Effective Molecular Identification of Ectomycorrhizal Fungi: Revisiting DNA Isolation Methods




Abstract
:1. Introduction
- Approaches to isolating DNA from ECM root tips differ significantly in general efficiency.
- The efficiency of certain DNA isolation methods depends on the properties of a specific fungal sample.
2. Materials and Methods
2.1. Materials
2.2. Cell Lysis and Lysate Purification
2.3. Isolation Assessment
2.4. Statistical Analysis
3. Results
3.1. ECM Root Tip Mass and Morphology
3.2. PCR Results and DNA Sequencing
3.3. Statistical Analysis
4. Discussion
4.1. The Efficiency of the DNA Isolation Approaches
4.2. Fungal Diversity as a Factor in DNA Isolation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Brundrett, M.C. Mycorrhizal associations and other means of nutrition of vascular plants: Understanding the global diversity of host plants by resolving conflicting information and developing reliable means of diagnosis. Plant Soil 2009, 320, 37–77. [Google Scholar] [CrossRef]
- Wang, B.; Qiu, Y.-L. Phylogenetic distribution and evolution of mycorrhizas in land plants. Mycorrhiza 2006, 16, 299–363. [Google Scholar] [CrossRef] [PubMed]
- Wurzburger, N.; Brookshire, E.N.J.; McCormack, M.L.; Lankau, R.A. Mycorrhizal fungi as drivers and modulators of terrestrial ecosystem processes. New Phytol. 2017, 213, 996–999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Read, D.J.; Perez-Moreno, J. Mycorrhizas and nutrient cycling in ecosystems—A journey towards relevance? New Phytol. 2003, 157, 475–492. [Google Scholar] [CrossRef]
- Wu, Q.S. Arbuscular Mycorrhizas and Stress Tolerance of Plants; Wu, Q.-S., Ed.; Springer: Singapore, 2017; ISBN 978-981-10-4114-3. [Google Scholar]
- Boroujeni, D.S.; Hemmatinezhad, B. Review of Application and Importance of Ectomycorrhiza Fungi and their Role in the Stability of Ecosystems. Biosci. Biotechnol. Res. Asia 2015, 12, 153–158. [Google Scholar] [CrossRef]
- Tedersoo, L.; May, T.W.; Smith, M.E. Ectomycorrhizal lifestyle in fungi: Global diversity, distribution, and evolution of phylogenetic lineages. Mycorrhiza 2010, 20, 217–263. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Mett, M.; Ishida, T.A.; Bahram, M. Phylogenetic relationships among host plants explain differences in fungal species richness and community composition in ectomycorrhizal symbiosis. New Phytol. 2013, 199, 822–831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez, C.W.; Langley, J.A.; Chapman, S.; McCormack, M.L.; Koide, R.T. The decomposition of ectomycorrhizal fungal necromass. Soil Biol. Biochem. 2016, 93, 38–49. [Google Scholar] [CrossRef] [Green Version]
- Sevag, M.G. A new physical de-proteination method for representation of biologically effective substances-isolation of carbohydrates in chicken-protein and pneumococci. Biochem. Z. 1934, 273, 419–429. [Google Scholar]
- Sevag, M.G.; Lackman, D.B.; Smolens, J. The Isolation of the Components of Streptococcal Nucleoproteins in serologically Active Form. J. Biol. Chem. 1938, 124, 425–436. [Google Scholar]
- Boom, R.; Sol, C.J.A.; Salimans, M.M.M.; Jansen, C.L.; Wertheim-Van Dillen, P.M.E.; Van Der Noordaa, J. Rapid and simple method for purification of nucleic acids. J. Clin. Microbiol. 1990, 28, 495–503. [Google Scholar] [PubMed]
- Manian, S.; Sreenivasaprasad, S.; Mills, P.R. DNA extraction method for PCR in mycorrhizal fungi. Lett. Appl. Microbiol. 2001, 33, 307–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karakousis, A.; Tan, L.; Ellis, D.; Alexiou, H.; Wormald, P.J. An assessment of the efficiency of fungal DNA extraction methods for maximizing the detection of medically important fungi using PCR. J. Microbiol. Methods 2006, 65, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Rezadoost, M.H.; Kordrostami, M.; Kumleh, H.H. An efficient protocol for isolation of inhibitor-free nucleic acids even from recalcitrant plants. 3 Biotech 2016, 6, 61. [Google Scholar] [CrossRef] [PubMed]
- Schrader, C.; Schielke, A.; Ellerbroek, L.; Johne, R. PCR inhibitors-occurrence, properties and removal. J. Appl. Microbiol. 2012, 113, 1014–1026. [Google Scholar] [CrossRef] [PubMed]
- Agerer, R. Studies on Ectomycorrhizae. II. Introducing remarks on characterization and identification. Mycotaxon 1986, 26, 473–492. [Google Scholar]
- Agerer, R. Colour Atlas of Ectomycorrhizae, 15th ed.; Agerer, R., Ed.; Einhorn-Verlag: Schwäbisch Gmünd, Germany, 2012; ISBN 3-921703-77-8. [Google Scholar]
- Dickie, I.A.; Richardson, S.J.; Wiser, S.K. Ectomycorrhizal fungal communities and soil chemistry in harvested and unharvested temperate Nothofagus rainforests. Can. J. For. Res. 2009, 39, 1069–1079. [Google Scholar] [CrossRef]
- Richard, F.; Roy, M.; Shahin, O.; Sthultz, C.; Duchemin, M.; Joffre, R.; Selosse, M.-A. Ectomycorrhizal communities in a Mediterranean forest ecosystem dominated by Quercus ilex: Seasonal dynamics and response to drought in the surface organic horizon. Ann. For. Sci. 2011, 68, 57–68. [Google Scholar] [CrossRef]
- White, T.; Bruns, T.D.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. PCR Protocols 1990, 18, 315–322. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Varma, A.; Padh, H.; Shrivastava, N. Plant genomic DNA isolation: An art or a science. Biotechnol. J. 2007, 2, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Shi, B.; Shin, Y.K.; Hassanali, A.A.; Singer, S.J. DNA Binding to the Silica Surface. J. Phys. Chem. B 2015, 119, 11030–11040. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Katzensteiner, K.; Schume, H.; Van Loo, M.; Godbold, D.L. Potassium fertilization affects the distribution of fine roots but does not change ectomycorrhizal community structure. Ann. For. Sci. 2016, 73, 691–702. [Google Scholar] [CrossRef] [Green Version]
- Bresinsky, A.; Rennschmid, A. Pigmentmerkmale, Organisationsstufen und systematische Gruppen bei Hoheren Pilzen. Ber. Dtsch. Bot. Ges. 1971, 84, 313–329. [Google Scholar]
- Vega Gutierrez, S.M.; Van Court, R.C.; Stone, D.W.; Konkler, M.J.; Groth, E.N.; Robinson, S.C. Relationship between Molarity and Color in the Crystal (‘Dramada’) Produced by Scytalidium cuboideum, in Two Solvents. Molecules 2018, 23, 2581. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Species | Family | Order | Division | |
|---|---|---|---|---|
| A | Elaphomyces muricatus | Elaphomycetaceae | Eurotiales | Ascomycota |
| B | Pachyphloeus sp. | Pezizaceae | Pezizales | Ascomycota |
| C | Genea hispidula | Pyronemataceae | Pezizales | Ascomycota |
| D | Humaria hemisphaerica | Pyronemataceae | Pezizales | Ascomycota |
| E | Tuber puberulum | Tuberaceae | Pezizales | Ascomycota |
| F | Amanita rubescens | Amanitaceae | Agaricales | Basidiomycota |
| G | Cortinarius torvus | Cortinariaceae | Agaricales | Basidiomycota |
| H | Laccaria amethystina | Hydnangiaceae | Agaricales | Basidiomycota |
| I | Piloderma sphaerosporum | Atheliaceae | Atheliales | Basidiomycota |
| J | Imleria badia | Boletaceae | Boletales | Basidiomycota |
| K | Xerocomellus cisalpinus | Boletaceae | Boletales | Basidiomycota |
| L | Paxillus involutus | Paxillaceae | Boletales | Basidiomycota |
| M | Suillus variegatus | Suillaceae | Boletales | Basidiomycota |
| N | Craterellus cornucopioides | Cantharellaceae | Cantharellales | Basidiomycota |
| O | Clavulina coralloides | Clavulinaceae | Cantharellales | Basidiomycota |
| P | Lactarius aurantiacus | Russulaceae | Russulales | Basidiomycota |
| Q | Russula nigricans | Russulaceae | Russulales | Basidiomycota |
| R | Tomentella sp.1 | Thelephoraceae | Thelephorales | Basidiomycota |
| S | Tomentella sp.2 | Thelephoraceae | Thelephorales | Basidiomycota |
| T | Tomentella terrestris | Thelephoraceae | Thelephorales | Basidiomycota |
| Species | Mass ± SD (g × 10−5) | Color | ||||
|---|---|---|---|---|---|---|
| Hue (°) | Saturation (%) | Value (%) | ||||
| A | Elaphomyces muricatus | 0.833 ± | 0.111 f * | 30 | 60 | 80 |
| B | Pachyphloeus sp. | 1.333 ± | 0.043 ef | 30 | 55 | 75 |
| C | Genea hispidula | 1.417 ± | 0.283 e | 30 | 65 | 65 |
| D | Humaria hemisphaerica | 1.944 ± | 0.053 de | 25 | 70 | 65 |
| E | Tuber puberulum | 1.028 ± | 0.086 f | 45 | 70 | 95 |
| F | Amanita rubescens | 2.056 ± | 0.411 de | 15 | 40 | 75 |
| G | Cortinarius torvus | 0.861 ± | 0.084 f | 40 | 40 | 80 |
| H | Laccaria amethystina | 1.694 ± | 0.261 de | 25 | 40 | 80 |
| I | Piloderma sphaerosporum | 3.472 ± | 1.157 c | 20 | 70 | 85 |
| J | Imleria badia | 4.500 ± | 0.811 b | 30 | 35 | 85 |
| K | Xerocomellus cisalpinus | 2.806 ± | 0.401 d | 30 | 35 | 80 |
| L | Paxillus involutus | 0.972 ± | 0.210 ef | 25 | 50 | 75 |
| M | Suillus variegatus | 9.556 ± | 2.415 a | 20 | 55 | 70 |
| N | Craterellus cornucopioides | 2.556 ± | 0.333 de | 40 | 40 | 85 |
| O | Clavulina coralloides | 1.389 ± | 0.231 e | 30 | 15 | 60 |
| P | Lactarius aurantiacus | 1.500 ± | 0.134 de | 45 | 75 | 95 |
| Q | Russula nigricans | 2.667 ± | 0.064 de | 25 | 50 | 75 |
| R | Tomentella sp.1 | 2.528 ± | 0.139 de | 0 | 25 | 45 |
| S | Tomentella sp.2 | 3.528 ± | 0.044 c | 0 | 5 | 30 |
| T | Tomentella terrestris | 1.444 ± | 0.076 e | 20 | 40 | 70 |
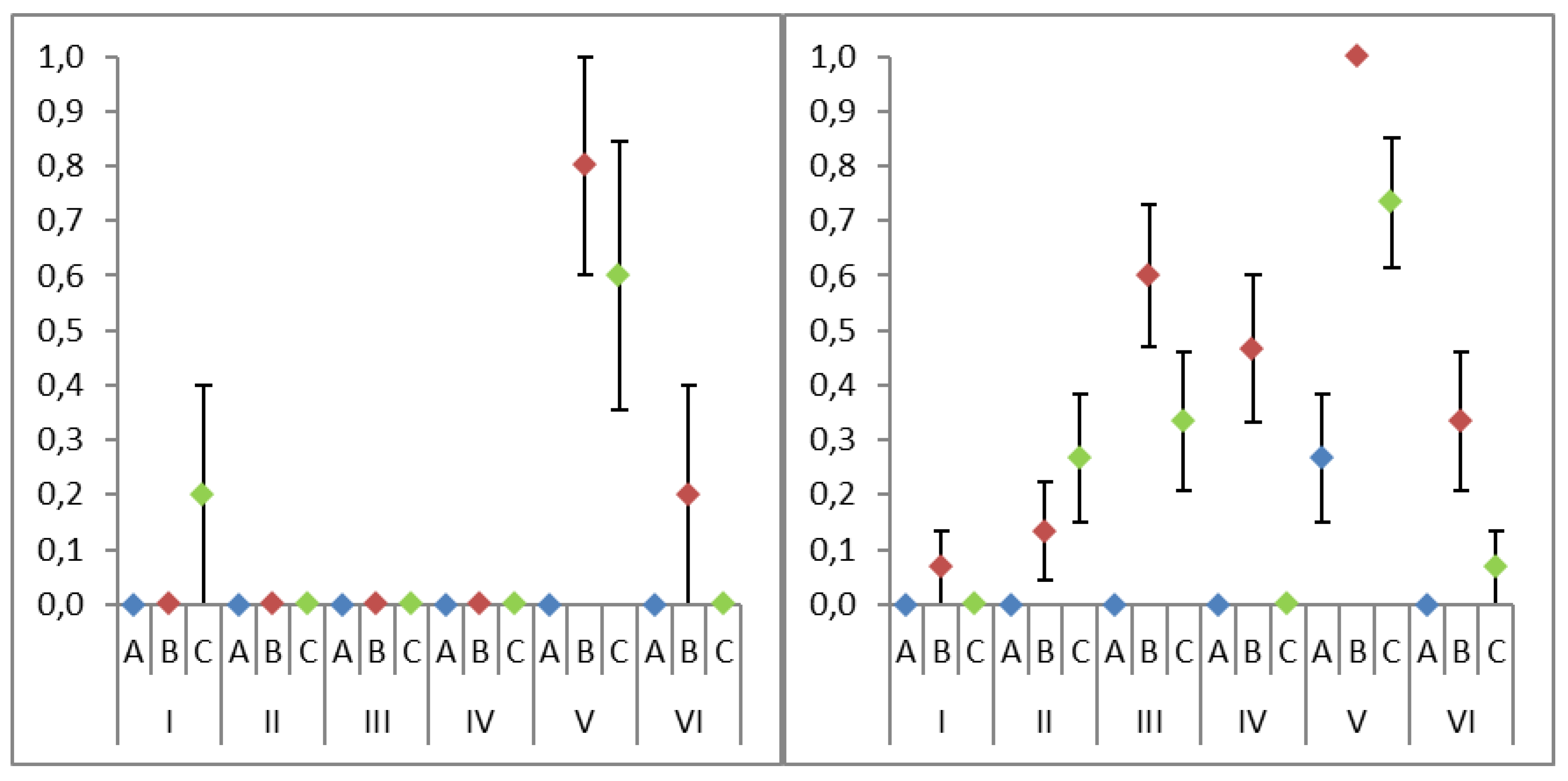
| Lysis | Purification | Number of Successful Ascomycota Amplifications | Number of Successful Basidiomycota Amplifications | Total Number of Successful Amplifications |
|---|---|---|---|---|
| I | A | 0 (0%) | 0 (0%) | 0 (0%) |
| B | 0 (0%) | 1 (7%) | 1 (5%) | |
| C | 1 (20%) | 0 (0%) | 1 (5%) | |
| II | A | 0 (0%) | 0 (0%) | 0 (0%) |
| B | 0 (0%) | 2 (13%) | 2 (10%) | |
| C | 0 (0%) | 4 (27%) | 4 (20%) | |
| III | A | 0 (0%) | 0 (0%) | 0 (0%) |
| B | 0 (0%) | 9 (60%) | 9 (45%) | |
| C | 0 (0%) | 5 (33%) | 5 (25%) | |
| IV | A | 0 (0%) | 0 (0%) | 0 (0%) |
| B | 0 (0%) * | 7 (47%) | 7 (35%) * | |
| C | 0 (0%) | 0 (0%) | 0 (0%) | |
| V | A | 0 (0%) | 4 (27%) | 4 (20%) |
| B | 4 (80%) * | 15 (100%) | 19 (95%) * | |
| C | 3 (60%) * | 11 (73%) * | 14 (70%) * | |
| VI | A | 0 (0%) | 0 (0%) | 0 (0%) |
| B | 1 (20%) | 5 (33%) | 6 (30%) | |
| C | 0 (0%) | 1 (7%) | 1 (5%) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Janowski, D.; Wilgan, R.; Leski, T.; Karliński, L.; Rudawska, M. Effective Molecular Identification of Ectomycorrhizal Fungi: Revisiting DNA Isolation Methods. Forests 2019, 10, 218. https://doi.org/10.3390/f10030218
Janowski D, Wilgan R, Leski T, Karliński L, Rudawska M. Effective Molecular Identification of Ectomycorrhizal Fungi: Revisiting DNA Isolation Methods. Forests. 2019; 10(3):218. https://doi.org/10.3390/f10030218
Chicago/Turabian StyleJanowski, Daniel, Robin Wilgan, Tomasz Leski, Leszek Karliński, and Maria Rudawska. 2019. "Effective Molecular Identification of Ectomycorrhizal Fungi: Revisiting DNA Isolation Methods" Forests 10, no. 3: 218. https://doi.org/10.3390/f10030218
APA StyleJanowski, D., Wilgan, R., Leski, T., Karliński, L., & Rudawska, M. (2019). Effective Molecular Identification of Ectomycorrhizal Fungi: Revisiting DNA Isolation Methods. Forests, 10(3), 218. https://doi.org/10.3390/f10030218