3.1. Isolation and Selection of Thermotolerant Yeast Strains
Natural habitats are a major source of useful microorganisms for biorefinery production [
36]. In this study, soil and plant samples from different locations in Northeastern Thailand were collected, and the isolation of thermotolerant yeasts was performed using the enrichment culture technique as described by Limtong
et al. [
13]. The utilization of this technique under selection pressures has been widely used to isolate several thermotolerant yeast strains [
13,
17]. It has been reported that the effects of high temperatures on microbial growth have often been aggravated by ethanol concentrations greater than 3% [
37]. Therefore, ethanol at 4% (
v/
v) and an incubation temperature of 35 °C served as selection pressures for the isolation of thermotolerant yeast strains in this work. As a result, sixty-two yeast isolates were obtained, and most of these yeasts were from soil samples. Their ability to grow and produce ethanol at high temperatures were tested by culturing the yeast strains in SSJ incubated at temperatures between 30 and 50 °C. The results showed that nine isolates of yeast, which were designated as DBKKU Y-53, DBKKU Y-55, DBKKU Y-58, DBKKU Y-102, DBKKU Y-103, DBKKU Y-104, DBKKU Y-105, DBKKU Y-106, and DBKKU Y-107, were able to grow and produce a relatively high level of ethanol at 45 °C. Of these, six isolates (
i.e., DBKKUY-102, DBKKUY-103, DBKKUY-104, DBKKUY-105, DBKKUY-106, DBKKUY-107) were capable of growth and ethanol production at 47 °C. Their ability to grow and produce ethanol at high temperatures were comparable to that of
K. marxianus ATCC 8554, which is known as a thermotolerant yeast. Based on growth performance of the isolated yeast strains at high temperatures (above 40 °C), we speculated that these nine isolates of yeast were thermotolerant yeasts [
38]. Therefore, all nine yeast isolates were selected for further study.
Successful isolation and selection of thermotolerant yeasts for ethanol production at high temperatures has been reported by several investigators. According to our literature review, SSJ has never been used directly as the substrate for yeast selection. Almost all previous research studies used glucose as a substrate, e.g., Ballesteros
et al. [
39] reported the screening and selection of thermotolerant yeast strains using glucose as the sole carbon source and found that
K. marxianus L.G. was the most effective strain for ethanol production at high temperatures. This strain produced an ethanol concentration of approximately 3.76% (
w/
v) when the fermentation was carried out at 42 °C. Banat
et al. [
40] isolated and selected the thermotolerant yeast
K. marxianus using an enrichment method with glucose as the carbon source and demonstrated that the selected
K. marxianus strain produced an ethanol concentration of approximately 5.6%–6.0% (
w/
v) at 45–50 °C when using glucose as the substrate and ethanol concentration of approximately 7.5%–8.0% (
w/
v) and 6.5%–7.0% (
w/
v) at 37 °C and 40 °C, respectively, when using molasses as a substrate. Kiran Sree
et al. [
12] screened thermotolerant yeast
S. cerevisiae VS3 using glucose as the sole carbon source, and they reported that the selected yeast strain produced an ethanol concentration of approximately 75 g·L
−1 from a glucose concentration of 150 g·L
−1 at 40 °C. Edgardo
et al. [
19] used glucose for the selection of thermotolerant yeast strains and obtained a good potential thermotolerant yeast
S. cerevisiae that was able to produce approximately 75% of the theoretical ethanol yield at 40 °C. In addition to using glucose as a substrate, other carbon sources, such as sugarcane juice [
13], xylose [
41], sugarcane blackstrap molasses [
42], and inulin [
43] have also been used for screening and selecting thermotolerant yeasts.
3.2. Identification of the Selected Thermotolerant Yeasts
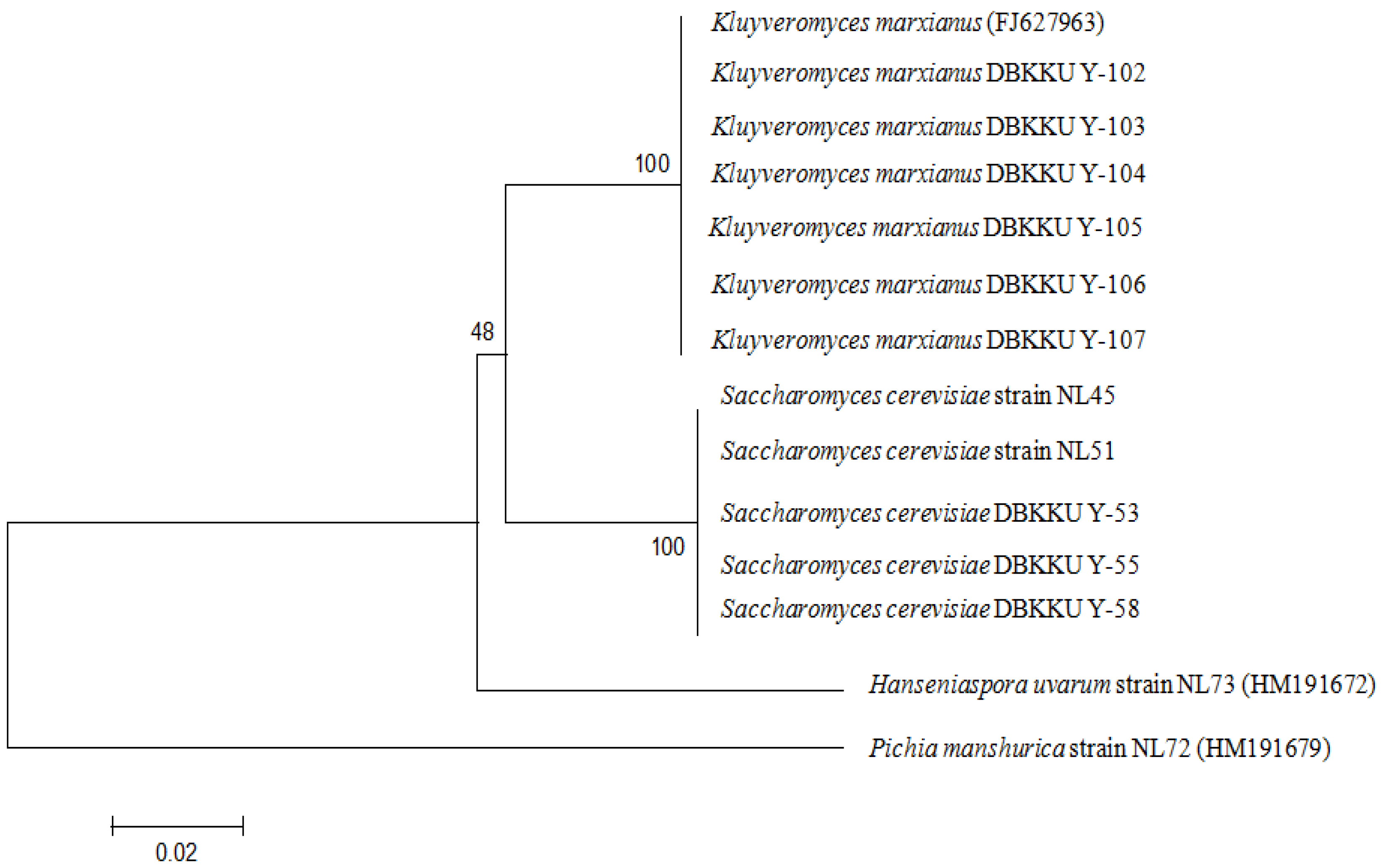
Based on the morphological and physiological characteristics, three isolated yeasts (
i.e., DBKKU Y-53, DBKKU Y-55, DBKKU Y-58) were found to be
Saccharomyces, whereas six isolated yeasts (
i.e., DBKKU Y-102, DBKKU Y-103, DBKKU Y-104, DBKKU Y-105, DBKKU Y-106, DBKKU Y-107) were revealed to be
Kluyveromyces. To confirm this finding, molecular taxonomic analyses based on the nucleotide sequences of the D1/D2 domain of the 26S rDNA gene were carried out. As a result, the nucleotide sequences from the yeast strains DBKKU Y-53, DBKKU Y-55, DBKKU Y-58 and from
S. cerevisiae (NL45 and NL51) were 99% identical, whereas strains DBKKU Y-102, DBKKU Y-103, DBKKU Y-104, DBKKU Y-105, DBKKU Y-106, DBKKU Y-107, and
K. marxianus (FJ627963) were 99% identical. Phylogenetic analysis confirmed that strains DBKKU Y-53, DBKKU Y-55, and DBKKU Y-58 were clustered in the same group as
S. cerevisiae whereas strains DBKKU Y-102, DBKKU Y-103, DBKKU Y-104, DBKKU Y-105, DBKKU Y-106, and DBKKU Y-107 were closely related to
K. marxianus (
Figure 1). Therefore, strains DBKKU Y-53, DBKKU Y-55, and DBKKU Y-58 were identified as
S. cerevisiae, whereas the other strains were
K. marxianus.
3.4. Factors Influencing Ethanol Production When Using Thermotolerant Yeast S. cerevisiae DBKKU Y-53
There are several factors influencing the growth and ethanol production of yeast, such as the incubation temperature, the pH of the fermentation medium, the cell concentration, the sugar concentration, the nitrogen sources, and the aeration [
13,
44]. Therefore, the effects of some of the major factors on ethanol production using SSJ by thermotolerant yeast
S. cerevisiae DBKKU Y-53 were investigated. In this work, the ethanol production efficiency of
S. cerevisiae DBKKU Y-53 was compared with that of
S. cerevisiae SC90, which is one of the industrial yeast strains widely used to produce ethanol on a commercial scale in Thailand.
The effect of temperature on ethanol production from SSJ using
S. cerevisiae DBKKU Y-53 and
S. cerevisiae SC90 was analyzed, and the results are summarized in
Table 3. There was no significantly difference in ethanol concentration produced by both strains at 30 °C. However, at 37 °C and 40 °C,
S. cerevisiae DBKKU Y-53 produced greater ethanol concentrations than
S. cerevisiae SC09. The maximum ethanol concentrations produced by
S. cerevisiae DBKKU Y-53 at 37 °C and 40 °C were 71.73 ± 2.62 g·L
−1 and 58.14 ± 7.71 g·L
−1, respectively. When the incubation temperature was shifted from 40 °C to 42 °C and 45 °C, ethanol concentrations and volumetric ethanol productivities produced by both strains were remarkably decreased. This might be due to the negative effect of high temperature on growth and metabolic processes in yeast cells. It has been reported that high temperature causes a modification of plasma membrane fluidity and a reduction in the effectiveness of the plasma membrane as a semipermeable barrier allowing leakage of essential cofactors and coenzymes required for the activity of enzymes involved in glucose metabolism and ethanol production [
44]. Roukas [
45] reported that high temperatures caused denaturation of cellular proteins, which resulted in the reduction of cell growth and fermentation activity. Moreover, the reduction in yeast growth at high temperatures was also due to the accumulation of intracellular ethanol, which modifies the cell membrane structure of yeast cell [
46]. Our results were in good agreement with Banat
et al. [
40], who observed a reduction in ethanol concentration produced by the thermotolerant yeast
K. marxianus at 40 °C. Kiran Sree
et al. [
12] reported the ethanol production from glucose (150 g·L
−1) using thermotolerant yeast
S. cerevisiae VS3 and observed a reduction in ethanol concentration when the incubation temperature shifted from 30 °C to 40 °C. Limtong
et al. [
13] demonstrated that the ethanol concentration produced by the newly isolated thermotolerant yeast
K. marxianus DMKU3-1042 decreased significantly when the incubation temperature was increased from 37 °C to 40 °C. Tofighi
et al. [
47] reported the reduction in cell mass productivity and ethanol fermentation ability of the thermotolerant yeast
S. cerevisiae when the incubation temperature increased to greater than 40 °C. Recently, Charoensopharat
et al. [
43] found that the ethanol concentration produced by the newly isolated thermotolerant yeast
K. marxianus decreased dramatically when ethanol fermentation was carried out at temperatures greater than 40 °C.
The pH of the medium is an important factor influencing ethanol yield. Generally, the optimum pH for yeast growth and ethanol production is in the range of 4.0 to 6.0 depending on growth conditions, such as the temperature, the presence of oxygen, the yeast species, and the type of raw material. For instance, the optimum pH for ethanol production from sugarcane juice at high temperatures using the thermotolerant yeast
K. marxianus DMKU 3-1042 was 5.0 [
13]. During ethanol fermentation, the pH of the fermentation medium is almost in the range of 4.0 to 5.5. This pH level typically prevents bacterial contamination during the fermentation process [
48]. In this study, the effect of pH on ethanol production by the thermotolerant yeast
S. cerevisiae DBKKU Y-53 and a reference strain was investigated, and the results are summarized in
Table 4 and
Table 5. The optimum pH for ethanol production from SSJ by both strains at 37 °C and 40 °C was 5.5, which is in agreement with reports by Ercan
et al. [
49] and Sign and Bishnoi [
50]. At a pH less than or greater than 5.5, ethanol concentrations and volumetric ethanol productivities tended to decrease. This might be related to the activity of enzymes involved in the ethanol production pathway. It has been reported that enzymes may be inactivated at a pH level that is less than or greater than the optimum value causing a reduction in cell growth and ethanol fermentation ability [
48]. The initial pH of the SSJ was in the range of 5.2–5.5. Therefore, a pH of 5.5, which gave the highest ethanol concentration, was selected for further study
The initial cell concentration affects not only the ethanol yield but also the substrate consumption rate and ethanol fermentation rate. Generally, high initial cell concentrations reduce the lag phase of growth and increase the sugar consumption and ethanol fermentation rate, leading to a high ethanol yield and productivity. In this study, the effect of initial cell concentrations (1 × 10
6, 1 × 10
7, 1 × 10
8 cells·mL
−1) on ethanol fermentation from SSJ containing 220 g·L
−1 of total sugars was investigated, and the results are summarized in
Table 4 and
Table 5. As a result, increasing the cell concentration resulted in an increase in the ethanol concentration and the volumetric ethanol productivity. The maximum ethanol concentrations and volumetric ethanol productivities produced by
S. cerevisiae DBKKU Y-53 and SC09 at 37 °C and 40 °C were achieved at an initial cell concentration of 1 × 10
8 cells·mL
−1, which is in good agreement with Charoensopharat
et al. [
43] and Laopaiboon
et al. [
51].
High sugar concentrations (more than 20%
w/
v) are not often used in industrial ethanol production because they may reduce the yeast cell viability, the substrate conversion rate, and the ethanol yield [
52,
53]. However, ethanol production with high sugar concentrations have also been reported, and the fermentation efficiencies vary depending on the yeast species and fermentation conditions. For example, Laopaiboon
et al. [
54] reported a maximum ethanol concentration of 120.68 g·L
−1 and volumetric ethanol productivity of 2.01 g·L
−1·h
−1 when
S. cerevisiae NP01 was used to produce ethanol from SSJ under a very high gravity fermentation. Charoensopharat
et al. [
43] reported a maximum ethanol concentration of 104.83 g·L
−1 and a volumetric ethanol productivity of 4.37 g·L
−1·h
−1 when
K. marxianus DBKKU Y-102 was used to produce ethanol from Jerusalem artichoke tubers at 37 °C during consolidated bioprocessing. To verify the effect of sugar concentration on ethanol production efficiency of the thermotolerant yeast
S. cerevisiae DBKKU Y-53, SSJ containing various sugar concentrations (200, 250, 300 g·L
−1) was tested. As shown in
Table 4 and
Table 5, increasing in the sugar concentration from 200 g·L
−1 to 250 g·L
−1 resulted in an increase in the ethanol concentration. However, at a sugar concentration of 300 g·L
−1 the ethanol concentration was remarkably decreased, and a large amount of sugar remained in the fermentation broth at both 37 °C and 40 °C. High sugar concentrations have been reported to cause negative effects on cell viability and morphology due to an increase in the osmotic pressure, which leads to a reduction in the cell biomass and ethanol yield [
53,
55]. The maximum ethanol concentrations produced by
S. cerevisiae DBKKU Y-53 and SC90 were achieved at a sugar concentration of 250 g·L
−1. Therefore, this sugar concentration was used for subsequent experiments.
Approximately 10% of yeast’s dry weight is nitrogen [
44]. Therefore, nitrogen is one of the important constituents for cell growth and synthesis of structural and functional proteins involved in metabolic processes. Various types of nitrogen sources have been used to supplement ethanol fermentation medium both in organic (yeast extract, corn steep liquor) and inorganic forms (ammonium sulfate, ammonium nitrate, urea, diammonium phosphate). In this study, the effects on ethanol production of using yeast extract, urea, and ammonium sulfate at different concentrations from SSJ by
S. cerevisiae DBKKU Y-53 and SC90 were determined, and the results are summarized in
Table 6 and
Table 7. Supplementation of urea in the SSJ was shown to significantly enhance the ethanol production by both strains of yeast at 37 °C and 40 °C, which is in good agreement with reports from Yue
et al. [
56]. There was no significant difference in ethanol production at 37 °C when comparing yeast extract supplementation and supplementation-free fermentations. However, at 40 °C, supplementation of yeast extract tended to result in a lower ethanol concentration compared with the control condition. Yeast extract has been reported to be a good organic nitrogen source for ethanol production using
S. cerevisiae. However, its availability to be utilized by yeast cells is depended on the fermentation conditions. For example, Laopaiboon
et al. [
54] reported that supplementation of yeast extract in SSJ improved the ethanol production by
S. cerevisiae NP01 for very high gravity fermentation. With respect to ammonium sulfate, supplementation of this nitrogen compound resulted in a reduction of the ethanol concentration in all conditions tested compared with the control without nitrogen supplementation. These results clearly indicate that ammonium sulfate was not a good nitrogen source for ethanol production at high temperatures using
S. cerevisiae DBKKU Y-53 and SC90 when SSJ was used as a substrate. One possibility is that this nitrogen compound was not taken up by the yeast cells during high temperature fermentation. Ter Schure
et al. [
57] and Magasanik and Kaiser [
58] reported that the uptake of nitrogen by yeast cells is regulated by the mechanism known as nitrogen catabolite repression (NCR). NCR enables yeast cells to select the best nitrogen sources by repressing the transcription of genes involved in the utilization of the poorer nitrogen [
59]. However, the effectiveness of NCR mechanism is influenced by many factors including fermentation temperature and the presence of ethanol. Normally, high temperatures and high ethanol concentrations cause the modification of the plasma membrane. Therefore, the nitrogen sources sensing system, which is mainly located in the plasma membrane of yeast cells, may be affected by these stress conditions, leading to the impairment of ammonium sulfate uptake [
60,
61]. In this study, urea proved to be the best nitrogen source for ethanol production from SSJ during high temperature fermentation using
S. cerevisiae DBKKU Y-53 and SC90; therefore, it was chosen for further analysis.
3.5. Ethanol Production in 2-L Bioreactor
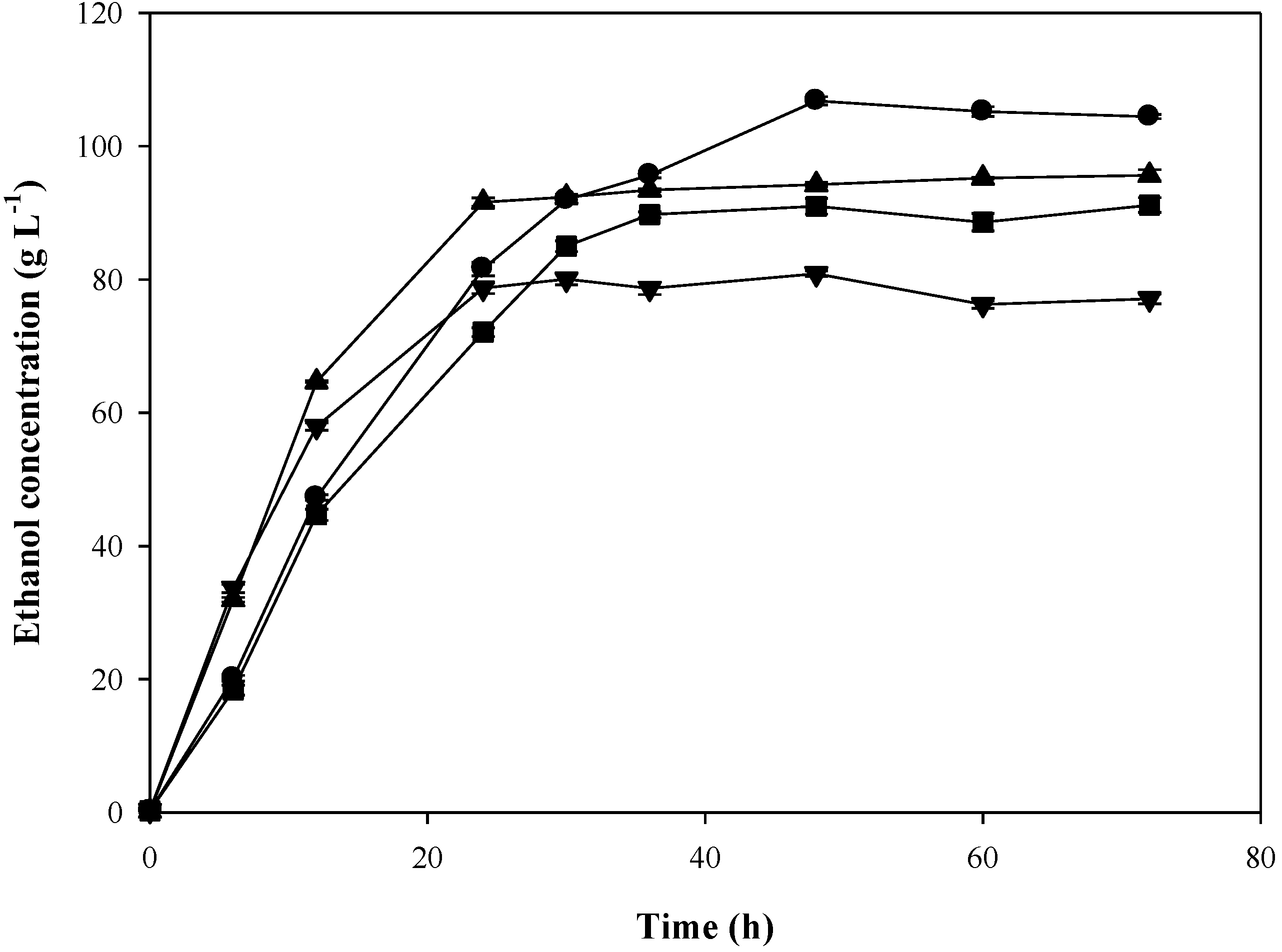
The time profiles of ethanol production from SSJ using
S. cerevisiae DBKKU Y-53 and SC90 at 37 °C and 40 °C in a 2-L bioreactor are illustrated in
Figure 2. During the first 12 h after fermentation, ethanol concentrations produced by both strains sharply increased and reached their maximum values at 24 h for
S. cerevisiae SC90 and 48 h for
S. cerevisiae DBKKU Y-53.
Table 8 summarizes the kinetic parameters of ethanol production from the SSJ at high temperatures using
S. cerevisiae DBKKU Y-53 and SC90. It can be seen from this finding that the newly isolated thermotolerant yeast
S. cerevisiae DBKKU Y-53 resulted in a greater ethanol concentration as well as sugar utilization capability compared with SC90.
A comparative analysis of the ethanol production from SSJ using the thermotolerant yeast
S. cerevisiae DBKKU Y-53 with values reported in the literatures using different raw materials and yeast strains was performed, and the results are summarized in
Table 9. The ethanol concentration and volumetric ethanol productivity produced by
S. cerevisiae DBKKU Y-53 were greater than values reported in other works, suggesting that the newly isolated thermotolerant yeast
S. cerevisiae DBKKU Y-53 was a good candidate for ethanol production from SSJ at high temperatures. Although several isolates of the thermotolerant yeast
K. marxianus have been reported to be capable of growth and ethanol production at temperatures greater than 45 °C, almost all of these isolates had relatively lower ethanol yields and were less tolerant to ethanol than
S. cerevisiae. Furthermore, these isolates also required oxygen during ethanol fermentation resulting in an increase in the operating cost. Therefore, the thermotolerant yeast
S. cerevisiae is more promising for ethanol production at high temperatures on a commercial scale compared with
K. marxianus.
This work will contribute a significant amount of information on ethanol production at an industrial scale. However, the ethanol production cost at a large-scale should be concerned. There are many techniques which can be employed to reduce the production cost; for example, cell recycling, various fermentation approaches, such as very high gravity fermentation, consolidated bioprocessing, and continuous ethanol fermentation using stirred tank bioreactor coupling with plug flow bioreactor [
17,
18,
43,
54].
3.6. Real-Time RT-PCR Analysis of Gene Expression
Stressful conditions during ethanol fermentation, such as high temperatures, and high concentrations of ethanol or sugar, have been reported to trigger the expression of several stress-responsive genes including those encoding HSPs, enzymes involved in protein degradation and in the glycolysis pathway, and other proteins involved in the synthesis of compatible solutes and reserve carbohydrates [
22,
23,
25,
66,
67]. In the present study, the expression levels of
hsp26,
hsp70,
hsp90,
hsp104,
cdc,
tps,
nth,
gsy, and
rsp in the thermotolerant yeast
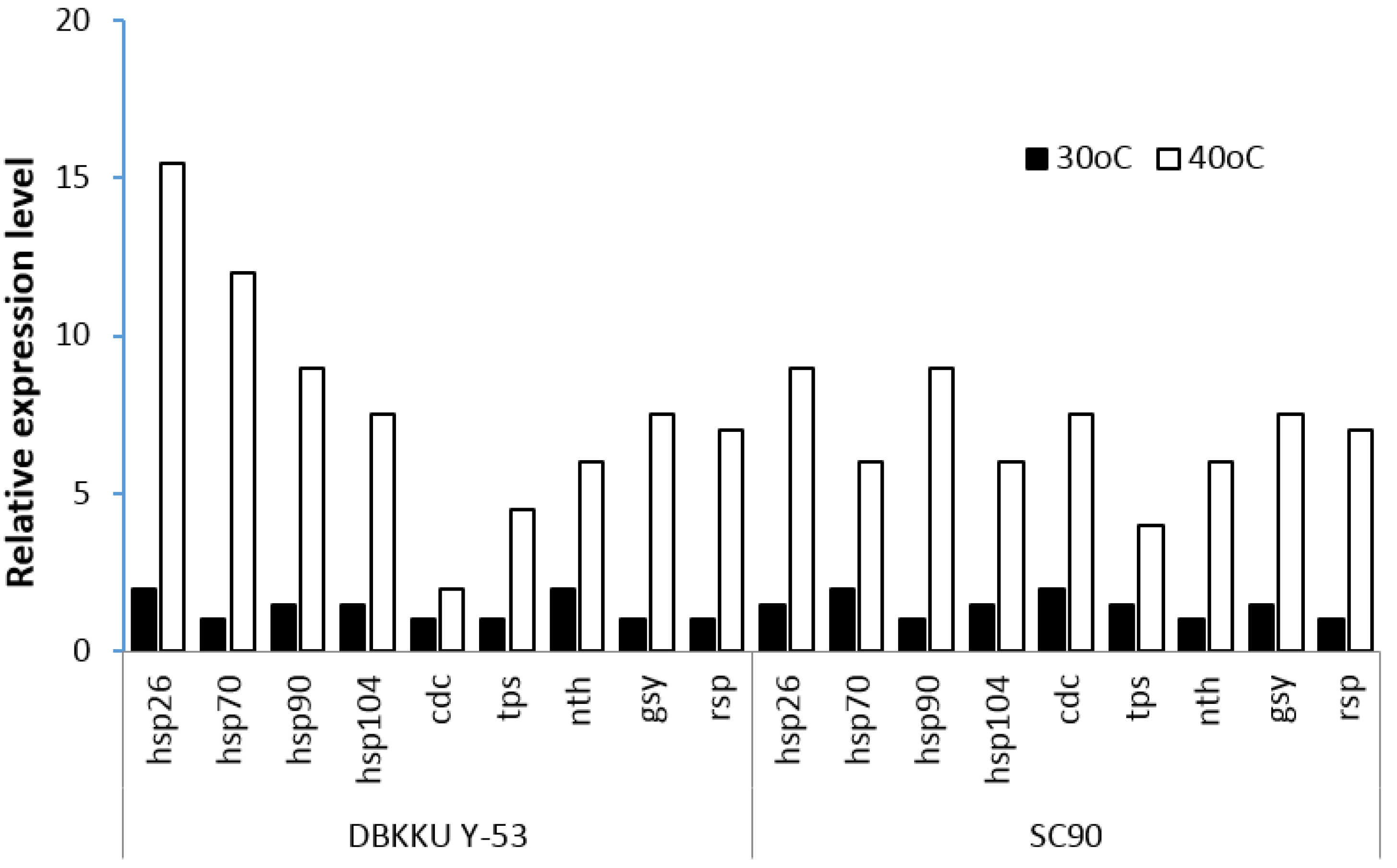
S. cerevisiae DBKKU Y-53 and SC90 were determined using real-time RT-PCR. As shown in
Figure 3, the expression of all genes was activated in both strains of yeast during ethanol fermentation at 40 °C. Although the expression levels of
tps,
nth,
gsy, and
rsp in
S. cerevisiae DBKKU Y-53 and SC90 were not dramatically different, the genes encoding HSPs (
hsp26,
hsp70,
hsp90,
hsp104) in DBKKU Y-53 were much greater than in SC90. This finding suggests that high growth and ethanol fermentation capabilities of DBKKU Y-53 at a high temperature might be related to increased expression levels of HSP genes. Conversely, the expression level of
cdc encoding pyruvate kinase, which is involved in ATP production, in SC90 was greater than that in DBKKU Y-53, suggesting that SC90 required more ATP for growth and ethanol production at high temperature. The results in this study were in good agreement with Auesukaree
et al. [
20], who observed high expression levels of genes encoding the small HSP, HSP70, HSP90, and HSP100 family and those genes encoding trehalose-6-phosphate synthase, neutral trehalase, and glycogen synthase in
S. cerevisiae after heat shock and long-term heat exposure at 37 °C. Piper
et al. [
68] reported that several HSPs are constitutively expressed at appropriate temperatures and play a crucial role in folding and assembling proteins. Many of the HSPs, such as HSP70, HSP90, and HSP104, play an important role as molecular chaperones. These molecular chaperones depend on the energy of ATP hydrolysis for function [
69]. Therefore, increasing the expression level of
cdc during ethanol fermentation at a high temperature may provide sufficient energy not only for growth and ethanol fermentation activity but may also function as molecular chaperones to protect the structural integrity of the proteins in yeast cells. In addition to the HSPs, the accumulation of trehalose has also been reported to be associated with heat stress protection and thermotolerance [
70]. Trehalose can stabilize the protein structure, reduce the aggregation of denatured proteins, and cooperate with HSPs to promote protein refolding [
71,
72,
73].
In
S. cerevisiae, ubiquitin ligase plays a key regulatory role in many cellular processes, such as trafficking, sorting, modifying gene expression, DNA repair, RNA transport as well as the degradation of a large number of proteins in multiple cellular compartments [
74]. This protein is also involved in the pathways responsible for the regulation of chromatin function and ultimately controls gene expression under limited nutrient conditions [
75]. Most recently, Shahsavarani
et al. [
76] also demonstrated that overexpression of
RSP5 encoding ubiquitin ligase improved the ability of
S. cerevisiae to tolerate high temperatures. An increase in the ubiquitin ligase, which was observed in this study by the high expression level of
rsp during ethanol fermentation at high temperature, might regulate the transcription of some genes and induce the heat stress response through the ubiquitination process. Therefore, the ability of
S. cerevisiae DBKKU Y-53 and SC90 to grow and produce a relatively high level of ethanol at a high temperature might be explained by this mechanism.
In the thermotolerant yeast
K. marxianus, the molecular mechanisms conferring thermotolerance are complicated and are controlled by multiple genes not only for HSPs biosynthesis but also for those genes encoding the proteins involved in DNA replication and repair, RNA processing, ribosome biogenesis, and carbohydrate metabolism process [
21]. The expression of
hsp genes and those functioning to prevent protein denaturation may be insufficient to allow growth and efficient ethanol fermentation of
S. cerevisiae at high temperatures. To gain a better understanding and provide useful information for fundamental and applied research for innovative applications, further studies are required to clarify the precise mechanism conferring thermotolerance in
S. cerevisiae.








{kind=link}
{kind=link}
{kind=link}