Initial Rhodonia placenta Gene Expression in Acetylated Wood: Group-Wise Upregulation of Non-Enzymatic Oxidative Wood Degradation Genes Depending on the Treatment Level

Abstract
:1. Introduction
2. Material and Methods
2.1. Wood Samples
2.2. Decay Test
2.3. RNA Purification and cDNA Synthesis
2.4. Quantitative Real-Time Polymerase Chain Reaction
2.5. Statistical Evaluation
3. Results
3.1. Mass Loss and Growth Rates
3.2. Expression of Target Genes
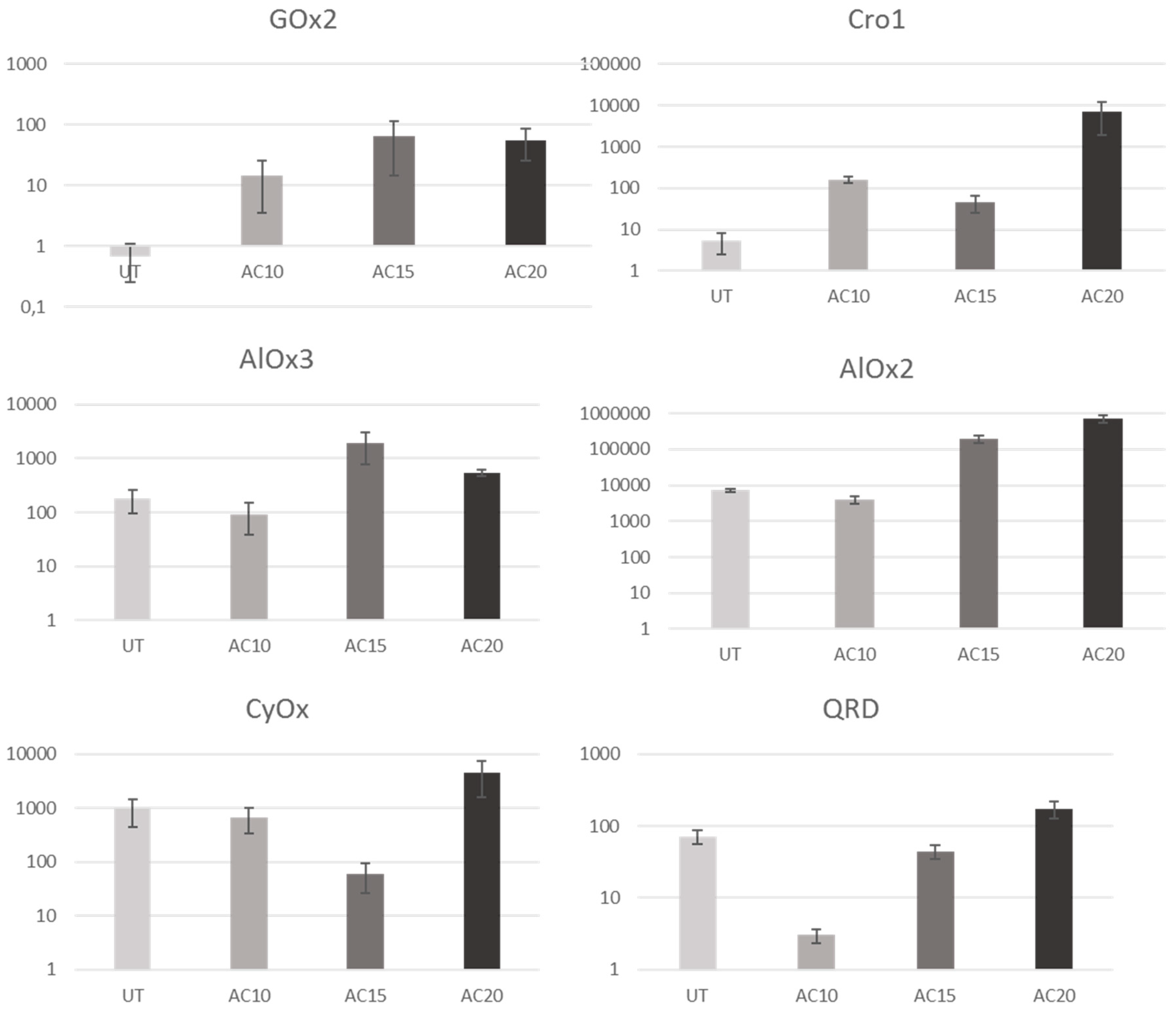
3.2.1. Upregulated Genes
3.2.2. Downregulated Genes
3.2.3. Effect of Acetylation on Overall Gene Expression
4. Discussion
4.1. Mass Loss and Growth Rates
4.2. Expression of Target Genes
4.2.1. Upregulated Genes
4.2.2. Downregulated Genes
4.2.3. Effect of Acetylation on Overall Gene Expression
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rowell, R.M. Wood Chemistry and Wood Composites; Rowell, R.M., Ed.; Taylor & Francis: Boca Raton, FL, USA, 2005; p. 703. [Google Scholar] [CrossRef]
- Huß, W.; Krötsch, S. Energieeffizientes Sanieren und Bauen mit Holz. LWF Aktuell 2013, 97, 4. [Google Scholar]
- Jakes, J.E.; Arzola, X.; Bergman, R.; Ciesielski, P.; Hunt, C.G.; Rahbar, N.; Tshabalala, M.; Wiedenhoeft, A.C.; Zelinka, S.L. Not Just Lumber—Using Wood in the Sustainable Future of Materials, Chemicals, and Fuels. JOM 2016, 68, 10. [Google Scholar] [CrossRef] [Green Version]
- Bergman, R.; Puettmann, M.; Taylor, A.; Skog, K. The Carbon Impacts of Wood Products. For. Prod. J. 2014, 64, 13. [Google Scholar] [CrossRef]
- Zabel, R.A.; Morrell, J.J. Wood Microbiology: Decay and Its Prevention; Academic Press: San Diego, CA, USA, 1992. [Google Scholar]
- Eaton, R.; Hale, M. Decay, Pests and Protection; Chapman & Hall: Cambridge, UK, 1993. [Google Scholar]
- Lebow, S. Alternatives to Chromated Copper Arsenate (CCA) for Residential Construction. In Proceedings of the Environmental Impacts of Preservative-Treated Wood, Orlando, FL, USA, 8–10 February 2004; p. 12. [Google Scholar]
- Hill, C. Wood Modification: Chemical, Thermal and other Processes; Stevens, C.V., Ed.; John Wiley and Sons, Ltd.: Hoboken, NJ, USA, 2006. [Google Scholar]
- Townsend, T.; Dubey, B.; Tolaymat, T.; Solo-Gabriele, H. Preservative leaching from weathered CCA-treated wood. J. Environ. Manag. 2005, 75, 105–113. [Google Scholar] [CrossRef]
- Mantanis, G.I. Chemical modification of wood by acetylation or furfurylation: A review of the present scaled-up technologies. Bioresources 2017, 12, 12. [Google Scholar] [CrossRef] [Green Version]
- Alfredsen, G.; Pilgård, A.; Fossdal, C.G. Characterisation of Postia placenta colonisation during 36 weeks in acetylated southern yellow pine sapwood at three acetylation levels including genomic DNA and gene expression quantification of the fungus. Holzforschung 2016, 70, 11. [Google Scholar] [CrossRef] [Green Version]
- Hosseinpourpia, R.; Mai, C. Mode of action of brown rot decay resistance of acetylated wood: Resistance to Fenton’s reagent. Wood Sci. Technol. 2016, 50, 413–426. [Google Scholar] [CrossRef]
- Beck, G.; Hegnar, O.A.; Fossdal, C.G.; Alfredsen, G. Acetylation of Pinus radiata delays hydrolytic depolymerisation by the brown-rot fungus Rhondonia placenta. Int. Biodeterior. Biodegrad. 2018, 135, 39–52. [Google Scholar] [CrossRef]
- Engelund, E.; Thygesen, L.G.; Svensson, S.; Hill, C. A critical discussion of the physics of wood-water interactions. Wood Sci. Technol. 2013, 47, 21. [Google Scholar] [CrossRef] [Green Version]
- Popescu, C.-M.; Hill, C.A.S.; Curling, S.; Ormondroyd, G.; Xie, Y. The water vapour sorption behaviour of acetylated birch wood: How acetylation affects the sorption isotherm and accessible hydroxyl content. J. Mater. Sci. 2014, 49, 2362–2371. [Google Scholar] [CrossRef]
- Beck, G.; Thybring, E.; Thygesen, L.; Hill, C. Characterization of moisture in acetylated and propionylated radiata pine using low-field nuclear magnetic resonance (LFNMR) relaxometry. Holzforschung 2017. [Google Scholar] [CrossRef]
- Stamm, A.J.; Baechler, R.H. Decay resistance and dimensional stability of five modified woods. For. Prod. J. 1960, 10, 22–26. [Google Scholar]
- Ibach, R.E.; Rowell, R.M. Improvements in Decay Resistance Based on Moisture Exclusion. Mol. Cryst. Liq. Cryst. Sci. Technol. Sect. A Mol. Cryst. Liq. Cryst. 2000, 353, 23–33. [Google Scholar] [CrossRef]
- Larsson Brelid, P.; Simonson, R.; Bergman, Ö.; Nilsson, T. Resistance of acetylated wood to biological degradation. Holz Als Roh- Und Werkst. 2000, 58, 331–337. [Google Scholar] [CrossRef]
- Hill, C. Why does acetylation protect wood from microbiological attack? Wood Mater. Sci. Eng. 2009, 4, 37–45. [Google Scholar] [CrossRef]
- Papadopoulos, A.N.; Hill, C.A.S. The biological effectiveness of wood modified with linear chain carboxylic acid anhydrides against Coniophora puteana. Holz Als Roh- Und Werkst. 2002, 60, 329–332. [Google Scholar] [CrossRef]
- Jakes, J.; Plaza, N.; Stone, D.S.; Hunt, C.; Glass, S.; Zelinka, S. Mechanism of Transport Through Wood Cell Wall Polymers. J. For. Prod. Ind. 2013, 2, 10–13. [Google Scholar]
- Xie, Y.; Xiao, Z.; Mai, C. Degradation of chemically modified Scots pine (Pinus sylvestris L.) with Fenton reagent. Holzforschung 2015, 69, 153. [Google Scholar] [CrossRef]
- Zelinka, S.L.; Ringman, R.; Pilgård, A.; Thybring, E.E.; Jakes, J.E.; Richter, K. The role of chemical transport in the brown-rot decay resistance of modified wood AU - Zelinka, S.L. Int. Wood Prod. J. 2016, 7, 66–70. [Google Scholar] [CrossRef]
- Ringman, R.; Pilgård, A.; Brischke, C.; Richter, K. Mode of action of brown rot decay resistance in modified wood: A review. Holzforschung 2014, 68, 239. [Google Scholar] [CrossRef]
- Hunt, C.; Zelinka, S.; Frihart, C.; Lorenz, L.; Yelle, D.; Gleber, S.-C.; Vogt, S.; Jakes, J.E. Acetylation increases relative humidity threshold for ion transport in wood cell walls—A means to understanding decay resistance. Int. Biodeterior. Biodegrad. 2018, 133, 230–237. [Google Scholar] [CrossRef]
- Zelinka, S.; Glass, S.; Stone, D. A percolation model for electrical conduction in wood with implications for wood-water relations. Wood Fiber Sci. J. Soc. Wood Sci. Technol. 2008, 40, 544–552. [Google Scholar]
- Zelinka, S.L.; Gleber, S.C.; Vogt, S.; López, G.M.R.; Jakes, J.E. Threshold for ion movements in wood cell walls below fiber saturation observed by X-ray fluorescence microscopy (XFM). Holzforschung 2015, 69, 441–448. [Google Scholar] [CrossRef]
- Kirk, P.M.; Cannon, P.F.; Minter, D.W.; Stalpers, J.A. Dictionary of the Funghi, 10th ed.; CAB International: Wallingford, UK, 2008. [Google Scholar]
- Eriksson, K.; Blanchette, R.A.; Ander, P. Microbial and Enzymatic Degradation of Wood and Wood Components; Springer: Berlin, Germany, 1990. [Google Scholar]
- Blanchette, R. Delignification by Wood-Decay Fungi. Annu. Rev. Phytopathol. 1991, 29, 18. [Google Scholar] [CrossRef]
- Daniel, G. Use of electron microscopy for aiding our understanding of wood biodegradation. FEMS Microbiol. Rev. 1994, 13, 199–233. [Google Scholar] [CrossRef]
- Riley, R.; Salamov, A.A.; Brown, D.W.; Nagy, L.G.; Floudas, D.; Held, B.W.; Levasseur, A.; Lombard, V.; Morin, E.; Otillar, R.; et al. Extensive sampling of basidiomycete genomes demonstrates inadequacy of the white-rot/brown-rot paradigm for wood decay fungi. Proc. Natl. Acad. Sci. USA 2014, 111, 9923–9928. [Google Scholar] [CrossRef] [Green Version]
- Liese, W. Ultrastructural Aspects of Woody Tissue Disintegration. Annu. Rev. Phytopathol. 1970, 8, 231–258. [Google Scholar] [CrossRef]
- Martin, F. Fair Trade in the Underworld: The Ectomycorrhizal Symbiosis. In Biology of the Fungal Cell; Howard, R.J., Gow, N.A.R., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 291–308. [Google Scholar] [CrossRef]
- Schmidt, O. Indoor wood-decay basidiomycetes: Damage, causal fungi, physiology, identification and characterization, prevention and control. Mycol. Prog. 2007, 6, 261. [Google Scholar] [CrossRef]
- Alfredsen, G.; Solheim, H.; Mohn Jenssen, K. Evaluation of decay fungi in Norwegian buildings. In Proceedings of the IRG Annual Meeting, IRG/WP 09-10701, Bangalore, India, 24–28 April 2015; p. 11. [Google Scholar]
- Vanden Wymelenberg, A.; Gaskell, J.; Mozuch, M.; Sabat, G.; Ralph, J.; Skyba, O.; Mansfield, S.D.; Blanchette, R.A.; Martinez, D.; Grigoriev, I.; et al. Comparative Transcriptome and Secretome Analysis of Wood Decay Fungi Postia placenta and Phanerochaete chrysosporium. Appl. Environ. Microbiol. 2010, 76, 3599–3610. [Google Scholar] [CrossRef] [Green Version]
- Goodell, B. Brown-Rot Fungal Degradation of Wood: Our Evolving View. In Wood Deterioration and Preservation; American Chemical Society: Washington, DC, USA, 2003; Volume 845, pp. 97–118. [Google Scholar]
- Filley, T.R.; Cody, G.D.; Goodell, B.; Jellison, J.; Noser, C.; Ostrofsky, A. Lignin demethylation and polysaccharide decomposition in spruce sapwood degraded by brown rot fungi. Org. Geochem. 2002, 33, 111–124. [Google Scholar] [CrossRef]
- Curling, S.F.; Clausen, C.A.; Winandy, J.E. Relationships between mechanical properties, weight loss, and chemical composition of wood during incipient brown-rot decay. For. Prod. J. 2002, 52, 34–39. [Google Scholar]
- Martínez, Á.T.; Speranza, M.; Ruiz-Dueñas, F.J.; Ferreira, P.; Camarero, S.; Guillén, F.; Martínez, M.J.; Gutiérrez, A.; Del Río, J.C. Biodegradation of lignocellulosics: Microbial, chemical, and enzymatic aspects of the fungal attack of lignin. Int. Microbiol. 2005, 8, 195–204. [Google Scholar] [PubMed]
- Niemenmaa, O.; Uusi-Rauva, A.; Hatakka, A. Demethoxylation of [O14CH3]-labelled lignin model compounds by the brown-rot fungi Gloeophyllum trabeum and Poria (Postia) placenta. Biodegradation 2007, 19, 555. [Google Scholar] [CrossRef] [PubMed]
- Arantes, V.; Goodell, B. Current Understanding of Brown-Rot Fungal Biodegradation Mechanisms: A Review. In Deterioration and Protection of Sustainable Biomaterials; American Chemical Society: Washington, DC, USA, 2014; Volume 1158, pp. 3–21. [Google Scholar]
- Baldrian, P.; Valášková, V. Degradation of cellulose by basidiomycetous fungi. FEMS Microbiol. Rev. 2008, 32, 501–521. [Google Scholar] [CrossRef] [Green Version]
- Arantes, V.; Jellison, J.; Goodell, B. Peculiarities of brown-rot fungi and biochemical Fenton reaction with regard to their potential as a model for bioprocessing biomass. Appl. Microbiol. Biotechnol. 2012, 94, 323–338. [Google Scholar] [CrossRef]
- Zhang, J.; Presley, G.N.; Hammel, K.E.; Ryu, J.-S.; Menke, J.R.; Figueroa, M.; Hu, D.; Orr, G.; Schilling, J.S. Localizing gene regulation reveals a staggered wood decay mechanism for the brown rot fungus Postia placenta. Proc. Natl. Acad. Sci. USA 2016, 113, 10968–10973. [Google Scholar] [CrossRef] [Green Version]
- Eastwood, D.C.; Floudas, D.; Binder, M.; Majcherczyk, A.; Schneider, P.; Aerts, A.; Asiegbu, F.O.; Baker, S.E.; Barry, K.; Bendiksby, M.; et al. The Plant Cell Wall–Decomposing Machinery Underlies the Functional Diversity of Forest Fungi. Science 2011, 333, 762–765. [Google Scholar] [CrossRef] [Green Version]
- Goodell, B.; Jellison, J.; Liu, J.; Daniel, G.; Paszczynski, A.; Fekete, F.; Krishnamurthy, S.; Jun, L.; Xu, G. Low molecular weight chelators and phenolic compounds isolated from wood decay fungi and their role in the fungal biodegradation of wood1This is paper 2084 of the Maine Agricultural and Forest Experiment Station.1. J. Biotechnol. 1997, 53, 133–162. [Google Scholar] [CrossRef]
- Paszczynski, A.; Crawford, R.; Funk, D.; Goodell, B. De novo synthesis of 4,5-dimethoxycatechol and 2, 5-dimethoxyhydroquinone by the brown rot fungus Gloeophyllum trabeum. Appl. Environ. Microbiol. 1999, 65, 674–679. [Google Scholar]
- Jensen, K.A.; Houtman, C.J.; Ryan, Z.C.; Hammel, K.E. Pathways for Extracellular Fenton Chemistry in the Brown Rot Basidiomycete Gloeophyllum trabeum. Appl. Environ. Microbiol. 2001, 67, 2705–2711. [Google Scholar] [CrossRef] [Green Version]
- Fenton, H. Oxidation of tartaric acid in the presence of iron. J. Chem. Soc. Trans. 1894, 65, 12. [Google Scholar] [CrossRef] [Green Version]
- Goodell, B.; Zhu, Y.; Kim, S.; Kafle, K.; Eastwood, D.C.; Daniel, G.; Jellison, J.; Yoshida, M.; Groom, L.; Pingali, S.V.; et al. Modification of the nanostructure of lignocellulose cell walls via a non-enzymatic lignocellulose deconstruction system in brown rot wood-decay fungi. Biotechnol. Biofuels 2017, 10, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, D.; Challacombe, J.; Morgenstern, I.; Hibbett, D.S.; Schmoll, M.; Kubicek, C.P.; Ferreira, P.; Ruiz-Dueñas, F.J.; Martinez, A.T.; Kersten, P.; et al. Genome, transcriptome, and secretome analysis of wood decay fungus Postia placenta supports unique mechanisms of lignocellulose conversion. Proc. Natl. Acad. Sci. USA 2009, 106, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jensen, K.; Ryan, Z.; Marty, A.; Cullen, D.; Hammel, K.E. An NADH: Quinone Oxidoreductase Active during Biodegradation by the Brown-Rot Basidiomycete Gloeophyllum trabeum. Appl. Environ. Microbiol. 2002, 68, 2699–2703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, W.; Jellison, J. Characterization of a transplasma membrane redox system of the brown rot fungus Gloeophyllum trabeum. Int. Biodeterior. Biodegrad. 2004, 53, 37–42. [Google Scholar] [CrossRef]
- Cohen, R.; Suzuki, M.R.; Hammel, K.E. Differential stress-induced regulation of two quinone reductases in the brown rot basidiomycete Gloeophyllum trabeum. Appl. Environ. Microbiol. 2004, 70, 324–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mueckler, M.; Caruso, C.; Baldwin, S.A.; Panico, M.; Blench, I.; Morris, H.R.; Allard, W.J.; Lienhard, G.E.; Lodish, H.F. Sequence and structure of a human glucose transporter. Science 1985, 229, 941. [Google Scholar] [CrossRef]
- Yan, N. Structural Biology of the Major Facilitator Superfamily Transporters. Annu. Rev. Biophys. 2015, 44, 257–283. [Google Scholar] [CrossRef]
- Wardman, P. Reduction Potentials of One-Electron Couples Involving Free Radicals in Aqueous Solution. J. Phys. Chem. Ref. Data 1989, 18, 1637–1755. [Google Scholar] [CrossRef] [Green Version]
- Wei, D.; Houtman, C.J.; Kapich, A.; Hunt, C.; Cullen, D.; Hammel, K.E. Laccase and Its Role in Production of Extracellular Reactive Oxygen Species during Wood Decay by the Brown Rot Basidiomycete Postia placenta. Appl. Environ. Microbiol. 2010, 76, 2091–2097. [Google Scholar] [CrossRef] [Green Version]
- Floudas, D.; Binder, M.; Riley, R.; Barry, K.; Blanchette, R.A.; Henrissat, B.; Martínez, A.T.; Otillar, R.; Spatafora, J.W.; Yadav, J.S.; et al. The Paleozoic Origin of Enzymatic Lignin Decomposition Reconstructed from 31 Fungal Genomes. Science 2012, 336, 1715–1719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levasseur, A.; Drula, E.; Lombard, V.; Coutinho, P.M.; Henrissat, B. Expansion of the enzymatic repertoire of the CAZy database to integrate auxiliary redox enzymes. Biotechnol. Biofuels 2013, 6, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takao, S. Organic Acid Production by Basidiomycetes. Appl. Microbiol. 1965, 13, 732. [Google Scholar] [PubMed]
- Akamatsu, Y.; Shimada, M. Partial purification and characterization of glyoxylate oxidase from the brown-rot basidiomycete Tyromyces palustris. Phytochemistry 1994, 37, 649–653. [Google Scholar] [CrossRef]
- Munir, E.; Yoon, J.-J.; Tokimatsu, T.; Hattori, T.; Shimada, M. New role for glyoxylate cycle enzymes in wood-rotting basidiomycetes in relation to biosynthesis of oxalic acid. J. Wood Sci. 2001, 47, 368–373. [Google Scholar] [CrossRef]
- Tokimatsu, T.; Nagai, Y.; Hattori, T.; Shimada, M. Purification and characteristics of a novel cytochrome c dependent glyoxylate dehydrogenase from a wood-destroying fungus Tyromyces palustris 1. FEBS Lett. 1998, 437, 117–121. [Google Scholar] [CrossRef] [Green Version]
- Pilgård, A.; Alfredsen, G.; Fossdal, C.G.; Long, I.C.J. The effects of acetylation on the growth of Postia placenta over 36 weeks. In Proceedings of the IRG Annual Meeting, Kuala Lumpur, Malaysia, 6–10 May 2012. IRG/WP 12-40589. [Google Scholar]
- Schmöllerl, B.; Alfredsen, G.; Fossdal, C.G.; Westin, M.; Steitz, A. Molecular investigation of Postia placenta growing in modified wood. In Proceedings of the IRG Annual Meeting, IRG/WP 11-10756, Queenstown, New Zealand, 8–12 May 2018; p. 10. [Google Scholar]
- Ringman, R.; Pilgård, A.; Richter, K. Effect of wood modification on gene expression during incipient Postia placenta decay. Int. Biodeterior. Biodegrad. 2014, 86, 86–91. [Google Scholar] [CrossRef]
- Ringman, R.; Pilgård, A.; Brischke, C.; Windeisen, E.; Richter, K. Incipient brown rot decay in modified wood: Patterns of mass loss, structural integrity, moisture and acetyl content in high resolution. Int. Wood Prod. J. 2017. [Google Scholar] [CrossRef]
- Bravery, A.F. A miniaturised wood-block test for the rapid evaluation of preservative fungicides. In Screening Techniques for Potential Wood Preservative Chemicals. Proceedings of the a Special Seminar Held in Association with the 10th Annual Meeting of IRG, Peebles, UK, 18−22 September 1978; IRG: Washington, DC, USA, 1978. [Google Scholar]
- Alfredsen, G.; Pilgård, A. Postia placenta decay of acetic anhydride modified wood—Effect of leaching. Wood Mater. Sci. Eng. 2014. [Google Scholar] [CrossRef]
- Schilling, J.S.; Duncan, S.M.; Presley, G.N.; Filley, T.R.; Jurgens, J.A.; Blanchette, R.A. Colocalizing incipient reactions in wood degraded by the brown rot fungus Postia placenta. Int. Biodeterior. Biodegrad. 2013, 83, 56–62. [Google Scholar] [CrossRef]
- Alfredsen, G.; Ringman, R.; Pilgård, A.; Fossdal, C.G. New insight regarding mode of action of brown rot decay of modified wood based on DNA and gene expression studies: A review. Int. Wood Prod. J. 2015, 6, 5–7. [Google Scholar] [CrossRef]
- Ringman, R.; Pilgård, A.; Kölle, M.; Richter, K. Expression patterns of Postia placenta genes involved in the chelator mediated Fenton degradation in modified wood. In Proceedings of the European Conference on Wood Modification, Arnhem, The Netherlands, 17–18 September 2019; p. 3. [Google Scholar]
- Ringman, R.; Pilgård, A.; Kölle, M.; Brischke, C.; Richter, K. Effects of thermal modification on Postia placenta wood degradation dynamics: Measurements of mass loss, structural integrity and gene expression. Wood Sci. Technol. 2016, 50, 385–397. [Google Scholar] [CrossRef]
- Alfredsen, G.; Fossdal, C.G.; Nahy, N.E.; Jellison, J.; Goodell, B. Furfurylated wood: Impact on Postia placenta gene expression and oxalate crystal formation. Holzforschung 2016, 70, 16. [Google Scholar] [CrossRef]
- Pilgård, A.; Schmöllerl, B.; Risse, M.; Fossdal, C.G.; Alfredsen, G. Profiling Postia placenta colonisation in modified wood—microscopy, DNA quantification and gene expression. In Proceedings of the Wood Science and Engineering Conference, Copenhagen, Denmark, 17–18 September 2019. [Google Scholar]
- Salame, T.M.; Knop, D.; Levinson, D.; Yarden, O.; Hadar, Y. Redundancy among manganese peroxidases in Pleurotus ostreatus. Appl. Environ. Microbiol. 2013, 79, 2405. [Google Scholar] [PubMed] [Green Version]
- Ringman, R.; Beck, G.; Pilgård, A. The Importance of Moisture for Brown Rot Degradation of Modified Wood: A Critical Discussion. Forests 2019, 10, 522. [Google Scholar] [CrossRef] [Green Version]
- Arantes, V.; Qian, Y.; Milagres, A.M.F.; Jellison, J.; Goodell, B. Effect of pH and oxalic acid on the reduction of Fe3+ by a biomimetic chelator and on Fe3+ desorption/adsorption onto wood: Implications for brown-rot decay. Int. Biodeterior. Biodegrad. 2009, 63, 478–483. [Google Scholar] [CrossRef]
- Narayanan, B.C.; Niu, W.; Han, Y.; Zou, J.; Mariano, P.S.; Dunaway-Mariano, D.; Herzberg, O. Structure and function of PA4872 from Pseudomonas aeruginosa, a novel class of oxaloacetate decarboxylase from the PEP mutase/isocitrate lyase superfamily. Biochemistry 2008, 47, 167–182. [Google Scholar] [CrossRef] [Green Version]
- Munir, E.; Yoon, J.; Tokimatsu, T.; Hattori, T.; Shimada, M. A physiological role for oxalic acid biosynthesis in the wood-rotting basidiomycete Fomitopsis palutris. Proc. Natl. Acad. Sci. USA 2001, 98, 11126–11130. [Google Scholar] [CrossRef] [Green Version]
- Schilling Jonathan, S.; Jellison, J. High-performance liquid chromatographic analysis of soluble and total oxalate in Ca- and Mg-amended liquid cultures of three wood decay fungi. Holzforschung 2004, 58, 682. [Google Scholar] [CrossRef]
- Zhang, J.; Schilling, J. Role of carbon source in the shift from oxidative to hydrolytic wood decomposition by Postia placenta. Fungal Genet. Biol. 2017, 106. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Mean WPG | Standard Deviation |
|---|---|
| 10 | 0.18 |
| 15 | 0.13 |
| 20 | 0.14 |
| Gene | JGI no. | Primer Sequence |
|---|---|---|
| β-tubulin (bT) | 113871 | CAGGATCTTGTCGCCGAGTAC/CCTCATACTCGCCCTCCTCTT |
| Quinone oxidoreductase (QRD) | 124517 | CGACGACAAGCCCAACAAG/GATGACGATGATGGCGATTTTAGG |
| Alcohol oxidase 1 (AlOx1) | 44331 | GGAGGTACAGACGGACGAAC/AGAGTCGACGACACCGTTCT |
| Alcohol oxidase 2 (AlOx2) | 129158 | TACTCGACGGCCCTCACTAT/CCGCTTGAGACTGAACACTG |
| Alcohol oxidase 3 (AlOx3) | 118723 | ACACCAAGGAGGACGACGAG/GACGAGCAAGGCAGACGAGTA |
| Putative quinate transporter (PQT) | 44553 | ACTGACCTTTTGCGCAGACT/CAATGTTGATTGTGGCGAAC |
| Laccase (Lac) | 111314 | CGGTGCTCTTGGCCACTTAG/CCATTGGTTATGGGCAGCTC |
| Copper radical oxidase (Cro1) | 56703 | CCTACCAGCTGCTTCCTGAC/AACGTTCGGCTGTATGAACC |
| Glucose oxidase (GOx2) | 108489 | GTCCGCTCTAACGTTGCTTC/CCGGCGTTATTGGAGAGATA |
| Glyoxylate dehydrogenase (GlyD) | 121561 | CGGAGCTGGACCTTTGTTAC/GCGCGAAGGCAAATCTAATA |
| Oxaloacetate dehydrogenase (CyOx) | 112832 | AAGGCGTTCTTCGAGGTCAT/AAAGCAGCAACCCGAGAAG |
| Treatment | Mass Loss (%) | Growth Rate [mm/day] | Growth Rate R2 Values |
|---|---|---|---|
| untreated | 9.36 | 0.23 | 0.9955 |
| 10% acetylated | 1.63 | 0.24 | 0.9955 |
| 15% acetylated | 0.25 | 0.22 | 0.91 |
| 20% acetylated | −0.16 | 0.25 | 0.9897 |
| AC10 | AC15 | AC20 | Gene | |
|---|---|---|---|---|
| UT | 2.08 × 10−9 | 7.33 × 10−9 | 2.27 × 10−9 | |
| AC10 | 7.3 × 10−6 | 9.81 × 10−9 | GlyD | |
| AC15 | 1.14 × 10−5 | |||
| UT | 3.60 × 10−5 | 5.85 × 10−4 | 4.07 × 10−5 | |
| AC10 | 3.48 × 10−4 | 5.19 × 10−4 | GOx2 | |
| AC15 | 1.37 × 10−1 | |||
| UT | 1.29 × 10−1 | 3.14 × 10−6 | 4.03 × 10−4 | |
| AC10 | 3.16 × 10−6 | 2.21 × 10−4 | CyOx | |
| AC15 | 4.53 × 10−5 | |||
| UT | 8.95 × 10−9 | 3.26 × 10−6 | 1.50 × 10−4 | |
| AC10 | 9.69 × 10−9 | 1.88 × 10−4 | Cro1 | |
| AC15 | 1.60 × 10−4 | |||
| UT | 5.64 × 10−7 | 6.44 × 10−7 | 1.21 × 10−6 | |
| AC10 | 7.38 × 10−6 | 3.18 × 10−4 | Lac1 | |
| AC15 | 1.41 × 10−3 | |||
| UT | 1.32 × 10−6 | 1.26 × 10−6 | 3.13 × 10−6 | |
| AC10 | 2.25 × 10−7 | 6.08 × 10−4 | PQT | |
| AC15 | 4.23 × 10−4 | |||
| UT | 7.86 × 10−3 | 7.71 × 10−5 | 1.19 × 10−12 | |
| AC10 | 5.48 × 10−5 | 3.75 × 10−14 | AlOx3 | |
| AC15 | 5.32 × 10−4 | |||
| UT | 4.70 × 10−11 | 1.59 × 10−10 | 3.72 × 10−10 | |
| AC10 | 1.24 × 10−10 | 3.51 × 10−10 | AlOx2 | |
| AC15 | 7.04 × 10−9 | |||
| UT | 1.83 × 10−12 | 3.5 × 10−1 | 1.71 × 10−11 | |
| AC10 | 1.39 × 10−8 | 6.56 × 10−12 | AlOx1 | |
| AC15 | 1.63 × 10−7 | |||
| UT | 1.26 × 10−11 | 4.99 × 10−6 | 3.48 × 10−8 | |
| AC10 | 1.79 × 10−8 | 1.52 × 10−10 | QRD | |
| AC15 | 2.27 × 10−9 | |||
| Highly significant | Significant | Not significant | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kölle, M.; Ringman, R.; Pilgård, A. Initial Rhodonia placenta Gene Expression in Acetylated Wood: Group-Wise Upregulation of Non-Enzymatic Oxidative Wood Degradation Genes Depending on the Treatment Level. Forests 2019, 10, 1117. https://doi.org/10.3390/f10121117
Kölle M, Ringman R, Pilgård A. Initial Rhodonia placenta Gene Expression in Acetylated Wood: Group-Wise Upregulation of Non-Enzymatic Oxidative Wood Degradation Genes Depending on the Treatment Level. Forests. 2019; 10(12):1117. https://doi.org/10.3390/f10121117
Chicago/Turabian StyleKölle, Martina, Rebecka Ringman, and Annica Pilgård. 2019. "Initial Rhodonia placenta Gene Expression in Acetylated Wood: Group-Wise Upregulation of Non-Enzymatic Oxidative Wood Degradation Genes Depending on the Treatment Level" Forests 10, no. 12: 1117. https://doi.org/10.3390/f10121117
APA StyleKölle, M., Ringman, R., & Pilgård, A. (2019). Initial Rhodonia placenta Gene Expression in Acetylated Wood: Group-Wise Upregulation of Non-Enzymatic Oxidative Wood Degradation Genes Depending on the Treatment Level. Forests, 10(12), 1117. https://doi.org/10.3390/f10121117