Climatic Change Can Influence Species Diversity Patterns and Potential Habitats of Salicaceae Plants in China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Spatial Data
2.2. Environmental Parameters
2.2.1. Current Environment Variables
2.2.2. Historical and Future Climate Scenarios
2.3. Building the Species Distribution Model (SDM)
2.4. Biodiversity Pattern Indices
2.5. Changes in Core Distribution Centers
2.6. Statistical Analysis
3. Results
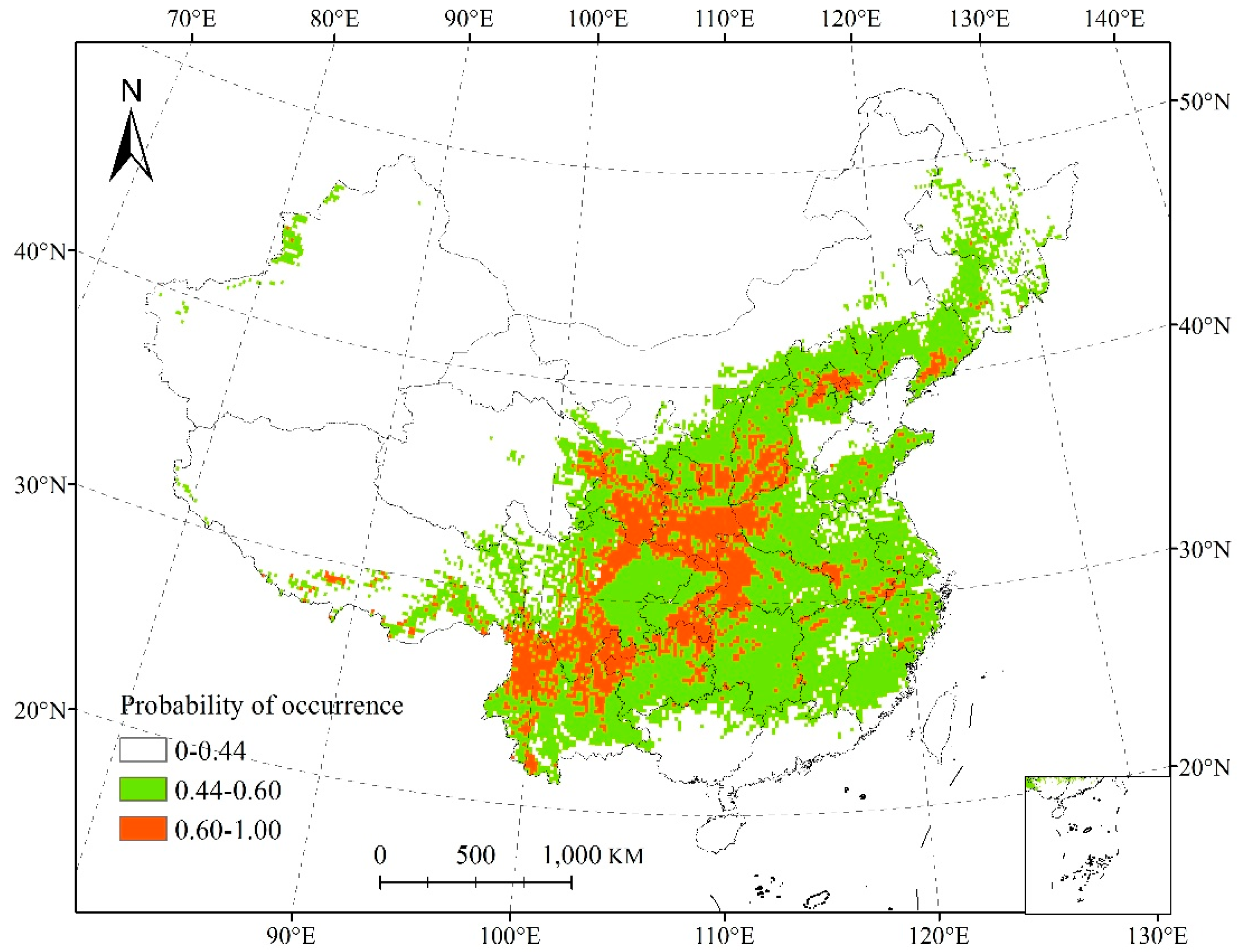
3.1. Species Distribution Model and Its Accuracy
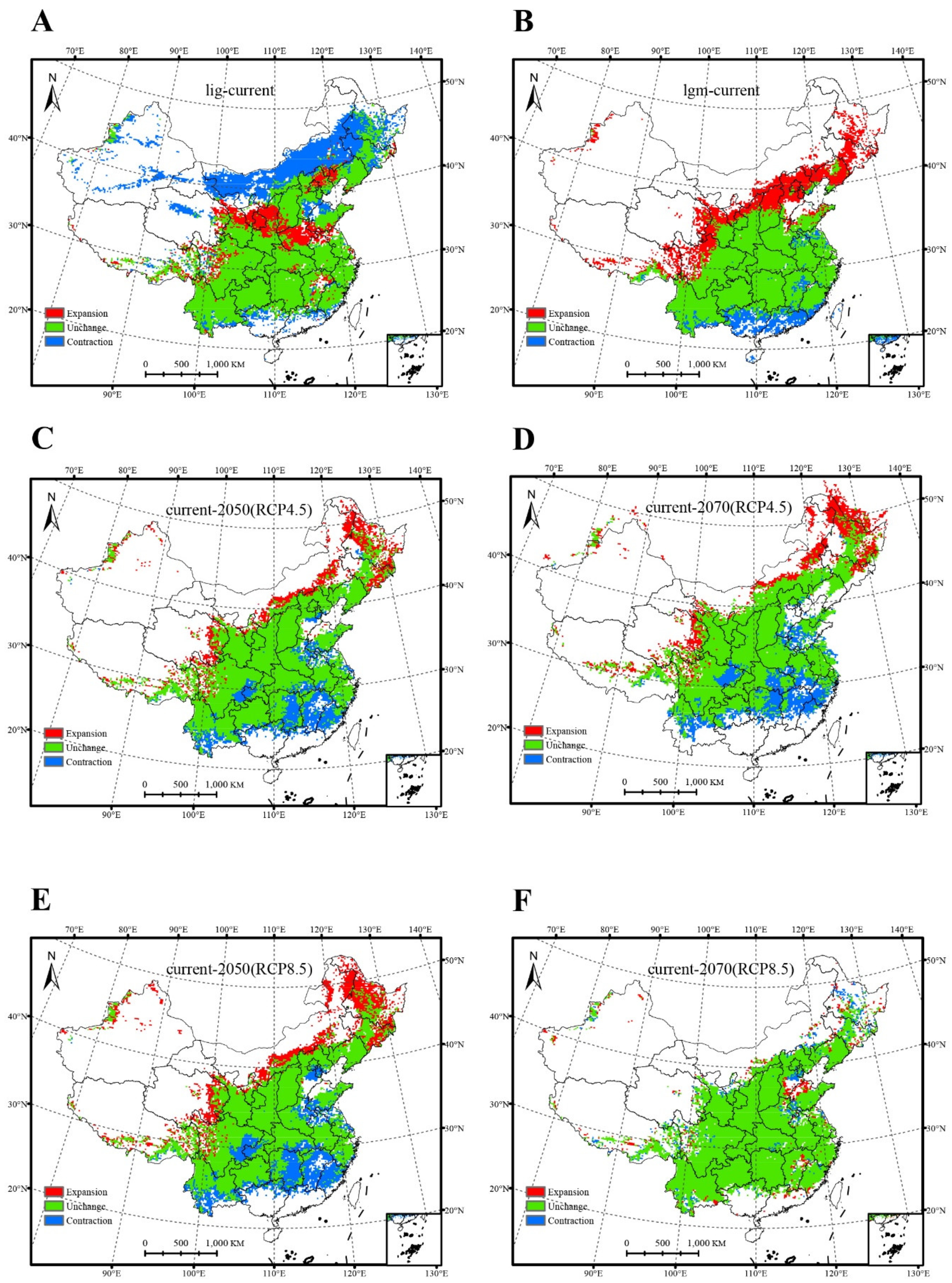
3.2. Changes in the Potential Range of Salicaceae
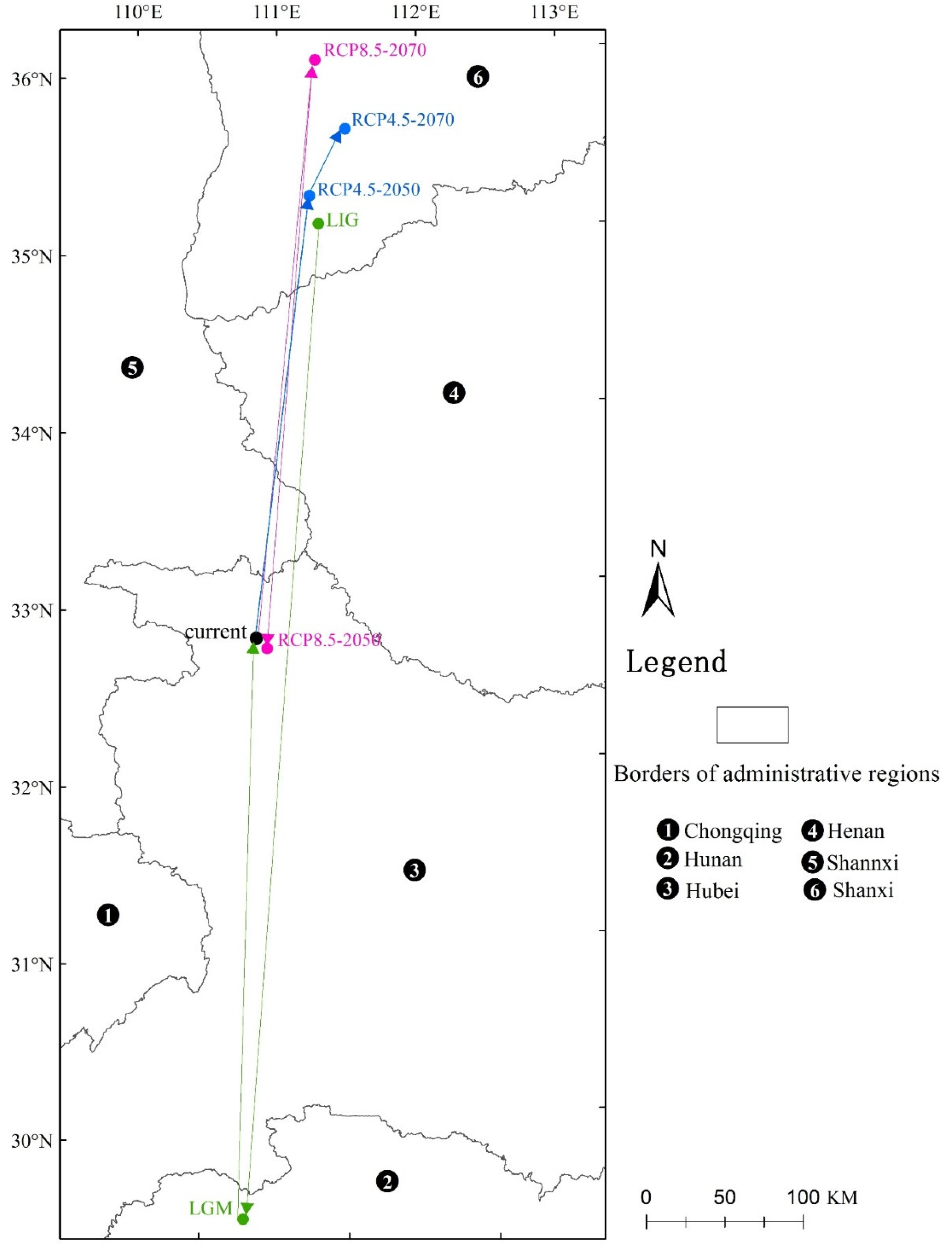
3.3. Core Distributional Shifts
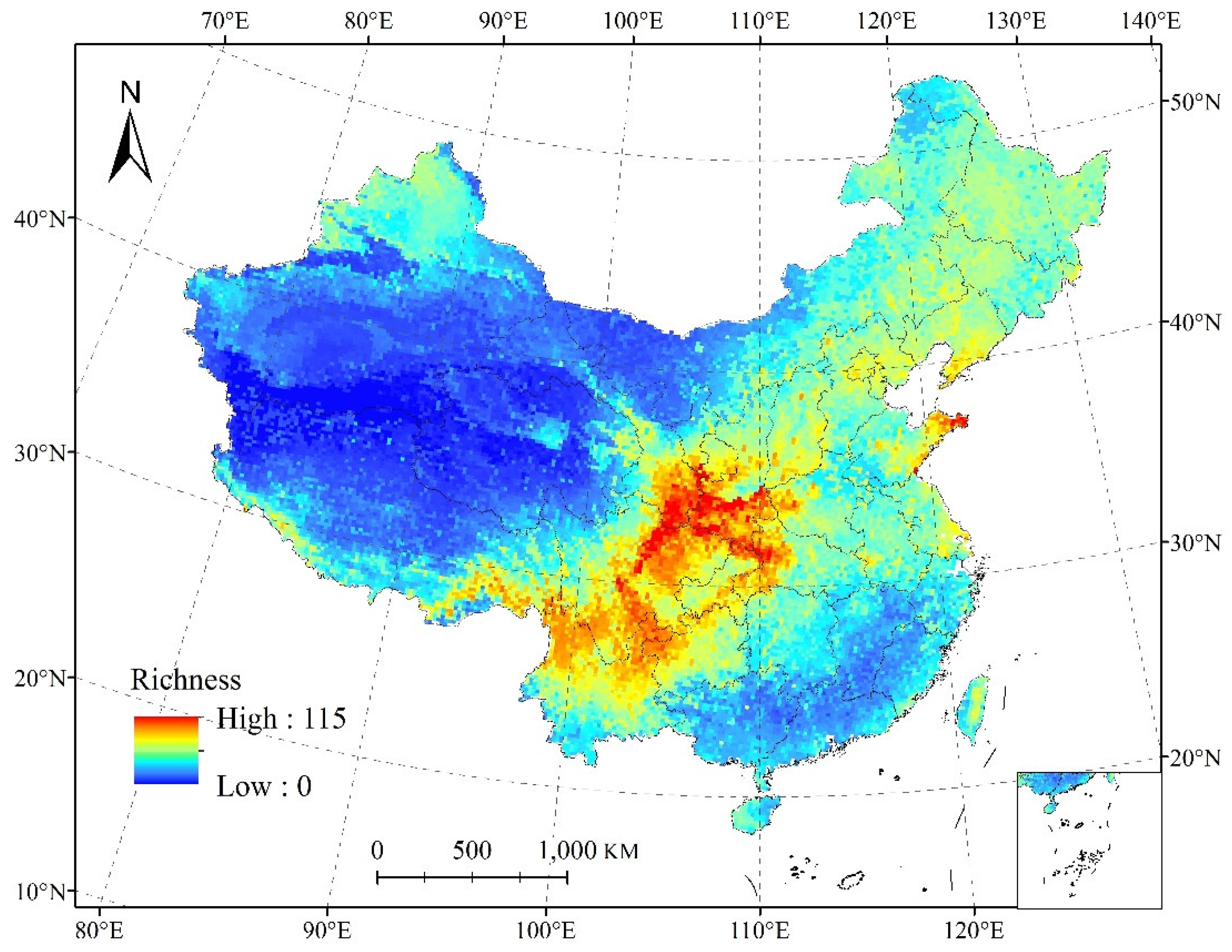
3.4. Relationships between Species Richness and Environmental Parameters
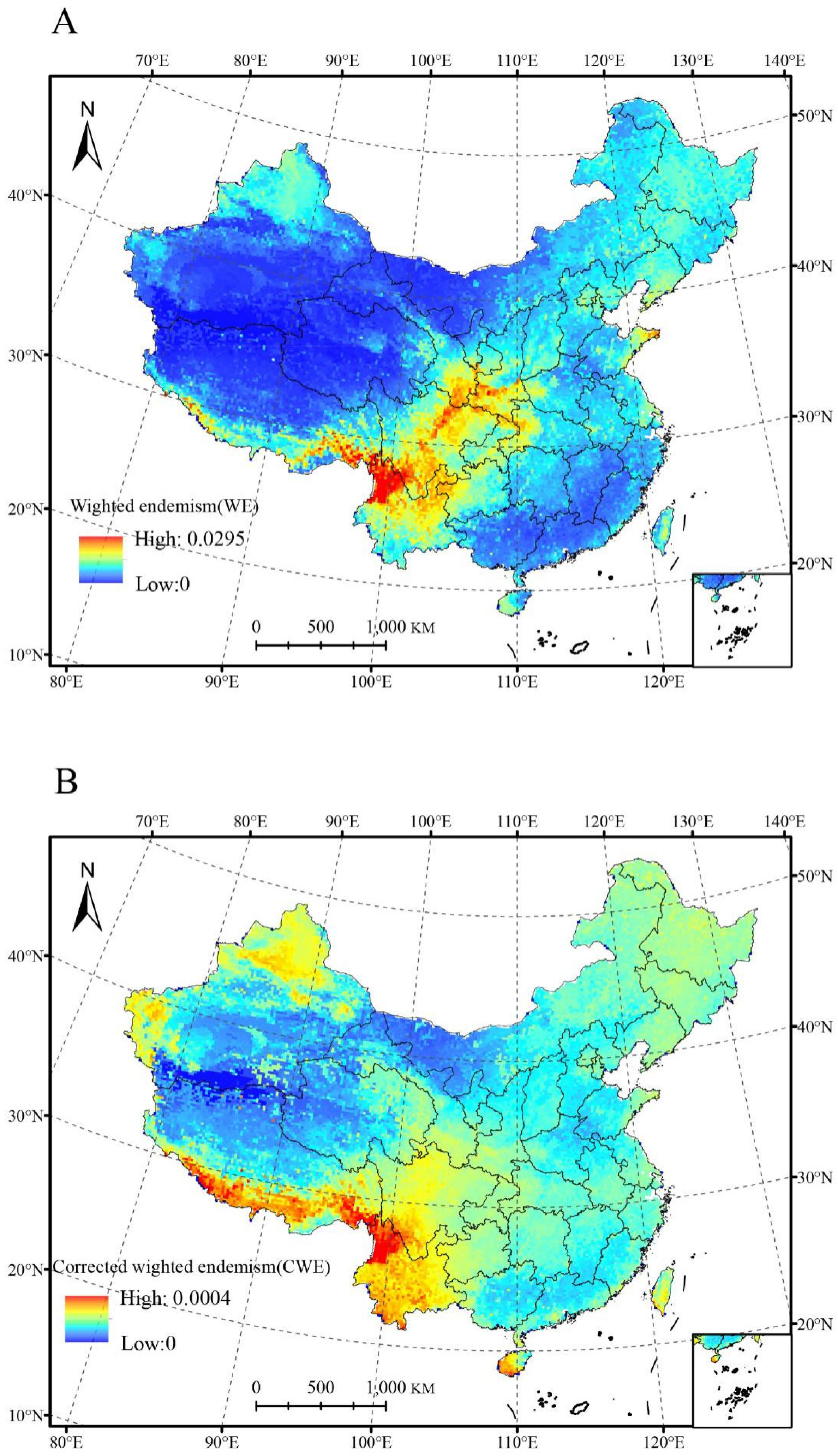
3.5. Relationships between Weighted Endemism (WE), Corrected-Weighted Endemism (CWE), and Environmental Factors
4. Discussion
4.1. Changes in the Potential Range of Salicaceae in China
4.2. Species Richness and Endemism Patterns of Salicaceae in China
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hong, Q.; Ricklefs, R.E. Large–scale processes and the asian bias in species diversity of temperate plants. Nature 2000, 407, 180–182. [Google Scholar]
- Buckley, L.B.; Davies, T.J.; Ackerly, D.D.; Kraft, N.J.; Harrison, S.P.; Anacker, B.L.; Cornell, H.V.; Damschen, E.I.; Grytnes, J.A.; Hawkins, B.A.; et al. Phylogeny, niche conservatism and the latitudinal diversity gradient in mammals. Proc. R. Soc. 2010, 277, 2131–2138. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.H. Why are there so many species in the tropics? J. Biogeogr. 2014, 41, 8–22. [Google Scholar] [CrossRef] [PubMed]
- Beck, J.; Ballesteros-Mejia, L.; Buchmann, C.M.; Dengler, J.; Fritz, S.A.; Gruber, B.; Hof, C.; Jansen, F.; Knapp, S.; Kreft, H.; et al. What’s on the horizon for macroecology? Ecography 2012, 35, 673–683. [Google Scholar] [CrossRef]
- Hawkins, B.A. Invited views in basic and applied ecology: Are we making progress toward understanding the global diversity gradient? Basic Appl. Ecol. 2004, 5, 1–3. [Google Scholar] [CrossRef]
- Brown, J.H.; Gillooly, J.F.; Allen, A.P.; Savage, V.M.; West, G.B. Toward a metabolic theory of ecology. Ecology 2004, 85, 1771–1789. [Google Scholar] [CrossRef]
- Colwell, R.K.; Rahbek, C.; Gotelli, N.J. The mid-domain effect and species richness patterns: What have we learned so far? Am. Nat. 2004, 163, E1–E23. [Google Scholar] [CrossRef] [PubMed]
- Joy, J.B. The global diversity of birds in space and time. Nature 2012, 491, 444–448. [Google Scholar]
- Jansson, R. Global variation in diversification rates of flowering plants: Energy vs. Climate change. Ecol. Lett. 2008, 11, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Mittelbach, G.G.; Schemske, D.W.; Cornell, H.V.; Allen, A.P.; Brown, J.M.; Bush, M.B.; Harrison, S.P.; Hurlbert, A.H.; Knowlton, N.; Lessios, H.A.; et al. Evolution and the latitudinal diversity gradient: Speciation, extinction and biogeography. Ecol. Lett. 2007, 10, 315–331. [Google Scholar] [CrossRef] [PubMed]
- Levsen, N.; Tiffin, P.; Olson, M. Pleistocene speciation in the genus populus (salicaceae). Syst. Biol. 2012, 61, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.B.; Chun, J.H. Retracted article: Habitat heterogeneity and climate explain plant diversity patterns along an extensive environmental gradient in the temperate forests of south korea. Folia Geobot. 2016, 1. [Google Scholar] [CrossRef]
- Veloz, S.D.; Williams, J.W.; Blois, J.L.; He, F.; Otto-Bliesner, B.; Liu, Z. No–analog climates and shifting realized niches during the late quaternary: Implications for 21st–century predictions by species distribution models. Glob. Chang. Biol. 2012, 18, 1698–1713. [Google Scholar] [CrossRef]
- Currie, D.J. Energy and large-scale patterns of animal and plant–species richness. Am. Nat. 1991, 137, 27–49. [Google Scholar] [CrossRef]
- Latham, R.E.; Ricklefs, R.E. Global patterns of tree species richness in moist forests: Energy-diversity theory does not account for variation in species richness. Oikos 1993, 67, 325–333. [Google Scholar] [CrossRef]
- Xu, X.; Zhao, H.; Zhang, X.; Hänninen, H.; Korpelainen, H.; Li, C. Different growth sensitivity to enhanced uv-b radiation between male and female populus cathayana. Tree Physiol. 2010, 30, 1489–1498. [Google Scholar] [CrossRef] [PubMed]
- Clarke, A.; Gaston, K.J. Climate, energy and diversity. Proc. R. Soc. 2006, 273, 2257–2266. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Dong, G.; Zhang, Y.; Zhang, F.; Wang, M. Patterns of species and phylogenetic diversity of pinus tabuliformis forests in the eastern loess plateau, china. For. Ecol. Manag. 2017, 394, 42–51. [Google Scholar] [CrossRef]
- Thuiller, W.; Lavergne, S.; Roquet, C.; Boulangeat, I.; Lafourcade, B.; Araujo, M.B. Consequences of climate change on the tree of life in europe. Nature 2011, 470, 531–534. [Google Scholar] [CrossRef] [PubMed]
- Pio, D.V.; Engler, R.; Linder, H.P.; Monadjem, A.; Cotterill, F.P.D.; Taylor, P.J.; Schoeman, M.C.; Price, B.W.; Villet, M.H.; Eick, G.; et al. Climate change effects on animal and plant phylogenetic diversity in southern africa. Glob. Chang. Biol. 2014, 20, 1538–1549. [Google Scholar] [CrossRef]
- Hultine, K.R.; Burtch, K.G.; Ehleringer, J.R. Gender specific patterns of carbon uptake and water use in a dominant riparian tree species exposed to a warming climate. Glob. Chang. Biol. 2013, 19, 3390–3405. [Google Scholar] [CrossRef] [PubMed]
- González-Orozco, C.E.; Pollock, L.J.; Thornhill, A.H.; Mishler, B.D.; Knerr, N.; Laffan, S.W.; Miller, J.T.; Rosauer, D.F.; Faith, D.P.; Nipperess, D.A.; et al. Phylogenetic approaches reveal biodiversity threats under climate change. Nat. Clim. Chang. 2016, 6, 1110–1114. [Google Scholar] [CrossRef]
- Bellard, C.; Bertelsmeier, C.; Leadley, P.; Thuiller, W.; Courchamp, F. Impacts of climate change on the future of biodiversity. Ecol. Lett. 2012, 15, 365–377. [Google Scholar] [CrossRef] [PubMed]
- Stocker, T.F.; Qin, D.; Plattner, G.K.; Tignor, M.; Allen, S.K.; Boschung, J.; Nauels, A.; Xia, Y.; Bex, B.; Midgley, B.M. IPCC, 2013: Climate change 2013: The physical science basis. Contribution of working group i to the fifth assessment report of the intergovernmental panel on climate change. Comput. Geom. 2013, 18, 95–123. [Google Scholar]
- Parmesan, C. Ecological and evolutionary responses to recent climate change. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 637–669. [Google Scholar] [CrossRef]
- Myers-Smith, I.H.; Forbes, B.C.; Wilmking, M.; Hallinger, M.; Lantz, T.; Blok, D.; Tape, K.D.; Macias-Fauria, M.; Sass-Klaassen, U.; Lévesque, E.; et al. Shrub expansion in tundra ecosystems: Dynamics, impacts and research priorities. Environ. Res. Lett. 2011, 6, 045509. [Google Scholar] [CrossRef]
- Jones, M.H. Sex- and habitat-specific responses of a high arctic willow, salix arctica, to experimental climate change. Oikos 1999, 87, 129–138. [Google Scholar] [CrossRef]
- Fang, Z.F.; Zhao, S.D.; Skvortsov, A.K. Salicaceae. Flora China 1999, 4, 139–274. [Google Scholar]
- Zhao, S.D. Distribution of willows (salix) in china. Acta Phytotaxon. Sin. 1987, 25, 114–124. [Google Scholar]
- Karp, A.; Shield, I. Bioenergy from plants and the sustainable yield challenge. New Phytol. 2008, 179, 15–32. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.H.; Sun, H.; Wen, J.; Yang, Y.P. Molecular phylogeny of salix l. (salicaceae) inferred from three chloroplast datasets and its systematic implications. Taxon 2010, 59, 29–37. [Google Scholar] [CrossRef]
- Wang, Q.; Su, X.; Shrestha, N.; Liu, Y.; Wang, S.; Xu, X.; Wang, Z. Historical factors shaped species diversity and composition of salix in eastern asia. Sci. Rep. 2017, 7, 42038. [Google Scholar] [CrossRef] [PubMed]
- Warren, D.L. In defense of ‘niche modeling’. Trends Ecol. Evol. 2012, 27, 497–500. [Google Scholar] [CrossRef] [PubMed]
- Peterson, A.T.; Soberón, J.; Pearson, R.G.; Anderson, R.P.; Martínez-Meyer, E.; Nakamura, M.; Araújo, M.B. Ecological niches and geographic distribution. Monogr. Popul. Biol. 2011, 49, 328. [Google Scholar]
- Chen, F.T. Phylogeography of Rehmannia (Scrophulariaceae); Northwest University: Xi’an, China, 2015. [Google Scholar]
- Wang, Q.; Wei, Y.K.; Huang, Y.B. Research on distribution pattern of subg. Salvia benth. (lamiaceae), an important group of medicinal plants in east asia. Acta Ecol. Sin. 2015, 5, 470–479. [Google Scholar]
- Beck, J.; Böller, M.; Erhardt, A.; Schwanghart, W. Spatial bias in the gbif database and its effect on modeling species’ geographic distributions. Ecol. Inform. 2014, 19, 10–15. [Google Scholar] [CrossRef]
- Fourcade, Y.; Engler, J.O.; Rödder, D.; Secondi, J. Mapping species distributions with maxent using a geographically biased sample of presence data: A performance assessment of methods for correcting sampling bias. PLoS ONE 2014, 9, e97122. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.G.; Slik, J.W.; Ma, K.P. Using species distribution modeling to delineate the botanical richness patterns and phytogeographical regions of china. Sci. Rep. 2016, 6, 22400. [Google Scholar] [CrossRef] [PubMed]
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Townsend Peterson, A. Original article: Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in madagascar. J. Biogeogr. 2007, 34, 102–117. [Google Scholar] [CrossRef]
- Stevens, G.C. The latitudinal gradient in geographical range: How so many species coexist in the tropics. Am. Nat. 1989, 133, 240–256. [Google Scholar] [CrossRef]
- Yan, Y.; Yang, X.; Tang, Z. Patterns of species diversity and phylogenetic structure of vascular plants on the qinghai-tibetan plateau. Ecol. Evol. 2013, 3, 4584–4595. [Google Scholar] [CrossRef] [PubMed]
- Nybakken, L.; Hörkkä, R.; Julkunen-Tiitto, R. Combined enhancements of temperature and uvb influence growth and phenolics in clones of the sexually dimorphic salix myrsinifolia. Physiol. Plant. 2012, 145, 551–564. [Google Scholar] [CrossRef] [PubMed]
- Randriamanana, T.R.; Nissinen, K.; Moilanen, J.; Nybakken, L.; Julkunen-Tiitto, R. Long-term uv-b and temperature enhancements suggest that females of salix myrsinifolia plants are more tolerant to uv-b than males. Environ. Exp. Bot. 2015, 109, 296–305. [Google Scholar] [CrossRef]
- Feng, L.; Hao, J.; Zhang, Y.; Sheng, Z. Sexual differences in defensive and protective mechanisms of populus cathayana exposed to high uv-b radiation and low soil nutrient status. Physiol. Plant. 2014, 151, 434–445. [Google Scholar] [CrossRef]
- Hageer, Y.; Esperónrodríguez, M.; Baumgartner, J.B.; Beaumont, L.J. Climate, soil or both? Which variables are better predictors of the distributions of australian shrub species? PeerJ 2017, 5, e3446. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Dong, T.; Duan, B.; Korpelainen, H.; Niinemets, Ü.; Li, C. Sexual competition and n supply interactively affect the dimorphism and competiveness of opposite sexes in populus cathayana. Plant Cell Environ. 2015, 38, 1285–1298. [Google Scholar] [CrossRef] [PubMed]
- Moor, H.; Hylander, K.; Norberg, J. Predicting climate change effects on wetland ecosystem services using species distribution modeling and plant functional traits. Ambio 2015, 44 (Suppl. 1), S113–S126. [Google Scholar] [CrossRef]
- Fitzpatrick, M.C.; Gotelli, N.J.; Ellison, A.M. Maxent versus maxlike: Empirical comparisons with ant species distributions. Ecosphere 2013, 4, art55. [Google Scholar] [CrossRef]
- Guisan, A.; Thuiller, W. Predicting species distribution: Offering more than simple habitat models. Ecol. Lett. 2005, 8, 993–1009. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Bertrand, R.; Perez, V.; Gégout, J.C. Disregarding the edaphic dimension in species distribution models leads to the omission of crucial spatial information under climate change: The case of quercus pubescensin france. Glob. Chang. Biol. 2012, 18, 2648–2660. [Google Scholar] [CrossRef]
- Araújo, M.B.; Peterson, A.T. Uses and misuses of bioclimatic envelope modeling. Ecology 2012, 93, 1527–1539. [Google Scholar] [CrossRef] [PubMed]
- Raes, N.; Steege, H.T. A null-model for significance testing of presence-only species distribution models. Ecography 2007, 30, 727–736. [Google Scholar] [CrossRef]
- Radosavljevic, A.; Anderson, R.P.; Araujo, M. Making better maxent models of species distributions: Complexity, overfitting and evaluation. J. Biogeogr. 2014, 41, 629–643. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of maxent for ecologists. Divers. Distrib. 2010, 17, 43–57. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.; Silander, J. A practical guide to maxent for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Jiménez-Valverde, A.; Lobo, J.M. Threshold criteria for conversion of probability of species presence to either–or presence–absence. Acta Oecol. 2007, 31, 361–369. [Google Scholar] [CrossRef]
- Liu, C.; Berry, P.M.; Dawson, T.P.; Pearson, R.G. Selecting thresholds of occurrence in the prediction of species distributions. Ecography 2005, 28, 385–393. [Google Scholar] [CrossRef]
- Crisp, M.D.; Laffan, S.; Linder, H.P.; Monro, A. Endemism in the australian flora. J. Biogeogr. 2001, 28, 183–198. [Google Scholar] [CrossRef]
- Brown, J.L.; Anderson, B. Sdmtoolbox: A python–based gis toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods Ecol. Evol. 2014, 5, 694–700. [Google Scholar] [CrossRef]
- Brown, J.L.; Bennett, J.R.; French, C.M. Sdmtoolbox 2.0: The next generation python-based gis toolkit for landscape genetic, biogeographic and species distribution model analyses. PeerJ 2017, 5, e4095. [Google Scholar] [CrossRef] [PubMed]
- Diniz-Filho, J.A.F.; Bini, L.M.; Hawkins, B.A. Spatial autocorrelation and red herrings in geographical ecology. Glob. Ecol. Biogeogr. 2003, 12, 53–64. [Google Scholar] [CrossRef]
- Dutilleul, P.; Clifford, P.; Richardson, S.; Hemon, D. Modifying the t test for assessing the correlation between two spatial processes. Biometrics 1993, 49, 305–314. [Google Scholar] [CrossRef]
- Clifford, P.; Richardson, S.; Hémon, D. Assessing the significance of the correlation between two spatial processes. Biometrics 1989, 45, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Karrenberg, S.; Edwards, P.J.; Kollmann, J. The life history of salicaceae living in the active zone of floodplains. Freshw. Biol. 2002, 47, 733–748. [Google Scholar] [CrossRef]
- Chao, N.; Liu, J. On the classification and distribution of the family salicaceae. J. Sichuan For. Sci. Technol. 1998, 9, 10–20. [Google Scholar]
- Jiang, D.; Wang, H.; Drange, H.; Lang, X. Last glacial maximum over china: Sensitivities of climate to paleovegetation and tibetan ice sheet. J. Geophys. Res. 2003, 108, 4102. [Google Scholar] [CrossRef]
- Fan, L.; Zheng, H.; Milne, R.I.; Zhang, L.; Mao, K. Strong population bottleneck and repeated demographic expansions of populus adenopoda (salicaceae) in subtropical china. Ann. Bot. 2018, 121, 665–679. [Google Scholar] [CrossRef] [PubMed]
- Barnosky, A.D.; Matzke, N.; Tomiya, S.; Wogan, G.O.; Swartz, B.; Quental, T.B.; Marshall, C.; Mcguire, J.L.; Lindsey, E.L.; Maguire, K.C. Has the earth’s sixth mass extinction already arrived? Nature 2011, 471, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Nyman, T.; Wang, D.C.; Argus, G.W.; Yang, Y.P.; Chen, J.H. Phylogeny of salix subgenus salix s.L. (salicaceae): Delimitation, biogeography, and reticulate evolution. BMC Evol. Biol. 2015, 15, 31. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.X.; Fu, C.X.; Comes, H.P. Plant molecular phylogeography in china and adjacent regions: Tracing the genetic imprints of quaternary climate and environmental change in the world’s most diverse temperate flora. Mol. Phylogenet. Evol. 2011, 59, 225–244. [Google Scholar] [CrossRef] [PubMed]
- Patricola, C.M.; Chang, P.; Saravanan, R. Impact of atlantic sst and high frequency atmospheric variability on the 1993 and 2008 midwest floods: Regional climate model simulations of extreme climate events. Clim. Chang. 2013, 129, 397–411. [Google Scholar] [CrossRef]
- Kodra, E.; Steinhaeuser, K.; Ganguly, A.R. Persisting cold extremes under 21st-century warming scenarios. Geophys. Res. Lett. 2011, 38, 16. [Google Scholar] [CrossRef]
- Planton, S.; Déqué, M.; Chauvin, F.; Terray, L. Expected impacts of climate change on extreme climate events. C. R. Geosci. 2008, 340, 564–574. [Google Scholar] [CrossRef]
- Khanum, R.; Mumtaz, A.S.; Kumar, S. Predicting impacts of climate change on medicinal asclepiads of pakistan using maxent modeling. Acta Oecol. 2013, 49, 23–31. [Google Scholar] [CrossRef]
- Leng, W.; He, H.S.; Liu, H. Response of larch species to climate changes. Plant Ecol. 2008, 1, 203–205. [Google Scholar] [CrossRef]
- Ying, L.; Liu, Y.; Chen, S.; Shen, Z.; Ecology, D.O. Simulation of the potential range of pistacia weinmannifolia in southwest china with climate change based on the maximum-entropy(maxent) model. Biodivers. Sci. 2016, 24, 453–461. [Google Scholar] [CrossRef]
- Guo, Y.L.; Wei, H.Y.; Lu, C.Y.; Zhang, H.L.; Gu, W. Predictions of potential geographical distribution of sinopodophyllum hexandrum under climate change. Chin. J. Plant Ecol. 2014, 38, 249–261. [Google Scholar]
- Cheaib, A.; Badeau, V.; Boe, J.; Chuine, I.; Delire, C.; Dufrene, E.; Francois, C.; Gritti, E.S.; Legay, M.; Page, C.; et al. Climate change impacts on tree ranges: Model intercomparison facilitates understanding and quantification of uncertainty. Ecol. Lett. 2012, 15, 533–544. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Yan, H. A study of the impacts of climate change on the geographic distribution of pinus koraiensis in china. Environ. Int. 2001, 27, 201–205. [Google Scholar] [CrossRef]
- Argus, G.W. The genus salix (salicaceae) in the southeastern united states. Syst. Bot. Monogr. 1986, 9, 1–170. [Google Scholar] [CrossRef]
- Bertrand, R.; Lenoir, J.; Piedallu, C.; Riofrio-Dillon, G.; de Ruffray, P.; Vidal, C.; Pierrat, J.C.; Gegout, J.C. Changes in plant community composition lag behind climate warming in lowland forests. Nature 2011, 479, 517–520. [Google Scholar] [CrossRef] [PubMed]
- Frei, E.; Bodin, J.; Walther, G.R. Plant species’ range shifts in mountainous areas—All uphill from here? Bot. Helv. 2010, 120, 117–128. [Google Scholar] [CrossRef]
- Walther, G.R.; Beißner, S.; Burga, C.A. Trends in the upward shift of alpine plants. J. Veg. Sci. 2005, 16, 541–548. [Google Scholar] [CrossRef]
- Bai, Y.; Wei, X.; Li, X. Distributional dynamics of a vulnerable species in response to past and future climate change: A window for conservation prospects. PeerJ 2018, 6, e4287. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Liu, C.; Wan, J.; Zhang, Z. Climate change may threaten habitat suitability of threatened plant species within chinese nature reserves. PeerJ 2016, 4, e2091. [Google Scholar] [CrossRef] [PubMed]
- Puga, N.D.; Corral, J.A.R.; Eguiarte, D.R.G.; Munguia, S.M.; Rosas, G.O.D. Climate change and its impact on environmental aptitude and geographical distribution of salvia hispanica l. In mexico. Interciencia 2016, 41, 407–413. [Google Scholar]
- Hu, X.G.; Jin, Y.; Wang, X.R.; Mao, J.F.; Li, Y. Predicting impacts of future climate change on the distribution of the widespread conifer platycladus orientalis. PLoS ONE 2015, 10, e0132326. [Google Scholar] [CrossRef] [PubMed]
- Garcia, K.; Lasco, R.; Ines, A.; Lyon, B.; Pulhin, F. Predicting geographic distribution and habitat suitability due to climate change of selected threatened forest tree species in the philippines. Appl. Geogr. 2013, 44, 12–22. [Google Scholar] [CrossRef]
- Bomhard, B.; Richardson, D.M.; Donaldson, J.S.; Hughes, G.O.; Midgley, G.F.; Raimondo, D.C.; Rebelo, A.G.; Rouget, M.; Thuiller, W. Potential impacts of future land use and climate change on the red list status of the proteaceae in the cape floristic region, south africa. Glob. Chang. Biol. 2005, 11, 1452–1468. [Google Scholar] [CrossRef]
- Midgley, G.F.; Hannah, L.; Millar, D.; Thuiller, W.; Booth, A. Developing regional and species-level assessments of climate change impacts on biodiversity in the cape floristic region. Biol. Conserv. 2003, 112, 87–97. [Google Scholar] [CrossRef]
- Fischer, J.; Lindenmayer, D.B. Landscape modification and habitat fragmentation: A synthesis. Glob. Ecol. Biogeogr. 2007, 16, 265–280. [Google Scholar] [CrossRef]
- Basile, M.; Valerio, F.; Balestrieri, R.; Posillico, M.; Bucci, R.; Altea, T.; De Cinti, B.; Matteucci, G. Patchiness of forest landscape can predict species distribution better than abundance: The case of a forest-dwelling passerine, the short-toed treecreeper, in central italy. PeerJ 2016, 4, e2398. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Fang, J.; Tang, Z.; Lin, X. Patterns, determinants and models of woody plant diversity in china. Proc. R. Soc. 2011, 278, 2122–2132. [Google Scholar] [CrossRef] [PubMed]
- Collinson, M.E. The early fossil history of salicaceae: A brief review. Proc. R. Soc. 1992, 98, 155–167. [Google Scholar] [CrossRef]
- Allen, A.P.; Gillooly, J.F.; Savage, V.M.; Brown, J.H. Kinetic effects of temperature on rates of genetic divergence and speciation. Proc. Natl. Acad. Sci. USA 2006, 103, 9130–9135. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Peng, G.Q.; Wu, C.C.; Han, Q.M. Global warming induces female cuttings of populus cathayana to allocate more biomass, c and n to aboveground organs than do male cuttings. Aust. J. Bot. 2010, 58, 519–526. [Google Scholar] [CrossRef]
- Chen, L.H.; Sheng, Z.; Zhao, H.X.; Korpelainen, H.; Li, C.Y. Sex-related adaptive responses to interaction of drought and salinity in populus yunnanensis. Plant Cell Environ. 2010, 33, 1767–1778. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Peng, G.; Wu, C.; Korpelainen, H.; Li, C. Drought inhibits photosynthetic capacity more in females than in males of populus cathayana. Tree Physiol. 2008, 28, 1751–1759. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.W. Paleobiogeographic relationships of angiosperms from the cretaceous and early tertiary of the north american area. Bot. Rev. 1990, 56, 279–417. [Google Scholar] [CrossRef]
- Ding, T.Y. Origin, divergence and geographical distribution of salicaceae. Acta Bot. Yunnanica 1995, 17, 277–290. [Google Scholar]
- O’Brien, E.M. Climatic gradients in woody plant species richness: Towards an explanation based on an analysis of southern africa’s woody flora. J. Biogeogr. 1993, 20, 181–198. [Google Scholar] [CrossRef]
- O’Brien, E.M. Water–energy dynamics, climate, and prediction of woody plant species richness: An interim general model. J. Biogeogr. 1998, 25, 379–398. [Google Scholar] [CrossRef]
- Evans, K.L.; Warren, P.H.; Gaston, K.J. Species–energy relationships at the macroecological scale: A review of the mechanisms. Biol. Rev. 2005, 80, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Wu, J.; Xing, Q.; Pan, Q.; Huang, J.; Yang, D.; Han, X. Primary production and rain use efficiency across a precipitation gradient on the mongolia plateau. Ecology 2008, 89, 2140–2153. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, R.J.; Nogués-Bravo, D.; Araújo, M.B. Geographical gradients of species richness: A test of the water-energy conjecture of hawkins et al. (2003) using european data for five taxa. Glob. Ecol. Biogeogr. 2006, 16, 76–78. [Google Scholar] [CrossRef]
- Zhang, S.; Chen, F.; Peng, S.; Ma, W.; Korpelainen, H.; Li, C. Comparative physiological, ultrastructural and proteomic analyses reveal sexual differences in the responses of populus cathayana under drought stress. Proteomics 2010, 10, 2661–2677. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Chen, K.; Jiang, H.; Yu, L.; Duan, B. Contrasting responses in the growth and energy utilization properties of sympatric populus and salix to different altitudes: Implications for sexual dimorphism in salicaceae. Physiol. Plant. 2017, 159, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Teramura, A.H. Effects of ultraviolet-b radiation on the growth and yield of crop plants. Physiol. Plant. 2010, 58, 415–427. [Google Scholar] [CrossRef]
- Keiller, D.R.; Holmes, M.G. Effects of long-term exposure to elevated uv-b radiation on the photosynthetic performance of five broad-leaved tree species. Photosynth. Res. 2001, 67, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Zhang, S. Sex-related responses to environmental changes in salicaceae. Mt. Res. 2017, 35, 645–652. [Google Scholar]
- Liu, B.; Liu, X.B.; Li, Y.S.; Herbert, S.J. Effects of enhanced uv-b radiation on seed growth characteristics and yield components in soybean. Field Crops Res. 2013, 154, 158–163. [Google Scholar] [CrossRef]
- Svenning, J.C.; Skov, F. The relative roles of environment and history as controls of tree species composition and richness in europe. J. Biogeogr. 2005, 32, 1019–1033. [Google Scholar] [CrossRef]
- Raes, N.; Roos, M.C.; Slik, J.W.F.; Van Loon, E.E.; Ter Steege, H. Botanical richness and endemicity patterns of borneo derived from species distribution models. Ecography 2009, 32, 180–192. [Google Scholar] [CrossRef]
- Stropp, J.; Ter Steege, H.; Malhi, Y. Disentangling regional and local tree diversity in the amazon. Ecography 2009, 32, 46–54. [Google Scholar] [CrossRef]
- Berthel, N.; Schwörer, C.; Tinner, W. Impact of holocene climate changes on alpine and treeline vegetation at sanetsch pass, bernese alps, switzerland. Rev. Palaeobot. Palynol. 2012, 174, 91–100. [Google Scholar] [CrossRef]
- Singh, S.; Kumar, P.; Rai, A. Ultraviolet radiation stress: Molecular and physiological adaptations in trees. In Abiotic Stress Tolerance in Plants; Springer: Dordrecht, The Netherlands, 2006. [Google Scholar]
- Osprey, S.M.; Butchart, N.; Knight, J.R.; Scaife, A.A.; Hamilton, K.; Anstey, J.A.; Schenzinger, V.; Zhang, C. An unexpected disruption of the atmospheric quasi-biennial oscillation. Science 2016, 353, 1424. [Google Scholar] [CrossRef] [PubMed]
- Dhomse, S.S.; Chipperfield, M.P.; Feng, W.; Hossaini, R.; Mann, G.W.; Santee, M.L. Revisiting the hemispheric asymmetry in midlatitude ozone changes following the mount pinatubo eruption: A 3-d model study. Geophys. Res. Lett. 2015, 42, 3038–3047. [Google Scholar] [CrossRef] [PubMed]
- Dhomse, S.S.; Chipperfield, M.P.; Damadeo, R.P.; Zawodny, J.M.; Haigh, J.D. On the ambiguous nature of the 11-year solar cycle signal in upper stratospheric ozone: Solar signal in upper stratosphere. Geophys. Res. Lett. 2016, 43, 7241–7249. [Google Scholar] [CrossRef]
- Caldwell, M.M. A steep latitudinal gradient of solar ultraviolet-b radiation in the arctic-alpine life zone. Ecology 1980, 61, 600–611. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Environment Variables | Resolution | Unit |
|---|---|---|---|
| Bio1 | Annual mean temperature | 30″ | °C × 10 |
| Bio3 | Isothermality (BIO2/BIO7) ∗ 100 | 30″ | – |
| Bio7 | Temperature annual range | 30″ | °C × 10 |
| Bio12 | Annual precipitation | 30″ | mm |
| Bio15 | Precipitation seasonality (Coefficient of variation) | 30″ | – |
| Elevation | Elevation | 30″ | m |
| S-CE | Cation exchange capacity (CEC) clay subsoil | 30″ | – |
| T-BS | Base saturation% topsoil | 30″ | % |
| T-C | Organic carbon pool topsoil | 30″ | – |
| T-N | Nitrogen % topsoil | 30″ | % |
| Drain | Soil drainage class | 30″ | – |
| UV-B1 | Annual mean UV-B | 15′ | J/m2/day |
| UV-B2 | Ultraviolet-B (UV-B) seasonality | 15′ | J/m2/day |
| UV-B3 | Mean UV-B of lightest month | 15′ | J/m2/day |
| UV-B4 | Mean UV-B of lowest month | 15′ | J/m2/day |
| Climate Scenario/Year | Area (×104 km2) | Proportion of Area (%) | ||||||
|---|---|---|---|---|---|---|---|---|
| Contraction | Expansion | Unchanged | Total | Contraction | Expansion | Unchanged | Total | |
| LIG a | 139.72 | 62.79 | 298.40 | −76.92 | 38.67 | 17.38 | 82.58 | −21.29 |
| LGM b | 37.25 | 106.88 | 254.34 | 69.63 | 10.31 | 29.58 | 70.39 | 19.27 |
| RCP4.5-2050 c | 65.63 | 59.09 | 295.56 | −6.54 | 18.16 | 16.35 | 81.80 | −1.81 |
| RCP4.5-2070 d | 73.18 | 67.82 | 288.01 | −5.36 | 20.25 | 18.77 | 79.71 | −1.48 |
| RCP8.5-2050 e | 82.06 | 78.06 | 279.09 | −4.00 | 22.71 | 21.60 | 77.24 | −1.11 |
| RCP8.5-2070 f | 20.21 | 20.18 | 341.05 | −0.04 | 5.59 | 5.58 | 94.39 | −0.01 |
| SR | WE | CWE | |
|---|---|---|---|
| Contemporary energy | |||
| Bio1 (°C) | 5.26%(+) *** | 3.73%(+) ** | 1.00%(+) |
| Bio7 (°C) | 5.85%(−) | 9.94%(−) | 6.53%(−) |
| Bio3 | 1.20%(+) | 0.92%(+) | 6.40%(+) |
| UVB1 (J m−2·day−1) | 4.80%(−) ** | 0.08%(−) *** | 1.90%(−) *** |
| UVB2 (J m−2·day−1) | 18.20(−) *** | 8.75%(−) | 1.20%(−) |
| UVB3 (J m−2·day−1) | 11.85%(−) *** | 2.72%(−) *** | 0.11%(−) ** |
| UVB4 (J m−2·day−1) | 0.03%(−) | 2.33%(−) *** | 6.8%(+) *** |
| Contemporary water availability | |||
| Bio12 (mm) | 10.22%(+) ** | 10.07%(+) ** | 5.90%(+) *** |
| Bio15 | 2.04%(−) ** | 2.21%(−) | 3.81%(−) |
| Contemporary soil conditions | |||
| S-CE | 9.71%(+) | 3.74%(+) | 0.01%(+) |
| T-N | 4.25%(+) *** | 8.01%(+) *** | 11.06%(+) *** |
| Drain | 0.12%(+) | 0.21%(−) | 0.02%(−) |
| T-BS | 0.10%(+) | 0.18%(+) | 1.95%(+) |
| Heterogeneity | |||
| Elevation | 6.59%(−) ** | 1.31%(−) | 0.04%(−) ** |
| Historical climate change | |||
| Bio1-Ano | 0.18%(−) *** | 0.45%(−) | 0.05%(−) |
| Bio3-Ano | 2.92%(+) ** | 3.18%(+) *** | 4.19%(+) ** |
| Bio7-Ano | 4.58%(−) ** | 1.22%(−) | 0.09%(−) |
| Bio12-Ano | 13.17%(+) | 6.30%(+) *** | 1.33%(+) *** |
| Bio15-Ano | 19.97%(−) ** | 12.63%(−) *** | 3.80%(−) *** |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Shi, M.; Huang, Y.; Chen, K.; Sun, H.; Chen, J. Climatic Change Can Influence Species Diversity Patterns and Potential Habitats of Salicaceae Plants in China. Forests 2019, 10, 220. https://doi.org/10.3390/f10030220
Li W, Shi M, Huang Y, Chen K, Sun H, Chen J. Climatic Change Can Influence Species Diversity Patterns and Potential Habitats of Salicaceae Plants in China. Forests. 2019; 10(3):220. https://doi.org/10.3390/f10030220
Chicago/Turabian StyleLi, Wenqing, Mingming Shi, Yuan Huang, Kaiyun Chen, Hang Sun, and Jiahui Chen. 2019. "Climatic Change Can Influence Species Diversity Patterns and Potential Habitats of Salicaceae Plants in China" Forests 10, no. 3: 220. https://doi.org/10.3390/f10030220
APA StyleLi, W., Shi, M., Huang, Y., Chen, K., Sun, H., & Chen, J. (2019). Climatic Change Can Influence Species Diversity Patterns and Potential Habitats of Salicaceae Plants in China. Forests, 10(3), 220. https://doi.org/10.3390/f10030220