
Fighting Phytopathogens with Engineered Inorganic-Based Nanoparticles



Abstract
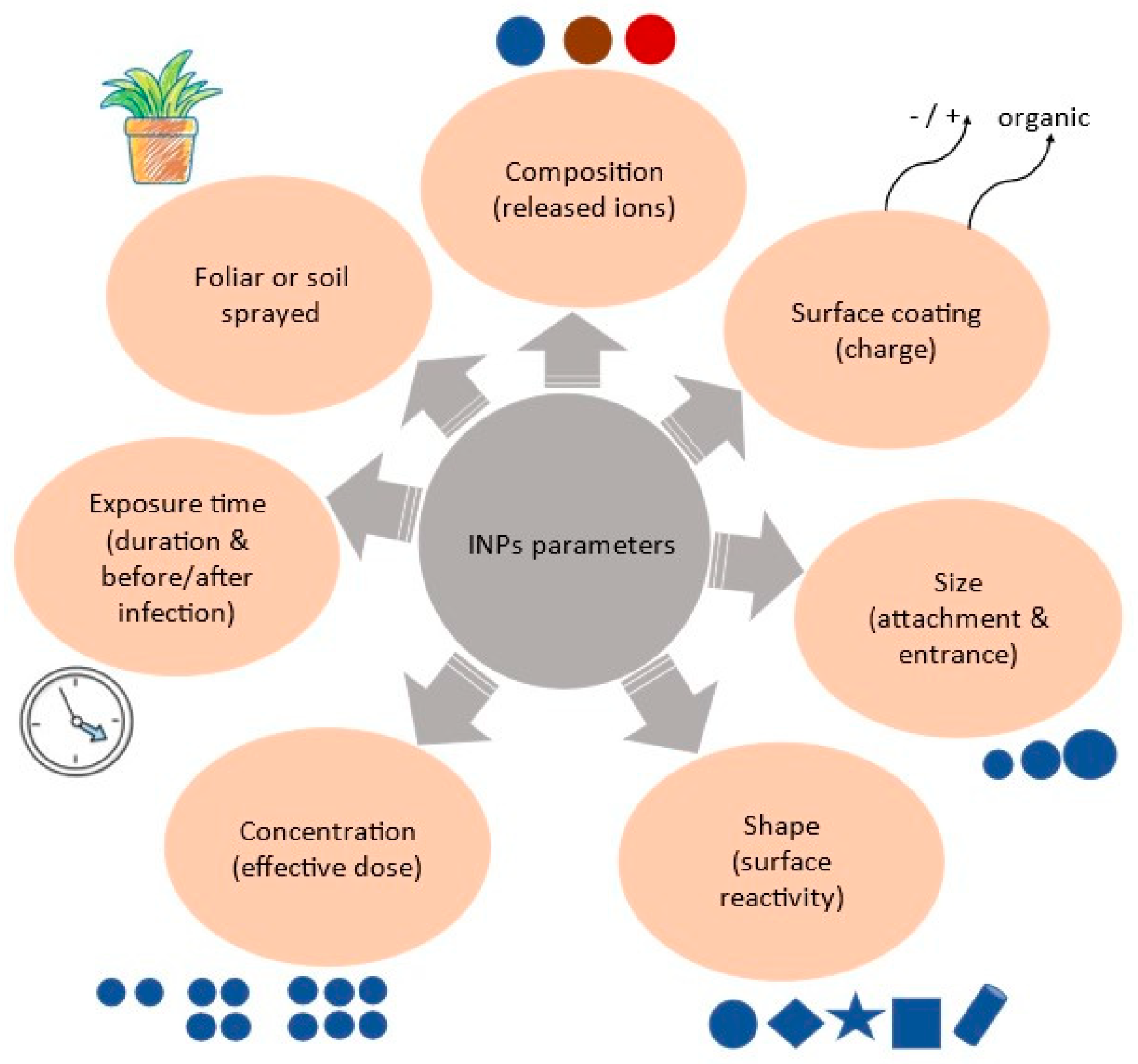
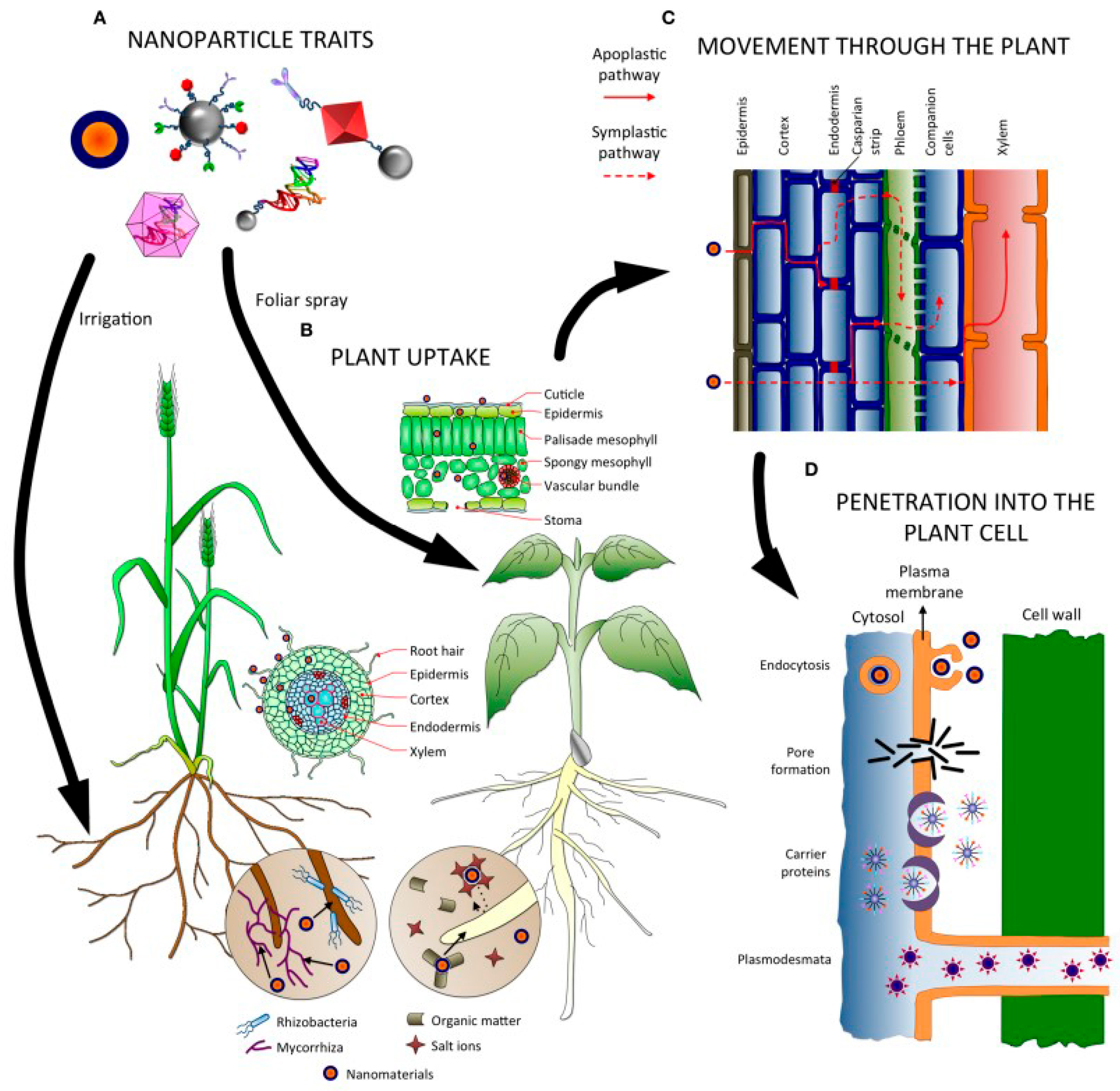
:1. Introduction
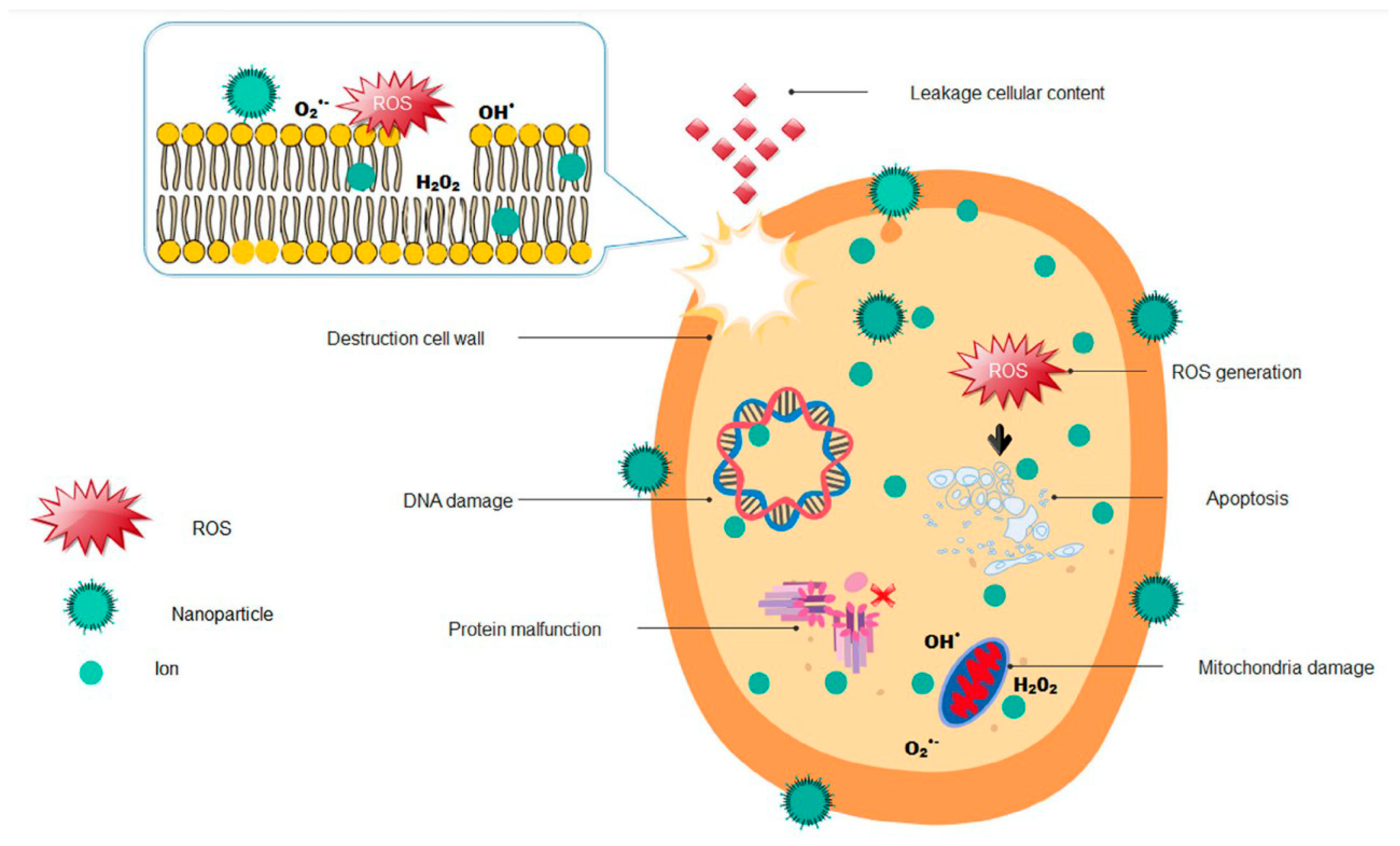
2. Mechanistic Aspects

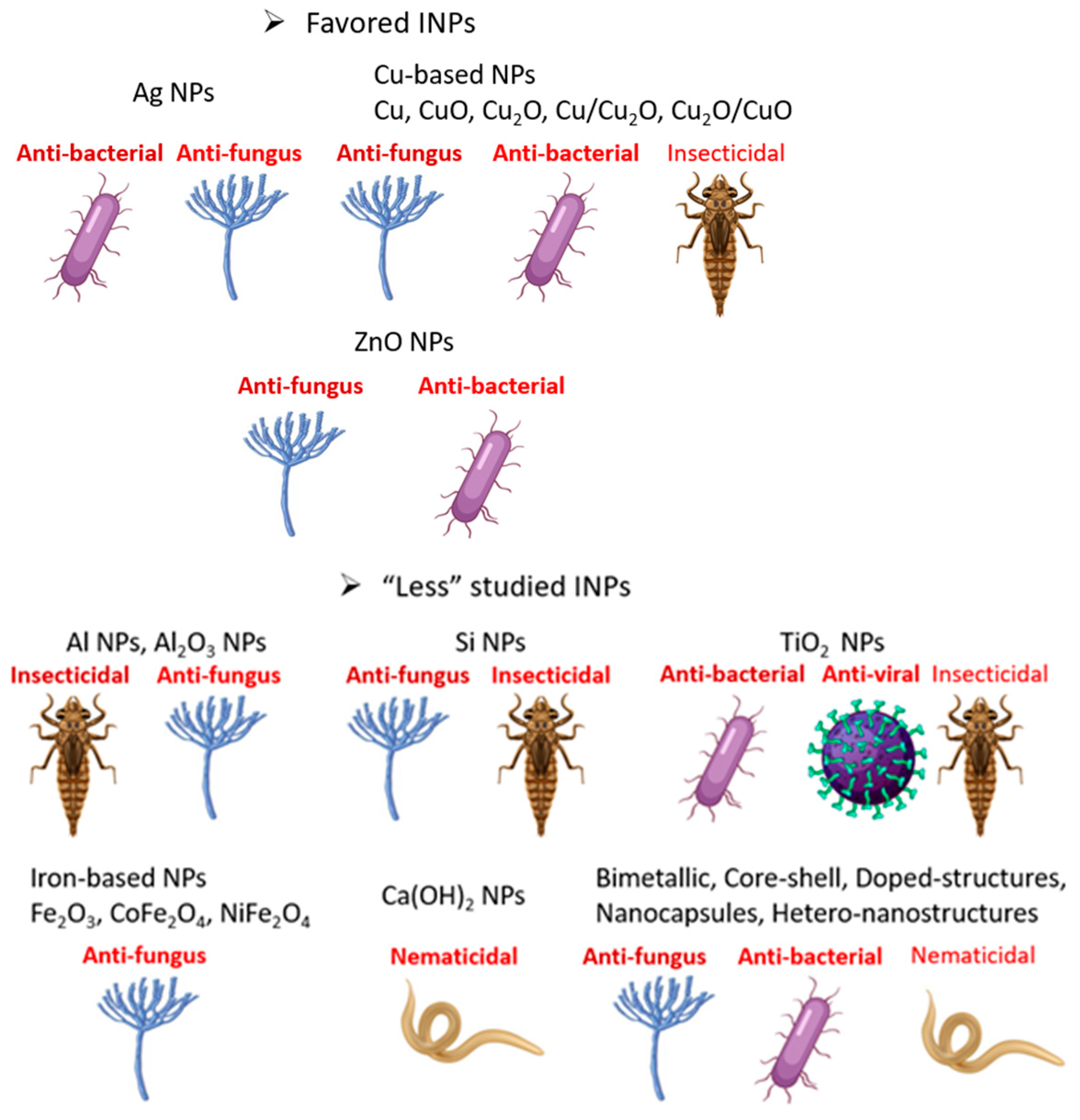
3. Favored INPs
3.1. Silver
3.1.1. Antifungal Effect
3.1.2. Antibacterial Effect
3.1.3. Antiviral Effect
3.2. Cu-Based NPs
3.2.1. Antifungal Effect
3.2.2. Antibacterial Effect
3.2.3. Insecticidal Effect
3.3. Zinc Oxide NPs
3.3.1. Antifungal Effect
3.3.2. Antibacterial Effect
3.3.3. Antiviral Effect
4. “Less” Studied INPs
- Antifungal effect
- Insecticidal effect
- Antibacterial effect
- Antiviral effect
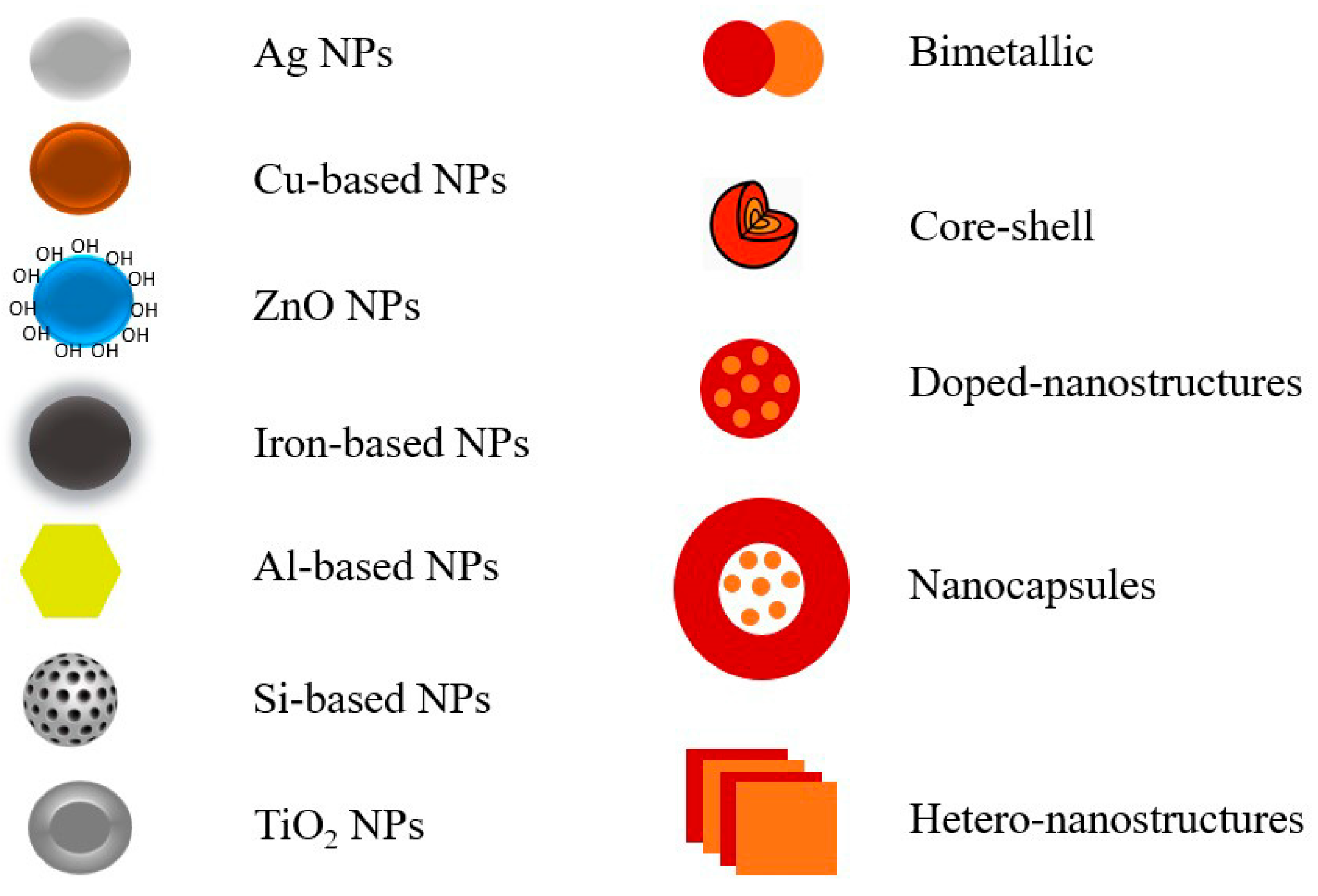
5. Advanced Inorganic-Based Nanostructures
6. Conclusions and Future Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Oerke, E.C.; Dehne, H.W. Global crop production and the efficacy of crop protection—Current situation and future trends. Eur. J. Plant Pathol. 1997, 103, 203–215. [Google Scholar] [CrossRef]
- Scherm, H.; Coakley, S.M. Plant pathogens in a changing world. Australas. Plant Pathol. 2003, 32, 157–165. [Google Scholar] [CrossRef]
- Ristaino, J.B.; Anderson, P.K.; Bebber, D.P.; Brauman, K.A.; Cunniffe, N.J.; Fedoroff, N.V.; Finegold, C.; Garrett, K.A.; Gilligan, C.A.; Jones, C.M.; et al. The persistent threat of emerging plant disease pandemics to global food security. Proc. Natl. Acad. Sci. USA 2021, 118, e2022239118. [Google Scholar] [CrossRef] [PubMed]
- Savary, S.; Ficke, A.; Aubertot, J.N.; Hollier, C. Crop losses due to diseases and their implications for global food production losses and food security. Food Secur. 2012, 4, 519–537. [Google Scholar] [CrossRef]
- Teng, P.S.; Shane, W.W.; MacKenzie, D.R. Crop losses due to plant pathogens. CRC Crit. Rev. Plant Sci. 1984, 2, 21–47. [Google Scholar] [CrossRef]
- Elkhouly, A.R.; Slama, T.A.; AlHireereeq, E.A. Survey of Global Crop Loss. Appl. Humanit. 2021. [Google Scholar]
- Abdollahdokht, D.; Gao, Y.; Faramarz, S.; Poustforoosh, A.; Abbasi, M.; Asadikaram, G.; Nematollahi, M.H. Conventional agrochemicals towards nano-biopesticides: An overview on recent advances. Chem. Biol. Technol. Agric. 2022, 9, 13. [Google Scholar] [CrossRef]
- Sharma, A.; Kumar, V.; Shahzad, B.; Tanveer, M.; Sidhu, G.P.S.; Handa, N.; Kohli, S.K.; Yadav, P.; Bali, A.S.; Parihar, R.D.; et al. Worldwide pesticide usage and its impacts on ecosystem. SN Appl. Sci. 2019, 1, 1446. [Google Scholar] [CrossRef] [Green Version]
- Raj, S.N.; Anooj, E.S.; Rajendran, K.; Vallinayagam, S. A comprehensive review on regulatory invention of nano pesticides in Agricultural nano formulation and food system. J. Mol. Struct. 2021, 1239, 130517. [Google Scholar] [CrossRef]
- Sekhon, B.S. Nanotechnology in agri-food production: An overview. Nanotechnol. Sci. Appl. 2014, 7, 31–53. [Google Scholar] [CrossRef] [Green Version]
- Kah, M.; Hofmann, T. Nanopesticide research: Current trends and future priorities. Environ. Int. 2014, 63, 224–235. [Google Scholar] [CrossRef]
- Kah, M.; Beulke, S.; Tiede, K.; Hofmann, T. Nanopesticides: State of knowledge, environmental fate, and exposure modeling. Crit. Rev. Environ. Sci. Technol. 2013, 43, 1823–1867. [Google Scholar] [CrossRef]
- Singh, H.; Sharma, A.; Bhardwaj, S.K.; Arya, S.K.; Bhardwaj, N.; Khatri, M. Recent advances in the applications of nano-agrochemicals for sustainable agricultural development. Environ. Sci. Process. Impacts 2021, 23, 213–239. [Google Scholar] [CrossRef]
- Horn, D.; Rieger, J. Organic nanoparticles in the aqueous phase—Theory, experiment, and use. Angew. Chem.-Int. Ed. 2001, 40, 4330–4361. [Google Scholar] [CrossRef]
- Hajipour, M.J.; Fromm, K.M.; Akbar Ashkarran, A.; Jimenez de Aberasturi, D.; Larramendi, I.R.; Rojo, T.; Serpooshan, V.; Parak, W.J.; Mahmoudi, M. Antibacterial properties of nanoparticles. Trends Biotechnol. 2012, 30, 499–511. [Google Scholar] [CrossRef] [Green Version]
- Schwab, F.; Zhai, G.; Kern, M.; Turner, A.; Schnoor, J.L.; Wiesner, M.R. Barriers, pathways and processes for uptake, translocation and accumulation of nanomaterials in plants—Critical review. Nanotoxicology 2016, 10, 257–278. [Google Scholar] [CrossRef]
- Iversen, T.G.; Skotland, T.; Sandvig, K. Endocytosis and intracellular transport of nanoparticles: Present knowledge and need for future studies. Nano Today 2011, 6, 176–185. [Google Scholar] [CrossRef]
- Salatin, S.; Dizaj, S.M.; Khosroushahi, A.Y. Effect of the surface modification, size, and shape on cellular uptake of nanoparticles. Cell Biol. Int. 2015, 39, 881–890. [Google Scholar] [CrossRef]
- Deng, J.; Gao, C. Recent advances in interactions of designed nanoparticles and cells with respect to cellular uptake, intracellular fate, degradation and cytotoxicity. Nanotechnology 2016, 27, 412002. [Google Scholar] [CrossRef]
- Suresh, A.K.; Pelletier, D.A.; Doktycz, M.J. Relating nanomaterial properties and microbial toxicity. Nanoscale 2013, 5, 463–474. [Google Scholar] [CrossRef]
- Sharon, M.; Choudhary, A.K.; Kumar, R. Nanotechnology in agricultural diseases and food safety. J. Phytol. 2010, 2, 83–92. [Google Scholar]
- Kettler, K.; Veltman, K.; van de Meent, D.; van Wezel, A.; Hendriks, A.J. Cellular uptake of nanoparticles as determined by particle properties, experimental conditions, and cell type. Environ. Toxicol. Chem. 2014, 33, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Foroozandeh, P.; Aziz, A.A. Insight into Cellular Uptake and Intracellular Trafficking of Nanoparticles. Nanoscale Res. Lett. 2018, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biondo, F.; Baldassarre, F.; Vergaro, V.; Ciccarella, G. Controlled biocide release from smart delivery systems: Materials engineering to tune release rate, biointeractions, and responsiveness. In Nanotechnology-Based Sustainable Alternatives for the Management of Plant Diseases; Balestra, G.M., Fortunati, E., Eds.; Elsevier: Amsterdam, The Netherlands, 2022; pp. 31–130. ISBN 978-0-12-823394-8. [Google Scholar]
- Donahue, N.D.; Handan, A.; Wilhelm, S. Concepts of nanoparticle cellular uptake, intracellular trafficking, and kinetics in nanomedicine. Adv. Drug Deliv. Rev. 2019, 143, 68–96. [Google Scholar] [CrossRef]
- Zhao, J.; Stenzel, M.H. Entry of nanoparticles into cells: The importance of nanoparticle properties. Polym. Chem. 2018, 9, 259–272. [Google Scholar] [CrossRef]
- Pérez-de-Luque, A. Interaction of Nanomaterials with Plants: What Do We Need for Real Applications in Agriculture? Front. Environ. Sci. 2017, 5, 12. [Google Scholar] [CrossRef] [Green Version]
- Bahrami, A.H.; Lipowsky, R.; Weikl, T.R. The role of membrane curvature for the wrapping of nanoparticles. Soft Matter 2016, 12, 581–587. [Google Scholar] [CrossRef] [Green Version]
- Slavin, Y.N.; Asnis, J.; Häfeli, U.O.; Bach, H. Metal nanoparticles: Understanding the mechanisms behind antibacterial activity. J. Nanotechnol. 2017, 15, 1–20. [Google Scholar] [CrossRef]
- Cruz-Luna, A.R.; Cruz-Martínez, H.; Vásquez-López, A.; Medina, D.I. Metal Nanoparticles as Novel Antifungal Agents for Sustainable Agriculture: Current Advances and Future Directions. J. Fungi 2021, 7, 1033. [Google Scholar] [CrossRef]
- Mohamed, M.A.; Abd–Elsalam, K.A. Nanoantimicrobials for Plant Pathogens Control: Potential Applications and Mechanistic Aspects. In Nanobiotechnology Applications in Plant Protection; Springer: Berlin/Heidelberg, Germany, 2018; pp. 87–109. ISBN 978-3-319-91160-1. [Google Scholar]
- Ghosh, S.K.; Bera, T. Unraveling the mechanism of nanoparticles for controlling plant pathogens and pests. In Advances in Nano-Fertilizers and Nano-Pesticides in Agriculture; Elsevier: Amsterdam, The Netherlands, 2021; ISBN 978-0-12-820444-3. [Google Scholar]
- Geoghegan, I.; Steinberg, G.; Gurr, S. The Role of the Fungal Cell Wall in the Infection of Plants Unique Aspects of the Cell Wall in Phytopathogenic Fungi. Trends Microbiol. 2017, 25, 957–967. [Google Scholar] [CrossRef]
- Alfei, S.; Schito, A.M. Positively Charged Polymers as Promising Devices against Multidrug Resistant Gram-Negative Bacteria: A Review. Polymers 2020, 12, 1195. [Google Scholar] [CrossRef]
- Hernández-Díaz, J.A.; Garza-García, J.J.; Zamudio-Ojeda, A.; León-Morales, J.M.; López-Velázquez, J.C.; García-Morales, S. Plant-mediated synthesis of nanoparticles and their antimicrobial activity against phytopathogens. J. Sci. Food Agric. 2020, 101, 1270–1287. [Google Scholar] [CrossRef]
- Pal, S.; Tak, Y.K.; Song, J.M. Does the Antibacterial Activity of Silver Nanoparticles Depend on the Shape of the Nanoparticle? A Study of the Gram-Negative Bacterium Escherichia coli. Appl. Environ. Microbiol. 2007, 73, 1712–1720. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; He, H.; Yu, Y.; Liu, S.; Zhang, C.; He, L. Morphology-dependent bactericidal activities of Ag/CeO2 catalysts against Escherichia coli. J. Inorg. Biochem. 2014, 135, 45–53. [Google Scholar] [CrossRef]
- Vargas-Hernandez, M.; Macias-Bobadilla, I.; Guevara-Gonzalez, R.G.; Rico-Garcia, E.; Ocampo-Velazquez, R.V.; Avila-Juarez, L.; Torres-Pacheco, I. Nanoparticles as Potential Antivirals in Agriculture. Agriculture 2020, 10, 444. [Google Scholar] [CrossRef]
- Arkhipenko, M.V.; Nikitin, N.A.; Baranov, O.A.; Evtushenko, E.A.; Atabekov, J.G.; Karpova, O.V. Surface charge mapping on virions and virus-like particles of helical plant viruses. Acta Nat. 2019, 11, 73–78. [Google Scholar] [CrossRef]
- Farooq, T.; Adeel, M.; He, Z.; Umar, M.; Shakoor, N.; da Silva, W.; Elmer, W.; White, J.C.; Rui, Y. Nanotechnology and Plant Viruses: An Emerging Disease Management Approach for Resistant Pathogens. ACS Nano 2021, 15, 6030–6037. [Google Scholar] [CrossRef]
- Cai, L.; Cai, L.; Jia, H.; Liu, C.; Wang, D.; Sun, X. Foliar exposure of Fe3O4 nanoparticles on Nicotiana benthamiana: Evidence for nanoparticles uptake, plant growth promoter and defense response elicitor against plant virus. J. Hazard. Mater. 2020, 393, 122415. [Google Scholar] [CrossRef]
- Zhao, X.; Cui, H.; Wang, Y.; Sun, C.; Cui, B.; Zeng, Z. Development strategies and prospects of nano-based smart pesticide formulation. J. Agric. Food Chem. 2017, 66, 6504–6512. [Google Scholar] [CrossRef]
- Benelli, G. Mode of action of nanoparticles against insects. Environ. Sci. Pollut. Res. 2018, 25, 12329–12341. [Google Scholar] [CrossRef]
- Tariq, M.; Mohammad, K.N.; Ahmed, B.; Siddiqui, M.A.; Lee, J. Biological Synthesis of Silver Nanoparticles and Prospects in Plant Disease Management. Molecules 2022, 27, 4754. [Google Scholar] [CrossRef] [PubMed]
- Co-Optimizing Solutions|Annex E|Nanotech Pesticides Possible Breakthroughs Nanotech Pesticides. 1–6. Available online: https://www.wbcsd.org/contentwbc/download/416/4430/1 (accessed on 5 February 2023).
- Mansoor, S.; Zahoor, I.; Baba, T.R.; Padder, S.A.; Bhat, Z.A.; Koul, A.M.; Jiang, L. Fabrication of Silver Nanoparticles against Fungal Pathogens. Front. Nanotechnol. 2021, 3, 679358. [Google Scholar] [CrossRef]
- Gharpure, S.; Akash, A.; Ankamwar, B. A Review on Antimicrobial Properties of Metal Nanoparticles. J. Nanosci. Nanotechnol. 2019, 20, 3303–3339. [Google Scholar] [CrossRef] [PubMed]
- Sotiriou, G.A.; Meyer, A.; Knijnenburg, J.T.N.; Panke, S.; Pratsinis, S.E. Quantifying the origin of released Ag+ ions from nanosilver. Langmuir 2012, 28, 15929–15936. [Google Scholar] [CrossRef]
- Dakal, T.C.; Kumar, A.; Majumdar, R.S.; Yadav, V. Mechanistic basis of antimicrobial actions of silver nanoparticles. Front. Microbiol. 2016, 7, 1831. [Google Scholar] [CrossRef] [Green Version]
- Morones, J.R.; Elechiguerra, J.L.; Camacho, A.; Holt, K.; Kouri, J.B.; Ramırez, J.T.; Yacaman, M.J. The bactericidal effect of silver nanoparticles. Nanotechnology 2005, 16, 2346–2353. [Google Scholar] [CrossRef] [Green Version]
- Elgorban, A.M.; El-Samawaty, A.E.-R.M.; Yassin, M.A.; Sayed, S.; Adil, S.F.; Elhindi, K.M.; Bakri, M.; Khan, M. Antifungal silver nanoparticles: Synthesis, characterization and biological evaluation. Biotechnol. Biotechnol. Equip. 2015. [Google Scholar] [CrossRef] [Green Version]
- Mendes, J.E.; Arunhosa, L.; Teixeira, J.A.; de Camargo, E.R.; de SOuza, C.P.; Pessoa, J.D.C. Antifungal activity of silver colloidal nanoparticles against phytopathogenic fungus (Phomopsis sp.) in soybean seeds. Int. J. Biol. Vet. Agric. Food Eng. 2014, 8, 928–933. [Google Scholar]
- Aguilar-Méndez, M.A.; Martín-Martínez, E.S.; Ortega-Arroyo, L.; Cobián-Portillo, G.; Sánchez-Espíndola, E. Synthesis and characterization of silver nanoparticles: Effect on phytopathogen Colletotrichum gloesporioides. J. Nanopart. Res. 2011, 13, 2525–2532. [Google Scholar] [CrossRef]
- Vrandečić, K.; Ćosić, J.; Ilić, J.; Ravnjak, B.; Selmani, A.; Galić, E.; Pem, B.; Barbir, R.; Vrček, I.V.; Vinković, T. Antifungal activities of silver and selenium nanoparticles stabilized with different surface coating agents. Pest Manag. Sci. 2020, 76, 2021–2029. [Google Scholar] [CrossRef]
- Kasprowicz, M.J.; Kozioł, M.; Gorczyca, A. The effect of silver nanoparticles on phytopathogenic spores of Fusarium culmorum. Can. J. Microbiol. 2010, 56, 247–253. [Google Scholar] [CrossRef]
- Tarazonaa, A.; Gómez, J.V.; Mateo, E.M.; Jiménez, M.; Mateo, F. Antifungal effect of engineered silver nanoparticles on phytopathogenic and toxigenic Fusarium spp. and their impact on mycotoxin accumulation. Int. J. Food Microbiol. 2019, 306, 108259. [Google Scholar] [CrossRef] [PubMed]
- Jo, Y.K.; Kim, B.H.; Jung, G. Antifungal activity of silver ions and nanoparticles on phytopathogenic fungi. Plant Dis. 2009, 93, 1037–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jo, Y.K.; Cromwell, W.; Jeong, H.K.; Thorkelson, J.; Roh, J.H.; Shin, D.B. Use of silver nanoparticles for managing Gibberella fujikuroi on rice seedlings. Crop Prot. 2015, 74, 65–69. [Google Scholar] [CrossRef]
- Dzimitrowicz, A.; Motyka, A.; Jamroz, P.; Lojkowska, E.; Babinska, W.; Terefinko, D.; Pohl, P.; Sledz, W. Application of silver nanostructures synthesized by cold atmospheric pressure plasma for inactivation of bacterial phytopathogens from the genera Dickeya and Pectobacterium. Materials 2018, 9, 331. [Google Scholar] [CrossRef] [Green Version]
- Dzimitrowicz, A.; Motyka-Pomagruk, A.; Cyganowski, P.; Babinska, W.; Terefinko, D.; Jamroz, P.; Lojkowska, E.; Pohl, P.; Sledz, W. Antibacterial Activity of Fructose-Stabilized Silver Nanoparticles Produced by Direct Current Atmospheric Pressure Glow Discharge towards Quarantine Pests. Nanomaterials 2018, 8, 751. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Li, S.; Luo, J.; Wang, R.; Ding, W. Enhancement of the Antibacterial Activity of Silver Nanoparticles against Phytopathogenic Bacterium Ralstonia solanacearum by Stabilization. J. Nanomater. 2016, 2016, 7135852. [Google Scholar] [CrossRef] [Green Version]
- Makarovsky, D.; Fadeev, L.; Salam, B.B.; Zelinger, E.; Matan, O.; Inbar, J.; Jurkevitch, E.; Gozin, M.; Burdman, S. Silver Nanoparticles Complexed with Bovine Submaxillary Mucin Possess Strong Antibacterial Activity and Protect against Seedling Infection. Appl. Environ. Microbiol. 2018, 84, e02212-17. [Google Scholar] [CrossRef] [Green Version]
- El-shazly, M.A.; Attia, Y.A.; Kabil, F.F.; Anis, E.; Hazman, M. Inhibitory Effects of Salicylic Acid and Silver Nanoparticles on Potato Virus Y-Infected Potato Plants in Egypt. Middle East J. Agric. Res. 2017, 6, 835–848. [Google Scholar]
- El-Dougdoug, N.K.; Bondok, A.M.; El-Dougdoug, K.A. Evaluation of Silver Nanoparticles as Antiviral Agent against ToMV and PVY in Tomato Plants. Middle East J. Appl. Sci. 2018, 8, 100–111. [Google Scholar]
- Shafie, R.M.; Salama, A.M.; Farroh, K.Y. Silver nanoparticles activity against Tomato spotted wilt virus. Middle East J. Agric. Res. 2018, 7, 1251–1267. [Google Scholar]
- El Gamal, A.Y.; Tohamy, M.R.; Abou-Zaid, M.I.; Atia, M.M.; El Sayed, T.; Farroh, K.Y. Silver nanoparticles as a viricidal agent to inhibit plant-infecting viruses and disrupt their acquisition and transmission by their aphid vector. Arch. Virol. 2022, 167, 85–97. [Google Scholar] [CrossRef]
- Vincent, M.; Hartemann, P.; Engels-Deutsch, M. Antimicrobial applications of copper. J. Hyg. Environ. Health 2016, 219, 585–591. [Google Scholar] [CrossRef]
- Ermini, M.L.; Voliani, V. Antimicrobial Nano-Agents: The Copper Age. ACS Nano 2021, 15, 6008–6029. [Google Scholar] [CrossRef]
- Lemire, J.A.; Harrison, J.J.; Turner, R.J. Antimicrobial activity of metals: Mechanisms, molecular targets and applications. Nat. Rev. Microbiol. 2013, 11, 371–384. [Google Scholar] [CrossRef]
- Giannousi, K.; Avramidis, I.; Dendrinou-Samara, C. Synthesis, characterization and evaluation of copper based nanoparticles as agrochemicals against Phytophthora infestans. RSC Adv. 2013, 3, 21743–21752. [Google Scholar] [CrossRef]
- Giannousi, K.; Pantazaki, A.; Dendrinou-Samara, C. Copper based Nanoparticles as Antimicrobials. In Nanostructures for Antimicrobial Therapy; Elsevier: Amsterdam, The Netherlands, 2017; pp. 515–529. [Google Scholar]
- NanoCu Commercial Product. Available online: https://product.statnano.com/product/10944/nanocu (accessed on 5 February 2023).
- Chawla, L.S.; Beers-Mulroy, B.; Tidmarsh, G.F. Therapeutic Opportunities for Hepcidin in Acute Care Medicine. Crit. Care Clin. 2019, 35, 357–374. [Google Scholar] [CrossRef] [Green Version]
- Majumdar, T.D.; Singh, M.; Thapa, M.; Dutta, M.; Mukherjee, A.; Ghosh, C.K. Size-dependent antibacterial activity of copper nanoparticles against Xanthomonas oryzae pv. oryzae—A synthetic and mechanistic approach. Colloid Interface Sci. Commun. 2019, 2019, 100190. [Google Scholar] [CrossRef]
- Chatterjee, A.K.; Chakraborty, R.; Basu, T. Mechanism of antibacterial activity of copper nanoparticles. Nanotechnology 2014, 25, 135101. [Google Scholar] [CrossRef]
- Chang, Y.N.; Zhang, M.; Xia, L.; Zhang, J.; Xing, G. The Toxic Effects and Mechanisms of CuO and ZnO Nanoparticles. Materials 2012, 5, 2850–2871. [Google Scholar] [CrossRef] [Green Version]
- Naz, S.; Gul, A.; Zia, M. Toxicity of copper oxide nanoparticles: A review study. IET Nanobiotechnol. 2019, 14. [Google Scholar] [CrossRef] [PubMed]
- Crisan, M.C.; Teodora, M.; Lucian, M. Copper Nanoparticles: Synthesis and Characterization, Physiology, Toxicity and Antimicrobial Applications. Appl. Sci. 2021, 12, 141. [Google Scholar] [CrossRef]
- Singh, M.; Jampaiah, D.; Kandjani, A.E.; Sabri, Y.M.; Della Gaspera, E.; Reineck, P.; Judd, M.; Langley, J.; Cox, N.; Van Embden, J.; et al. Oxygen-deficient photostable Cu2O for enhanced visible light photocatalytic activity. Nanoscale 2018, 10, 6039–6050. [Google Scholar] [CrossRef] [PubMed]
- Malandrakis, A.A.; Kavroulakis, N.; Chrysikopoulos, C.V. Use of copper, silver and zinc nanoparticles against foliar and soil-borne plant pathogens. Sci. Total Environ. 2019, 670, 292–299. [Google Scholar] [CrossRef]
- Quiterio-Gutiérrez, T.; Ortega-Ortiz, H.; Cadenas-Pliego, G.; Hernández-Fuentes, A.D.; Sandoval-Rangel, A.; Benavides-Mendoza, A.; Cabrera-De La Fuente, M.; Juárez-Maldonado, A. The application of selenium and copper nanoparticles modifies the biochemical responses of tomato plants under stress by Alternaria Solani. Int. J. Mol. Sci. 2019, 20, 1950. [Google Scholar] [CrossRef] [Green Version]
- Maqsood, S.; Qadir, S.; Hussain, A.; Asghar, A.; Saleem, R.; Zaheer, S.; Nayyar, N. Antifungal Properties of Copper Nanoparticles against Aspergillus niger. Sch. Int. J. Biochem. 2020, 3, 87–91. [Google Scholar] [CrossRef]
- Pham, N.D.; Duong, M.M.; Le, M.V.; Hoang, H.A.; Pham, L.K.O. Preparation and characterization of antifungal colloidal copper nanoparticles and their antifungal activity against Fusarium oxysporum and Phytophthora capsici. C. R. Chim. 2019, 22, 786–793. [Google Scholar] [CrossRef]
- Mohamed, E.A. Non-Dependency of In Vitro Fungicidal Efficiency of Copper Nanoparticles against Fusarium oxysporum upon Particle Size. J. Plant Pathol. Microbiol. 2018, 9, 1000432. [Google Scholar] [CrossRef]
- Ntasiou, P.; Kaldeli Kerou, A.; Karamanidou, T.; Vlachou, A.; Tziros, G.T.; Tsouknidas, A.; Karaoglanidis, G.S. Synthesis and Characterization of Novel Copper Nanoparticles for the Control of Leaf Spot and Anthracnose Diseases of Olive. Nanomaterials 2021, 11, 1667. [Google Scholar] [CrossRef]
- Kanhed, P.; Birla, S.; Gaikwad, S.; Gade, A.; Seabra, A.B.; Rubilar, O.; Duran, N.; Rai, M. In vitro antifungal efficacy of copper nanoparticles against selected crop pathogenic fungi. Mater. Lett. 2014, 115, 13–17. [Google Scholar] [CrossRef]
- Van Viet, P.; Nguyen, H.T.; Cao, T.M.; Van Hieu, L. Fusarium Antifungal Activities of Copper Nanoparticles Synthesized by a Chemical Reduction Method. J. Nanomater. 2016, 2016, 1957612. [Google Scholar] [CrossRef] [Green Version]
- Bramhanwade, K.; Shende, S.; Bonde, S.; Gade, A.; Rai, M. Fungicidal activity of Cu nanoparticles against Fusarium causing crop diseases. Environ. Chem. Lett. 2016, 14, 229–235. [Google Scholar] [CrossRef]
- Giannousi, K.; Sarafidis, G.; Mourdikoudis, S.; Pantazaki, A.; Dendrinou-Samara, C. Selective synthesis of Cu2O and Cu/Cu2O NPs: Antifungal activity to yeast saccharomyces cerevisiae and DNA interaction. Inorg. Chem. 2014, 53, 9657–9666. [Google Scholar] [CrossRef]
- Hermida-Montero, L.A.; Pariona, N.; Mtz-Enriquez, A.I.; Carrión, G.; Paraguay-Delgado, F.; Rosas-Saito, G. Aqueous-phase synthesis of nanoparticles of copper/copper oxides and their antifungal effect against Fusarium oxysporum. J. Hazard. Mater. 2019, 380, 120850. [Google Scholar] [CrossRef]
- Chakraborty, P.; Adhikary, J.; Chatterjee, S.; Biswas, B.; Chattopadhyay, T. Facile syntheses of copper sulfide nanoparticles: Antibacterial and antifungal activity study. Rasayan J. Chem. 2016, 9, 77–83. [Google Scholar]
- Varympopi, A.; Dimopoulou, A.; Theologidis, I.; Karamanidou, T.; Kaldeli Kerou, A.; Vlachou, A.; Karfaridis, D.; Papafotis, D.; Hatzinikolaou, D.G.; Tsouknidas, A.; et al. Bactericides based on copper nanoparticles restrain growth of important plant pathogens. Pathogens 2020, 9, 1024. [Google Scholar] [CrossRef]
- Giannousi, K.; Lafazanis, K.; Arvanitidis, J.; Pantazaki, A.; Dendrinou-Samara, C. Hydrothermal synthesis of copper based nanoparticles: Antimicrobial screening and interaction with DNA. J. Inorg. Biochem. 2014, 133, 24–32. [Google Scholar] [CrossRef]
- Gkanatsiou, C.; Karamanoli, K.; Menkissoglu-Spiroudi, U.; Dendrinou-Samara, C. Composition effect of Cu-based nanoparticles on phytopathogenic bacteria. Antibacterial studies and phytotoxicity evaluation. Polyhedron 2019, 170, 395–403. [Google Scholar] [CrossRef]
- Rose, M.J.; Aron, S.A.; Janicki, B.W. Effect of various nonionic surfactants on growth of Escherichia coli. J. Bacteriol. 1966, 91, 1863–1868. [Google Scholar] [CrossRef] [Green Version]
- Majumdar, T.D.; Ghosh, C.K.; Mukherjee, A. Dual Role of Copper Nanoparticles in Bacterial Leaf Blight-Infected Rice: A Therapeutic and Metabolic Approach. ACS Agric. Sci. Technol. 2021, 1, 160–172. [Google Scholar] [CrossRef]
- Ayoub, H.; Khairy, M.; Elsaid, S.; Rashwan, F.A.; Abdel-Hafezb, H.F. Pesticidal Activity of Nanostructured Metal Oxides for Generation of Alternative Pesticide Formulations. J. Agric. Food Chem. 2018, 66, 5491–5498. [Google Scholar] [CrossRef] [PubMed]
- Sabir, S.; Arshad, M.; Chaudhari, S.K. Zinc oxide nanoparticles for revolutionizing agriculture: Synthesis and applications. Sci. World J. 2014, 2014, 925494. [Google Scholar] [CrossRef] [Green Version]
- Sun, Q.; Li, J.; Le, T. Zinc Oxide Nanoparticle as a Novel Class of Antifungal Agents: Current Advances and Future Perspectives. J. Agric. Food Chem. 2018, 66, 11209–11220. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Liu, Y.; Zhang, P.; Zhou, P.; Wu, Z.; Lou, B.; Jiang, Y.; Shakoor, N.; Li, M.; Li, Y.; et al. Engineered Zn-based nano-pesticides as an opportunity for treatment of phytopathogens in agriculture. NanoImpact 2022, 28, 100420. [Google Scholar] [CrossRef] [PubMed]
- Cabot, C.; Martos, S.; Llugany, M.; Gallego, B.; Tolrà, R.; Poschenrieder, C. A Role for Zinc in Plant Defense against Pathogens and Herbivores. Front. Plant Sci. 2019, 10, 1171. [Google Scholar] [CrossRef]
- Król, A.; Pomastowski, P.; Rafińska, K.; Railean-Plugaru, V.; Buszewski, B. Zinc oxide nanoparticles: Synthesis, antiseptic activity and toxicity mechanism. Adv. Colloid Interface Sci. 2017, 249, 37–52. [Google Scholar] [CrossRef]
- Arciniegas-Grijalba, P.A.; Patiño-Portela, M.C.; Mosquera-Sánchez, L.P.; Guerrero-Vargas, J.A.; Rodríguez-Páez, J.E. ZnO nanoparticles (ZnO-NPs) and their antifungal activity against coffee fungus Erythricium salmonicolor. Appl. Nanosci. 2017, 7, 225–241. [Google Scholar] [CrossRef] [Green Version]
- Savi, G.D.; Piacentini, K.C.; de Souza, S.R.; Costa, M.E.B.; Santos, C.M.R.; Scussel, V.M. Efficacy of zinc compounds in controlling Fusarium head blight and deoxynivalenol formation in wheat (Triticum aestivum L.). Int. J. Food Microbiol. 2015, 205, 98–104. [Google Scholar] [CrossRef]
- Dimkpa, C.O.; McLean, J.E.; Britt, D.W.; Anderson, A.J. Antifungal activity of ZnO nanoparticles and their interactive effect with a biocontrol bacterium on growth antagonism of the plant pathogen Fusarium graminearum. BioMetals 2013, 26, 913–924. [Google Scholar] [CrossRef]
- Pariona, N.; Paraguay-Delgado, F.; Basurto-Cereceda, S.; Morales-Mendoza, J.E.; Hermida-Montero, L.A.; Mtz-Enriquez, A.I. Shape-dependent antifungal activity of ZnO particles against Phytopathogenic fungi. Appl. Nanosci. 2019, 10, 435–443. [Google Scholar] [CrossRef]
- Mosquera-Sánchez, L.P.; Arciniegas-Grijalba, P.A.; Patiño-Portela, M.C.; Guerra-Sierra, B.E.; Muñoz-Florez, J.E.; Rodríguez-Páez, J.E. Antifungal effect of zinc oxide nanoparticles (ZnO-NPs) on Colletotrichum sp., causal agent of anthracnose in coffee crops. Biocatal. Agric. Biotechnol. 2020, 25, 101579. [Google Scholar] [CrossRef]
- Arciniegas-Grijalba, P.A.; Patiño-Portela, M.C.; Mosquera-Sánchez, L.P.; Guerra Sierra, B.E.; Muñoz-Florez, J.E.; Erazo-Castillo, L.A.; Rodríguez-Páez, J.E. ZnO-based nanofungicides: Synthesis, characterization and their effect on the coffee fungi Mycena citricolor and Colletotrichum sp. Mater. Sci. Eng. C 2019, 98, 808–825. [Google Scholar] [CrossRef]
- Akpomie, K.G.; Ghosh, S.; Gryzenhout, M.; Conradie, J. One-pot synthesis of zinc oxide nanoparticles via chemical precipitation for bromophenol blue adsorption and the antifungal activity against Filamentous fungi. Sci. Rep. 2021, 11, 8305. [Google Scholar] [CrossRef]
- Gondal, M.A.; Alzahrani, A.J.; Randhawa, M.A.; Siddiqui, M.N. Morphology and antifungal effect of nano-ZnO and nano-Pd-doped nano-ZnO against Aspergillus and Candida. J. Environ. Sci. Health-Part A Toxic/Hazard. Subst. Environ. Eng. 2012, 47, 1413–1418. [Google Scholar] [CrossRef]
- Hui, A.; Liu, J.; Ma, J. Synthesis and morphology-dependent antimicrobial activity of cerium doped flower-shaped ZnO crystallites under visible light irradiation. Colloids Surf. A Physicochem. Eng. Asp. 2016, 506, 519–525. [Google Scholar] [CrossRef]
- Parveen, A.; Siddiqui, Z.A. Zinc oxide nanoparticles affect growth, photosynthetic pigments, proline content and bacterial and fungal diseases of tomato. Arch. Phytopathol. Plant Prot. 2021, 54, 1519–1538. [Google Scholar] [CrossRef]
- Khan, M.R.; Siddiqui, Z.A. Role of zinc oxide nanoparticles in the management of disease complex of beetroot (Beta vulgaris L.) caused by Pectobacterium betavasculorum, Meloidogyne incognita and Rhizoctonia solani. Hortic. Environ. Biotechnol. 2021, 62, 225–241. [Google Scholar] [CrossRef]
- Elsharkawy, M.; Derbalah, A.; Hamza, A.; El-Shaer, A. Zinc oxide nanostructures as a control strategy of bacterial speck of tomato caused by Pseudomonas syringae in Egypt. Environ. Sci. Pollut. Res. 2020, 27, 19049–19057. [Google Scholar] [CrossRef]
- Cai, L.; Liu, C.; Fan, G.; Liu, C.; Sun, X. Preventing viral disease by ZnONPs through directly deactivating TMV and activating plant immunity in: Nicotiana benthamiana. Environ. Sci. Nano 2019, 6, 3653–3669. [Google Scholar] [CrossRef]
- Sofy, A.R.; Sofy, M.R.; Hmed, A.A.; Dawoud, R.A.; Alnaggar, A.E.A.M.; Soliman, A.M.; El-Dougdoug, N.K. Ameliorating the adverse effects of tomato mosaic tobamovirus infecting tomato plants in Egypt by boosting immunity in tomato plants using zinc oxide nanoparticles. Molecules 2021, 26, 1337. [Google Scholar] [CrossRef]
- Tryfon, P.; Antonoglou, O.; Vourlias, G.; Mourdikoudis, S.; Menkissoglu-Spiroudi, U.; Dendrinou-Samara, C. Tailoring Ca-Based Nanoparticles by Polyol Process for Use as Nematicidals and pH Adjusters in Agriculture. ACS Appl. Nano Mater. 2019, 2, 3870–3881. [Google Scholar] [CrossRef]
- Parveen, S.; Wani, A.H.; Shah, M.A.; Devi, H.S.; Bhat, M.Y.; Koka, J.A. Preparation, characterization and antifungal activity of iron oxide nanoparticles. Microb. Pathog. 2018, 115, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Sharma, A.; Sharma, M.; Bhalla, N.; Estrela, P.; Jain, A.; Thakur, P.; Thakur, A. Nanomaterial Fungicides: In Vitro and In Vivo Antimycotic Activity of Cobalt and Nickel Nanoferrites on Phytopathogenic Fungi. Glob. Chall. 2017, 1, 1700041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shenashen, M.; Derbalah, A.; Hamza, A.; Mohamed, A.; El Safty, S. Antifungal activity of fabricated mesoporous alumina nanoparticles against root rot disease of tomato caused by Fusarium oxysporium. Pest Manag. Sci. 2017, 73, 1121–1126. [Google Scholar] [CrossRef] [PubMed]
- Derbalah, A.; Shenashen, M.; Hamza, A.; Mohamed, A.; El Safty, S. Antifungal activity of fabricated mesoporous silica nanoparticles against early blight of tomato. Egypt. J. Basic Appl. Sci. 2018, 5, 145–150. [Google Scholar] [CrossRef] [Green Version]
- Buteler, M.; Lopez Garcia, G.; Stadler, T. Potential of nanostructured alumina for leaf-cutting ants Acromyrmex lobicornis (Hymenoptera: Formicidae) management. Austral Entomol. 2018, 57, 292–296. [Google Scholar] [CrossRef]
- Stadler, T.; Buteler, M.; Weaver, D.K. Novel use of nanostructured alumina as an insecticide. Pest Manag. Sci. 2010, 66, 577–579. [Google Scholar] [CrossRef]
- Rouhani, M.; Samih, M.A.; Kalantari, S. Insecticidal effect of silica and silver nanoparticles on the cowpea seed beetle, Callosobruchus maculatus F.(Col.: Bruchidae). J. Entomol. Res. 2012, 4, 297–305. [Google Scholar]
- Debnath, N.; Das, S.; Seth, D.; Chandra, R.; Bhattacharya, S.C.; Goswami, A. Entomotoxic effect of silica nanoparticles against Sitophilus oryzae (L.). J. Pest Sci. 2011, 84, 99–105. [Google Scholar] [CrossRef]
- Gutiérrez-Ramírez, J.A.; Betancourt-Galindo, R.; Aguirre-Uribe, L.A.; Cerna-Chávez, E.; Sandoval-Rangel, A.; Castro Del Ángel, E.; Chacón-Hernández, J.C.; García-López, J.I.; Hernández-Juárez, A. Insecticidal effect of zinc oxide and titanium dioxide nanoparticles against bactericera cockerelli sulc. (hemiptera: Triozidae) on tomato solanum lycopersicum. Agronomy 2021, 11, 1460. [Google Scholar] [CrossRef]
- Norman, D.J.; Chen, J. Effect of foliar application of titanium dioxide on bacterial blight of geranium and Xanthomonas leaf spot of Poinsettia. HortScience 2011, 46, 426–428. [Google Scholar] [CrossRef] [Green Version]
- Elsharkawy, M.; Derbalah, A. Antiviral activity of titanium dioxide nanostructures as a control strategy for broad bean strain virus in faba bean. Pest Manag. Sci. 2019, 75, 828–834. [Google Scholar] [CrossRef]
- Antonoglou, O.; Moustaka, J.; Adamakis, I.D.S.; Sperdouli, I.; Pantazaki, A.A.; Moustakas, M.; Dendrinou-Samara, C. Nanobrass CuZn Nanoparticles as Foliar Spray Nonphytotoxic Fungicides. ACS Appl. Mater. Interfaces 2018, 10, 4450–4461. [Google Scholar] [CrossRef]
- Tryfon, P.; Kamou, N.N.; Mourdikoudis, S.; Menkissoglu-Spiroudi, U.; Dendrinou-Samara, C. CuZn and ZnO Nanoflowers as Nano-Fungicides against Botrytis cinerea and Sclerotinia sclerotiorum: Phytoprotection, Translocation, and Impact after Foliar Application. Materials 2021, 14, 7600. [Google Scholar] [CrossRef]
- Gkanatsiou, C.; Ntalli, N.; Menkissoglu-Spiroudi, U.; Dendrinou-Samara, C. Essential Metal-Based Nanoparticles (Copper/Iron NPs) as Potent Nematicidal Agents against Meloidogyne spp. J. Nanotechnol. Res. 2019, 1, 44–58. [Google Scholar] [CrossRef] [Green Version]
- Nomoev, A.V.; Bardakhanov, S.P.; Schreiber, M.; Bazarova, D.G.; Romanov, N.A.; Baldanov, B.B.; Radnaev, B.R.; Syzrantsev, V.V. Structure and mechanism of the formation of core-shell nanoparticles obtained through a one-step gas-phase synthesis by electron beam evaporation. Beilstein J. Nanotechnol. 2015, 6, 874–880. [Google Scholar] [CrossRef]
- Ozcan, A.; Young, M.; Lee, B.; Liao, Y.Y.; Da Silva, S.; Godden, D.; Colee, J.; Huang, Z.; Mendis, H.C.; Campos, M.G.N.; et al. Copper-fixed quat: A hybrid nanoparticle for application as a locally systemic pesticide (LSP) to manage bacterial spot disease of tomato. Nanoscale Adv. 2021, 3, 1473–1483. [Google Scholar] [CrossRef]
- Young, M.; Ozcan, A.; Myers, M.E.; Johnson, E.G.; Graham, J.H.; Santra, S. Multimodal Generally Recognized as Safe ZnO/Nanocopper Composite: A Novel Antimicrobial Material for the Management of Citrus Phytopathogens. J. Agric. Food Chem. 2018, 66, 6604–6608. [Google Scholar] [CrossRef]
- Liu, B.; Xue, Y.; Zhang, J.; Han, B.; Zhang, J.; Suo, X.; Mu, L.; Shi, H. Visible-light-driven TiO2/Ag3PO4 heterostructures with enhanced antifungal activity against agricultural pathogenic fungi Fusarium graminearum and mechanism insight. Environ. Sci. Nano 2017, 4, 255–264. [Google Scholar] [CrossRef]
- Liu, B.; Mu, L.; Zhang, J.; Han, X.; Shi, H. TiO2/Cu2(OH)2CO3 nanocomposite as efficient antimicrobials for inactivation of crop pathogens in agriculture. Mater. Sci. Eng. C 2020, 107, 110344. [Google Scholar] [CrossRef]
- Boxi, S.S.; Mukherjee, K.; Paria, S. Ag doped hollow TiO2 nanoparticles as an effective green fungicide against Fusarium solani and Venturia inaequalis phytopathogens. Nanotechnology 2016, 27, 85103. [Google Scholar] [CrossRef] [PubMed]
- Paret, M.L.; Vallad, G.E.; Averett, D.R.; Jones, J.B.; Olson, S.M. Photocatalysis: Effect of light-activated nanoscale formulations of TiO2 on Xanthomonas perforans and control of bacterial spot of tomato. Phytopathology 2013, 103, 228–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tryfon, P.; Kamou, N.N.; Ntalli, N.; Mourdikoudis, S.; Karamanoli, K.; Karfaridis, D.; Menkissoglu-Spiroudi, U.; Dendrinou-Samara, C. Coated Cu-doped ZnO and Cu nanoparticles as control agents against plant pathogenic fungi and nematodes. NanoImpact 2022, 28, 100430. [Google Scholar] [CrossRef] [PubMed]
- Enayati, S.; Davari, M.; Habibi-Yangjeh, A.; Ebadollahi, A.; Feizpoor, S. Enhancement of the antifungal properties of Zataria 653 multiflora essential oil through nanoencapsulation with ZnO nanomaterial. Res. Sq. 2021. [Google Scholar] [CrossRef]
- Akhtari, A.; Davari, M.; Habibi-Yangjeh, A.; Ebadollahi, A.; Feizpour, S. Antifungal Activities of Pure and ZnO-Encapsulated Essential Oil of Zataria multiflora on Alternaria solani as the Pathogenic Agent of Tomato Early Blight Disease. Front. Plant Sci. 2022, 13, 932475. [Google Scholar] [CrossRef]
- Giannousi, K.; Menelaou, M.; Arvanitidis, J.; Angelakeris, M.; Pantazaki, A.; Dendrinou-Samara, C. Hetero-nanocomposites of magnetic and antifungal nanoparticles as a platform for magnetomechanical stress induction in Saccharomyces cerevisiae. J. Mater. Chem. B 2015, 3, 5341–5351. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nanoparticle Properties | Antimicrobial Properties | Ref | |||||
|---|---|---|---|---|---|---|---|
| Composition | Preparation | Size | Shape | Effective Parameter | Specie | Evaluation Method | |
| Ag | Chemical reduction | 40–60 nm | Spherical | Concentration | Rhizoctonia solani | In vitro | [51] |
| Ag | Chemical/Turkevich | 52 nm | Spherical | Concentration | Phomopsis spp. | In vitro & in planta | [52] |
| Ag-gelatin | Chemical reduction | 5–24 nm | Spherical | Concentration | Colletotrichum gloesporioides | In vitro | [53] |
| Ag-PVP | Modified Tollens’ method | - | - | Concentration | Sclerotinia sclerotiorum | In vitro | [54] |
| Ag | High-voltage arc discharge method | 20 nm | - | Exposure time | Fusarium culmorum | In vitro | [55] |
| Ag | Dual reduction | 30 nm | - | Concentration, exposure time | Fusarium spp. | In vitro | [56] |
| Ag | Electrolysis | - | - | Application time (before infection) | Bipolaris sorokiniana, Magnaporthe grisea | In vitro | [57] |
| Ag | Chemical reduction | 7.5 nm | - | Concentration, exposure time | Gibberella fujikuroi | In vitro & in planta | [58] |
| Ag-SDS | Direct-current, Atmospheric-Pressure, Glow Discharge (dc-APGD) | 28 nm | - | Concentration | Dickeya spp., Pectobacterium spp., Erwinia amylovora, Clavibacter michiganensis, Ralstonia solanacearum, Xanthomonas campestris | In vitro | [59,60] |
| Ag | Chemical reduction | 10–100 nm | Spherical | Exposure time | Ralstonia solanacearum | In planta | [61] |
| Ag-bovine submaxillary mucin | Chemical synthesis | 5–20 nm | - | Size, concentration | Acidovorax sp., Xanthomonas sp., Clavibacter sp. | In vitro, pot experiments | [62] |
| Ag | Chemical reduction | 12 nm | Spherical | Exposure time | Potato virus Y (PVY) | In planta | [63] |
| Ag | Commercial | - | - | Concentration, application time (before infection) | Potato virus Y (PVY), Tomato mosaic virus (ToMV) | In planta | [64] |
| Ag | Co-precipitation | 12.6 nm, 8 nm | - | Concentration, application time (after infection) | Tomato spotted wilt virus (TSWV), Bean yellow mosaic virus (BYMV) | In planta | [65,66] |
| Pegylated Cu2O, Cu/Cu2O | Hydrothermal | 11–55 nm | Spherical | Concentration, composition phase Cu2O | Phytophthora infestans | In vitro, in planta (field exper.) | [70] |
| Cu, CuO | Commercial | 25 nm, <50 nm | - | Concentration, sensitivity in target site | Botrytis cinerea, Alternaria alternata, Monilia fructicola, Colletotrichum gloeosporioides, Fusarium solani, Fusarium oxysporum, Verticillium dahliae | In vitro & in planta | [80] |
| Cu | Bifunctional molecule- assisted method | 50 nm | Spherical | Concentration | Alternaria solani | In planta | [81] |
| Cu | Chemical reduction | 53 nm | Spherical | Concentration, exposure time | Aspergillus niger, Fusarium oxysporum, Phytophthora capsici | In vitro | [82,83] |
| Cu | Chemical reduction | 345 nm | Polygonal | Shape | Fusarium oxysporum | In vitro | [84] |
| Cu-animal protein, non-ionic polymer, ionic polymer | Modified wet chemistry | 5–10 nm | Spherical | Size, concentration, application time (developmental stage) | Fusicladium oleagineum Colletotrichum spp. | In vitro, in planta | [85] |
| Cu-CTAB | Chemical reduction | 3–10 nm | Spherical | Size | Phoma destructiva, Curvularia lunata, Alternaria alternata Fusarium oxysporum | In vitro | [86] |
| Cu-CTAB | Chemical reduction | 20–50 nm | Spherical | Concentration | Fusarium equiseti, Fusarium oxysporum, Fusarium culmorum | In vitro | [87,88] |
| Cu2O@OAm, Cu/Cu2O@ OAm | Solvothermal | 30 nm, 170 nm | Spherical, nanorods | Concentration, composition phase Cu2O | Saccharomyces cerevisiae | In vitro, in planta | [89] |
| Cu/Cu2O@ PEG 8000 | Aqueous-phase synthesis | 42 nm | Spherical | Concentration, composition phase Cu2O | Fusarium oxysporum | In vitro | [90] |
| CuS | Pyrolytic technique | - | Spherical, granular | Shape | Fusarium spp. | In vitro | [91] |
| CuO | Modified wet chemistry | 5 nm | Spherical | Concentration, zeta-potential | Agrobacterium tumefaciens, Dickeya dadantii, Erwinia amylovora, Pectobacterium carotovorum, Pseudomonas corrugata, Pseudomonas savastanoi, Xanthomonas campestris | In vitro | [92] |
| Cu2O@ PEG 8000 | Hydrothermal | 16 nm | Spherical | Size | Xanthomonas campestris, Escherichia coli, Bacillus subtilis, Bacillus cereus, Staphylococcus aureus | In vitro | [93] |
| Cu@ Tween 20 | Hydrothermal | 46 nm | Spherical | Concentration, metallic core Cu | Erwinia amylovora, Xanthomonas campestris, Pseudomonas syringae | In vitro, in planta | [94] |
| Cu | Chemical reduction | 18–33 nm | - | Size, concentration | Xanthomonas oryzae | In planta | [96] |
| CuO | Direct precipitation | 20 nm | Flower-like | Exposure time, morphology | Spodoptera littoralis | In planta | [97] |
| ZnO | Sol-gel | 20–35 nm | Spherical | Concentration | Erythricium salmonicolor | In vitro | [103] |
| ZnO | Microwave synthesis | 30 nm | Spherical | - | Fusarium graminearum | In planta | [104] |
| ZnO | Commercial | - | - | Concentration | Fusarium graminearum | In vitro | [105] |
| ZnO | Colloidal, Hydrothermal synthesis | Diam. 246 nm, Thick. 48 nm | Platelet | Shape | Fusarium solani, Colletotrichum gloesporioids | In vitro | [106] |
| ZnO | Controlled precipitation | 20–70 nm | - | Concentration, exposure time | Colletotrichum sp. | In vitro | [107] |
| ZnO | Solvothermal | <100 nm | Spheroidal | Composition | Mycena citricolor | In vitro | [108] |
| ZnO | One-pot chemical precipitation | 65 nm | Irregular, porous structure | Shape | Alternaria alternata, Fusarium verticilliodes | In vitro | [109] |
| Pd or Ce-doped ZnO | Sol-gel, precipitation, microwave-assisted hydrothermal | 55–100 nm | Flower-like | Composition, shape, concentration | Candida albicans, Aspergillus niger, Aspergillus flavus | In vitro | [110,111] |
| ZnO | Commercial | <100 nm | - | Concentration, foliar spray | Pseudomonas syringae, Xanthomonas campestris, Pectobacterium carotovorum, Pectobacterium betavasculorum, Ralstonia solanacearum | In vitro, in planta | [112,113] |
| ZnO | Chemical/Bath Deposition | - | Nanorods | Concentration | Pseudomonas syringae | In planta, In vitro | [114] |
| ZnO | Sol-gel | 55 nm | Spherical | Exposure time | Tobacco mosaic Virus (TMV) | In planta | [115] |
| ZnO | Commercial | - | - | Concentration, foliar spray | Tomato Mosaic Virus (ToMV) | In planta | [116] |
| Fe2O3 | Wet chemistry (green approach) | 10–30 nm | Spherical | Concentration, species sensitivity | Trichothecium roseum, Cladosporium herbarum, Penicillium chrysogenum, Alternaria alternata, Aspergillus niger | In vitro | [118] |
| CoFe2O4, NiFe2O4 | Co-precipitation | 25 nm | Spherical | Concentration | Fusarium oxysporum | In planta | [119] |
| Al-based | Microemulsion | 100–250 nm | Spherical, mesoporous | Concentration | Fusarium oxysporum | In vitro, in planta | [120] |
| Si-based | One-pot direct template | 20–150 μm | Mesoporous | Concentration | Alternaria solani | In vitro, in planta | [121] |
| Al2O3 | Glycine-Nitrate combustion synthesis | 10 μm | Amorphous | Concentration, exposure time | Acromyrmex lobicornis, Sitophilus oryzae, Rhyzopertha dominica | In planta | [122,123] |
| SiO2 | Sol-gel | 20–60 nm | Spherical | Concentration | Callosobruchus maculates & Sitophilus oryzae | In planta | [124,125] |
| TiO2 | Controlled precipitation | 76 nm long, 8 nm wide | Needle | Concentration, foliar spray | Bactericera cockerelli | In vitro, in planta | [126] |
| TiO2 | Commercial | - | - | Concentration, foliar spray | Xanthomonas spp. | In planta | [127] |
| TiO2-oleic acid | Commercial | 3–5 μm | Hollow | Shape, foliar spray | Broad bean strain virus (BBSV) | In planta | [128] |
| CuZn-glycol | Microwave-assisted Polyol Process (MW-PP) | 20 nm | - | Concentration | Saccharomyces cerevisiae | In planta | [129] |
| CuZn-glycol | Solvothermal | 35 nm | Nanoflower | Concentration | Botrytis cinerea, Sclerotinia sclerotiorum | In planta | [130] |
| CuFe- pegyllated | Chemical reduction, Hydrothermal | 40 nm | - | Composition (Cu released ions) | Meloidogyne spp. | In vitro | [131] |
| Si-Cu-Quat | Sequential addition, Sol-gel | 50–600 nm (silica core), <10 nm (Cu NPs) | Core-shell | Composition | Xanthomonas perforans | In planta | [133] |
| ZnO-nCuSi | Sol-gel | 600–1100 nm | Core-shell | Composition | Xanthomonas alfalfae subsp. Citrumelonis, Pseudomonas syringae pv. syringae, Clavibacter michiganensis subsp. michiganensis | In vitro, in planta | [134] |
| TiO2/Ag3PO4, TiO2/Cu2(OH)2CO3 | Solvothermal, In situ precipitation | 2–5 nm of dopants | Microspheres, NPs | Composition, exposure time | Fusarium graminearum | In vitro | [135,136] |
| Ag-doped TiO2 | Sol-gel | - | Hollow sphere | Visible light exposure duration | Fusarium solani | In vitro | [137] |
| Zn-doped TiO2 | Commercial | 7 nm | - | Concentration, exposure time | Xanthomonas perforans | In vitro, in planta | [138] |
| Cu-doped ZnO | Microwave-assisted polyol process | 12 nm | Spheroidal | Concentration | Botrytis cinerea, Sclerotinia sclerotiorum, Meloidogyne javanica | In vitro, in planta | [139] |
| ZnO-ZmEO | Precipitation | - | Nanocapsule | Concentration, exposure time | Alternaria solani | In vitro | [141] |
| Cu2O/ NiFe2O4 | Solvothermal | 30 nm Cu2O, 9 nm NiFe2O4 | Spherical | Concentration, exposure time | Saccharomyces cerevisiae | In vitro | [142] |
| ZnO-ZmEO | Precipitation | - | Nanocapsule | Concentration, exposure time | Fusarium spp. | In vitro | [140] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kanakari, E.; Dendrinou-Samara, C. Fighting Phytopathogens with Engineered Inorganic-Based Nanoparticles. Materials 2023, 16, 2388. https://doi.org/10.3390/ma16062388
Kanakari E, Dendrinou-Samara C. Fighting Phytopathogens with Engineered Inorganic-Based Nanoparticles. Materials. 2023; 16(6):2388. https://doi.org/10.3390/ma16062388
Chicago/Turabian StyleKanakari, Eirini, and Catherine Dendrinou-Samara. 2023. "Fighting Phytopathogens with Engineered Inorganic-Based Nanoparticles" Materials 16, no. 6: 2388. https://doi.org/10.3390/ma16062388
APA StyleKanakari, E., & Dendrinou-Samara, C. (2023). Fighting Phytopathogens with Engineered Inorganic-Based Nanoparticles. Materials, 16(6), 2388. https://doi.org/10.3390/ma16062388