
A Flavonoid-Rich Extract of Sambucus nigra L. Reduced Lipid Peroxidation in a Rat Experimental Model of Gentamicin Nephrotoxicity

 ,
,  ,
,  , ,
, ,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection and Processing of Elderflowers
2.2. Quantitative Analysis of Phenolic Compounds in Elderflowers
2.3. Protocol for Obtaining Elderflower Extract for In Vivo Administration
2.4. Experimental Design
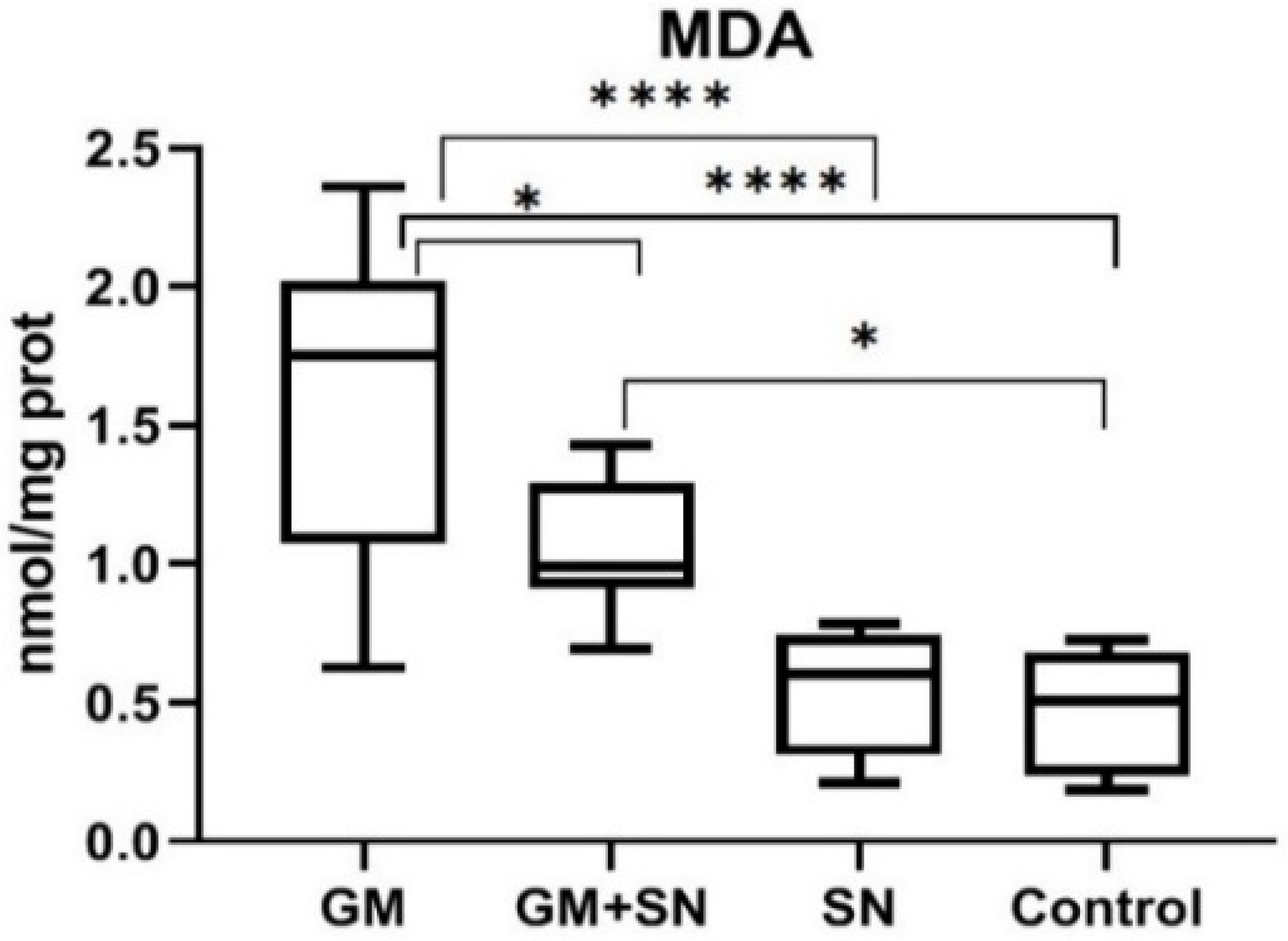
2.5. Analysis of Lipid Peroxidation
2.6. Analysis of Antioxidant Enzyme Activity
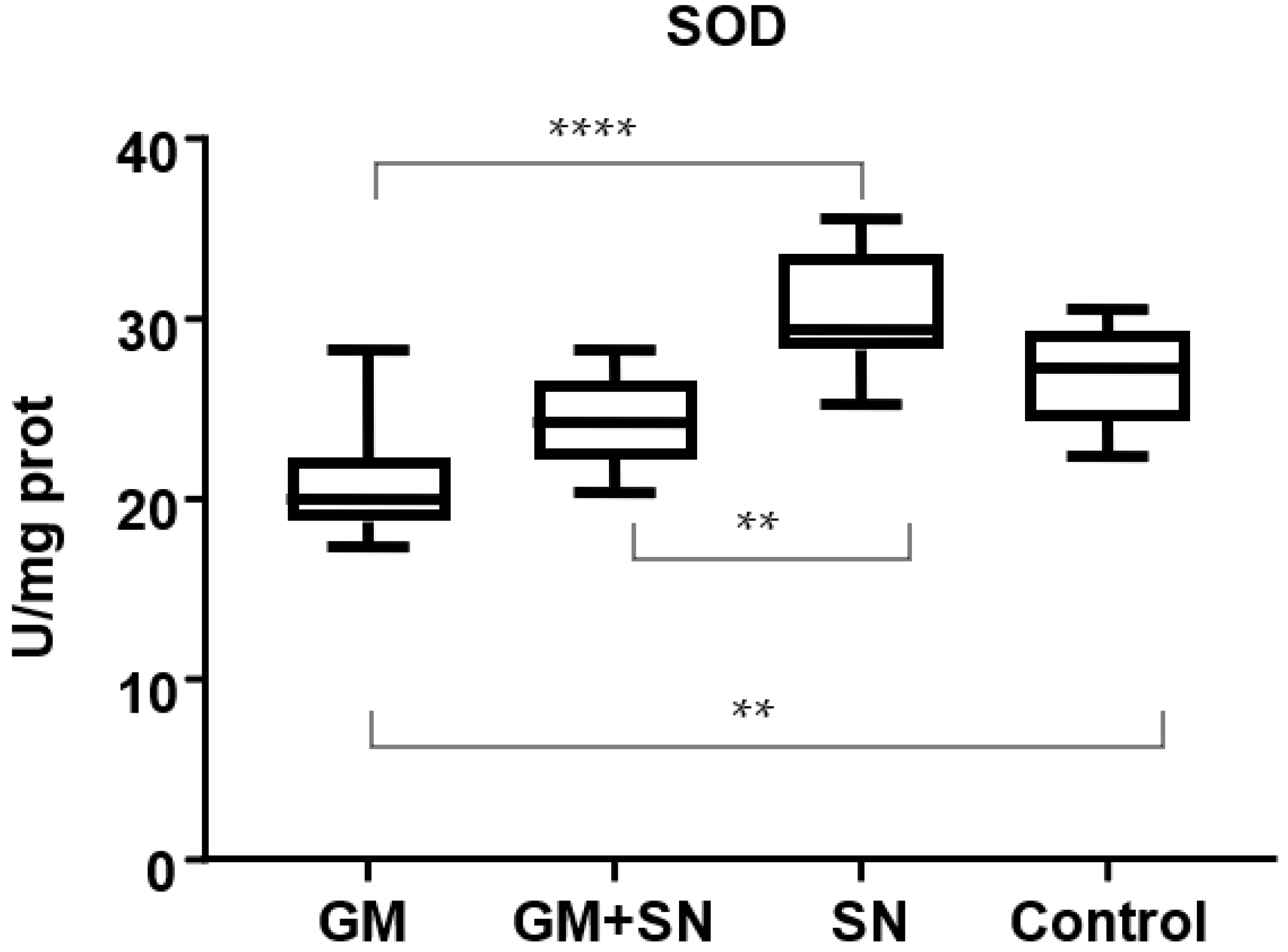
2.6.1. Superoxide Dismutase (SOD)
2.6.2. Glutathione Peroxidase (GPX)
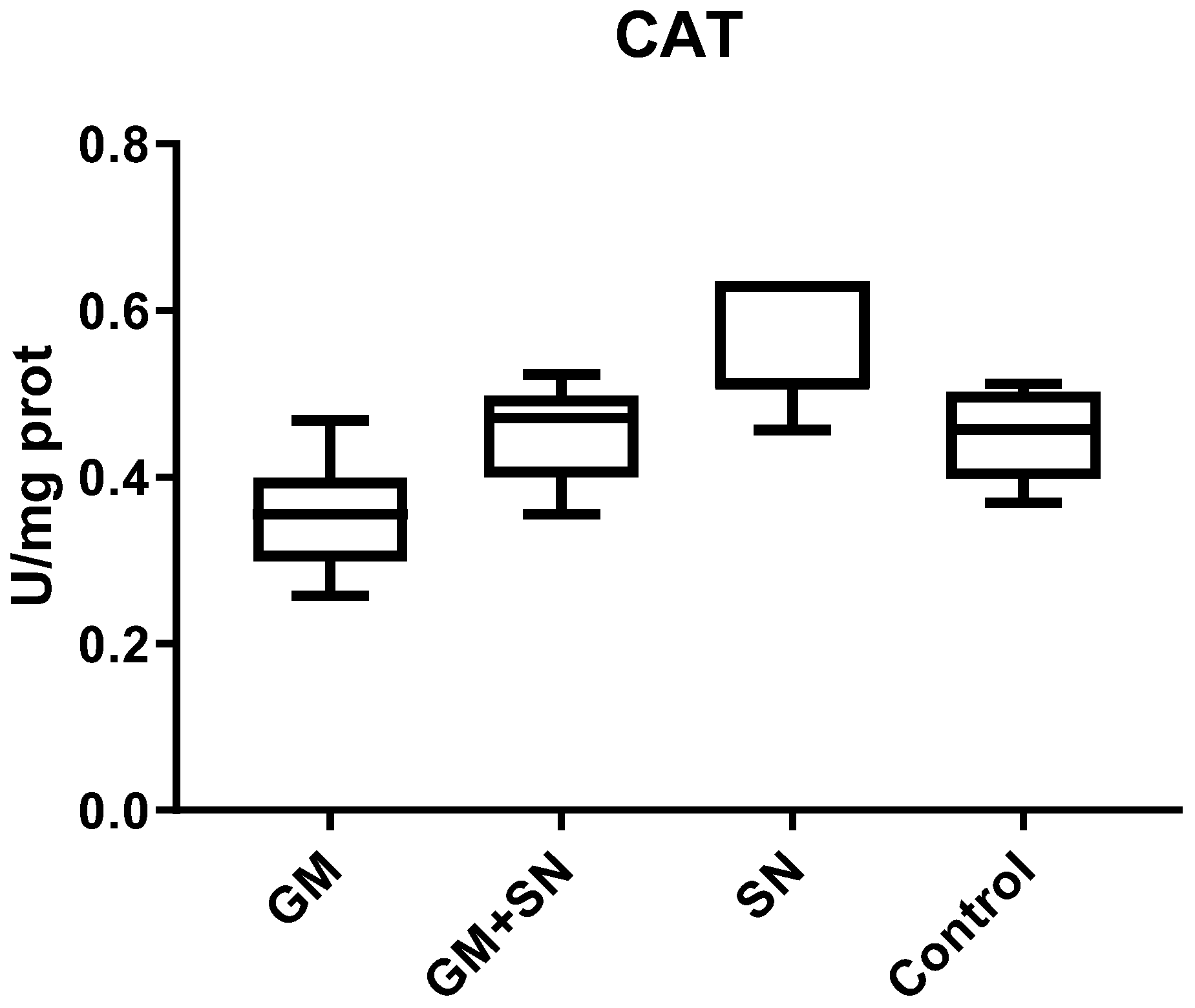
2.6.3. Catalase (CAT)
2.7. Statistical Analysis
3. Results and Discussion
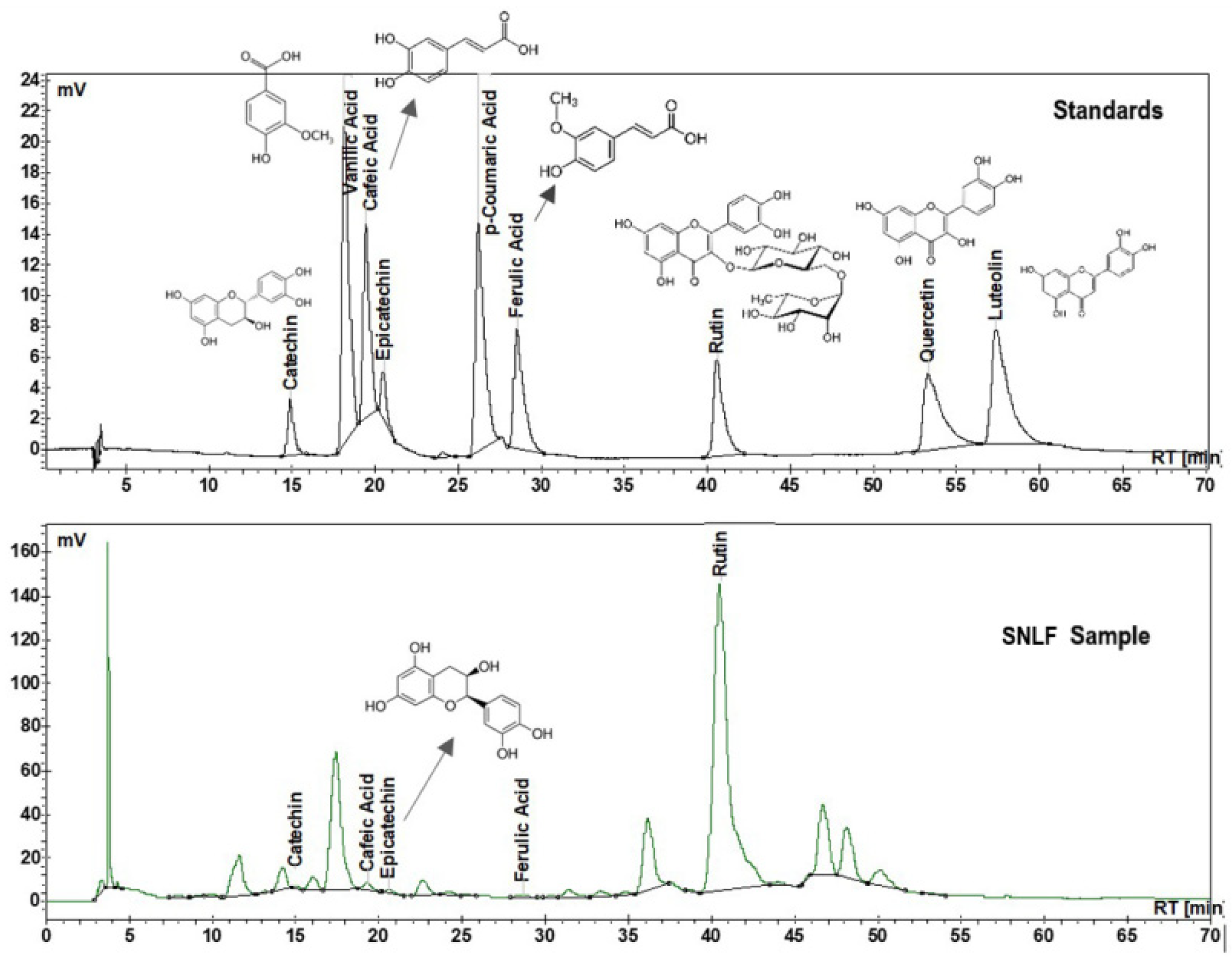
3.1. Quantitative Evaluation of Phenolic Compounds in the Analyzed SN Ethanolic Extract
3.2. Influence of SN Extract Administered In Vivo on Oxidative Stress Parameters
3.2.1. In Vivo Influence of SN Extract on Lipid Peroxidation
3.2.2. In Vivo Influence of SN Extract on Superoxide Dismutase, Catalase, and Glutathione Peroxidase
3.3. Non-Enzymatic Antioxidant Effects of Bioflavonoids from Elderflower Extract and Their Main Compound—Rutin
3.4. Role of Caffeic Acid and Ferulic Acid
3.5. New Research Direction for Elderflowers Extract
3.5.1. Action Mechanism of Active Compounds
3.5.2. Interference between SN Extract and Antimicrobial Effect of GM
3.5.3. Pharmacological Characteristics of SN Extract
3.5.4. Efficiency of SN Extract in Other Nephrotoxicity Models
3.5.5. Antiviral Effect in SARS-CoV-2 Infection
3.6. Elderflowers—Promising Medical and Nutritional Intervention in Kidney Diseases
3.7. Limits of the Study
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ANOVA | One-way analysis of variance |
| CAT | Catalase |
| CA | Caffeic acid |
| DPPH | 2,2-Diphenyl-1-picrylhydrazyl radical |
| EDTA | Ethylenediaminetetraacetic acid |
| EC50 | Half maximal effective concentration |
| FA | Ferulic acid |
| FRAP | Ferric reducing antioxidant power |
| GM | Gentamicin |
| GPX | Glutathione peroxidase |
| GSH | Glutathione |
| HCAs | Hydroxycinnamic acids |
| HPLC | High performance liquid chromatography |
| iNOS | Inducible nitric oxide synthase |
| MDA | Malondialdehyde |
| NADPH | Nicotinamide adenine dinucleotide phosphate |
| NIHCE | UK National Institute of Health and Care Excellence |
| OS | Oxidative stress |
| PBS | Phosphate-buffered saline solution |
| RNS | Reactive nitrogen species |
| ROS | Reactive oxygen species |
| RS | Reactive species |
| SEM | Standard error of the mean |
| SN | Sambucus nigra L. elderflowers |
| SOD | Superoxide dismutase |
References
- McWilliam, S.J.; Antoine, D.J.; Smyth, R.L.; Pirmohamed, M. Aminoglycoside-induced nephrotoxicity in children. Pediatr. Nephrol. 2016, 32, 2015–2025. [Google Scholar] [CrossRef]
- Cebrero-Cangueiro, T.; Marín, R.; Labrador-Herrera, G.; Smani, Y.; Cordero, E.; Pachón, J.; Pachón-Ibáñez, M.E. In vitro Activity of Pentamidine Alone and in Combination With Aminoglycosides, Tigecycline, Rifampicin, and Doripenem against Clinical Strains of Carbapenemase-Producing and/or Colistin-Resistant Enterobacteriaceae. Front. Cell. Infect. Microbiol. 2018, 8, 363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silverblatt, F.J.; Kuehn, C. Autoradiography of gentamicin uptake by the rat proximal tubule cell. Kidney Int. 1979, 15, 335–345. [Google Scholar] [CrossRef] [Green Version]
- Sandoval, R.; Leiser, J.; Molitoris, B.A. Aminoglycoside antibiotics traffic to the Golgi complex in LLC-PK1 cells. J. Am. Soc. Nephrol. 1998, 9, 167–174. [Google Scholar] [CrossRef]
- Ozbek, E. Induction of Oxidative Stress in Kidney. Int. J. Nephrol. 2012, 2012, 465897. [Google Scholar] [CrossRef] [Green Version]
- Casanova, A.G.; Vicente-Vicente, L.; Hernández-Sánchez, M.T.; Pescador, M.; Prieto, M.; Martínez-Salgado, C.; Morales, A.I.; López-Hernández, F.J. Key role of oxidative stress in animal models of aminoglycoside nephrotoxicity revealed by a systematic analysis of the antioxidant-to-nephroprotective correlation. Toxicology 2017, 385, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Elgebaly, H.A.; Mosa, N.M.; Germoush, M.O.; Brahim, A.C. The Nephro—Protective Effects of Olive Oil and Bee Honey against Gentamicin—Induced Nephrotoxicity in Rabbits. Aljouf Univ. Med. J. 2016, 300, 1–7. [Google Scholar] [CrossRef]
- Codea, R.; Mircean, M.; Nagy, A.; Sarpataky, O.; Sevastre, B.; Stan, R.L.; Hangan, A.C.; Popovici, C.; Neagu, D.; Purdoiu, R.C.; et al. Melatonine and erythropoietin prevents gentamicin induced nephrotoxicity in rats. Farmacia 2019, 67, 392–397. [Google Scholar] [CrossRef]
- Vargas, F.; Romecín, P.; Guillen, A.I.G.; Wangesteen, R.; Vargas-Tendero, P.; Paredes, M.D.; Atucha, N.M.; García-Estañ, J. Flavonoids in Kidney Health and Disease. Front. Physiol. 2018, 9, 394. [Google Scholar] [CrossRef] [Green Version]
- Ungur, R.; Buzatu, R.; Lacatus, R.; Purdoiu, R.C.; Petrut, G.; Codea, R.; Sarpataky, O.; Biris, A.; Popovici, C.; Mircean, M.; et al. Evaluation of the Nephroprotective Effect of Sambucus nigra Total Extract in a Rat Experimental Model of Gentamicine Nephrotoxicity. Rev. Chim. 2019, 70, 1971–1974. [Google Scholar] [CrossRef]
- Mahboubi, M. Sambucus nigra (black elder) as alternative treatment for cold and flu. Adv. Tradit. Med. 2020, 21, 405–414. [Google Scholar] [CrossRef]
- Viapiana, A.; Wesolowski, M. The Phenolic Contents and Antioxidant Activities of Infusions of Sambucus nigra L. Mater. Veg. 2017, 72, 82–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikulic-Petkovsek, M.; Samoticha, J.; Eler, K.; Stampar, F.; Veberic, R. Traditional Elderflower Beverages: A Rich Source of Phenolic Compounds with High Antioxidant Activity. J. Agric. Food Chem. 2015, 63, 1477–1487. [Google Scholar] [CrossRef] [PubMed]
- Gong, B.; Gou, X.; Han, T.; Qi, Y.; Ji, X.; Bai, J. Protective effects of rutin on kidney in type 1 diabetic mice. Pak. J. Pharm. Sci. 2020, 33, 597–603. [Google Scholar]
- Khajevand-Khazaei, M.-R.; Mohseni-Moghaddam, P.; Hosseini, M.; Gholami, L.; Baluchnejadmojarad, T.; Roghani, M. Rutin, a quercetin glycoside, alleviates acute endotoxemic kidney injury in C57BL/6 mice via suppression of inflammation and up-regulation of antioxidants and SIRT1. Eur. J. Pharmacol. 2018, 833, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Dawidowicz, A.L.; Wianowska, D.; Baraniak, B. The antioxidant properties of alcoholic extracts from Sambucus nigra L. (antioxidant properties of extracts). LWT Food Sci. Technol. 2006, 39, 308–315. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Christensen, K.B.; Olsen, L.C.B.; Christensen, L.P.; Grevsen, K.; Færgeman, N.J.; Kristiansen, K.; Young, J.F.; Oksbjerg, N. Bioactive Components from Flowers of Sambucus nigra L. Increase Glucose Uptake in Primary Porcine Myotube Cultures and Reduce Fat Accumulation in Caenorhabditis elegans. J. Agric. Food Chem. 2013, 61, 11033–11040. [Google Scholar] [CrossRef]
- Salem, A.M.; Ragheb, A.S.; Hegazy, M.G.A.; Matboli, M.; Eissa, S. Caffeic Acid Modulates miR-636 Expression in Diabetic Nephropathy Rats. Indian J. Clin. Biochem. 2018, 34, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Sanjeev, S.; Bidanchi, R.M.; Murthy, M.K.; Gurusubramanian, G.; Roy, V. Influence of ferulic acid consumption in ameliorating the cadmium-induced liver and renal oxidative damage in rats. Environ. Sci. Pollut. Res. 2019, 26, 20631–20653. [Google Scholar] [CrossRef]
- Mikulic-Petkovsek, M.; Ivancic, A.; Schmitzer, V.; Veberic, R.; Stampar, F. Comparison of major taste compounds and antioxidative properties of fruits and flowers of different Sambucus species and interspecific hybrids. Food Chem. 2016, 200, 134–140. [Google Scholar] [CrossRef]
- Filip, M.; Silaghi-Dumitrescu, L.; Prodan, D.; Sarosi, C.; Moldovan, M.; Cojocaru, I. Analytical Approaches for Characterization of Teeth Whitening Gels Based on Natural Extracts. Key Eng. Mater. 2017, 752, 24–28. [Google Scholar] [CrossRef]
- Huang, H.-S.; Yu, H.-S.; Yen, C.-H.; Liaw, E.-T. HPLC-DAD-ESI-MS Analysis for Simultaneous Quantitation of Phenolics in Taiwan Elderberry and Its Anti-Glycation Activity. Molecules 2019, 24, 3861. [Google Scholar] [CrossRef] [Green Version]
- Mota, A.H.; Andrade, J.M.; Rodrigues, M.J.; Custódio, L.; Bronze, M.R.; Duarte, N.; Baby, A.; Rocha, J.; Gaspar, M.M.; Simões, S.; et al. Synchronous insight of in vitro and in vivo biological activities of Sambucus nigra L. extracts for industrial uses. Ind. Crop. Prod. 2020, 154, 112709. [Google Scholar] [CrossRef]
- Koleva, P.; Tsanova-Savova, S.; Paneva, S.; Velikov, S.; Savova, Z. Polyphenols content of selected medical plants and food supplements present at Bulgarian market. Pharmacia 2021, 68, 819–826. [Google Scholar] [CrossRef]
- Kim, T.; Shin, H.; Park, S.; Kim, H.; Chung, D. Development and Validation of a Method for Determining the Quercetin-3-O-glucuronide and Ellagic Acid Content of Common Evening Primrose (Oenothera biennis) by HPLC-UVD. Molecules 2021, 26, 267. [Google Scholar] [CrossRef] [PubMed]
- Saad, W.M.M.; Ridwan, R.; Lasim, N.S.M.; Rapi, N.L.M.; Salim, F. Determination and quantification of p-coumaric acid in pineapples (Ananas comosus) extracts using gradient mode RP-HPLC. Pharmacogn. Res. 2019, 11, 67. [Google Scholar] [CrossRef]
- Sá, R.R.; Matos, R.A.; Silva, V.C.; Caldas, J.D.C.; Sauthier, M.C.D.S.; dos Santos, W.N.L.; Magalhães, H.I.F.; Junior, A.D.F.S. Determination of bioactive phenolics in herbal medicines containing Cynara scolymus, Maytenus ilicifolia Mart ex Reiss and Ptychopetalum uncinatum by HPLC-DAD. Microchem. J. 2017, 135, 10–15. [Google Scholar] [CrossRef]
- Moura, H.F.S.; Dias, F.D.S.; e Souza, L.B.S.; de Magalhães, B.E.A.; Tannus, C.D.A.; de Carvalho, W.C.; Brandão, G.C.; dos Santos, W.N.L.; Korn, M.G.A.; dos Santos, D.C.M.B.; et al. Evaluation of multielement/proximate composition and bioactive phenolics contents of unconventional edible plants from Brazil using multivariate analysis techniques. Food Chem. 2021, 363, 129995. [Google Scholar] [CrossRef]
- Han, Y.; Lu, J.S.; Xu, Y.; Zhang, L.; Hong, B.F. Rutin ameliorates renal fibrosis and proteinuria in 5/6-nephrectomized rats by anti-oxidation and inhibiting activation of TGFβ1-smad signaling. Int. J. Clin. Exp. Pathol. 2015, 8, 4725–4734. [Google Scholar]
- Yagmurca, M.; Yasar, Z.; Bas, O. Effects of quercetin on kidney injury induced by doxorubicin. Bratisl Lek List. 2015, 116, 486–489. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, C.-L.; Peng, C.-C.; Chen, K.-C.; Peng, R.Y. Rutin (Quercetin Rutinoside) Induced Protein-Energy Malnutrition in Chronic Kidney Disease, but Quercetin Acted Beneficially. J. Agric. Food Chem. 2013, 61, 7258–7267. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.-C.; Hsieh, C.-L.; Ker, Y.-B.; Wang, H.-Y.; Chen, K.-C.; Peng, R.Y. Selected nutraceutic screening by therapeutic effects on doxorubicin-induced chronic kidney disease. Mol. Nutr. Food Res. 2012, 56, 1541–1558. [Google Scholar] [CrossRef]
- Al-Rejaie, S.S.; Abuohashish, H.M.; Alkhamees, O.A.; Aleisa, A.M.; Alroujayee, A.S. Gender difference following high cholesterol diet induced renal injury and the protective role of rutin and ascorbic acid combination in Wistar albino rats. Lipids Heal. Dis. 2012, 11, 41. [Google Scholar] [CrossRef] [Green Version]
- Bledsoe, G.; Shen, B.; Yao, Y.-Y.; Hagiwara, M.; Mizell, B.; Teuton, M.; Grass, D.; Chao, L.; Chao, J. Role of Tissue Kallikrein in Prevention and Recovery of Gentamicin-Induced Renal Injury. Toxicol. Sci. 2008, 102, 433–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thibault, N.; Grenier, L.; Simard, M.; Bergeron, M.G.; Beauchamp, D. Attenuation by daptomycin of gentamicin-induced experimental nephrotoxicity. Antimicrob. Agents Chemother. 1994, 38, 1027–1035. [Google Scholar] [CrossRef] [Green Version]
- Medscape. Drugs & Diseases—Gentamicin. Available online: https://reference.medscape.com/drug/gentak-garamycin-gentamicin-342517 (accessed on 27 December 2021).
- Prescribers’ Digital Reference—Gentamicin Sulfate—Drug Summary. Available online: https://www.pdr.net/drug-summary/Gentamicin-Injection-10-mg-mL-gentamicin-sulfate-3300 (accessed on 27 December 2021).
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Conti, M.; Morand, P.C.; Levillain, P.; Lemonnier, A. Improved fluorometric determination of malonaldehyde. Clin. Chem. 1991, 37, 1273–1275. [Google Scholar] [CrossRef]
- Flohé, L. Superoxide dismutase assays. Methods Enzym. 1984, 105, 93–104. [Google Scholar] [CrossRef]
- Titheradge, M.A. The Enzymatic Measurement of Nitrate and Nitrite. Methods Mol. Biol. 1998, 100, 83–91. [Google Scholar] [CrossRef]
- Pippenger, C.E.; Browne, R.W.; Armstrong, D. Regulatory Antioxidant Enzymes. Methods Mol. Biol. 1998, 108, 299–314. [Google Scholar] [CrossRef] [PubMed]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxidative Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef]
- Chainy, G.B.N.; Sahoo, D.K. Hormones and oxidative stress: An overview. Free Radic. Res. 2020, 54, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.L.; Norhaizan, M.E.; Liew, W.-P.-P. Nutrients and Oxidative Stress: Friend or Foe? Oxidative Med. Cell. Longev. 2018, 2018, 9719584. [Google Scholar] [CrossRef] [PubMed]
- Gangwar, R.S.; Bevan, G.H.; Palanivel, R.; Das, L.; Rajagopalan, S. Oxidative stress pathways of air pollution mediated toxicity: Recent insights. Redox Biol. 2020, 34, 101545. [Google Scholar] [CrossRef] [PubMed]
- Ungur, R.; Dronca, M.; Crăciun, E.C.; Rusu, R.L.; Văleanu, M.; Onac, I.; Borda, I.M.; Laszlo, I. Improvement of total antioxidant status, a possible bioeffect of the ultrasound therapy—A pilot study. Rev. Romana Med. Lab. 2011, 19, 177–183. [Google Scholar]
- Mironczuk-Chodakowska, I.; Witkowska, A.; Zujko, M.E. Endogenous non-enzymatic antioxidants in the human body. Adv. Med. Sci. 2018, 63, 68–78. [Google Scholar] [CrossRef]
- Walker, P.D.; Shah, S.V. Evidence suggesting a role for hydroxyl radical in gentamicin-induced acute renal failure in rats. J. Clin. Investig. 1988, 81, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Mccord, J.M. Iron, free radicals, and oxidative injury. J. Nutr. 2004, 134, 3171S–3172S. [Google Scholar] [CrossRef]
- Ratliff, B.B.; Abdulmahdi, W.; Pawar, R.; Wolin, M.S. Oxidant Mechanisms in Renal Injury and Disease. Antioxid. Redox Signal 2016, 25, 119–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eid, R.; Arab, N.T.; Greenwood, M.T. Iron mediated toxicity and programmed cell death: A review and a re-examination of existing paradigms. Biochim. Biophys. Acta BBA-Bioenerg. 2017, 1864, 399–430. [Google Scholar] [CrossRef]
- Morales, A.I.; Detaille, D.; Prieto, M.; Puente, A.; Briones, E.; Arévalo, M.; Leverve, X.; Lopez-Novoa, J.M.; El-Mir, M.-Y. Metformin prevents experimental gentamicin-induced nephropathy by a mitochondria-dependent pathway. Kidney Int. 2010, 77, 861–869. [Google Scholar] [CrossRef]
- Nakazawa, T.; Miyanoki, Y.; Urano, Y.; Uehara, M.; Saito, Y.; Noguchi, N. Effect of vitamin E on 24(S)-hydroxycholesterol-induced necroptosis-like cell death and apoptosis. J. Steroid Biochem. Mol. Biol. 2016, 169, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Negrette-Guzmán, M.; García-Niño, W.R.; Tapia, E.; Zazueta, C.; Huerta-Yepez, S.; León-Contreras, J.C.; Hernández-Pando, R.; Aparicio-Trejo, O.E.; Madero, M.; Pedraza-Chaverri, J. Curcumin Attenuates Gentamicin-Induced Kidney Mitochondrial Alterations: Possible Role of a Mitochondrial Biogenesis Mechanism. Evid.-Based Complement. Altern. Med. 2015, 2015, 917435. [Google Scholar] [CrossRef] [PubMed]
- Helal, M.G.; Zaki, M.M.A.F.; Said, E. Nephroprotective effect of saxagliptin against gentamicin-induced nephrotoxicity, emphasis on anti-oxidant, anti-inflammatory and anti-apoptic effects. Life Sci. 2018, 208, 64–71. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, A.K. Chemistry and Biological Activities of Flavonoids: An Overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hfaiedh, M.; Brahmi, D.; Zourgui, M.N.; Zourgui, L. Phytochemical analysis and nephroprotective effect of cactus (Opuntia ficus-indica) cladodes on sodium dichromate-induced kidney injury in rats. Appl. Physiol. Nutr. Metab. 2019, 44, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Yu, J.S.; Lee, S.R.; Hwang, G.S.; Kang, K.S.; Park, J.G.; Kim, H.Y.; Kim, K.H.; Yamabe, N. Beneficial Effects of Bioactive Compounds in Mulberry Fruits against Cisplatin-Induced Nephrotoxicity. Int. J. Mol. Sci. 2018, 19, 1117. [Google Scholar] [CrossRef] [Green Version]
- Marțiș, G.S.; Mureșan, V.; Marc, R.M.; Mureșan, C.C.; Pop, C.R.; Buzgău, G.; Mureșan, A.E.; Ungur, R.A.; Muste, S. The Physicochemical and Antioxidant Properties of Sambucus nigra L. and Sambucus nigra Haschberg during Growth Phases: From Buds to Ripening. Antioxidants 2021, 10, 1093. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, S.S.; da Silva, P.M.C.; Silva, A.; Nunes, F.M. Effect of harvesting year and elderberry cultivar on the chemical composition and potential bioactivity: A three-year study. Food Chem. 2019, 302, 125366. [Google Scholar] [CrossRef] [PubMed]
- Topolska, J.; Kostecka-Gugała, A.; Ostachowicz, B.; Latowski, D. Selected metal content and antioxidant capacity of Sambucus nigra flowers from the urban areas versus soil parameters and traffic intensity. Environ. Sci. Pollut. Res. 2019, 27, 668–677. [Google Scholar] [CrossRef]
- Singureanu, V.; Ungur, R.; Onac, I.; Kovacs, M.H.; Moldovan, G.; Singureanu, V. Automatic Germination Evaluation and Qualitative Analysis of Essential Oil of Mentha ×piperita L. under the Influence of High Frequency Pulsatile Electro-magnetic and Ultrasound Pulsatile Fields. Not. Bot. Horti Agrobot. 2015, 43, 146–152. [Google Scholar] [CrossRef] [Green Version]
- Onac, I.; Singureanu, V.; Moldovan, G.; Ungur, R. High Frequency Pulsatile Electromagnetic Fields and Ultrasound Pulsatile Fields Impact on Germination Dynamic at Ocimum basilicum L. and O. basilicum var. purpurascens Benth., Observed with Open Source Software. Not. Bot. Horti Agrobot. 2016, 44, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Senica, M.; Stampar, F.; Veberic, R.; Mikulic-Petkovsek, M. The higher the better? Differences in phenolics and cyanogenic glycosides in Sambucus nigra leaves, flowers and berries from different altitudes. J. Sci. Food Agric. 2016, 97, 2623–2632. [Google Scholar] [CrossRef]
- Muhammad, A.; Arthur, D.E.; Babangida, S.; Erukainure, O.; Malami, I.; Sani, H.; Abdulhamid, A.W.; Ajiboye, I.O.; Saka, A.A.; Hamza, N.M.; et al. Modulatory role of rutin on 2,5-hexanedione-induced chromosomal and DNA damage in rats: Validation of computational predictions. Drug Chem. Toxicol. 2018, 43, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Schafer, E.W.; Bowles, W.A.; Hurlbut, J. The acute oral toxicity, repellency, and hazard potential of 998 chemicals to one or more species of wild and domestic birds. Arch. Environ. Contam. Toxicol. 1983, 12, 355–382. [Google Scholar] [CrossRef] [PubMed]
- Zieger, E.; Tice, R. Chlorogenic Acid and Caffeic Acid: Review of Toxicological Literature. Integrated Laboratory System. Available online: https://ntp.niehs.nih.gov/ntp/htdocs/chem_background/exsumpdf/chlorogenicacid_508.pdf (accessed on 29 December 2021).
- Zduńska, K.; Dana, A.; Kolodziejczak, A.; Rotsztejn, H. Antioxidant Properties of Ferulic Acid and Its Possible Application. Ski. Pharmacol. Physiol. 2018, 31, 332–336. [Google Scholar] [CrossRef] [PubMed]
- Tada, Y.; Tayama, K.; Aoki, N. Acute oral toxicity of ferulic acid, natural food additive, in rats. Ann. Rep. Tokyo Metr. Lab. 1999, 50, 311–313. [Google Scholar]
- Shin, H.A.; Shin, Y.S.; Kang, S.U.; Kim, J.-H.; Oh, Y.-T.; Park, K.H.; Lee, B.H.; Kim, C.-H. Radioprotective effect of epicatechin in cultured human fibroblasts and zebrafish. J. Radiat. Res. 2013, 55, 32–40. [Google Scholar] [CrossRef] [Green Version]
- Yusuf, A.J.; Abdullahi, M.I.; Musa, A.M.; Abubakar, H.; Amali, A.M.; Nasir, A.H. Potential inhibitors of SARS CoV-2 from Neocarya macrophylla: Chemoinformatic and Molecular modeling studies against three key targets. Turk. J. Pharm. Sci. 2021, 2, 100038. [Google Scholar] [CrossRef]
- Otunctemur, A.; Ozbek, E.; Cekmen, M.; Cakir, S.S.; Dursun, M.; Polat, E.C.; Somay, A.; Ozbay, N. Protective Effect of Montelukast Which Is Cysteinyl-Leukotriene Receptor Antagonist on Gentamicin-Induced Nephrotoxicity and Oxidative Damage in Rat Kidney. Ren. Fail. 2013, 35, 403–410. [Google Scholar] [CrossRef]
- Yarijani, Z.M.; Najafi, H.; Shackebaei, D.; Madani, S.H.; Modarresi, M.; Jassemi, S.V. Amelioration of renal and hepatic function, oxidative stress, inflammation and histopathologic damages by Malva sylvestris extract in gentamicin induced renal toxicity. Biomed. Pharmacother. 2019, 112, 108635. [Google Scholar] [CrossRef]
- El-Kashef, D.H.; El-Kenawi, A.; Suddek, G.M.; Salem, H. Protective effect of allicin against gentamicin-induced nephrotoxicity in rats. Int. Immunopharmacol. 2015, 29, 679–686. [Google Scholar] [CrossRef]
- Khattab, H.A.; Wazzan, M.A.; Al-Ahdab, M.A. Nephroprotective potential of artichoke leaves extract against gentamicin in rats: Antioxidant mechanisms. Pak. J. Pharm. Sci. 2016, 29, 1775–1782. [Google Scholar] [PubMed]
- Hajihashemi, S.; Jafarian, T.; Ahmadi, M.; Rahbari, A.; Ghanbari, F. Ameliorative Effects of Zataria Multiflora Hydro-Alcoholic extract on Gentamicin Induced Nephrotoxicity in Rats. Drug Res. 2018, 68, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Changizi-Ashtiyani, S.; Seddigh, A.; Najafi, H.; Hossaini, N.; Avan, A.; Akbary, A.; Manian, M.; Nedaeinia, R. Pimpinella anisum L. ethanolic extract ameliorates the gentamicin—Induced nephrotoxicity in rats. Nephrology 2017, 22, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Heidarian, E.; Jafari-Dehkordi, E.; Valipour, P.; Ghatreh-Samani, K.; Ashrafi-Eshkaftaki, L. Nephroprotective and Anti-Inflammatory Effects of Pistacia atlantica Leaf Hydroethanolic Extract against Gentamicin-Induced Nephrotoxicity in Rats. J. Diet. Suppl. 2017, 14, 489–502. [Google Scholar] [CrossRef]
- Cekmen, M.; Otunctemur, A.; Ozbek, E.; Cakir, S.S.; Dursun, M.; Polat, E.C.; Somay, A.; Ozbay, N. Pomegranate Extract Attenuates Gentamicin-Induced Nephrotoxicity in Rats by Reducing Oxidative Stress. Ren. Fail. 2012, 35, 268–274. [Google Scholar] [CrossRef] [Green Version]
- Yildirim, B.A.; Kordali, S.; Kapakin, K.A.T.; Yildirim, F.; Senocak, E.A.; Altun, S. Effect of Helichrysum plicatum DC. subsp. plicatum ethanol extract on gentamicin-induced nephrotoxicity in rats. J. Zhejiang Univ. Sci. B 2017, 18, 501–511. [Google Scholar] [CrossRef] [Green Version]
- Naidu, M.; Shifow, A.A.; Kumar, K.V.; Ratnakar, K. Ginkgo biloba extract ameliorates gentamicin-induced nephrotoxicity in rats. Phytomedicine 2000, 7, 191–197. [Google Scholar] [CrossRef]
- El-Ashmawy, I.M.; El-Nahas, A.F.; Salama, O.M. Grape Seed Extract Prevents Gentamicin-Induced Nephrotoxicity and Genotoxicity in Bone Marrow Cells of Mice. Basic Clin. Pharmacol. Toxicol. 2006, 99, 230–236. [Google Scholar] [CrossRef]
- Feyissa, T.; Asres, K.; Engidawork, E. Renoprotective effects of the crude extract and solvent fractions of the leaves of Euclea divinorum Hierns against gentamicin-induced nephrotoxicity in rats. J. Ethnopharmacol. 2013, 145, 758–766. [Google Scholar] [CrossRef]
- El Gamal, A.; Alsaid, M.S.; Raish, M.; Al-Sohaibani, M.; Al-Massarani, S.M.; Ahmad, A.; Hefnawy, M.; Al-Yahya, M.; Basoudan, O.A.; Rafatullah, S. Beetroot (Beta vulgaris L.) Extract Ameliorates Gentamicin-Induced Nephrotoxicity Associated Oxidative Stress, Inflammation, and Apoptosis in Rodent Model. Mediat. Inflamm. 2014, 2014, 983952. [Google Scholar] [CrossRef] [Green Version]
- Sawardekar, S.; Patel, T. Evaluation of the effect of Boerhavia diffusa on gentamicin-induced nephrotoxicity in rats. J. Ayurveda Integr. Med. 2015, 6, 95–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ajami, M.; Eghtesadi, S.; Pazoki-Toroudi, H.; Habibey, R.; Ebrahimi, S.A. Effect of crocus sativus on gentamicin induced nephrotoxicity. Biol. Res. 2010, 43, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Galal, H.M.; El-Rady, N.M.A. Aqueous garlic extract supresses experimental gentamicin induced renal pathophysiology mediated by oxidative stress, inflammation and Kim-1. Pathophysiology 2019, 26, 271–279. [Google Scholar] [CrossRef] [PubMed]
- Arjinajarn, P.; Pongchaidecha, A.; Chueakula, N.; Jaikumkao, K.; Chatsudthipong, V.; Mahatheeranont, S.; Norkaew, O.; Chattipakorn, N.; Lungkaphin, A. Riceberry bran extract prevents renal dysfunction and impaired renal organic anion transporter 3 (Oat3) function by modulating the PKC/Nrf2 pathway in gentamicin-induced nephrotoxicity in rats. Phytomedicine 2016, 23, 1753–1763. [Google Scholar] [CrossRef]
- Jain, A.; Nahata, A.; Singhai, A.K. Effect of Tephrosia purpurea (L.) Pers. Leaves on Gentamicin-Induced Nephrotoxicity in Rats. Sci. Pharm. 2013, 81, 1071–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajihashemi, S.; Ahmadi, M.; Chehrei, A.; Ghanbari, F. Ameliorative effect of cotreatment with the methanolic leaf extract of Urtica dioica on acute kidney injury induced by gentamicin in rats. Avicenna J. Phytomed. 2019, 10, 273–286. [Google Scholar] [CrossRef]
- Vysakh, A.; Abhilash, S.; Kuriakose, J.; Midhun, S.J.; Jyothis, M.; Latha, M.S. Protective effect of Rotula aquatica Lour against gentamicin induced oxidative stress and nephrotoxicity in Wistar rats. Biomed. Pharmacother. 2018, 106, 1188–1194. [Google Scholar] [CrossRef]
- Raeeszadeh, M.; Rezaee, M.; Akbari, A.; Khademi, N. The comparison of the effect of Origanum vulgare L. extract and vitamin C on the gentamycin-induced nephrotoxicity in rats. Drug Chem. Toxicol. 2021, 1–8. [Google Scholar] [CrossRef]
- Veljković, M.; Pavlović, D.R.; Stojiljković, N.; Ilić, S.; Jovanovic, I.; Ulrih, N.P.; Rakić, V.; Velickovic, L.J.; Sokolovic, D. Bilberry: Chemical Profiling, in Vitro and in Vivo Antioxidant Activity and Nephroprotective Effect against Gentamicin Toxicity in Rats. Phytother. Res. 2016, 31, 115–123. [Google Scholar] [CrossRef]
- Ademiluyi, A.O.; Oboh, G.; Ogunsuyi, O.B.; Akinyemi, A. Attenuation of gentamycin-induced nephrotoxicity in rats by dietary inclusion of ginger (Zingiber officinale) and turmeric (Curcuma longa) rhizomes. Nutr. Heal. 2012, 21, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Stoilova, I.; Wilker, M.; Stoyanova, A.; Krastanov, A.; Stanchev, V. Antioxidant activity of extract from elder flower (Sambucus nigra L.). Herba Pol. 2007, 53, 45–54. [Google Scholar]
- Młynarczyk, K.; Walkowiak-Tomczak, D.; Staniek, H.; Kidoń, M.; Łysiak, G.P. The Content of Selected Minerals, Bioactive Compounds, and the Antioxidant Properties of the Flowers and Fruit of Selected Cultivars and Wildly Growing Plants of Sambucus nigra L. Molecules 2020, 25, 876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morsy, M.A.; Ibrahim, S.A.; Amin, E.F.; Kamel, M.Y.; Rifaai, R.A.; Hassan, M.K. Sildenafil Ameliorates Gentamicin-Induced Nephrotoxicity in Rats: Role of iNOS and eNOS. J. Toxicol. 2014, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Miao, L.; Clair, D.K.S. Regulation of superoxide dismutase genes: Implications in disease. Free Radic. Biol. Med. 2009, 47, 344–356. [Google Scholar] [CrossRef] [Green Version]
- Yoshioka, Y.; Kitao, T.; Kishino, T.; Yamamuro, A.; Maeda, S. Nitric Oxide Protects Macrophages from Hydrogen Peroxide-Induced Apoptosis by Inducing the Formation of Catalase. J. Immunol. 2006, 176, 4675–4681. [Google Scholar] [CrossRef] [Green Version]
- Rindler, P.M.; Plafker, S.M.; Szweda, L.I.; Kinter, M. High Dietary Fat Selectively Increases Catalase Expression within Cardiac Mitochondria. J. Biol. Chem. 2013, 288, 1979–1990. [Google Scholar] [CrossRef] [Green Version]
- Dobashi, K.; Asayama, K.; Nakane, T.; Kodera, K.; Hayashibe, H.; Nakazawa, S. Induction of glutathione peroxidase in response to inactivation by nitric oxide. Free. Radic. Res. 2001, 35, 319–327. [Google Scholar] [CrossRef]
- Medić, B.; Stojanović, M.; Rovčanin, B.; Kekic, D.; Škodrić, S.R.; Jovanović, G.B.; Vujović, K.S.; Divac, N.; Stojanović, R.; Radenković, M.; et al. Pioglitazone attenuates kidney injury in an experimental model of gentamicin-induced nephrotoxicity in rats. Sci. Rep. 2019, 9, 13689. [Google Scholar] [CrossRef]
- Martínez-Salgado, C.; Eleno, L.; Tavares, P.; Rodríguez-Barbero, A.; García-Criado, J.; Bolaños, J.P.; López-Novoa, J.M. Involvement of reactive oxygen species on gentamicin-induced mesangial cell activation. Kidney Int. 2002, 62, 1682–1692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahmoud, A.M.; Ahmed, O.; Galaly, S.R. Thymoquinone and curcumin attenuate gentamicin-induced renal oxidative stress, inflammation and apoptosis in rats. EXCLI J. 2014, 13, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Ghaznavi, H.; Fatemi, I.; Kalantari, H.; Hosseini Tabatabaei, S.M.T.; Mehrabani, M.; Gholamine, B.; Kalantar, M.; Mehrzadi, S.; Goudarzi, M. Ameliorative effects of gallic acid on gentamicin-induced nephrotoxicity in rats. J. Asian Nat. Prod. Res. 2018, 20, 1182–1193. [Google Scholar] [CrossRef] [PubMed]
- Yazar, E.; Elmas, M.; Altunok, V.; Sivrikaya, A.; Oztekin, E.; Birdane, Y.O. Effects of aminoglycoside antibiotics on renal an-tioxidants, malondialdehyde levels, and some serum biochemical parameters. Can. J. Vet. Res. 2003, 67, 239–240. [Google Scholar]
- Ramsammy, L.; Ling, K.-Y.; Josepovitz, C.; Levine, R.; Kaloyanides, G.J. Effect of gentamicin on lipid peroxidation in rat renal cortex. Biochem. Pharmacol. 1985, 34, 3895–3900. [Google Scholar] [CrossRef]
- Randjelovic, P.; Veljkovic, S.; Stojiljkovic, N.; Sokolovic, D.; Ilic, I. Gentamicinnephrotoxicity in animals: Current knowledge and future perspectives. EXCLI J. 2017, 16, 388–399. [Google Scholar] [PubMed]
- Pan, H.; Shen, K.; Wang, X.; Meng, H.; Wang, C.; Jin, B. Protective Effect of Metalloporphyrins against Cisplatin-Induced Kidney Injury in Mice. PLoS ONE 2014, 9, e86057. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, T.; Kuragano, T.; Nanami, M.; Nagasawa, Y.; Hasuike, Y. Misdistribution of iron and oxidative stress in chronic kidney disease. Free. Radic. Biol. Med. 2018, 133, 248–253. [Google Scholar] [CrossRef] [PubMed]
- El-Far, M.A.; Bakr, M.A.; Farahat, S.E.; El-Fattah, E.A.A. Glutathione peroxidase activity in patients with renal disorders. Clin. Exp. Nephrol. 2005, 9, 127–131. [Google Scholar] [CrossRef]
- Banning, A.; Deubel, S.; Kluth, D.; Zhou, Z.; Brigelius-Flohé, R. The GI-GPx Gene Is a Target for Nrf2. Mol. Cell. Biol. 2005, 25, 4914–4923. [Google Scholar] [CrossRef] [Green Version]
- Amiri, F.S. Intracellular organelles in health and kidney disease. Nephrol. Ther. 2019, 15, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Ueda, N.; Guidet, B.; Shah, S.V. Gentamicin-induced mobilization of iron from renal cortical mitochondria. Am. J. Physiol. 1993, 265, F435–F439. [Google Scholar] [CrossRef] [PubMed]
- Glorieux, C.; Zamocky, M.; Sandoval, J.M.; Verrax, J.; Calderon, P.B. Regulation of catalase expression in healthy and cancerous cells. Free Radic. Biol. Med. 2015, 87, 84–97. [Google Scholar] [CrossRef] [PubMed]
- Su, K.-Y.; Yu, C.Y.; Chen, Y.-W.; Huang, Y.-T.; Chen, C.-T.; Wu, H.-F.; Chen, Y.-L.S. Rutin, a Flavonoid and Principal Component of Saussurea involucrata, Attenuates Physical Fatigue in a Forced Swimming Mouse Model. Int. J. Med. Sci. 2014, 11, 528–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kandemir, F.M.; Ozkaraca, M.; Yildirim, B.A.; Hanedan, B.; Kirbas, A.; Kilic, K.; Aktas, E.; Benzer, F. Rutin attenuates gentamicin-induced renal damage by reducing oxidative stress, inflammation, apoptosis, and autophagy in rats. Ren. Fail. 2015, 37, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Simos, Y.V.; Verginadis, I.I.; Toliopoulos, I.K.; Velalopoulou, A.P.; Karagounis, I.V.; Karkabounas, S.C.; Evangelou, A.M. Effects of catechin and epicatechin on superoxide dismutase and glutathione peroxidase activity, in vivo. Redox Rep. 2012, 17, 181–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zelko, I.N.; Mariani, T.J.; Folz, R.J. Superoxide dismutase multigene family: A comparison of the CuZn-SOD (SOD1), Mn-SOD (SOD2), and EC-SOD (SOD3) gene structures, evolution, and expression. Free Radic. Biol. Med. 2002, 33, 337–349. [Google Scholar] [CrossRef]
- Kitada, M.; Xu, J.; Ogura, Y.; Monno, I.; Koya, D. Manganese Superoxide Dismutase Dysfunction and the Pathogenesis of Kidney Disease. Front. Physiol. 2020, 11, 755. [Google Scholar] [CrossRef] [PubMed]
- Marklund, S.L. Extracellular superoxide dismutase and other superoxide dismutase isoenzymes in tissues from nine mammalian species. Biochem. J. 1984, 222, 649–655. [Google Scholar] [CrossRef] [Green Version]
- Sitarek, P.; Skała, E.; Wysokińska, H.; Wielanek, M.; Szemraj, J.; Toma, M.; Śliwiński, T. The Effect of Leonurus sibiricus Plant Extracts on Stimulating Repair and Protective Activity against Oxidative DNA Damage in CHO Cells and Content of Phenolic Compounds. Oxidative Med. Cell. Longev. 2015, 2016, 5738193. [Google Scholar] [CrossRef] [Green Version]
- Walker, P.D.; Shah, S.V. Gentamicin enhanced production of hydrogen peroxide by renal cortical mitochondria. Am. J. Physiol. Physiol. 1987, 253, C495–C499. [Google Scholar] [CrossRef] [PubMed]
- Liochev, S.I.; Fridovich, I. The Haber-Weiss cycle—70 years later: An alternative view. Redox Rep. 2002, 7, 55–57. [Google Scholar] [CrossRef] [PubMed]
- Kakhlon, O.; Cabantchik, Z.I. The labile iron pool: Characterization, measurement, and participation in cellular processes. Free Radic. Biol. Med. 2002, 33, 1037–1046. [Google Scholar] [CrossRef]
- Zhou, D.R.; Eid, R.; Miller, K.A.; Boucher, E.; Mandato, C.A.; Greenwood, M.T. Intracellular second messengers mediate stress inducible hormesis and Programmed Cell Death: A review. Biochim. Biophys. Acta BBA-Bioenerg. 2019, 1866, 773–792. [Google Scholar] [CrossRef] [PubMed]
- Dinte, E.; Vostinaru, O.; Samoila, O.; Sevastre, B.; Bodoki, E. Ophthalmic Nanosystems with Antioxidants for the Prevention and Treatment of Eye Diseases. Coatings 2020, 10, 36. [Google Scholar] [CrossRef] [Green Version]
- Młynarczyk, K.; Walkowiak-Tomczak, D.; Łysiak, G.P. Bioactive properties of Sambucus nigra L. as a functional ingredient for food and pharmaceutical industry. J. Funct. Foods 2018, 40, 377–390. [Google Scholar] [CrossRef] [PubMed]
- Nijveldt, R.J.; van Nood, E.; van Hoorn, D.E.C.; Boelens, P.G.; van Norren, K.; van Leeuwen, P.A.M. Flavonoids: A review of probable mechanisms of action and potential applications. Am. J. Clin. Nutr. 2001, 74, 418–425. [Google Scholar] [CrossRef]
- Robak, J.; Gryglewski, R.J. Flavonoids are scavengers of superoxide anions. Biochem. Pharmacol. 1988, 37, 837–841. [Google Scholar] [CrossRef]
- Dugas, J.A.J.; Castañeda-Acosta, J.; Bonin, G.C.; Price, K.L.; Fischer, N.H.; Winston, G.W. Evaluation of the Total Peroxyl Radical-Scavenging Capacity of Flavonoids: Structure−Activity Relationships. J. Nat. Prod. 2000, 63, 327–331. [Google Scholar] [CrossRef]
- Hanasaki, Y.; Ogawa, S.; Fukui, S. The correlation between active oxygens scavenging and antioxidative effects of flavonoids. Free Radic. Biol. Med. 1994, 16, 845–850. [Google Scholar] [CrossRef]
- Ho, G.T.T.; Wangensteen, H.; Barsett, H. Elderberry and Elderflower Extracts, Phenolic Compounds, and Metabolites and Their Effect on Complement, RAW 264.7 Macrophages and Dendritic Cells. Int. J. Mol. Sci. 2017, 18, 584. [Google Scholar] [CrossRef] [Green Version]
- Tajner-Czopek, A.; Gertchen, M.; Rytel, E.; Kita, A.; Kucharska, A.Z.; Sokół-Łętowska, A. Study of Antioxidant Activity of Some Medicinal Plants Having High Content of Caffeic Acid Derivatives. Antioxidants 2020, 9, 412. [Google Scholar] [CrossRef]
- Karamać, M.; Koleva, L.; Kancheva, V.D.; Amarowicz, R. The Structure–Antioxidant Activity Relationship of Ferulates. Molecules 2017, 22, 527. [Google Scholar] [CrossRef] [Green Version]
- Genaro-Mattos, T.; Maurício, D.Q.; Rettori, D.; Alonso, A.; Hermes-Lima, M. Antioxidant Activity of Caffeic Acid against Iron-Induced Free Radical Generation—A Chemical Approach. PLoS ONE 2015, 10, e0129963. [Google Scholar] [CrossRef] [Green Version]
- Latunde-Dada, G.O. Ferroptosis: Role of lipid peroxidation, iron and ferritinophagy. Biochim. Biophys. Acta BBA-Gen. Subj. 2017, 1861, 1893–1900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srinivasan, M.; Sudheer, A.R.; Menon, V.P. Ferulic Acid: Therapeutic Potential Through Its Antioxidant Property. J. Clin. Biochem. Nutr. 2007, 40, 92–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, L.; Qi, Y.; Liu, Z.; Xi, Y.; Xue, W. Effect of tannic acid on blood components and functions. Colloids Surf. B Biointerfaces 2019, 184, 110505. [Google Scholar] [CrossRef] [PubMed]
- Mörtberg, J.; Blombäck, M.; Wallén, Å.; He, S.; Jacobson, S.H.; Spaak, J. Increased fibrin formation and impaired fibrinolytic capacity in severe chronic kidney disease. Blood Coagul. Fibrinolysis 2016, 27, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.; Qi, X.; Xiang, Y.; You, S.; Cai, E.; Gao, T.; Tong, X.; Hu, R.; Shen, J.; Deng, H. Facile formation of injectable quaternized chitosan/tannic acid hydrogels with antibacterial and ROS scavenging capabilities for diabetic wound healing. Int. J. Biol. Macromol. 2022, 195, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Gulcin, İ. Antioxidants and antioxidant methods: An updated overview. Arch. Toxicol. 2020, 94, 651–715. [Google Scholar] [CrossRef] [Green Version]
- Jing, J.; He, Y.; Wang, Y.; Zeng, M. Inhibitory effects of Portulaca oleracea L. and selected flavonoid ingredients on heterocyclic amines in roast beef patties and Density Function Theory calculation of binding between heterocyclic amines intermediates and flavonoids. Food Chem. 2020, 336, 127551. [Google Scholar] [CrossRef]
- Spiegel, M.; Kapusta, K.; Kołodziejczyk, W.; Saloni, J.; Żbikowska, B.; Hill, G.A.; Sroka, Z. Antioxidant Activity of Selected Phenolic Acids–Ferric Reducing Antioxidant Power Assay and QSAR Analysis of the Structural Features. Molecules 2020, 25, 3088. [Google Scholar] [CrossRef]
- Sardana, A.; Kalra, S.; Khanna, D.; Balakumar, P. Nephroprotective effect of catechin on gentamicin-induced experimental nephrotoxicity. Clin. Exp. Nephrol. 2014, 19, 178–184. [Google Scholar] [CrossRef]
- Tanabe, K.; Tamura, Y.; Lanaspa, M.A.; Miyazaki, M.; Suzuki, N.; Sato, W.; Maeshima, Y.; Schreiner, G.F.; Villarreal, F.J.; Johnson, R.J.; et al. Epicatechin limits renal injury by mitochondrial protection in cisplatin nephropathy. Am. J. Physiol. Physiol. 2012, 303, F1264–F1274. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Jin, W.W.; Huang, M.; Ji, H.; Capen, D.E.; Xia, Y.; Yuan, J.; Păunescu, T.G.; Lu, H.A.J. Gentamicin-Induced Acute Kidney Injury in an Animal Model Involves Programmed Necrosis of the Collecting Duct. J. Am. Soc. Nephrol. 2020, 31, 2097–2115. [Google Scholar] [CrossRef] [PubMed]
- Akyol, S.; Ugurcu, V.; Altuntas, A.; Hasgul, R.; Cakmak, O.; Akyol, O. Caffeic Acid Phenethyl Ester as a Protective Agent against Nephrotoxicity and/or Oxidative Kidney Damage: A Detailed Systematic Review. Sci. World J. 2014, 2014, 561971. [Google Scholar] [CrossRef] [Green Version]
- Erseçkin, V.; Mert, H.; Irak, K.; Yildirim, S.; Mert, N. Nephroprotective effect of ferulic acid on gentamicin-induced nephrotoxicity in female rats. Drug Chem. Toxicol. 2020, 1–7. [Google Scholar] [CrossRef]
- Laaroussi, H.; Bakour, M.; Ousaaid, D.; Ferreira-Santos, P.; Genisheva, Z.; El Ghouizi, A.; Aboulghazi, A.; Teixeira, J.A.; Lyoussi, B. Protective Effect of Honey and Propolis against Gentamicin-Induced Oxidative Stress and Hepatorenal Damages. Oxidative Med. Cell. Longev. 2021, 2021, 9719906. [Google Scholar] [CrossRef] [PubMed]
- Kohanski, M.A.; Dwyer, D.J.; Hayete, B.; Lawrence, C.A.; Collins, J.J. A Common Mechanism of Cellular Death Induced by Bactericidal Antibiotics. Cell 2007, 130, 797–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bustos, P.; Ponzio, R.D.; Páez, P.L.; Cabrera, J.L.; Virgolini, M.B.; Ortega, M.G. Flavonoids as protective agents against oxidative stress induced by gentamicin in systemic circulation. Potent protective activity and microbial synergism of luteolin. Food Chem. Toxicol. 2018, 118, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Miroshnichenko, A.G.; Perfiliev, V.Y.; Lysenko, I.V.; Zhernakova, N.I. Interaction between some antibiotics and antioxidant. Res. Results Pharmacol. 2017, 3, 100–112. [Google Scholar] [CrossRef]
- Angiolella, L.; Sacchetti, G.; Efferth, T. Antimicrobial and Antioxidant Activities of Natural Compounds. Evid.-Based Complement. Altern. Med. 2018, 2018, 1945179. [Google Scholar] [CrossRef] [PubMed]
- Faleiro, M.L.; Miguel, G. Antimicrobial and Antioxidant Activities of Natural Compounds: Enhance the Safety and Quality of Food. Foods 2020, 9, 1145. [Google Scholar] [CrossRef]
- Krawitz, C.; Abu Mraheil, M.; Stein, M.; Imirzalioglu, C.; Domann, E.; Pleschka, S.; Hain, T. Inhibitory activity of a standardized elderberry liquid extract against clinically-relevant human respiratory bacterial pathogens and influenza A and B viruses. BMC Complement. Altern. Med. 2011, 11, 16. [Google Scholar] [CrossRef] [Green Version]
- Hearst, C.; Mccollum, G.; Nelson, D.W.; Ballard, L.M.; Millar, B.C.; Goldsmith, C.E.; Rooney, P.J.; Loughrey, A.C.; Moore, J.E.; Rao, J.R. Antibacterial activity of elder (Sambucus nigra L.) flower or berry against hospital pathogens. J. Med. Plants Res. 2010, 4, 1805–1809. [Google Scholar]
- Năsui, B.A.; Ungur, R.A.; Talaba, P.; Varlas, V.N.; Ciuciuc, N.; Silaghi, C.A.; Silaghi, H.; Opre, D.; Pop, A.L.l. Is Alcohol Consumption Related to Lifestyle Factors in Romanian University Students? Int. J. Environ. Res. Public Health 2021, 18, 1835. [Google Scholar] [CrossRef]
- Popović, B.; Šutić, I.; Marković, N.B. Nephrotoxic drugs. Acta Med. Croat. 2016, 70, 309–314. [Google Scholar]
- Irsay, L.; Checicheș, A.; Perja, D.; Borda, I.M.; Dogaru, G.; Onac, I.; Ungur, R.; Ciortea, V. Pharmacological pain management in patients with chronic kidney disease. Balneo Res. J. 2019, 10, 12–16. [Google Scholar] [CrossRef]
- Matzkin, E.G.; Curry, E.J.; Kong, Q.; Rogers, M.; Henry, M.; Smith, E.L. Efficacy and Treatment Response of Intra-articular Corticosteroid Injections in Patients With Symptomatic Knee Osteoarthritis. J. Am. Acad. Orthop. Surg. 2017, 25, 703–714. [Google Scholar] [CrossRef]
- Bannuru, R.R.; Osani, M.C.; Vaysbrot, E.E.; Arden, N.K.; Bennell, K.; Bierma-Zeinstra, S.M.A.; Kraus, V.B.; Lohmander, L.S.; Abbott, J.H.; Bhandari, M.; et al. OARSI guidelines for the non-surgical management of knee, hip, and polyarticular osteoarthritis. Osteoarthr. Cartil. 2019, 27, 1578–1589. [Google Scholar] [CrossRef] [Green Version]
- Ungur, R.A.; Florea, A.; Tăbăran, A.F.; Scurtu, I.C.; Onac, I.; Borda, I.M.; Irsay, L.; Ciortea, V.M.; Dronca, E.; Zdrenghea, M.T.; et al. Chondroprotective effects of pulsed shortwave therapy in rabbits with experimental osteoarthritis. Rom. J. Morphol. Embryol. 2017, 58, 465–472. [Google Scholar] [PubMed]
- Alschuler, L.; Weil, A.; Horwitz, R.; Stamets, P.; Chiasson, A.M.; Crocker, R.; Maizes, V. Integrative considerations during the COVID-19 pandemic. Explore 2020, 16, 354–356. [Google Scholar] [CrossRef] [PubMed]
- Barak, V. The effect of Sambucol, a black elderberry-based, natural product, on the production of human cytokines: I. Inflammatory cytokines. Eur. Cytokine Netw. 2001, 12, 290–296. [Google Scholar] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Polyphenolic Compounds | Amount of Polyphenolic Compounds (μg mL−1) | |
|---|---|---|
| X 1 ± St. Dev. 2 | ±St. Dev. 2 | |
| Flavanols | ||
| Catechin | 3.9 ± 0.3 | |
| Epicatechin | 9.0 ± 1.1 | |
| Flavonols | ||
| Quercetin-3-O-rutinoside (rutin) | 463.2 ± 0.0 | |
| Hydroxycinnamic acids | ||
| Caffeic acid | 3.6 ± 0.1 | |
| Ferulic acid | 1.5 ± 0.3 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ungur, R.A.; Borda, I.M.; Codea, R.A.; Ciortea, V.M.; Năsui, B.A.; Muste, S.; Sarpataky, O.; Filip, M.; Irsay, L.; Crăciun, E.C.; et al. A Flavonoid-Rich Extract of Sambucus nigra L. Reduced Lipid Peroxidation in a Rat Experimental Model of Gentamicin Nephrotoxicity. Materials 2022, 15, 772. https://doi.org/10.3390/ma15030772
Ungur RA, Borda IM, Codea RA, Ciortea VM, Năsui BA, Muste S, Sarpataky O, Filip M, Irsay L, Crăciun EC, et al. A Flavonoid-Rich Extract of Sambucus nigra L. Reduced Lipid Peroxidation in a Rat Experimental Model of Gentamicin Nephrotoxicity. Materials. 2022; 15(3):772. https://doi.org/10.3390/ma15030772
Chicago/Turabian StyleUngur, Rodica Ana, Ileana Monica Borda, Răzvan Andrei Codea, Viorela Mihaela Ciortea, Bogdana Adriana Năsui, Sevastița Muste, Orsolya Sarpataky, Miuța Filip, Laszlo Irsay, Elena Cristina Crăciun, and et al. 2022. "A Flavonoid-Rich Extract of Sambucus nigra L. Reduced Lipid Peroxidation in a Rat Experimental Model of Gentamicin Nephrotoxicity" Materials 15, no. 3: 772. https://doi.org/10.3390/ma15030772
APA StyleUngur, R. A., Borda, I. M., Codea, R. A., Ciortea, V. M., Năsui, B. A., Muste, S., Sarpataky, O., Filip, M., Irsay, L., Crăciun, E. C., Căinap, S., Jivănescu, D. B., Pop, A. L., Singurean, V. E., Crișan, M., Groza, O. B., & Martiș, G. S. (2022). A Flavonoid-Rich Extract of Sambucus nigra L. Reduced Lipid Peroxidation in a Rat Experimental Model of Gentamicin Nephrotoxicity. Materials, 15(3), 772. https://doi.org/10.3390/ma15030772