Shallow Genetic Structure among the European Populations of the Six-Toothed Bark Beetle Ips sexdentatus (Coleoptera, Curculionidae, Scolytinae)

 ,
,  ,
,  ,
,  ,
, 
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling—DNA Analysis
2.2. Data Analysis
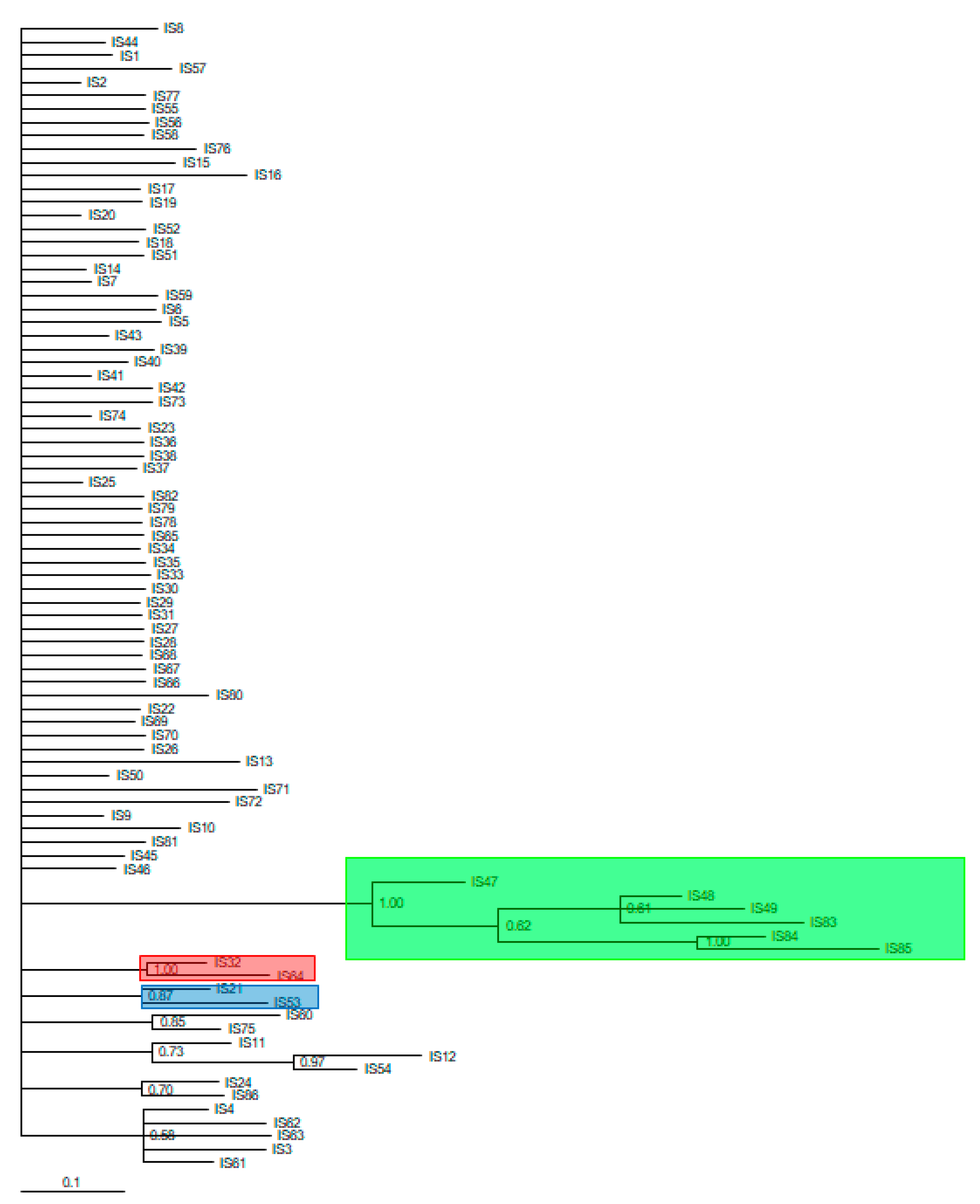
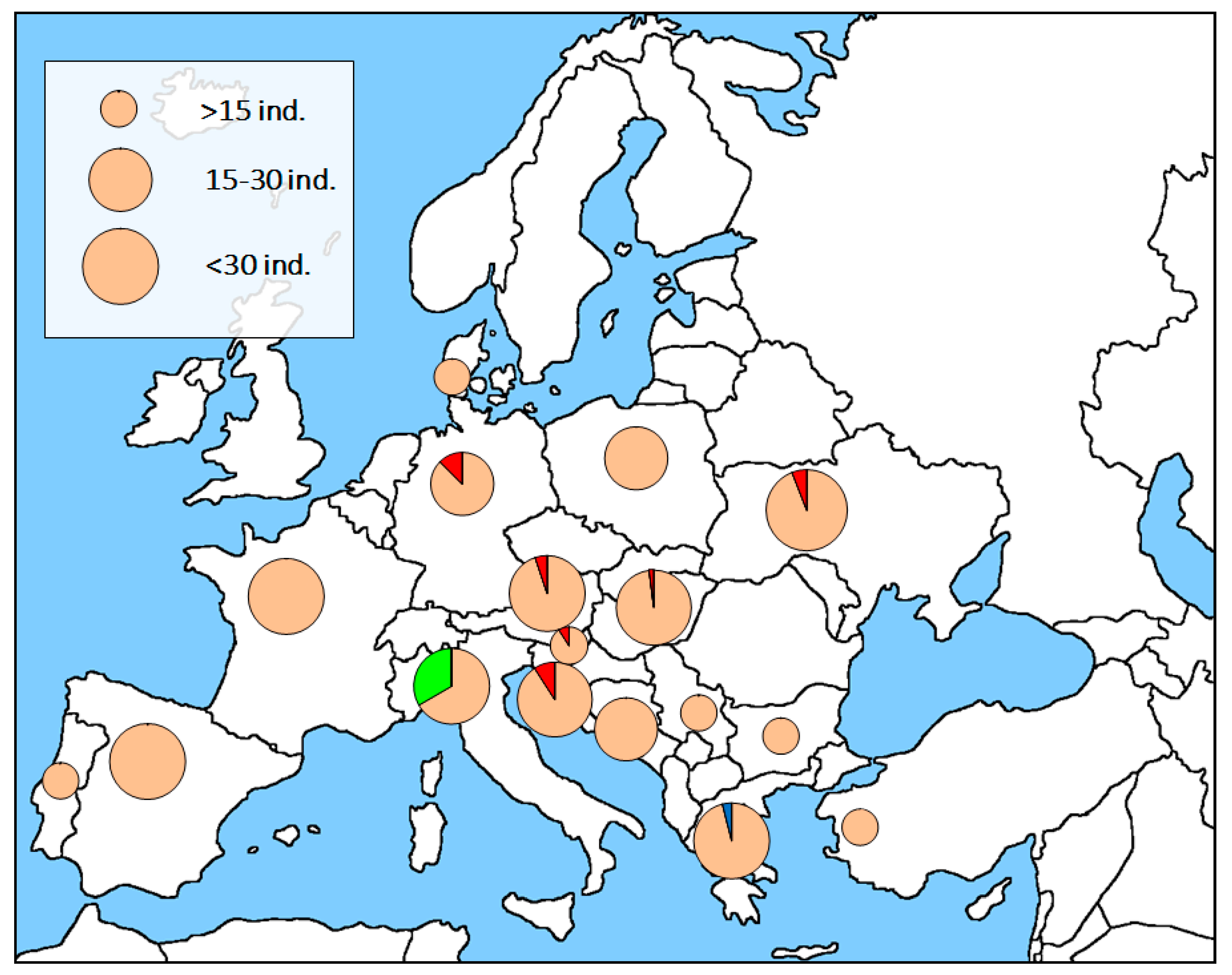
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Taberlet, P.; Fumagalli, L.; Wust-Saucy, A.-G.; Cosson, J.-F. Comparative phylogeography and postglacial colonization routes in Europe. Mol. Ecol. 1998, 7, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, G.M. The genetic legacy of the Quaternary ice ages. Nature 2000, 405, 907–913. [Google Scholar] [CrossRef]
- Stewart, J.R.; Lister, A.M.; Barnes, I.; Dalén, L. Refugia revisited: Individualistic responses of species in space and time. Proc. R. Soc. Lond. B Biol. Sci. 2010, 277, 661–671. [Google Scholar] [CrossRef] [PubMed]
- Tzedakis, P.C.; Emerson, B.C.; Hewitt, G.M. Cryptic or mystic? Glacial tree refugia in northern Europe. Trends Ecol. Evol. 2013, 28, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Kajtoch, Ł.; Cieślak, E.; Varga, Z.; Paul, W.; Mazur, M.A.; Sramkó, G.; Kubisz, D. Phylogeographic patterns of steppe species in Eastern Central Europe: A review and the implications for conservation. Biodivers. Conserv. 2016, 25, 2309–2339. [Google Scholar] [CrossRef]
- Médail, F.; Diadema, K. Glacial refugia influence plant diversity patterns in the Mediterranean Basin. J. Biogeogr. 2009, 36, 1333–1345. [Google Scholar] [CrossRef]
- Theissinger, K.; Balint, M.; Feldheim, K.A.; Haase, P.; Johannesen, J.; Laube, I.; Pauls, S.U. Glacial survival and post-glacial recolonization of an arctic-alpine freshwater insect (Arcynopteryx dichroa, Plecoptera, Perlodidae) in Europe. J. Biogeogr. 2012, 40, 236–248. [Google Scholar] [CrossRef]
- Bertheau, C.; Bankhead-Dronnet, S.; Martin, C.; Lieutier, F.; Roux-Morabito, G. Lack of genetic differentiation after host range extension argues for the generalist nature of Pityogenes chalcographus (Curculionidae: Scolytinae). Ann. For. Sci. 2012, 69, 313–323. [Google Scholar] [CrossRef]
- Daneck, H.; Fer, T.; Marhold, K. Glacial survival in northern refugia? Phylogeography of the temperate shrub Rosa pendulina L. (Rosaceae): AFLP vs. chloroplast DNA variation. Biol. J. Linn. Soc. 2016, 119, 704–718. [Google Scholar] [CrossRef]
- Rousselet, J.; Zhao, R.; Argal, D.; Simonato, M.; Battisti, A.; Roques, A.; Kerdelhue, C. The role of topography in structuring the demographic history of the pine processionary moth, Thaumetopoea pityocampa (Lepidoptera, Notodontidae). J. Biogeogr. 2010, 37, 1478–1490. [Google Scholar] [CrossRef]
- Brockerhoff, E.G.; Bain, J.; Kimberley, M.; Knižek, M. Interception frequency of exotic ark and ambrosia beetles (Coleoptera: Scolytinae) and relationship with establishment in New Zealand and worldwide. Can. J. For. Res. 2006, 36, 289–298. [Google Scholar] [CrossRef]
- Fettig, C.J.; Klepzig, K.D.; Billings, R.F.; Munson, A.S.; Nebeker, T.E.; Negrón, J.F.; Nowak, J.T. The effectiveness of vegetation management practices for prevention and control of bark beetle infestations in coniferous forests of the western and southern United States. For. Ecol. Manag. 2007, 238, 24–53. [Google Scholar] [CrossRef]
- Abdulah, H.; Darvishzadeh, R.; Skidmore, A.K.; Groen, T.A.; Heurich, M. European spruce bark beetle (Ips typographus L.) green attack affects foliar reflectance and biochemical properties. Int. J. Appl. Earth Obs. Geoinf. 2018, 64, 199–209. [Google Scholar] [CrossRef]
- Kurz, W.A.; Dymond, C.C.; Stinson, G. Mountain pine beetle and forest carbon: Feedback to climate change. Nature 2008, 454, 987–990. [Google Scholar] [CrossRef] [PubMed]
- Waring, K.M.; Reboletti, D.M.; Mork, L.A.; Huang, C.-H.; Hofstetter, R.W.; Garcia, A.M.; Fulé, P.Z.; Davis, T.S. Modeling the impacts of two bark beetle species under a warming climate in the southwestern USA: Ecological and economic consequences. Environ. Manag. 2009, 44, 824–835. [Google Scholar] [CrossRef] [PubMed]
- Vega, F.E.; Hofstetter, R.W. Bark Beetles: Biology and Ecology of Native and Invasive Species, 1st ed.; Elsevier Academic Press: Oxford, UK, 2015. [Google Scholar]
- Sallé, A.; Arthofer, W.; Lieutier, F.; Stauffer, C.; Kerdelhué, C. Phylogeography of a host-specific insect: Genetic structure of Ips typographus in Europe does not reflect past fragmentation of its host. Biol. J. Linn. Soc. 2007, 90, 239–246. [Google Scholar] [CrossRef]
- Cullingham, C.I.; Roe, A.D.; Sperling, F.A.H.; Coltman, D.W. Phylogeographic insights into an irruptive pest outbreak. Ecol. Evol. 2012, 2, 908–919. [Google Scholar] [CrossRef] [PubMed]
- Bertheau, C.; Schuler, H.; Arthofer, W.; Avtzis, D.N.; Mayer, F.; Krumböck, S.; Moodley, Y.; Stauffer, C. Divergent evolutionary histories of two sympatric spruce bark beetle species. Mol. Ecol. 2013, 22, 3318–3332. [Google Scholar] [CrossRef] [PubMed]
- Kerdelhué, C.; Roux-Morabito, G.; Forichon, J.; Chambon, J.-M.; Robert, A.; Lieutier, F. Population genetic structure of Tomicus piniperda L. (Curculionidae: Scolytinae) on different pines species and validation of T. destruens. Mol. Ecol. 2002, 11, 483–494. [Google Scholar]
- Gallego, D.; Cánovas, F.; Esteve, M.A.; Galián, J. Descriptive biogeography of Tomicus (Coleoptera: Scolytidae) species in Spain. J. Biogeogr. 2004, 31, 2011–2014. [Google Scholar] [CrossRef]
- Faccoli, M.; Piscedda, A.; Salvato, P.; Simonato, M.; Masutti, L.; Battisti, A. Genetic structure and phylogeography of pine shoot beetle populations (Tomicus destruens and T. piniperda, Coleoptera, Scolytidae) in Italy. Ann. For. Sci. 2005, 62, 361–368. [Google Scholar] [CrossRef]
- Horn, A.; Stauffer, C.; Lieutier, F.; Kerdelhué, C. Complex postglacial history of the temperate bark beetle Tomicus piniperda L. (Coleoptera, Scolytinae). Heredity 2009, 103, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Rener, I.; Maja, J. The six-toothed bark beetle Ips sexdentatus (Boerner, 1767) (Coleoptera: Scolytidae) in the fire site Kojnik. J. For. Sci. 2001, 2, 154–155. [Google Scholar]
- Ozcan, G.E.; Eroglu, M.; Akinci, H.A. Use of pheromone-baited traps for monitoring Ips sexdentatus (Boerner) (Coleoptera: Curculionidae) in oriental spruce stands. Afr. J. Biotechnol. 2011, 10, 16351–16360. [Google Scholar]
- Pfeffer, A. Zentral- und westpaläarktische Borken- und Kernkäfer (Coleoptera: Scolytidae, Platypodidae); Pro Entomologica, Naturhistorisches Museum: Basel, Switzerland, 1995. [Google Scholar]
- Alonso-Zarazaga, M.; Barrios, H.; Borovec, R.; Bouchard, P.; Caldara, R.; Colonnelli, E.; Gültekin, L.; Hlaváč, P.; Korotyaev, B.; Lyal, C.; et al. Cooperative Catalogue of Palaearctic Coleoptera Curculionoidea; Sociedad Entomológica Aragonesa (S.E.A.): Zaragoza, Spain, 2017. [Google Scholar]
- Paiva, M.R.; Pessoa, M.F.; Vité, J.P. Reduction in the pheromone attractant response of Orthotomicus erosus (Woll.) and Ips sexdentatus Boern. (Col., Scolytidae). J. Appl. Entom. 1988, 106, 198–200. [Google Scholar] [CrossRef]
- Lombardero, M.J.; Ayres, M.P. Factors influencing bark beetle outbreaks after forest fires on the Iberian Peninsula. Environ. Entomol. 2011, 40, 1007–1018. [Google Scholar] [CrossRef] [PubMed]
- Faccoli, M. European Bark and Ambrosia Beetles: Types, Characteristics and Identification of Mating Systems; World Biodiversity Association Handbooks 5: Verona, Italy, 2015. [Google Scholar]
- Rossi, J.P.; Samalens, J.-C.; Guyon, D.; van Halder, I.; Jactel, H.; Menassieu, P.; Piou, D. Multiscale spatial variation of the bark beetle Ips sexdentatus damage in a pine plantation forest (Landers de Gascogne, southwestern France). For. Ecol. Manag. 2009, 257, 1551–1557. [Google Scholar] [CrossRef]
- Pineau, X.; Bourguignon, M.; Jactel, H.; Lieutier, F.; Sallé, A. Pyrrhic victory for bark beetles: Successful standing tree colonization triggers strong intraspecific competition for offspring of Ips sexdentatus. For. Ecol. Manag. 2017, 399, 188–196. [Google Scholar] [CrossRef]
- Schebeck, M.; Dowle, E.J.; Schuler, H.; Avtzis, D.N.; Bertheau, C.; Feder, J.L.; Ragland, G.J.; Stauffer, C. Pleistocene climate cycling and host plant association shaped the demographic history of the bark beetle Pityogenes chalcographus. Sci. Rep. 2018, 8, 14207. [Google Scholar] [CrossRef] [PubMed]
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgind, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, N.; Tamura, K.; Aotsuka, T. PCR error and molecular population genetics. Biochem. Gen. 1999, 37, 317–321. [Google Scholar] [CrossRef]
- Nei, M. Molecular Evolutionary Genetics; Columbia University Press: New York, NY, USA, 1987. [Google Scholar]
- Tajima, F. Evolutionary relationship of DNA sequences in finite populations. Genetics 1983, 105, 437–460. [Google Scholar] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Avtzis, D.N.; Bertheau, C.; Stauffer, C. What is next in bark beetle phylogeography? Insects 2012, 3, 453–472. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Nylander, J.; Ronquist, F.; Huelsenbeck, J.; Nieves-Aldrey, J. Bayesian phylogenetic analysis of combined data. Syst. Biol. 2004, 53, 47–67. [Google Scholar] [CrossRef]
- Hasegawa, M.; Kishino, K.; Yano, T. Dating the human-ape splitting by a molecular clock of mitochondrial DNA. J. Mol. Evol. 1985, 22, 160–174. [Google Scholar] [CrossRef]
- Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 1989, 123, 585–595. [Google Scholar]
- Fu, Y.-X.; Li, W.H. Statistical test of neutrality of mutations. Genetics 1993, 133, 693–709. [Google Scholar]
- Panchal, M. The automation of Nested Clade Phylogeographic Analysis. Bioinformatics 2007, 23, 509–510. [Google Scholar] [CrossRef] [PubMed]
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1659. [Google Scholar] [CrossRef] [PubMed]
- Posada, D.; Crandall, K.A.; Templeton, A.R. GeoDis: A program for the cladistic nested analysis of the geographical distribution of genetic haplotypes. Mol. Ecol. 2000, 9, 487–488. [Google Scholar] [CrossRef]
- Dupanloup, I.; Schneider, S.; Excoffier, L. A simulated annealing approach to define the genetic structure of populations. Mol. Ecol. 2002, 11, 2571–2581. [Google Scholar] [CrossRef] [PubMed]
- Wright, S. Isolation by distance. Genetics 1943, 28, 114–138. [Google Scholar] [PubMed]
- Safner, T.; Miller, M.P.; McRae, B.H.; Fortin, M.J.; Manel, S. Comparison of Bayesian clustering and edge detection methods for inferring boundaries in landscape genetics. Int. J. Mol. Sci. 2011, 12, 865–889. [Google Scholar] [CrossRef] [PubMed]
- Mantel, N. The detection of disease clustering and a generalized regression approach. Cancer Res. 1967, 27, 209–220. [Google Scholar] [PubMed]
- Miller, M.P. Alleles in space: Computer software for the joint analysis of interindividual spatial and genetic information. J. Hered. 2005, 96, 722–724. [Google Scholar] [CrossRef]
- Vandergast, A.G.; Perry, W.M.; Lugo, R.V.; Hathaway, S.A. Genetic landscapes GIS Toolbox: Tools to map patterns of genetic divergence and diversity. Mol. Ecol. Resour. 2011, 11, 158–161. [Google Scholar] [CrossRef]
- Stauffer, C.; Lakatos, F.; Hewitt, G.M. Phylogeography and postglacial colonization routes of Ips typographus (Coleoptera, Scolytidae). Mol. Ecol. 1999, 8, 763–773. [Google Scholar] [CrossRef]
- Avtzis, D.N.; Arthofer, W.; Stauffer, C. Sympatric occurrence of diverged mtDNA lineages of Pityogenes chalcographus (Coleoptera, Scolytinae) in Europe. Biol. J. Linn. Soc. 2008, 94, 331–340. [Google Scholar] [CrossRef]
- Gómez, A.; Lunt, D.H. Refugia within Refugia: Patterns of Phylogeographic Concordance in the Iberian Peninsula. In Phylogeography of Southern European Refugia; Weiss, S., Ferrand, N., Eds.; Springer: Dordrecht, The Netherland, 2007; pp. 155–188. [Google Scholar]
- Poulakakis, N.; Kapli, P.; Lymberakis, P.; Trichas, A.; Vardinoyiannis, K.; Sfenthourakis, S.; Mylonas, M. A review of phylogeographic analyses of animal taxa from the Aegean and surrounding regions. J. Zool. Syst. Evol. Res. 2014, 53, 18–32. [Google Scholar] [CrossRef]
- Fontanella, F.M. Mitochondrial DNA panmixia in the little brown seastar Echinaster spinulosus suggests a recent population expansion. DNA Seq. 2016, 27, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Neethling, M.; Matthee, C.A.; Bowie, R.C.K.; von der Heyden, S. Evidence of panmixia despite barriers to gene flow in the southern African endemic, Caffrogobius caffer (Teleostei: Gobiidae). BMC Evol. Biol. 2008, 8, 325. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, H.; Nishikawa, M.; Sota, T. Loss of flight promotes beetle diversification. Nat. Commun. 2012, 3, 648. [Google Scholar] [CrossRef] [PubMed]
- Jactel, H. Individual variability of the flight potential of Ips sexdentatus Boern. (Coleoptera: Scolytidae) in relation to day of emergence, sex, size and lipid content. Can. Entomol. 1993, 125, 919–930. [Google Scholar] [CrossRef]
- Schmitt, T. Molecular biogeography of Europe: Pleistocene cycles and postglacial trends. Front. Zool. 2007, 4, 11. [Google Scholar] [CrossRef]
- Kelley, S.T.; Farrell, B.D.; Mitton, J.B. Effects of specialization on genetic differentiation in sister species of bark beetles. Heredity 2000, 84, 218–227. [Google Scholar] [CrossRef]
- Rassati, D.; Faccoli, M.; Marini, L.; Haack, R.A.; Battisti, A.; Toffolo, E.P. Exploring the role of wood waste landfills in early detection of non-native wood-boring beetles. J. Pest Sci. 2015, 88, 563–572. [Google Scholar] [CrossRef]
- Robinet, C.; Imbert, C.-E.; Rousselet, J.; Sauvard, D.; Garcia, J.; Goussard, F.; Roques, A. Human-mediated long-distance jumps of the pine processionary moth in Europe. Biol. Invasions 2012, 14, 1557–1569. [Google Scholar] [CrossRef]
- Lacković, N.; Pernek, M.; Bertheau, C.; Franjević, D.; Stauffer, C.; Avtzis, D.N. Limited genetic structure of gypsy moth populations reflecting a recent history in Europe. Insects 2018, 9, 143. [Google Scholar] [CrossRef] [PubMed]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Avtzis, D.N.; Lakatos, F.; Gallego, D.; Pernek, M.; Faccoli, M.; Wegensteiner, R.; Stauffer, C. Shallow Genetic Structure among the European Populations of the Six-Toothed Bark Beetle Ips sexdentatus (Coleoptera, Curculionidae, Scolytinae). Forests 2019, 10, 136. https://doi.org/10.3390/f10020136
Avtzis DN, Lakatos F, Gallego D, Pernek M, Faccoli M, Wegensteiner R, Stauffer C. Shallow Genetic Structure among the European Populations of the Six-Toothed Bark Beetle Ips sexdentatus (Coleoptera, Curculionidae, Scolytinae). Forests. 2019; 10(2):136. https://doi.org/10.3390/f10020136
Chicago/Turabian StyleAvtzis, Dimitrios N., Ferenc Lakatos, Diego Gallego, Milan Pernek, Massimo Faccoli, Rudolf Wegensteiner, and Christian Stauffer. 2019. "Shallow Genetic Structure among the European Populations of the Six-Toothed Bark Beetle Ips sexdentatus (Coleoptera, Curculionidae, Scolytinae)" Forests 10, no. 2: 136. https://doi.org/10.3390/f10020136
APA StyleAvtzis, D. N., Lakatos, F., Gallego, D., Pernek, M., Faccoli, M., Wegensteiner, R., & Stauffer, C. (2019). Shallow Genetic Structure among the European Populations of the Six-Toothed Bark Beetle Ips sexdentatus (Coleoptera, Curculionidae, Scolytinae). Forests, 10(2), 136. https://doi.org/10.3390/f10020136