodiesel Properties
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Piloto-Rodríguez, R.; Sánchez-Borroto, Y.; Melo-Espinosa, E.A.; Verhelst, S. Assessment of diesel engine performance when fueled with biodiesel from algae and microalgae: An overview. Renew. Sustain. Energy Rev. 2017, 69, 833–842. [Google Scholar] [CrossRef]
- Velasquez-Orta, S.B.; Lee, J.G.M.; Harvey, A.P. Evaluation of FAME production from wet marine and freshwater microalgae by in situ transesterification. Biochem. Eng. J. 2013, 76, 83–89. [Google Scholar] [CrossRef]
- Maeda, Y.; Yoshino, T.; Matsunaga, T.; Matsumoto, M.; Tanaka, T. Marine microalgae for production of biofuels and chemicals. Curr. Opin. Biotechnol. 2018, 50, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Richmond, A. Biological principles of mass cultivation. Handb. Microalgal Cult. Biotechnol. Appl. Phycol. 2004, 125–177. [Google Scholar]
- Li, Z.; Haifeng, L.; Zhang, Y.; Shanshan, M.; Baoming, L.; Zhidan, L.; Na, D.; Minsheng, L.; Buchun, S.; Jianwen, L. Effects of strain, nutrients concentration and inoculum size on microalgae culture for bioenergy from post hydrothermal liquefaction wastewater. Int. J. Agric. Biol. Eng. 2017, 10, 194–204. [Google Scholar]
- Bartley, M.L.; Boeing, W.J.; Dungan, B.N.; Holguin, F.O.; Schaub, T. pH effects on growth and lipid accumulation of the biofuel microalgae Nannochloropsis salina and invading organisms. J. Appl. Phycol. 2014, 26, 1431–1437. [Google Scholar] [CrossRef]
- Sánchez, S.; Martínez, M.E.; Espinola, F. Biomass production and biochemical variability of the marine microalga Isochrysis galbana in relation to culture medium. Biochem. Eng. J. 2000, 6, 13–18. [Google Scholar] [CrossRef]
- Cao, S.; Zhang, X.; Fan, X.; Qiao, H.; Liang, C.; Xu, D.; Mou, S.; Wang, W.; Ye, N. Phylogeny and characterisation of Nannochloropsis oceanica var. sinensis var. nov. (Eustigmatophyceae), a new oleaginous alga from China. Phycologia 2013, 52, 573–577. [Google Scholar]
- Yamamoto, M.; Nozaki, H.; Kawano, S. Evolutionary relationships among multiple modes of cell division in the genus Nannochloris (Chlorophyta) revealed by genome size, actin gene multiplicity, and phylogeny. J. Phycol. 2001, 37, 106–120. [Google Scholar] [CrossRef]
- Suda, S.; Atsumi, M.; Miyashita, H. Taxonomic characterization of a marine Nannochloropsis species, N. oceanica sp. nov. (Eustigmatophyceae). Phycologia 2002, 41, 273–279. [Google Scholar] [CrossRef]
- Ma, X.-N.; Chen, T.-P.; Yang, B.; Liu, J.; Chen, F. Lipid production from Nannochloropsis. Mar. Drugs 2016, 14, 61. [Google Scholar] [CrossRef]
- Mata, T.M.; Martins, A.A.; Caetano, N.S. Microalgae for biodiesel production and other applications: A review. Renew. Sustain. Energy Rev. 2010, 14, 217–232. [Google Scholar] [CrossRef] [Green Version]
- Kumar, B.R.; Saravanan, S. Use of higher alcohol biofuels in diesel engines: A review. Renew. Sustain. Energy Rev. 2016, 60, 84–115. [Google Scholar] [CrossRef]
- Tracy, N.I.; Crunkleton, D.W.; Price, G.L. Gasoline production from phytol. Fuel 2010, 89, 3493–3497. [Google Scholar] [CrossRef]
- Abdel-Aal, E.I.; Haroon, A.M.; Mofeed, J. Successive solvent extraction and GC–MS analysis for the evaluation of the phytochemical constituents of the filamentous green alga Spirogyra longata. Egypt. J. Aquat. Res. 2015, 41, 233–246. [Google Scholar] [CrossRef]
- Abrantes, J.L.; Barbosa, J.; Cavalcanti, D.; Pereira, R.C.; Fontes, C.L.F.; Teixeira, V.L.; Souza, T.L.M.; Paixão, I.C. The Effects of the Diterpenes Isolated from the Brazilian Brown Algae Dictyota pfaffii and Dictyota menstrualis against the Herpes Simplex Type-1 Replicative Cycle; Georg Thieme Verlag: Stuttgart, Germany, 2010. [Google Scholar]
- Mewalal, R.; Rai, D.K.; Kainer, D.; Chen, F.; Külheim, C.; Peter, G.F.; Tuskan, G.A. Plant-derived terpenes: A feedstock for specialty biofuels. Trends Biotechnol. 2017, 35, 227–240. [Google Scholar] [CrossRef] [PubMed]
- Rossetti, I.; Gambaro, C.; Calemma, V. Hydrocracking of long chain linear paraffins. Chem. Eng. J. 2009, 154, 295–301. [Google Scholar] [CrossRef]
- Richmond, A. Handbook of Microalgal Culture: Biotechnology and Applied Phycology; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Mühling, M.; Fuller, N.J.; Millard, A.; Somerfield, P.J.; Marie, D.; Wilson, W.H.; Scanlan, D.J.; Post, A.F.; Joint, I.; Mann, N.H. Genetic diversity of marine Synechococcus and co-occurring cyanophage communities: Evidence for viral control of phytoplankton. Environ. Microbiol. 2005, 7, 499–508. [Google Scholar] [CrossRef]
- Ganuza, E.; Tonkovich, A.L. Method of Treating Bacterial Contamination in a Microalgae Culture with pH Shock. Google Patents US9181523B1, 10 November 2015. [Google Scholar]
- Stewart, W.D.P. Algal Physiology and Biochemistry; University of California Press: Berkeley, CA, USA, 1974; Volume 10. [Google Scholar]
- Holzinger, A.; Tschaikner, A.; Remias, D. Cytoarchitecture of the desiccation-tolerant green alga Zygogonium ericetorum. Protoplasma 2010, 243, 15–24. [Google Scholar] [CrossRef]
- Youssef, M.; Valdez-Ojeda, R.; Ku-Cauich, J.R.; Escobedo-Gracia Medrano, R. Enhanced protocol for isolation of plant genomic DNA. J. Agric. Environ. Sci. 2015, 4, 172–180. [Google Scholar] [CrossRef]
- Lee, S.-R.; Oak, J.H.; Chung, I.K.; Lee, J.A. Effective molecular examination of eukaryotic plankton species diversity in environmental seawater using environmental PCR, PCR-RFLP, and sequencing. J. Appl. Phycol. 2010, 22, 699–707. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Le Rouzic, B.; Bertru, G. Phytoplankton community growth in enrichment bioassays: Possible role of the nutrient intracellular pools. Acta Oecol. 1997, 18, 121–133. [Google Scholar] [CrossRef]
- Doan, T.T.Y.; Sivaloganathan, B.; Obbard, J.P. Screening of marine microalgae for biodiesel feedstock. Biomass Bioenergy 2011, 35, 2534–2544. [Google Scholar] [CrossRef]
- Demirbaş, A. Biodiesel fuels from vegetable oils via catalytic and non-catalytic supercritical alcohol transesterifications and other methods: A survey. Energy Conv. Manag. 2003, 44, 2093–2109. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Wahlen, B.D.; Willis, R.M.; Seefeldt, L.C. Biodiesel production by simultaneous extraction and conversion of total lipids from microalgae, cyanobacteria, and wild mixed-cultures. Biores. Technol. 2011, 102, 2724–2730. [Google Scholar] [CrossRef]
- Volkman, J.K.; Jeffrey, S.W.; Nichols, P.D.; Rogers, G.I.; Garland, C.D. Fatty acid and lipid composition of 10 species of microalgae used in mariculture. J. Exp. Mar. Biol. Ecol. 1989, 128, 219–240. [Google Scholar] [CrossRef]
- Iverson, S.J.; Lang, S.L.C.; Cooper, M.H. Comparison of the bligh and dyer and folch methods for total lipid determination in a broad range of marine tissue. Lipids 2001, 36, 1283–1287. [Google Scholar] [CrossRef]
- Kang, W.; Ji, Z.; Wang, J. Composition of the essential oil of Adiantum flabellulatum. Chem. Nat. Compd. 2009, 45, 575–577. [Google Scholar] [CrossRef]
- Poirier, M.-A. Stable Alkoxylated Fatty Acid Alkyl Esters from Transesterification-Alkoxylation of Vegetable Oils. Google Patents US8523960B2, 3 September 2013. [Google Scholar]
- Araujo, G.S.; Matos, L.J.; Fernandes, J.O.; Cartaxo, S.J.; Goncalves, L.R.; Fernandes, F.A.; Farias, W.R. Extraction of lipids from microalgae by ultrasound application: Prospection of the optimal extraction method. Ultrason. Sonochem. 2013, 20, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Ganuza, E.; Sellers, C.E.; Bennett, B.W.; Lyons, E.M.; Carney, L.T. A novel treatment protects chlorella at commercial scale from the predatory bacterium Vampirovibrio chlorellavorus. Front. Microbiol. 2016, 7, 848. [Google Scholar] [CrossRef] [PubMed]
- Vidyashankar, S.; VenuGopal, K.S.; Swarnalatha, G.V.; Kavitha, M.D.; Chauhan, V.S.; Ravi, R.; Bansal, A.K.; Singh, R.; Pande, A.; Ravishankar, G.A. Characterization of fatty acids and hydrocarbons of chlorophycean microalgae towards their use as biofuel source. Biomass Bioenergy 2015, 77, 75–91. [Google Scholar] [CrossRef]
- Ramos-Higuera, E.; Alcocer, J.; Ortega-Mayagoitia, E.; Camacho, A. Nitrógeno: Elemento limitante para el crecimiento fitoplanctónico en un lago oligotrófico tropical. Hidrobiológica 2008, 18, 105–113. [Google Scholar]
- Chu, S. The influence of the mineral composition of the medium on the growth of planktonic algae: Part I. Methods and culture media. J. Ecol. 1942, 284–325. [Google Scholar] [CrossRef]
- Abirami, S.; Murugesan, S.; Sivamurugan, V.; Sivaswamy, S.N. Screening and optimization of culture conditions of Nannochloropsis gaditana for omega 3 fatty acid production. J. Appl. Biol. Biotechnol. 2017, 5, 13–17. [Google Scholar]
- Santhanam, P.; Kumar, S.D.; Ananth, S.; Jeyanthi, S.; Sasirekha, R.; Premkumar, C. Effect of culture conditions on growth and lipid content of marine microalga Nannochloropsis sp. strain (PSDK11). Indian J. Geo-Mar. Sci. 2017, 46, 2332–2338. [Google Scholar]
- Van den Hoek, C.; Stam, W.; Olsen, J. The emergence of a new chlorophytan system, and Dr. Kornmann’s contribution thereto. Helgoländer Meeresuntersuchungen 1988, 42, 339–383. [Google Scholar] [CrossRef]
- Shubert, E.; Gärtner, G. Nonmotile coccoid and colonial green algae. In Freshwater Algae of North America, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 315–373. [Google Scholar]
- Lubián, L.M. Nannochloropsis gaditana sp. nov., una nueva Eustigmatophyceae marina. Lazaroa 1982, 4, 287–293. [Google Scholar]
- Henley, W.J.; Hironaka, J.L.; Guillou, L.; Buchheim, M.A.; Buchheim, J.A.; Fawley, M.W.; Fawley, K.P. Phylogenetic analysis of the ‘Nannochloris-like’ algae and diagnoses of Picochlorum oklahomensis gen. et sp. nov. (Trebouxiophyceae, Chlorophyta). Phycologia 2004, 43, 641–652. [Google Scholar] [CrossRef]
- Fawley, K.P.; Fawley, M.W. Observations on the diversity and ecology of freshwater Nannochloropsis (Eustigmatophyceae), with descriptions of new taxa. Protist 2007, 158, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Antia, N.J.; Cheng, J.Y. The keto-carotenoids of two marine coccoid members of the Eustigmatophyceae. Br. Phycol. J. 1982, 17, 39–50. [Google Scholar] [CrossRef] [Green Version]
- South, G.; Whittick, A. Physiology and biochemistry. In An Introduction to Phycology; Blackwell Scientific Publications: Oxford, UK, 1987. [Google Scholar]
- Liu, X.; Zhu, H.; Liu, B.; Liu, G.; Hu, Z. Phylogeny and morphology of genus Nephrocytium (Sphaeropleales, Chlorophyceae, Chlorophyta) from China. Phytotaxa 2017, 319, 84–92. [Google Scholar] [CrossRef]
- Komárek, J. Änderungen in der taxonomie der chlorokokkalalgen. Algolog. Stud. /Arch. Hydrobiol. 1979, 24, 239–263. [Google Scholar]
- Volkman, J.; Gllan, F.; Johns, R.; Eglinton, G. Sources of neutral lipids in a temperate intertidal sediment. Geochim. Cosmochim. Acta 1981, 45, 1817–1828. [Google Scholar] [CrossRef]
- Yao, L.; Gerde, J.A.; Lee, S.-L.; Wang, T.; Harrata, K.A. Microalgae lipid characterization. J. Agric. Food Chem. 2015, 63, 1773–1787. [Google Scholar] [CrossRef] [PubMed]
- Giakoumis, E.G. A statistical investigation of biodiesel physical and chemical properties, and their correlation with the degree of unsaturation. Renew. Energy 2013, 50, 858–878. [Google Scholar] [CrossRef]
- Bondioli, P.; Della Bella, L.; Rivolta, G.; Chini Zittelli, G.; Bassi, N.; Rodolfi, L.; Casini, D.; Prussi, M.; Chiaramonti, D.; Tredici, M.R. Oil production by the marine microalgae Nannochloropsis sp. F&M-M24 and Tetraselmis suecica F&M-M33. Biores. Technol. 2012, 114, 567–572. [Google Scholar] [CrossRef]
- El Shoubaky, G.A.; Salem, E.A. Terpenes and sterols composition of marine brown algae Padina pavonica (Dictyotales) and Hormophysa triquetra (Fucales). Int. J. Pharmacogn. Phytochem. Res. 2014, 6, 894–900. [Google Scholar]
- Talebi, A.F.; Mohtashami, S.K.; Tabatabaei, M.; Tohidfar, M.; Bagheri, A.; Zeinalabedini, M.; Hadavand Mirzaei, H.; Mirzajanzadeh, M.; Malekzadeh Shafaroudi, S.; Bakhtiari, S. Fatty acids profiling: A selective criterion for screening microalgae strains for biodiesel production. Algal Res. 2013, 2, 258–267. [Google Scholar] [CrossRef]
- Abomohra, A.E.-F.; El-Sheekh, M.; Hanelt, D. Screening of marine microalgae isolated from the hypersaline Bardawil lagoon for biodiesel feedstock. Renew. Energy 2017, 101, 1266–1272. [Google Scholar] [CrossRef]
- Stevenson, R.J.; Bothwell, M.L.; Lowe, R.L.; Thorp, J.H. Algal Ecology: Freshwater Benthic Ecosystem; Academic Press: Cambridge, MA, USA, 1996. [Google Scholar]
- Patterson, G.W.; Krauss, R.W. Sterols of Chlorella. I. The naturally occurring sterols of Chlorella vulgaris, C. ellipsoidea, and C. saccharophila. Plant Cell Physiol. 1965, 6, 211–220. [Google Scholar] [CrossRef]
- Orcutt, D.; Patterson, G. Sterol, fatty acid and elemental composition of diatoms grown in chemically defined media. Comp. Biochem. Physiol. Part B Comp. Biochem. 1975, 50, 579–583. [Google Scholar] [CrossRef]
- Valdez-Ojeda, R.; González-Muñoz, M.; Us-Vázquez, R.; Narváez-Zapata, J.; Chavarria-Hernandez, J.C.; López-Adrián, S.; Barahona-Pérez, F.; Toledano-Thompson, T.; Garduño-Solórzano, G.; Medrano, R.M.E.-G. Characterization of five fresh water microalgae with potential for biodiesel production. Algal Res. 2015, 7, 33–44. [Google Scholar] [CrossRef]
- Serrano, M.; Oliveros, R.; Sánchez, M.; Moraschini, A.; Martínez, M.; Aracil, J. Influence of blending vegetable oil methyl esters on biodiesel fuel properties: Oxidative stability and cold flow properties. Energy 2014, 65, 109–115. [Google Scholar] [CrossRef]
- Altaie, M.A.H.; Janius, R.B.; Taufiq-Yap, Y.H.; Rashid, U. Basic properties of methyl palmitate-diesel blends. Fuel 2017, 193, 1–6. [Google Scholar] [CrossRef]
- Batts, B.; Fathoni, A.Z. A literature review on fuel stability studies with particular emphasis on diesel oil. Energy Fuels 1991, 5, 2–21. [Google Scholar] [CrossRef]
- Fajar, R.; Sugiarto, B. Predicting fuel properties of partially hydrogenated jatropha methyl esters used for biodiesel formulation to meet the fuel specification of automobile and engine manufacturers. Kasetsart J. Nat. Sci. 2012, 46, 629–637. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fraction | % of Total Extract | ||
|---|---|---|---|
| NSRE-1 | NRRE-1 | NSRE-2 | |
| 1 | 5.8 | 8.1 | 20.1 |
| 2 | 8.9 | 10.7 | 7.1 |
| 3 | 8.5 | 6 | 23.1 |
| 4 | 5.9 | 16.4 | 19.6 |
| 5 | - | 10 | 8.4 |
| 6 | 71 | 48.8 | 21.7 |
| Compound (IUPAC) | Molecular Formula | % of Total Peak Area (wt % Fractions) | ||
|---|---|---|---|---|
| NSRE-1 | NRRE-1 | NSRE-2 | ||
| Hydrocarbons | ||||
| Tetradecane | C14H36 | 5.64 (3.6) | ||
| Pentadecane | C15H32 | 4.89 (8.3) | 4.35 (4.5) | |
| 1-Heptadecene | C17H34 | 23.75 (59.2) | ||
| Heptadecane | C17H36 | 6.91 (11.8) | 8.48 (5.5) | 5.97 (6.1) |
| Octadecane | C18H38 | 4.86 (8.3) | 5.72 (5.9) | |
| Nonadecane | C19H40 | 4.66 (3.0) | 4.3 (1.1) | |
| Eicosane | C20H42 | 7.8 (5.0) | 8.78 | |
| Heneicosane | C21H44 | 7.28 (4.7) | 8.31 (2.1) | |
| Tetracosane | C24H50 | 3.95 (6.7) | 7.76 (2.0) | |
| Hexacosane | C26H54 | 4.86 (5.0) | ||
| Heptacosane | C27H56 | 3.37 (5.7) | ||
| Neophytadiene | C20H38 | 64.22 (78.2) | 64.98 (61.8) | |
| Total (wt %) | 33.8 | 59.9 | 46.9 | |
| FAMEs (TAG and FFA fractions) | ||||
| Hexadecanoic acid, methyl ester (Methyl palmitate, C16:0) | C17H34O2 | 13.58 (4.16) | 13.85 (28.48) | 7.82 (10.82) |
| 9,12-Octadecadienoic acid (Z,Z)-, methyl ester (Methyl linoleate, C18:2) | C19H34O2 | 4.92 (10.11) | 12.34 (26.35) | |
| Octadecenoic acid. Methyl stearate (C18:0) | 5.4 (10.4) | |||
| (Z)-9-Octadecenoic acid, methyl ester (Methyl oleate, C18:1) | C19H36O2 | 7.37 (15.15) | ||
| Total (wt %) | 9.5 | 24.7 | 23 | |
| Oxygenated compounds | ||||
| 3-Hydroxy-butanoic acid | C4H8O3 | 10.78 (2.2) | ||
| Bis (2-ethylhexyl) hexadecanoic acid ester | C22H42O4 | 45.19 (12.9) | 4.93 (9.14) | 39.03 (5.6) |
| Phytol | C20H40O | 3.02 (10.0) | ||
| 6,10,14-Trimethyl-2-pentadecanone | C18H36O | 10.54 (2.7) | ||
| 2-Palmitoylglycerol | C19H38O4 | 9.94 (10.19) | ||
| Campesterol | C28H48O | 6.58 (12.20) | ||
| Clionasterol | C29H50O | 26.79 (49.68) | 8.25 (2.4) | |
| (Z)-13-Docosenamide | C22H43NO | 61.35 (40.88) | ||
| 2,2,4-Trimethyl-1,3-pentanediol 1-isobutyrate | C12H24O3 | 5.01 (9.24) | ||
| 2,4-Di-t-butylphenol | C14H22O | 5.73 (18.97) | ||
| Total (wt %) | 56.8 | 15.3 | 30.1 | |
| % oxygen | 19.2 | 6 | 12.5 | |
| H/C atomic ratio | 1.9 | 1.7 | 1.9 | |
| Compound (IUPAC) | Molecular Formula | % of Total Peak Area |
|---|---|---|
| Dodecane | C12H26 | 82.9 |
| 2,6,10-Trimethyldodecane | C15H32 | 1.8 |
| Hexadecane | C16H34 | 1.8 |
| Heptadecane | C17H36 | 5.9 |
| Octadecane | C18H38 | 4.8 |
| 10-Methylnonadecane | C20H46 | 2.7 |
| Property (Units) | NSRE-1 | NRRE-1 | NSRE-2 | ASTM D6751 | EN 14214 |
|---|---|---|---|---|---|
| ρ @15°C (kg/m3) | 886 a | 880.4 | 883.0 | 860–900 | |
| ρ @40°C (kg/m3) | 850.8 | 855.5 | 856.3 | ||
| CN | 74.5 | 66.6 | 65.9 | 47 min | 51 min |
| ν @40°C (mm2/s) | 4.7 a | 4.5 | 4.2 | 1.9–6.0 | 3.5–5.0 |
| HHV (MJ/kg) | 38.4 a | 39.7 | 39.5 | ||
| IV (g I/100 g biodiesel) | 0.0 | 44.6 | 45.4 | 120 max | |
| CFPP (°C) | 31.6 | 17.9 | 23.4 | ||
| OSI (h) | 27.4 a | 5.4 | 7.1 | 3 h | 6 h |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Rosales, A.R.; Ancona-Canché, K.; Chavarria-Hernandez, J.C.; Barahona-Pérez, F.; Toledano-Thompson, T.; Garduño-Solórzano, G.; López-Adrian, S.; Canto-Canché, B.; Polanco-Lugo, E.; Valdez-Ojeda, R. Fatty Acids, Hydrocarbons and Terpenes of Nannochloropsis and Nannochloris Isolates with Potential for Biofuel Production. Energies 2019, 12, 130. https://doi.org/10.3390/en12010130
López-Rosales AR, Ancona-Canché K, Chavarria-Hernandez JC, Barahona-Pérez F, Toledano-Thompson T, Garduño-Solórzano G, López-Adrian S, Canto-Canché B, Polanco-Lugo E, Valdez-Ojeda R. Fatty Acids, Hydrocarbons and Terpenes of Nannochloropsis and Nannochloris Isolates with Potential for Biofuel Production. Energies. 2019; 12(1):130. https://doi.org/10.3390/en12010130
Chicago/Turabian StyleLópez-Rosales, Alan Rodrigo, Katia Ancona-Canché, Juan Carlos Chavarria-Hernandez, Felipe Barahona-Pérez, Tanit Toledano-Thompson, Gloria Garduño-Solórzano, Silvia López-Adrian, Blondy Canto-Canché, Erik Polanco-Lugo, and Ruby Valdez-Ojeda. 2019. "Fatty Acids, Hydrocarbons and Terpenes of Nannochloropsis and Nannochloris Isolates with Potential for Biofuel Production" Energies 12, no. 1: 130. https://doi.org/10.3390/en12010130
APA StyleLópez-Rosales, A. R., Ancona-Canché, K., Chavarria-Hernandez, J. C., Barahona-Pérez, F., Toledano-Thompson, T., Garduño-Solórzano, G., López-Adrian, S., Canto-Canché, B., Polanco-Lugo, E., & Valdez-Ojeda, R. (2019). Fatty Acids, Hydrocarbons and Terpenes of Nannochloropsis and Nannochloris Isolates with Potential for Biofuel Production. Energies, 12(1), 130. https://doi.org/10.3390/en12010130