Differences in the Spatial Structure of Two Pinus cembra L. Populations in the Carpathian Mountains



Abstract
:1. Introduction
2. Methods
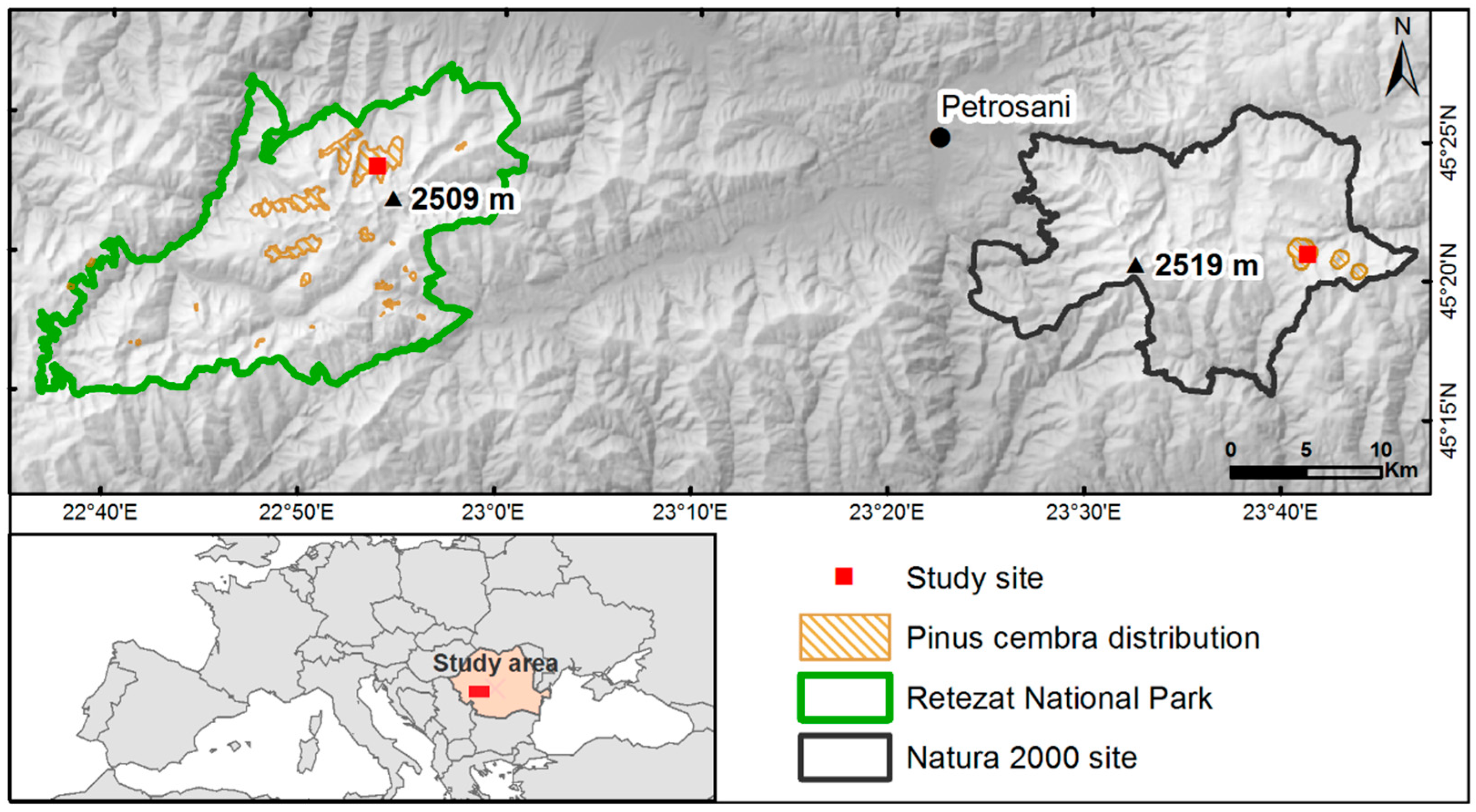
2.1. Study Area
2.2. Spatial Patterns Analysis
2.3. Modelling the Spatial Relationship between Tree Sizes
2.4. Spatial Correlation and Autocorrelation
3. Results
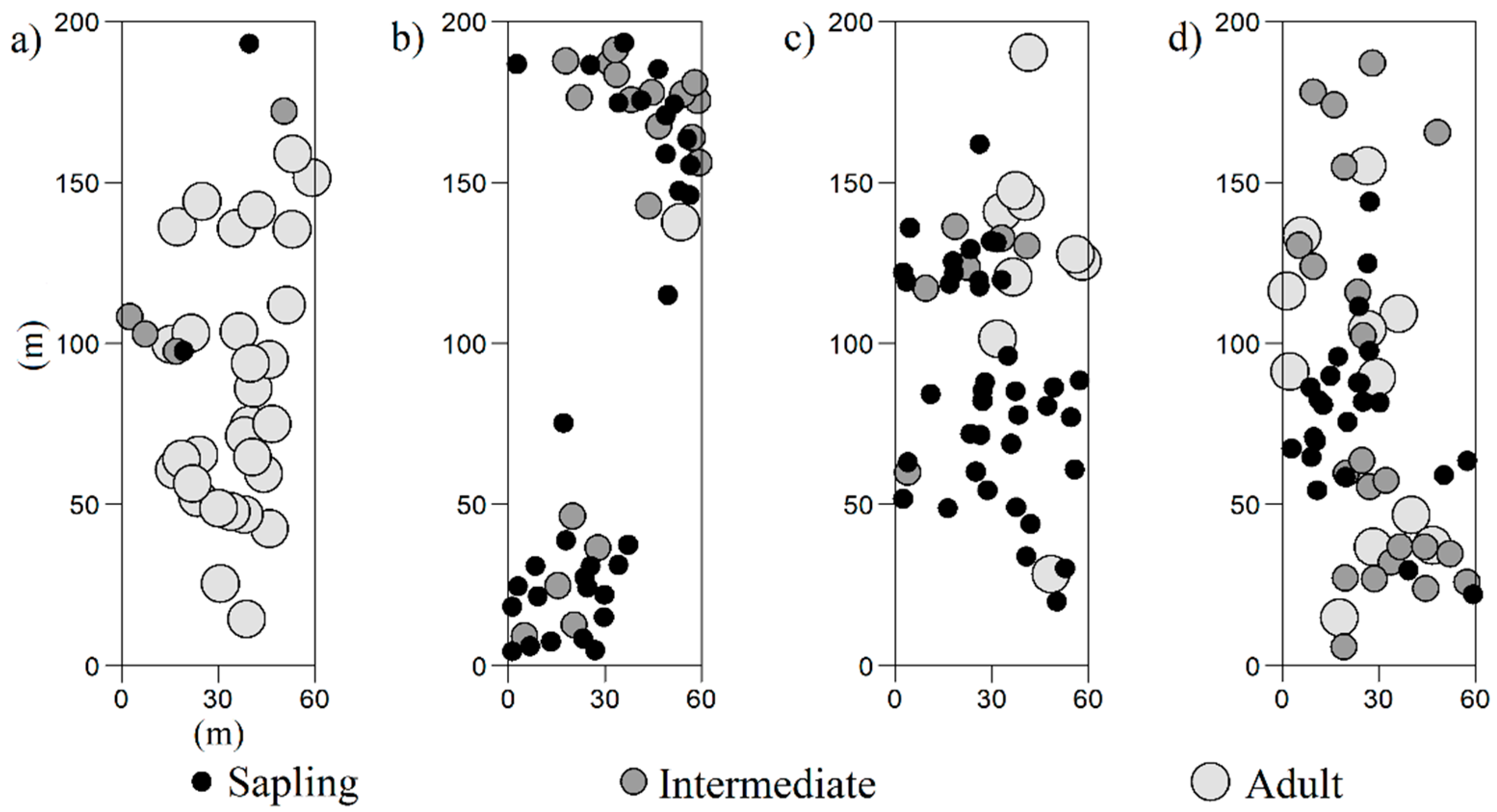
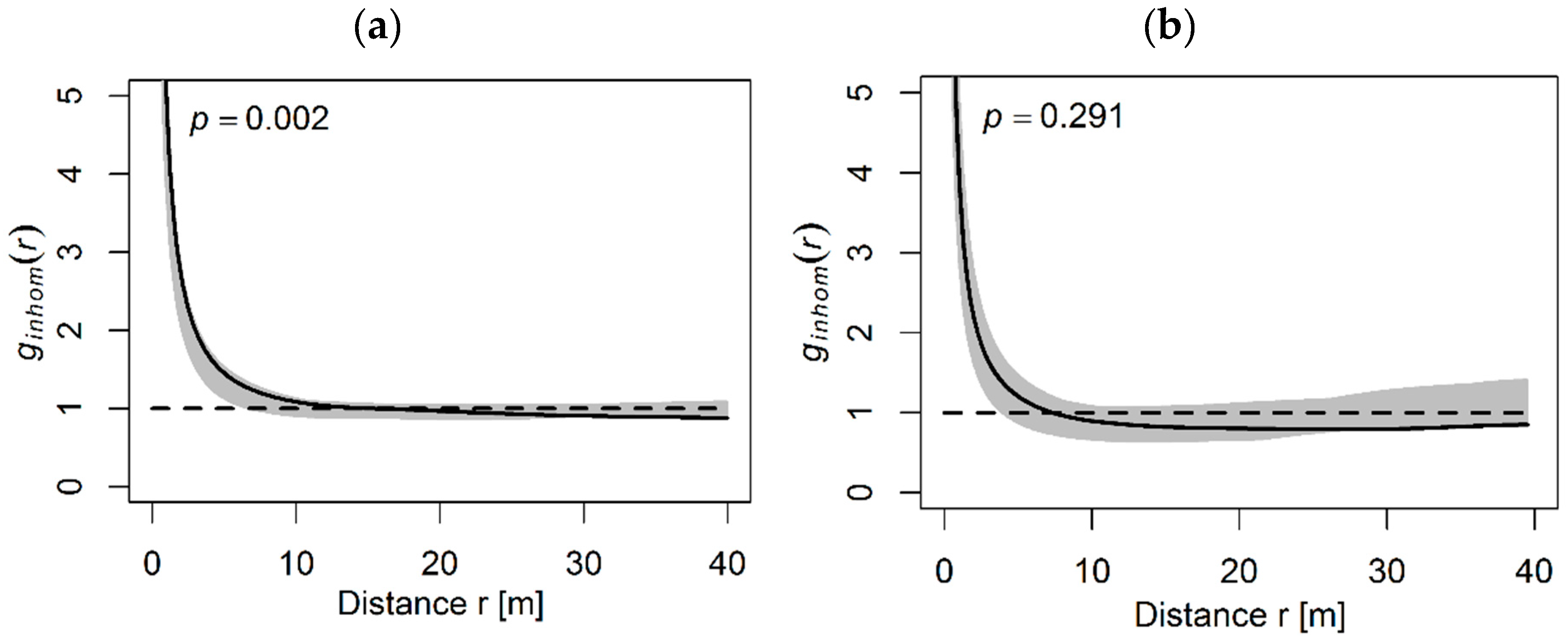
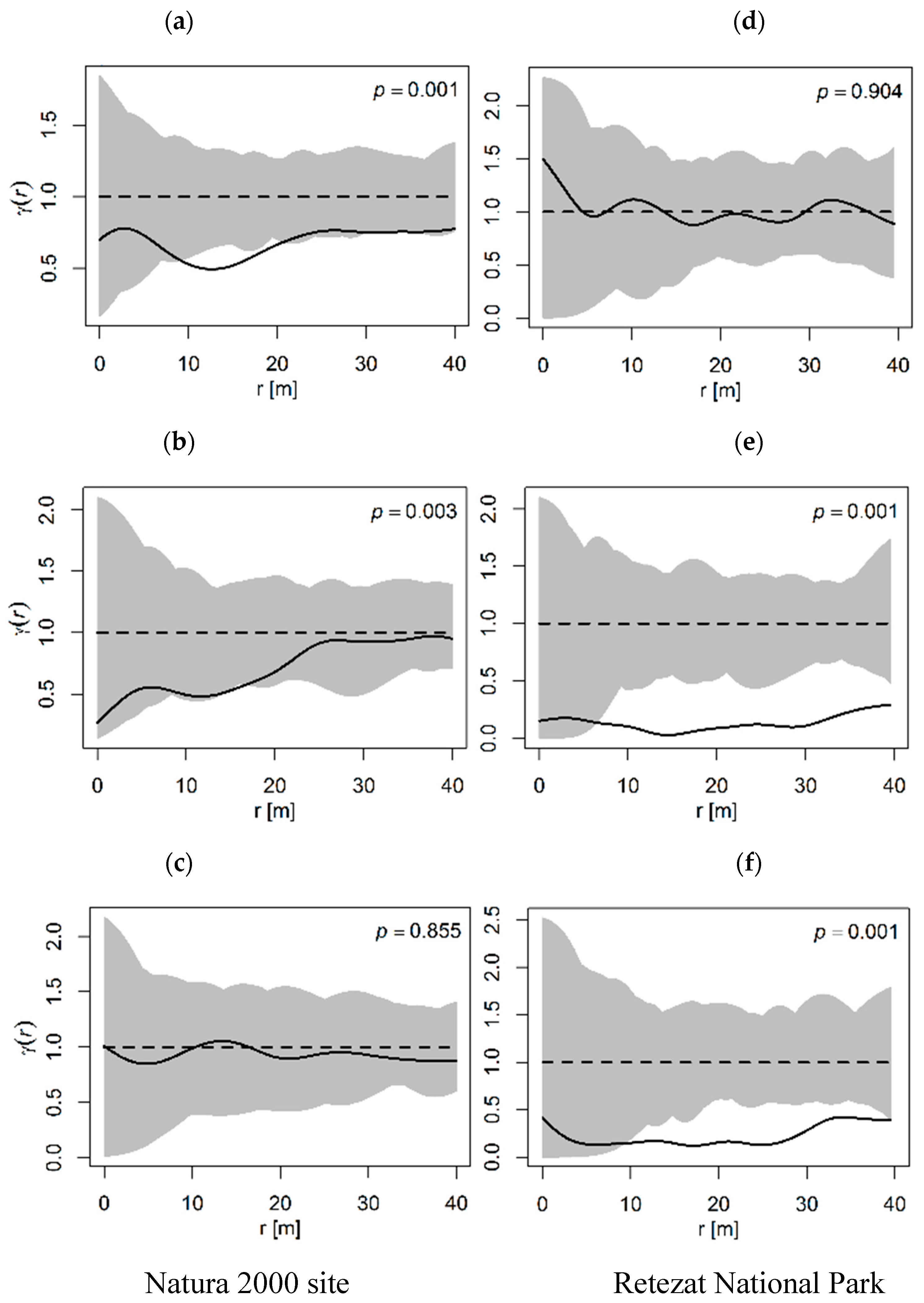
3.1. General Spatial Distribution Pattern
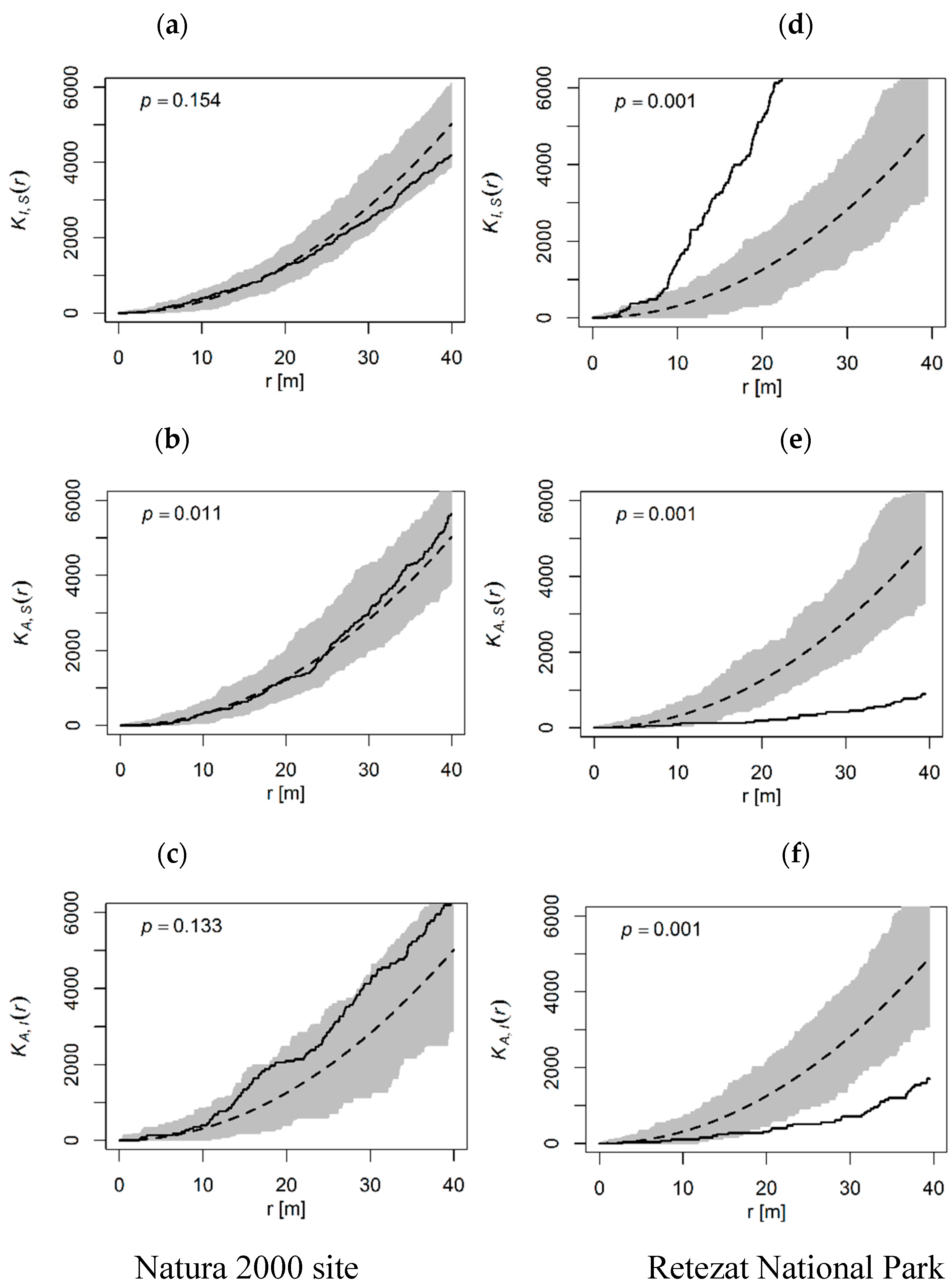
3.2. Spatial Distribution of the Tree Size Classes
3.3. Size Hierarchies
4. Discussion
4.1. Spatial Patterns at the Treeline
4.2. Spatial Structure and Facilitation
4.3. Local Size Hierarchies
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hanski, I. Metapopulation Dynamics. Nature 1998, 396, 41–49. [Google Scholar] [CrossRef]
- Nathan, R.; Muller-Landau, H.C. Spatial patterns of seed dispersal, their determinants and consequences for recruitment. Trends Ecol. Evol. 2000, 15, 278–285. [Google Scholar] [CrossRef] [Green Version]
- MacArthur, R.H. Geographical Ecology. Patterns in the Distribution of Species; Harper and Row: New York, NY, USA, 1972. [Google Scholar]
- Levin, S.A. The problem of pattern and scale in ecology: the Robert H. MacArthur award lecture. Ecology 1992, 73, 1943–1967. [Google Scholar] [CrossRef]
- Callaway, R.M.; Walker, L.R. Competition and facilitation: a synthetic approach to interactions in plant communities. Ecology 1997, 78, 1958. [Google Scholar] [CrossRef]
- Getzin, S.; Dean, C.; He, F.; Trofymow, J.A.; Wiegand, K.; Wiegand, T. Spatial patterns and competition of tree species in a Douglas-fir chronosequence on Vancouver Island. Ecography 2006, 29, 671–682. [Google Scholar] [CrossRef] [Green Version]
- Bruno, J.F.; Stachowicz, J.J.; Bertness, M.D. Inclusion of facilitation into ecological theory. Trends Ecol. Evol. 2003, 18, 119–125. [Google Scholar] [CrossRef]
- Janzen, D.H. Herbivores and the number of tree species in tropical forests. Am. Nat. 1970, 104, 501–528. [Google Scholar] [CrossRef]
- Connell, J.H. On the role of natural enemies in preventing competitive exclusion in some marine animals and in rain forest trees. In Dynamics of Populations; Boer, P.J.D., Gradwell, G.R., Eds.; Centre for Agricultural Publication and Documentation: Wageningen, The Netherlands, 1971; pp. 298–312. [Google Scholar]
- Martínez, I.; González Taboada, F.; Wiegand, T.; Obeso, J.R. Spatial patterns of seedling-adult associations in a temperate forest community. For. Ecol. Manag. 2013, 296, 74–80. [Google Scholar] [CrossRef]
- Hille Ris Lambers, J.; Clark, J.S.; Beckage, B. Density-dependent mortality and the latitudinal gradient in species diversity. Nature 2002, 417, 732–735. [Google Scholar] [CrossRef]
- Camarero, J.J.; Gutiérrez, E.; Fortin, M.-J.; Ribbens, E. Spatial patterns of tree recruitment in a relict population of Pinus uncinata: forest expansion through stratified diffusion. J. Biogeogr. 2005, 32, 1979–1992. [Google Scholar] [CrossRef]
- Wiegand, T.; Moloney, K.A. Handbook of Spatial Point-Pattern Analysis in Ecology; Chapman and Hall/CRC: Boca Raton, FL, USA, 2014. [Google Scholar]
- Casalegno, S.; Amatulli, G.; Camia, A.; Nelson, A.; Pekkarinen, A. Vulnerability of Pinus cembra L. in the Alps and the Carpathian Mountains under present and future climates. For. Ecol. Manag. 2010, 259, 750–761. [Google Scholar] [CrossRef]
- Farjon, A.; Pinus Cembra. The IUCN Red List of Threatened Species 2017: E.T42349A95684563. Available online: https://doi.org/10.2305/IUCN.UK.2017-2.RLTS.T42349A95684563.en (accessed on 12 September 2018).
- Blada, I. Pinus cembra distribution in the Romanian Carpathians. Ann. For. Res. 2008, 51, 115–132. [Google Scholar]
- Höhn, M.; Gugerli, F.; Abran, P.; Bisztray, G.; Buonamici, A.; Cseke, K.; Hufnagel, L.; Quintela-Sabarís, C.; Sebastiani, F.; Vendramin, G.G. Variation in the chloroplast DNA of Swiss stone pine (Pinus cembra L.) reflects contrasting post-glacial history of populations from the Carpathians and the Alps. J. Biogeogr. 2009, 36, 1798–1806. [Google Scholar] [CrossRef]
- Dzialuk, A.; Chybicki, I.; Gout, R.; Mączka, T.; Fleischer, P.; Konrad, H.; Curtu, A.L.; Sofletea, N.; Valadon, A. No reduction in genetic diversity of Swiss stone pine (Pinus cembra L.) in Tatra Mountains despite high fragmentation and small population size. Conserv. Genet. 2014, 15, 1433–1445. [Google Scholar] [CrossRef] [Green Version]
- Blada, I. Zâmbrul: I. Importanță și Areal; Editura Silvică, Seria ‘Manuale, tratate, monografii’: Voluntari, Romania, 2015. (In Romanian) [Google Scholar]
- Vucetich, J.A.; Waite, T.A. Spatial patterns of demography and genetic processes across the species’ range: null hypotheses for landscape conservation genetics. Conserv. Genet. 2003, 4, 639–645. [Google Scholar] [CrossRef]
- Hampe, A.; Petit, R.J. Conserving biodiversity under climate change: The rear edge matters. Ecol. Lett. 2005, 8, 461–467. [Google Scholar] [CrossRef]
- Iojă, C.I.; Pătroescu, M.; Rozylowicz, L.; Popescu, V.D.; Vergheleţ, M.; Zotta, M.I.; Felciuc, M. The efficacy of Romania’s protected areas network in conserving biodiversity. Biol. Conserv. 2010, 143, 2468–2476. [Google Scholar] [CrossRef]
- Neuschulz, E.L.; Merges, D.; Bollmann, K.; Gugerli, F.; Böhning-Gaese, K. Biotic interactions and seed deposition rather than abiotic factors determine recruitment at elevational range limits of an alpine tree. J. Ecol. 2018, 106, 948–959. [Google Scholar] [CrossRef]
- Evans, D. Building the European Union’s Natura 2000 network. Nat. Conserv. 2012, 1, 11–26. [Google Scholar] [CrossRef] [Green Version]
- Lingua, E.; Cherubini, P.; Motta, R.; Nola, P. Spatial structure along an altitudinal gradient in the Italian central Alps suggests competition and facilitation among coniferous species. J. Veg. Sci. 2008, 19, 425–436. [Google Scholar] [CrossRef] [Green Version]
- Körner, C.; Paulsen, J. A world-wide study of high altitude treeline temperatures. J. Biogeogr. 2004, 31, 713–732. [Google Scholar] [CrossRef]
- Holtmeier, F.-K. Mountain Timberlines: Ecology, Patchiness, and Dynamics, 2nd ed.; Preface. Advances in global change research; Springer: Dordrecht, The Netherlands, 2009. [Google Scholar]
- Watson, J.E.M.; Dudley, N.; Segan, D.B.; Hockings, M. The performance and potential of protected areas. Nature 2014, 515, 67–73. [Google Scholar] [CrossRef]
- Gray, C.L.; Hill, S.L.L.; Newbold, T.; Hudson, L.N.; Börger, L.; Contu, S.; Hoskins, A.J.; Ferrier, S.; Purvis, A.; Scharlemann, J.P.W. Local biodiversity is higher inside than outside terrestrial protected areas worldwide. Nat. Commun. 2016, 7, 12306. [Google Scholar] [CrossRef] [Green Version]
- Gehrig-Fasel, J.; Guisan, A.; Zimmermann, N.E. Tree line shifts in the Swiss Alps: Climate Change or Land Abandonment? J. Veg. Sci. 2007, 18, 571–582. [Google Scholar] [CrossRef]
- Shandra, O.; Weisberg, P.; Martazinova, V. Influences of climate and land use history on forest and timberline dynamics in the Carpathian Mountains during the twentieth century. In The Carpathians: Integrating Nature and Society Towards Sustainability; Kozak, J., Ostapowicz, K., Bytnerowicz, A., Wyżga, B., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 209–223. [Google Scholar]
- Mihai, B.; Savulescu, I.; Sandric, I. Change detection analysis (1986–2002) of vegetation cover in Romania: a study of alpine, subalpine, and forest landscapes in the Iezer Mountains, Southern Carpathians. Mt. Res. Dev. 2007, 27, 250–258. [Google Scholar] [CrossRef]
- Urdea, P.; Törok-Oance, M.; Ardelean, M.; Vuia, F.; Voiculescu, M. Geomorphological aspects of the human impact in the alpine area of Southern Carpathians (Romania). Hrvat. Geogr. Glas. 2009, 71, 19–31. [Google Scholar] [CrossRef]
- Oberhuber, W. Influence of climate on radial growth of Pinus cembra within the Alpine timberline Ecotone. Tree Physiol. 2004, 24, 291–301. [Google Scholar] [CrossRef]
- Popa, I.; Kern, Z. Long-term summer temperature reconstruction inferred from tree-ring records from the Eastern Carpathians. Clim. Dyn. 2009, 32, 1107–1117. [Google Scholar] [CrossRef]
- Carrer, M.; Urbinati, C. Spatial analysis of structural and tree-ring related parameters in a timberline forest in the Italian Alps. J. Veg. Sci. 2001, 12, 643–652. [Google Scholar] [CrossRef]
- Urdea, P.; Vuia, F. Aspects of the periglaciar relief in the Parâng Mountains. Rev. Geomorfol. 2000, 2, 35–39. Available online: http://www.geomorfologie.ro/wp-content/uploads/2015/07/Revista-de-geomorfologie-nr.-2-2000-p.-35-39.pdf (accessed on 14 November 2018).
- Vespremeanu-Stroe, A.; Urdea, P.; Popescu, R.; Vasile, M. Rock glacier activity in the Retezat Mountains, Southern Carpathians, Romania. Permafr. Periglac. Process. 2012, 23, 127–137. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org (accessed on 20 August 2018).
- Getzin, S.; Wiegand, T.; Wiegand, K.; He, F. Heterogeneity influences spatial patterns and demographics in forest stands. J. Ecol. 2008, 96, 807–820. [Google Scholar] [CrossRef] [Green Version]
- Baddeley, A.J.; Møller, J.; Waagepetersen, R. Non- and semi-parametric estimation of interaction in inhomogeneous point patterns. Stat. Neerl. 2000, 54, 329–350. [Google Scholar] [CrossRef]
- Wiegand, T.; Gunatilleke, S.; Gunatilleke, N. Species associations in a heterogeneous Sri Lankan Dipterocarp forest. Am. Nat. 2007, 170, E77–E95. [Google Scholar] [CrossRef]
- Baddeley, A.; Rubak, E.; Turner, R. Spatial Point Pattern Analysis, Model-Fitting, Simulation, Tests. 2019, p. 830. Available online: ftp://ftp.math.ethz.ch/sfs/pub/Software/R-CRAN/web/packages/spatstat/spatstat.pdf (accessed on 15 January 2019).
- Stoyan, D.; Penttinen, A. Recent applications of point process methods in forestry statistics. Stat. Sci. 2000, 15, 61–78. [Google Scholar]
- Diggle, P.J.; Cox, T.F. Some distance-based tests of independence for sparsely-sampled multivariate spatial point patterns. Int. Stat. Rev. Rev. Int. De Stat. 1983, 51, 11. [Google Scholar] [CrossRef]
- Dixon, P.M. Nearest-neighbor contingency table analysis of spatial segregation for several species. Écoscience 2002, 9, 142–151. [Google Scholar] [CrossRef]
- Szmyt, J. Spatial statistics in ecological analysis: from indices to functions. Silva Fenn. 2014, 48. [Google Scholar] [CrossRef]
- Wälder, K.; Wälder, O. Analysing interaction effects in forests using the mark correlation function. Iforest Biogeosci. For. 2008, 1, 34–38. [Google Scholar] [CrossRef] [Green Version]
- Illian, J.; Penttinen, A.; Stoyan, H.; Stoyan, D. Statistical Analysis and Modelling of Spatial Point Patterns: Illian/Statistical Analysis and Modelling of Spatial Point Patterns; John Wiley & Sons, Ltd.: Chichester, UK, 2008. [Google Scholar] [CrossRef]
- Pommerening, A.; Särkkä, A. What mark variograms tell about spatial plant interactions. Ecol. Model. 2013, 251, 64–72. [Google Scholar] [CrossRef]
- Wälder, O.; Stoyan, D. On variograms in point process statistics. Biom. J. 1996, 38, 895–905. [Google Scholar] [CrossRef]
- Suzuki, S.N.; Kachi, N.; Suzuki, J.-I. Development of a local size hierarchy causes regular spacing of trees in an even-aged Abies forest: analyses using spatial autocorrelation and the mark correlation function. Ann. Bot. 2008, 102, 435–441. [Google Scholar] [CrossRef]
- Loosmore, N.B.; Ford, E.D. Statistical inference using the G or K point pattern spatial statistics. Ecology 2006, 87, 1925–1931. [Google Scholar] [CrossRef]
- Barbeito, I.; Brücker, R.L.; Rixen, C.; Bebi, P. Snow fungi-induced mortality of Pinus cembra at the alpine treeline: evidence from plantations. Arct. Antarct. Alp. Res. 2013, 45, 455–470. [Google Scholar] [CrossRef]
- Barbeito, I.; Dawes, M.A.; Rixen, C.; Senn, J.; Bebi, P. Factors driving mortality and growth at treeline: A 30-Year Experiment of 92 000 Conifers. Ecology 2012, 93, 389–401. [Google Scholar] [CrossRef]
- Kueppers, L.M.; Conlisk, E.; Castanha, C.; Moyes, A.B.; Germino, M.J.; de Valpine, P.; Torn, M.S.; Mitton, J.B. Warming and provenance limit tree recruitment across and beyond the elevation range of subalpine forest. Glob. Chang. Biol. 2017, 23, 2383–2395. [Google Scholar] [CrossRef]
- Asselin, H.; Fortin, M.-J.; Bergeron, Y. Spatial distribution of late-successional coniferous species regeneration following disturbance in southwestern Québec boreal forest. For. Ecol. Manag. 2001, 140, 29–37. [Google Scholar] [CrossRef]
- Vittoz, P.; Rulence, B.; Largey, T.; Freléchoux, F. Effects of climate and land-use change on the establishment and growth of cembran pine (Pinus cembra L.) over the altitudinal treeline ecotone in the Central Swiss Alps. Arct. Antarct. Alp. Res. 2008, 40, 225–232. [Google Scholar] [CrossRef]
- Grace, J. Impacts of climate change on the tree line. Ann. Bot. 2002, 90, 537–544. [Google Scholar] [CrossRef]
- Mihai, B.; Săvulescu, I.; Rujoiu-Mare, M.; Nistor, C. Recent forest cover changes (2002–2015) in the Southern Carpathians: a case study of the Iezer Mountains, Romania. Sci. Total Environ. 2017, 599–600, 2166–2174. [Google Scholar] [CrossRef]
- Smithers, B.V.; North, M.P.; Millar, C.I.; Latimer, A.M. Leap frog in slow motion: divergent responses of tree species and life stages to climatic warming in Great Basin subalpine forests. Glob. Chang. Biol. 2018, 24, e442–e457. [Google Scholar] [CrossRef]
- Närmann, F.; Küfmann, C.; Neuschulz, E.L. Präferenzen des Tannenhähers Nucifraga caryocatactes beim Anlegen von Samenverstecken. Ornithol. Anz. 2017, 55, 89–98. [Google Scholar]
- Rolland, C.; Petitcolas, V.; Michalet, R. Changes in radial tree growth for Picea abies, Larix decidua, Pinus cembra and Pinus uncinata near the alpine timberline since 1750. Trees 1998, 13, 40–53. [Google Scholar] [CrossRef]
- Theurillat, J.-P.; Guisan, A. Potential impact of climate change on vegetation in the European Alps: A Review. Clim. Chang. 2001, 50, 77–109. [Google Scholar] [CrossRef]
- Motta, R.; Nola, P. Growth trends and dynamics in sub-alpine forest stands in the Varaita Valley (Piedmont, Italy) and their relationships with human activities and global change. J. Veg. Sci. 2001, 12, 219–230. [Google Scholar] [CrossRef]
- Szmyt, J.; Stoyan, D. Spatial autocorrelation of tree attributes in naturally regenerated managed beech (Fagus sylvatica) forests in the Beskid Niski Mountains, Southern Poland. Dendrobiology 2014, 71, 129–136. [Google Scholar] [CrossRef]
- Reed, D.D.; Burkhart, H.E. Spatial autocorrelation of individual tree characteristics in Loblolly pine stands. For. Sci. 1985, 31, 575–587. [Google Scholar] [CrossRef]
- Freeman, E.A.; Ford, E.D. Effects of data quality on analysis of ecological pattern using the K(d) statistical function. Ecology 2002, 83, 35–46. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plots | Natura 2000 Site | Retezat National Park |
|---|---|---|
| # sapling (10–130 cm height) | 64 | 33 |
| # intermediate (1–15 cm DBH) | 28 | 24 |
| # adult (>15 cm DBH) | 20 | 31 |
| DBH Intermediate (cm) (mean, min-max) | 5.4 (1.9–14.3) | 6.1 (0.6–14.9) |
| DBH Adult (cm) (mean, min-max) | 44.2 (15.2–111.4) | 32.6 (16.5–55.4) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beloiu, M.; Beierkuhnlein, C. Differences in the Spatial Structure of Two Pinus cembra L. Populations in the Carpathian Mountains. Forests 2019, 10, 326. https://doi.org/10.3390/f10040326
Beloiu M, Beierkuhnlein C. Differences in the Spatial Structure of Two Pinus cembra L. Populations in the Carpathian Mountains. Forests. 2019; 10(4):326. https://doi.org/10.3390/f10040326
Chicago/Turabian StyleBeloiu, Mirela, and Carl Beierkuhnlein. 2019. "Differences in the Spatial Structure of Two Pinus cembra L. Populations in the Carpathian Mountains" Forests 10, no. 4: 326. https://doi.org/10.3390/f10040326
APA StyleBeloiu, M., & Beierkuhnlein, C. (2019). Differences in the Spatial Structure of Two Pinus cembra L. Populations in the Carpathian Mountains. Forests, 10(4), 326. https://doi.org/10.3390/f10040326