Long-Term Recordings of Multiple, Single-Neurons for Clinical Applications: The Emerging Role of the Bioactive Microelectrode

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

2. The Brain’s Initial Reaction to the Trauma of Insertion



2.1. The Cellular Response

2.2. Neuronal Cell Death and Migration
2.3. Evaluating the Success of Response Reduction

2.4. Effect of Initial Microelectrode Insertion on the Glia Response

3. Bioactive Interventions to Modulate Gliosis
3.1. Increasing Adhesion between the Microelectrode and the Neural Tissue
3.2. Polymer Coatings for Microelectrodes

3.3. Enhancing Microelectrodes with Bioactive Molecules

4. Altering the Physical Structure of the Microelectrodes
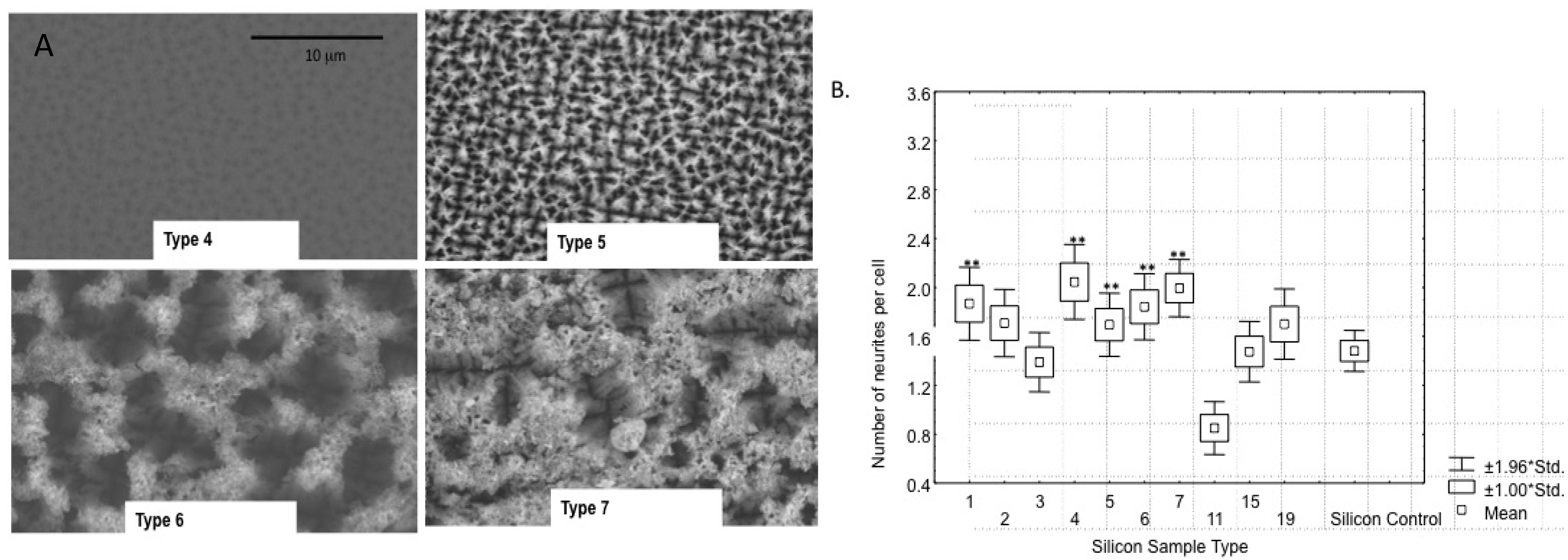
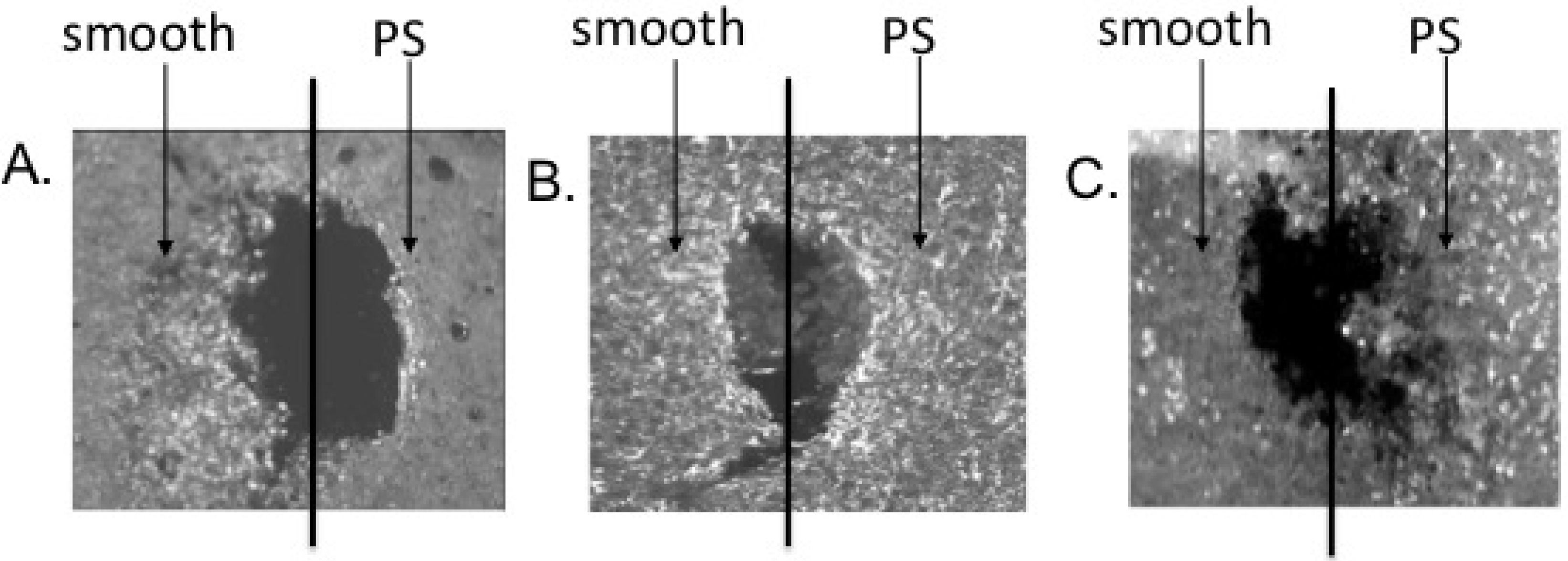
4.1. Modifying Surface Structure
4.2. Flexible microelectrodes: Matching the Brain’s Mechanical Properties

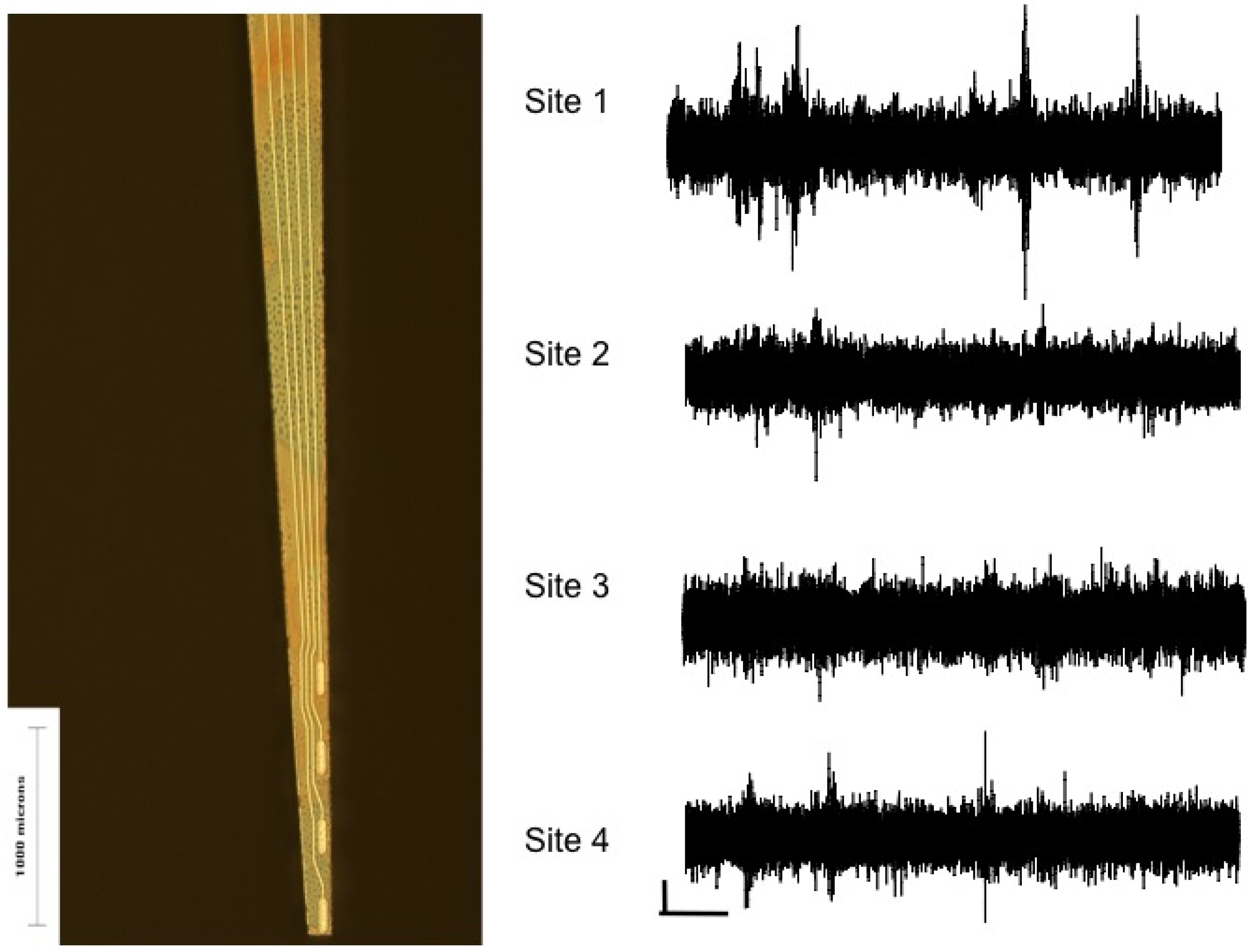
4.3 Floating Arrays vs. Untethering the Microelectrode


5. Conclusions
Acknowledgements
References and Notes
- Chapin, J.K.; Moxon, K.A.; Markowitz, R.S.; Nicolelis, M.A. Real-time control of a robot arm using simultaneously recorded neurons in the motor cortex. Nat. Neurosci. 1999, 2, 664–670. [Google Scholar] [CrossRef] [PubMed]
- Hochberg, L.R.; Serruya, M.D.; Friehs, G.M.; Mukand, J.A.; Saleh, M.; Caplan, A.H.; Branner, A.; Chen, D.; Penn, R.D.; Donoghue, J.P. Neuronal ensemble control of prosthetic devices by a human with tetraplegia. Nature 2006, 442, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Winter, J.O.; Cogan, S.F.; Rizzo, J.F., 3rd. Neurotrophin-eluting hydrogel coatings for neural stimulating electrodes. J. Biomed. Mater. Res. B Appl. Biomater. 2007, 81, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Bell, T.E.; Wise, K.D.; Anderson, D.J. A flexible micromachined electrode array for a cochlear prosthesis. Sens. Actuators A 1998, 66, 63–69. [Google Scholar] [CrossRef]
- Moxon, K.A.; Morizio, J.; Chapin, J.K.; Nicolelis, M.A.L.; Wolf, P.D. Designing a brain-machine interface for neuroprosthetic control. In Neural Prostheses for Restoration of Sensory and Motor Function; Chapin, J.K., Moxon, K.A., Eds.; CRC Press: Boca Raton, FL, USA, 2000; pp. 179–220. [Google Scholar]
- Polikov, V.S.; Tresco, P.A.; Reichert, W.M. Response of brain tissue to chronically implanted neural electrodes. J. Neurosci. Methods 2005, 148, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Grill, W.M.; Norman, S.E.; Bellamkonda, R.V. Implanted neural interfaces: biochallenges and engineered solutions. Annu. Rev. Biomed. Eng. 2009, 11, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Moxon, K.A. Multichannel electrode design: Considerations for different applications. In Methods for Neural Ensemble Recordings; Nicolelis, M.A.L., Ed.; CRC Press: Boca Raton, FL, USA, 1999; pp. 25–45. [Google Scholar]
- Kennedy, P.R.; Bakay, R.A. Restoration of neural output from a paralyzed patient by a direct brain connection. Neuroreport 1998, 9, 1707–1711. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, P.R.; Bakay, R.A.; Moore, M.M.; Adams, K.; Goldwaithe, J. Direct control of a computer from the human central nervous system. IEEE Trans. Rehabil. Eng. 2000, 8, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Moxon, K.A.; Kuzmick, V.; Lafferty, J.; Serfass, A.; Szperka, D.; Zale, B.; Johnson, J.; Nagvajara, P. Real-time seizure detection system using multiple single-neuron recordings. In Proceedings of the 23rd Annual International Conference of the IEEE Engineering in Medicine and Biology Society, Istanbul, Turkey, October 25-28, 2001.
- Suner, S.; Fellows, M.R.; Vargas-Irwin, C.; Nakata, G.K.; Donoghue, J.P. Reliability of signals from a chronically implanted, silicon-based electrode array in non-human primate primary motor cortex. IEEE Trans. Neural. Syst. Rehabil. Eng. 2005, 13, 524–541. [Google Scholar] [CrossRef] [PubMed]
- Lebedev, M.A.; Nicolelis, M.A. Brain-machine interfaces: Past, present and future. Trends Neurosci. 2006, 29, 536–546. [Google Scholar] [CrossRef] [PubMed]
- Kipke, D.R.; Vetter, R.J.; Williams, J.C.; Hetke, J.F. Silicon-substrate intracortical microelectrode arrays for long-term recording of neuronal spike activity in cerebral cortex. IEEE Trans. Neural Syst. Rehabil. Eng. 2003, 11, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Szarowski, D.H.; Andersen, M.D.; Retterer, S.; Spence, A.J.; Isaacson, M.; Craighead, H.G.; Turner, J.N.; Shain, W. Brain responses to micro-machined silicon devices. Brain Res. 2003, 983, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Vetter, R.J.; Williams, J.C.; Hetke, J.F.; Nunamaker, E.A.; Kipke, D.R. Chronic neural recording using silicon-substrate microelectrode arrays implanted in cerebral cortex. IEEE Trans. Biomed. Eng. 2004, 51, 896–904. [Google Scholar] [CrossRef] [PubMed]
- Moxon, K.A.; Kalkhoran, N.M.; Markert, M.; Sambito, M.A.; McKenzie, J.L.; Webster, J.T. Nanostructured surface modification of ceramic-based microelectrodes to enhance biocompatibility for a direct brain-machine interface. IEEE Trans. Biomed. Eng. 2004, 51, 881–889. [Google Scholar] [CrossRef] [PubMed]
- Hsu, J.M.; Rieth, L.; Normann, R.A.; Tathireddy, P.; Solzbacher, F. Encapsulation of an integrated neural interface device with Parylene C. IEEE Trans. Biomed. Eng. 2009, 56, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Seymour, J.P.; Elkasabi, Y.M.; Chen, H.Y.; Lahann, J.; Kipke, D.R. The insulation performance of reactive parylene films in implantable electronic devices. Biomaterials 2009, 30, 6158–6167. [Google Scholar] [CrossRef] [PubMed]
- Moxon, K.A.; Leiser, S.C.; Gerhardt, G.A.; Barbee, K.A.; Chapin, J.K. Ceramic-based multisite electrode arrays for chronic single-neuron recording. IEEE Trans. Biomed. Eng. 2004, 51, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Biran, R.; Martin, D.C.; Tresco, P.A. Neuronal cell loss accompanies the brain tissue response to chronically implanted silicon microelectrode arrays. Exp. Neurol. 2005, 195, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Schultz, R.L.; Willey, T.J. The ultrastructure of the sheath around chronically implanted electrodes in brain. J. Neurocytol. 1976, 5, 621–642. [Google Scholar] [CrossRef] [PubMed]
- Edell, D.J.; Toi, V.V.; McNeil, V.M.; Clark, L.D. Factors influencing the biocompatibility of insertable silicon microshafts in cerebral cortex. IEEE Trans. Biomed. Eng. 1992, 39, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Fitch, M.T.; Silver, J. Activated macrophages and the blood-brain barrier: inflammation after CNS injury leads to increases in putative inhibitory molecules. Exp. Neurol. 1997, 148, 587–603. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.M.; Rodriguez, A.; Chang, D.T. Foreign body reaction to biomaterials. Semin. Immunol. 2008, 20, 86–100. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; McCreery, D.B.; Carter, R.R.; Bullara, L.A.; Yuen, T.G.; Agnew, W.F. Stability of the interface between neural tissue and chronically implanted intracortical microelectrodes. IEEE Trans. Rehabil. Eng. 1999, 7, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Giulian, D.; Chen, J.; Ingeman, J.E.; George, J.K.; Noponen, M. The role of mononuclear phagocytes in wound-healing after traumatic injury to adult mammalian brain. J. Neurosci. 1989, 9, 4416–4429. [Google Scholar] [PubMed]
- Kahn, M.A.; Ellison, J.A.; Speight, G.J.; de Vellis, J. CNTF regulation of astrogliosis and the activation of microglia in the developing rat central nervous system. Brain Res. 1995, 685, 55–67. [Google Scholar] [CrossRef] [PubMed]
- Rostworowski, M.; Balasingam, V.; Chabot, S.; Owens, T.; Yong, V.W. Astrogliosis in the neonatal and adult murine brain post-trauma: elevation of inflammatory cytokines and the lack of requirement for endogenous interferon-gamma. J. Neurosci. 1997, 17, 3664–3674. [Google Scholar] [PubMed]
- Fawcett, J.W.; Asher, R.A. The glial scar and central nervous system repair. Brain Res. Bull. 1999, 49, 377–391. [Google Scholar] [CrossRef] [PubMed]
- Raivich, G.; Haas, S.; Werner, A.; Klein, M.A.; Kloss, C.; Kreutzberg, G.W. Regulation of MCSF receptors on microglia in the normal and injured mouse central nervous system: A quantitative immunofluorescence study using confocal laser microscopy. J. Comp. Neurol. 1998, 395, 342–358. [Google Scholar] [CrossRef] [PubMed]
- Elkabes, S.; DiCicco-Bloom, E.M.; Black, I.B. Brain microglia/macrophages express neurotrophins that selectively regulate microglial proliferation and function. J. Neurosci. 1996, 16, 2508–2521. [Google Scholar] [PubMed]
- Goss, J.R.; O'Malley, M.E.; Zou, L.; Styren, S.D.; Kochanek, P.M.; DeKosky, S.T. Astrocytes are the major source of nerve growth factor upregulation following traumatic brain injury in the rat. Exp. Neurol. 1998, 149, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Raivich, G.; Bohatschek, M.; Kloss, C.U.; Werner, A.; Jones, L.L.; Kreutzberg, G.W. Neuroglial activation repertoire in the injured brain: graded response, molecular mechanisms and cues to physiological function. Brain Res. Brain Res. Rev. 1999, 30, 77–105. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, K.; Honda, S.; Tohyama, Y.; Imai, Y.; Kohsaka, S.; Kurihara, T. Neurotrophin secretion from cultured microglia. J. Neurosci. Res. 2001, 65, 322–331. [Google Scholar] [CrossRef] [PubMed]
- Woodroofe, M.N.; Sarna, G.S.; Wadhwa, M.; Hayes, G.M.; Loughlin, A.J.; Tinker, A.; Cuzner, M.L. Detection of interleukin-1 and interleukin-6 in adult rat brain, following mechanical injury, by in vivo microdialysis: Evidence of a role for microglia in cytokine production. J. Neuroimmunol. 1991, 33, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Sheng, W.S.; Hu, S.; Kravitz, F.H.; Peterson, P.K.; Chao, C.C. Tumor necrosis factor alpha upregulates human microglial cell production of interleukin-10 in vitro. Clin. Diagn. Lab Immunol. 1995, 2, 604–608. [Google Scholar] [PubMed]
- Kossmann, T.; Hans, V.H.; Imhof, H.G.; Stocker, R.; Grob, P.; Trentz, O.; Morganti-Kossmann, C. Intrathecal and serum interleukin-6 and the acute-phase response in patients with severe traumatic brain injuries. Shock 1995, 4, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Babcock, A.A.; Kuziel, W.A.; Rivest, S.; Owens, T. Chemokine expression by glial cells directs leukocytes to sites of axonal injury in the CNS. J. Neurosci. 2003, 23, 7922–7930. [Google Scholar] [PubMed]
- Minghetti, L.; Levi, G. Microglia as effector cells in brain damage and repair: Focus on prostanoids and nitric oxide. Prog. Neurobiol. 1998, 54, 99–125. [Google Scholar] [CrossRef] [PubMed]
- Koyama, Y.; Takemura, M.; Fujiki, K.; Ishikawa, N.; Shigenaga, Y.; Baba, A. BQ788, an endothelin ET(B) receptor antagonist, attenuates stab wound injury-induced reactive astrocytes in rat brain. Glia 1999, 26, 268–271. [Google Scholar] [CrossRef] [PubMed]
- Fitch, M.T.; Doller, C.; Combs, C.K.; Landreth, G.E.; Silver, J. Cellular and molecular mechanisms of glial scarring and progressive cavitation: In vivo and in vitro analysis of inflammation-induced secondary injury after CNS trauma. J. Neurosci. 1999, 19, 8182–8198. [Google Scholar] [PubMed]
- Giulian, D.; Li, J.; Leara, B.; Keenen, C. Phagocytic microglia release cytokines and cytotoxins that regulate the survival of astrocytes and neurons in culture. Neurochem. Int. 1994, 25, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Svensson, M.; Eriksson, N.P.; Aldskogius, H. Evidence for activation of astrocytes via reactive microglial cells following hypoglossal nerve transection. J. Neurosci. Res. 1993, 35, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Balasingam, V.; Tejada-Berges, T.; Wright, E.; Bouckova, R.; Yong, V.W. Reactive astrogliosis in the neonatal mouse brain and its modulation by cytokines. J. Neurosci. 1994, 14, 846–856. [Google Scholar] [PubMed]
- Eddleston, M.; Mucke, L. Molecular profile of reactive astrocytes--implications for their role in neurologic disease. Neuroscience 1993, 54, 15–36. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.Y.; Lieska, N.; Kriho, V.; Wu, C.M.; Pappas, G.D. A subpopulation of reactive astrocytes at the immediate site of cerebral cortical injury. Exp. Neurol. 1997, 146, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Raghupathi, R.; Graham, D.I.; McIntosh, T.K. Apoptosis after traumatic brain injury. J. Neurotrauma 2000, 17, 927–938. [Google Scholar] [CrossRef] [PubMed]
- McConnell, G.C.; Rees, H.D.; Levey, A.I.; Gutekunst, C.A.; Gross, R.E.; Bellamkonda, R.V. Implanted neural electrodes cause chronic, local inflammation that is correlated with local neurodegeneration. J. Neural. Eng. 2009, 6, 56003. [Google Scholar] [CrossRef]
- Turner, J.N.; Shain, W.; Szarowski, D.H.; Andersen, M.; Martins, S.; Isaacson, M.; Craighead, H. Cerebral astrocyte response to micromachined silicon implants. Exp. Neurol. 1999, 156, 33–49. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, C.E.; Leach, J.B. Neural tissue engineering: strategies for repair and regeneration. Annu. Rev. Biomed. Eng. 2003, 5, 293–347. [Google Scholar] [CrossRef] [PubMed]
- Leung, B.K.; Biran, R.; Underwood, C.J.; Tresco, P.A. Characterization of microglial attachment and cytokine release on biomaterials of differing surface chemistry. Biomaterials 2008, 29, 3289–3297. [Google Scholar] [CrossRef] [PubMed]
- Porada, I.; Bondar, I.; Spatz, W.B.; Kruger, J. Rabbit and monkey visual cortex: More than a year of recording with up to 64 microelectrodes. J. Neurosci. Methods 2000, 95, 13–28. [Google Scholar] [CrossRef] [PubMed]
- Merrill, D.R.; Tresco, P.A. Impedance characterization of microarray recording electrodes in vitro. Conf. Proc. IEEE Eng. Med. Biol. Soc. 2004, 6, 4349–4352. [Google Scholar] [PubMed]
- Williams, J.C.; Hippensteel, J.A.; Dilgen, J.; Shain, W.; Kipke, D.R. Complex impedance spectroscopy for monitoring tissue responses to inserted neural implants. J. Neural Eng. 2007, 4, 410–423. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.D.; Kao, O.E.; Kipke, D.R. Spatiotemporal pH dynamics following insertion of neural microelectrode arrays. J. Neurosci. Methods 2007, 160, 276–287. [Google Scholar] [CrossRef] [PubMed]
- He, W.; McConnell, G.C.; Bellamkonda, R.V. Nanoscale laminin coating modulates cortical scarring response around implanted silicon microelectrode arrays. J. Neural Eng. 2006, 3, 316–326. [Google Scholar] [CrossRef] [PubMed]
- He, W.; McConnell, G.C.; Schneider, T.M.; Bellamkonda, R.V. A novel anti-inflammatory surface for neural electrodes. Adv. Mater. 2007, 19, 3529–3533. [Google Scholar] [CrossRef]
- Bjornsson, C.S.; Oh, S.J.; Al-Kofahi, Y.A.; Lim, Y.J.; Smith, K.L.; Turner, J.N.; De, S.; Roysam, B.; Shain, W.; Kim, S.J. Effects of insertion conditions on tissue strain and vascular damage during neuroprosthetic device insertion. J. Neural Eng. 2006, 3, 196–207. [Google Scholar] [CrossRef] [PubMed]
- Kralik, J.D.; Dimitrov, D.F.; Krupa, D.J.; Katz, D.B.; Cohen, D.; Nicolelis, M.A. Techniques for long-term multisite neuronal ensemble recordings in behaving animals. Methods 2001, 25, 121–150. [Google Scholar] [CrossRef] [PubMed]
- Paralikar, K.J.; Clement, R.S. Collagenase-aided intracortical microelectrode array insertion: Effects on insertion force and recording performance. IEEE Trans. Biomed. Eng. 2008, 55, 2258–2267. [Google Scholar] [CrossRef] [PubMed]
- Rousche, P.J.; Normann, R.A. A method for pneumatically inserting an array of penetrating electrodes into cortical tissue. Ann. Biomed. Eng. 1992, 20, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Stice, P.; Gilletti, A.; Panitch, A.; Muthuswamy, J. Thin microelectrodes reduce GFAP expression in the implant site in rodent somatosensory cortex. J. Neural Eng. 2007, 4, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Rosenthal, F. Extracellular potential fields of single PT-neurons. Brain Res. 1972, 36, 251–263. [Google Scholar] [CrossRef] [PubMed]
- Buzsaki, G.; Kandel, A. Somadendritic backpropagation of action potentials in cortical pyramidal cells of the awake rat. J. Neurophysiol. 1998, 79, 1587–1591. [Google Scholar] [PubMed]
- Mountcastle, V.B.; Davies, P.W.; Berman, A.L. Response properties of neurons of cat's somatic sensory cortex to peripheral stimuli. J. Neurophysiol. 1957, 20, 374–407. [Google Scholar] [PubMed]
- Zhong, Y.; Yu, X.; Gilbert, R.; Bellamkonda, R.V. Stabilizing electrode-host interfaces: A tissue engineering approach. J. Rehabil. Res. Dev. 2001, 38, 627–632. [Google Scholar] [PubMed]
- Subbaroyan, J.; Martin, D.C.; Kipke, D.R. A finite-element model of the mechanical effects of implantable microelectrodes in the cerebral cortex. J. Neural Eng. 2005, 2, 103–113. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Bellamkonda, R.V.; Sun, W.; Levenston, M.E. Biomechanical analysis of silicon microelectrode-induced strain in the brain. J. Neural Eng. 2005, 2, 81–89. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Bellamkonda, R.V. Nanoscale neuro-integrative coatings for neural implants. Biomaterials 2005, 26, 2983–2990. [Google Scholar] [CrossRef] [PubMed]
- McConnell, G.C.; Schneider, T.M.; Owens, D.J.; Bellamkonda, R.V. Extraction force and cortical tissue reaction of silicon microelectrode arrays implanted in the rat brain. IEEE Trans. Biomed. Eng. 2007, 54, 1097–1107. [Google Scholar] [CrossRef] [PubMed]
- Straley, K.S.; Heilshorn, S.C. Design and adsorption of modular engineered proteins to prepare customized, neuron-compatible coatings. Front Neuroengineering 2009, 2, 9. [Google Scholar]
- Chen, J.; Wise, K.D.; Hetke, J.F.; Bledsoe, S.C., Jr. A multichannel neural probe for selective chemical delivery at the cellular level. IEEE Trans. Biomed. Eng. 1997, 44, 760–769. [Google Scholar] [CrossRef] [PubMed]
- Rathnasingham, R.; Kipke, D.R.; Bledsoe, S.C., Jr.; McLaren, J.D. Characterization of implantable microfabricated fluid delivery devices. IEEE Trans. Biomed. Eng. 2004, 51, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Retterer, S.T.; Smith, K.L.; Bjornsson, C.S.; Neeves, K.B.; Spence, A.J.; Turner, J.N.; Shain, W.; Isaacson, M.S. Model neural prostheses with integrated microfluidics: A potential intervention strategy for controlling reactive cell and tissue responses. IEEE Trans. Biomed. Eng. 2004, 51, 2063–2073. [Google Scholar] [CrossRef] [PubMed]
- Retterer, S.T.; Smith, K.L.; Bjornsson, C.S.; Turner, J.N.; Isaacson, M.S.; Shain, W. Constant pressure fluid infusion into rat neocortex from implantable microfluidic devices. J. Neural. Eng. 2008, 5, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Willerth, S.M.; Sakiyama-Elbert, S.E. Approaches to neural tissue engineering using scaffolds for drug delivery. Adv. Drug Deliv. Rev. 2007, 59, 325–338. [Google Scholar] [CrossRef] [PubMed]
- Dipaolo, B.C. Biodegradable Polymer Scaffolding for Improved Neural Electrode Biocompatibility; Drexel University: Philadelphia, PA, USA, 2003. [Google Scholar]
- Azemi, E.; Stauffer, W.R.; Gostock, M.S.; Lagenaur, C.F.; Cui, X.T. Surface immobilization of neural adhesion molecule L1 for improving the biocompatibility of chronic neural probes: In vitro characterization. Acta Biomater. 2008, 4, 1208–1217. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Lee, V.A.; Raphael, Y.; Wiler, J.A.; Hetke, J.F.; Anderson, D.J.; Martin, D.C. Surface modification of neural recording electrodes with conducting polymer/biomolecule blends. J. Biomed. Mater. Res. 2001, 56, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Green, R.A.; Lovell, N.H.; Wallace, G.G.; Poole-Warren, L.A. Conducting polymers for neural interfaces: challenges in developing an effective long-term implant. Biomaterials 2008, 29, 3393–3399. [Google Scholar] [CrossRef] [PubMed]
- Diaz, A.F.; Castillo, J.I.; Logan, J.A.; Lee, W.Y. Electrochemistry of conducting polypyrrole films. J. Electroanal. Chem. 1981, 129, 115–132. [Google Scholar] [CrossRef]
- Wong, J.Y.; Langer, R.; Ingber, D.E. Electrically conducting polymers can noninvasively control the shape and growth of mammalian cells. Proc. Natl. Acad. Sci. USA 1994, 91, 3201–3204. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Kim, S.J. Method for unsupervised classification of multiunit neural signal recording under low signal-to-noise ratio. IEEE Trans. Biomed. Eng. 2003, 50, 421–431. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Martin, D.C.; Cui, X.; Shenai, M. Surface modification of neural probes with conducting polymer poly(hydroxymethylated-3,4-ethylenedioxythiophene) and its biocompatibility. Appl. Biochem. Biotechnol. 2006, 128, 117–130. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, K.A.; Uram, J.D.; Yang, J.; Martin, D.C.; Kipke, D.R. Chronic neural recordings using silicon microelectrode arrays electrochemically deposited with a poly(3,4-ethylenedi-oxythiophene) (PEDOT) film. J. Neural. Eng. 2006, 3, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Cui, Q.; Pollett, M.A.; Symons, N.A.; Plant, G.W.; Harvey, A.R. A new approach to CNS repair using chimeric peripheral nerve grafts. J. Neurotrauma 2003, 20, 17–31. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Wiler, J.A.; Anderson, D.J.; Kipke, D.R.; Martin, D.C. Conducting polymers on hydrogel-coated neural electrode provide sensitive neural recordings in auditory cortex. Acta Biomater. 2009. [Google Scholar] [CrossRef]
- Green, R.A.; Williams, C.M.; Lovell, N.H.; Poole-Warren, L.A. Novel neural interface for implant electrodes: improving electroactivity of polypyrrole through MWNT incorporation. J. Mater. Sci. Mater. Med. 2008, 19, 1625–1629. [Google Scholar] [CrossRef] [PubMed]
- Tomobe, Y.I.; Hama, H.; Sakurai, T.; Fujimori, A.; Abe, Y.; Goto, K. Anticoagulant factor protein S inhibits the proliferation of rat astrocytes after injury. Neurosci. Lett. 1996, 214, 57–60. [Google Scholar] [CrossRef] [PubMed]
- Lindholm, D.; Castren, E.; Kiefer, R.; Zafra, F.; Thoenen, H. Transforming growth factor-beta 1 in the rat brain: Increase after injury and inhibition of astrocyte proliferation. J. Cell Biol. 1992, 117, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Klaver, C.L.; Caplan, M.R. Bioactive surface for neural electrodes: Decreasing astrocyte proliferation via transforming growth factor-beta1. J. Biomed. Mater. Res. A 2007, 81, 1011–1016. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, P.R. The cone electrode—A long-term electrode that records from neurites grown onto its recording surface. J. Neurosci. Methods 1989, 29, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, P.R.; Bakay, R.A.; Sharpe, S.M. Behavioral correlates of action potentials recorded chronically inside the cone electrode. Neuroreport 1992, 3, 605–608. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, P.R.; Bakay, R.A. Activity of single action potentials in monkey motor cortex during long-term task learning. Brain Res. 1997, 760, 251–254. [Google Scholar] [CrossRef] [PubMed]
- Jun, S.B.; Hynd, M.R.; Dowell-Mesfin, N.M.; Al-Kofahi, Y.; Roysam, B.; Shain, W.; Kim, S.J. Modulation of cultured neural networks using neurotrophin release from hydrogel-coated microelectrode arrays. J. Neural. Eng. 2008, 5, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Shain, W.; Spataro, L.; Dilgen, J.; Haverstick, K.; Retterer, S.; Isaacson, M.; Saltzman, M.; Turner, J.N. Controlling cellular reactive responses around neural prosthetic devices using peripheral and local intervention strategies. IEEE Trans. Neural Syst. Rehabil. Eng. 2003, 11, 186–188. [Google Scholar] [CrossRef] [PubMed]
- Spataro, L.; Dilgen, J.; Retterer, S.; Spence, A.J.; Isaacson, M.; Turner, J.N.; Shain, W. Dexamethasone treatment reduces astroglia responses to inserted neuroprosthetic devices in rat neocortex. Exp. Neurol. 2005, 194, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Bellamkonda, R.V. Dexamethasone-coated neural probes elicit attenuated inflammatory response and neuronal loss compared to uncoated neural probes. Brain Res. 2007, 1148, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Martin, D.C. Sustained release of dexamethasone from hydrophilic matrices using PLGA nanoparticles for neural drug delivery. Biomaterials 2006, 27, 3031–3037. [Google Scholar] [PubMed]
- Zhong, Y.; Bellamkonda, R.V. Controlled release of anti-inflammatory agent alpha-MSH from neural implants. J. Control Release 2005, 106, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Serbest, G.; Horwitz, J.; Jost, M.; Barbee, K. Mechanisms of cell death and neuroprotection by poloxamer 188 after mechanical trauma. FASEB J. 2006, 20, 308–310. [Google Scholar] [PubMed]
- Serbest, G.; Horwitz, J.; Barbee, K. The effect of poloxamer-188 on neuronal cell recovery from mechanical injury. J. Neurotrauma 2005, 22, 119–132. [Google Scholar] [CrossRef] [PubMed]
- Raffa, V.; Pensabene, V.; Menciassi, A.; Dario, P. Design criteria of neuron/electrode interface. The focused ion beam technology as an analytical method to investigate the effect of electrode surface morphology on neurocompatibility. Biomed. Microdevices 2007, 9, 371–383. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, J.L.; Waid, M.C.; Shi, R.; Webster, T.J. Decreased functions of astrocytes on carbon nanofiber materials. Biomaterials 2004, 25, 1309–1317. [Google Scholar] [CrossRef] [PubMed]
- Canham, L.T. Bioactive silicon structure fabrication through nanoetching techniques. Adv. Mater. 1995, 7, 1033–1037. [Google Scholar] [CrossRef]
- Moxon, K.A.; Hallman, S.; Aslani, A.; Kalkhoran, N.M.; Lelkes, P.I. Bioactive properties of nanostructured porous silicon for enhancing electrode to neuron interfaces. J. Biomater. Sci. Polym. Ed. 2007, 18, 1263–1281. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Singh, A.; He, J.P.; Massia, S.; Kim, B.; Raupp, G. Polyimide based neural implants with stiffness improvement. Sens. Actuators B 2004, 102, 67–72. [Google Scholar] [CrossRef]
- Takeuchi, S.; Ziegler, D.; Yoshida, Y.; Mabuchi, K.; Suzuki, T. Parylene flexible neural probes integrated with microfluidic channels. Lab Chip 2005, 5, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Rousche, P.J.; Pellinen, D.S.; Pivin, D.P., Jr.; Williams, J.C.; Vetter, R.J.; Kipke, D.R. Flexible polyimide-based intracortical electrode arrays with bioactive capability. IEEE Trans. Biomed. Eng. 2001, 48, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Spence, A.J.; Neeves, K.B.; Murphy, D.; Sponberg, S.; Land, B.R.; Hoy, R.R.; Isaacson, M.S. Flexible multielectrodes can resolve multiple muscles in an insect appendage. J. Neurosci. Methods 2007, 159, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Ommaya, A.K. Trauma to the nervous system. Ann. R Coll. Surg. Engl. 1966, 39, 317–347. [Google Scholar] [PubMed]
- Kozai, T.D.; Kipke, D.R. Insertion shuttle with carboxyl terminated self-assembled monolayer coatings for implanting flexible polymer neural probes in the brain. J. Neurosci. Methods 2009. [Google Scholar] [CrossRef]
- Hetke, J.F.; Lund, J.L.; Najafi, K.; Wise, K.D.; Anderson, D.J. Silicon ribbon cables for chronically implantable microelectrode arrays. IEEE Trans. Biomed. Eng. 1994, 41, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Biran, R.; Martin, D.C.; Tresco, P.A. The brain tissue response to implanted silicon microelectrode arrays is increased when the device is tethered to the skull. J. Biomed. Mater. Res. A 2007, 82, 169–178. [Google Scholar] [CrossRef] [PubMed]
- Loeb, G.E.; Peck, R.A.; Moore, W.H.; Hood, K. BION system for distributed neural prosthetic interfaces. Med. Eng. Phys. 2001, 23, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Cameron, T.; Liinamaa, T.L.; Loeb, G.E.; Richmond, F.J. Long-term biocompatibility of a miniature stimulator implanted in feline hind limb muscles. IEEE Trans. Biomed. Eng. 1998, 45, 1024–1035. [Google Scholar] [CrossRef] [PubMed]
- Wu, A.C.; Spuhler, P.S.; Unlu, M.S.; Sahin, M. Floating Microfabricated Wireless Neuro- Stimulators. In Annual Fall Meeting of Biomedical Engineering Society, Los Angeles, CA, USA, September 26-29, 2007.
- Obeid, I.; Morizio, J.C.; Moxon, K.A.; Nicolelis, M.A.; Wolf, P.D. Two multichannel integrated circuits for neural recording and signal processing. IEEE Trans. Biomed. Eng. 2003, 50, 255–258. [Google Scholar] [CrossRef] [PubMed]
- Canham, L.T. Silicon quantum wire array fabrication by electrochemical and chemical dissolution of wafers. Appl. Phys. Lett. 1990, 57, 1046–1048. [Google Scholar] [CrossRef]
- Lehmann, V.; Gosele, U. Porous silicon formation—A quantum wire effect. Appl. Phys. Lett. 1991, 58, 856–858. [Google Scholar] [CrossRef]
- Lee, J.S.; Cho, N.H. Nanostructural and photoluminescence features of nanoporous silicon prepared by anodic etching. Appl. Surf. Sci. 2002, 190, 171–175. [Google Scholar] [CrossRef]
- Huang, M.J.; Yang, C.R.; Chiou, Y.C.; Lee, R.T. Fabrication of nanoporous antireflection surfaces on silicon. Sol. Energ. Mat. Sol. C. 2008, 92, 1352–1357. [Google Scholar] [CrossRef]
- Martinez-Duart, J.M.; Martin-Palma, R.J. Photodetectors and solar cells based on porous silicon. Phys. Status Solidi B 2002, 232, 81–88. [Google Scholar] [CrossRef]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Moxon, K.A.; Hallman, S.; Sundarakrishnan, A.; Wheatley, M.; Nissanov, J.; Barbee, K.A. Long-Term Recordings of Multiple, Single-Neurons for Clinical Applications: The Emerging Role of the Bioactive Microelectrode. Materials 2009, 2, 1762-1794. https://doi.org/10.3390/ma2041762
Moxon KA, Hallman S, Sundarakrishnan A, Wheatley M, Nissanov J, Barbee KA. Long-Term Recordings of Multiple, Single-Neurons for Clinical Applications: The Emerging Role of the Bioactive Microelectrode. Materials. 2009; 2(4):1762-1794. https://doi.org/10.3390/ma2041762
Chicago/Turabian StyleMoxon, Karen A., Stefanie Hallman, Aswin Sundarakrishnan, Margaret Wheatley, Jonathan Nissanov, and Kenneth A. Barbee. 2009. "Long-Term Recordings of Multiple, Single-Neurons for Clinical Applications: The Emerging Role of the Bioactive Microelectrode" Materials 2, no. 4: 1762-1794. https://doi.org/10.3390/ma2041762
APA StyleMoxon, K. A., Hallman, S., Sundarakrishnan, A., Wheatley, M., Nissanov, J., & Barbee, K. A. (2009). Long-Term Recordings of Multiple, Single-Neurons for Clinical Applications: The Emerging Role of the Bioactive Microelectrode. Materials, 2(4), 1762-1794. https://doi.org/10.3390/ma2041762