Genome Size Variation within Species of Chinese Jujube (Ziziphus jujuba Mill.) and Its Wild Ancestor Sour Jujube (Z. acidojujuba Cheng et Liu)

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Flow Cytometry Measurement
2.3. Fruit Morphological Analysis
2.4. Fruit Chemical Analysis
2.5. Statistical Analyses
3. Results
3.1. Validation of the Accuracy of the Genome Size Estimation Method
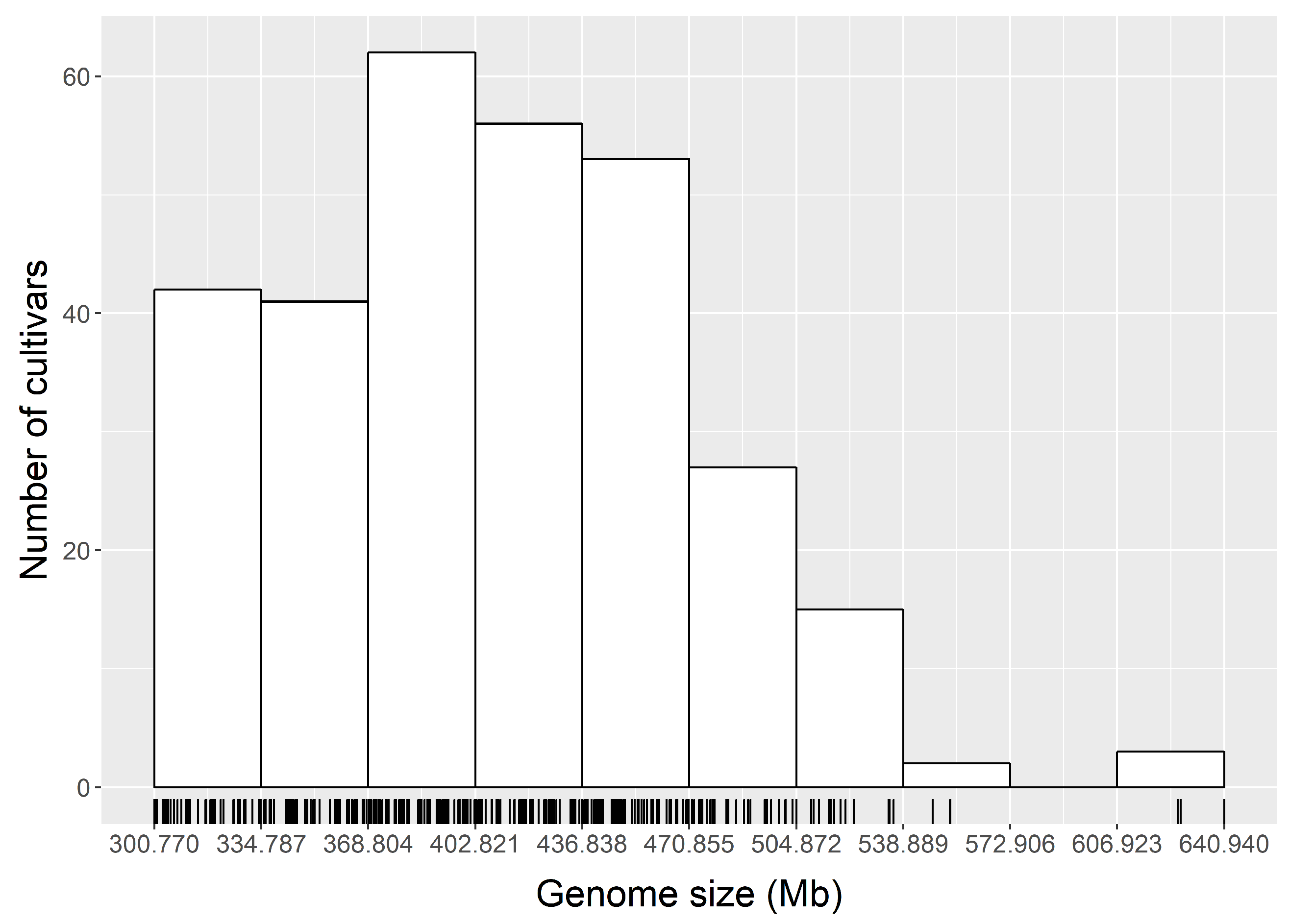
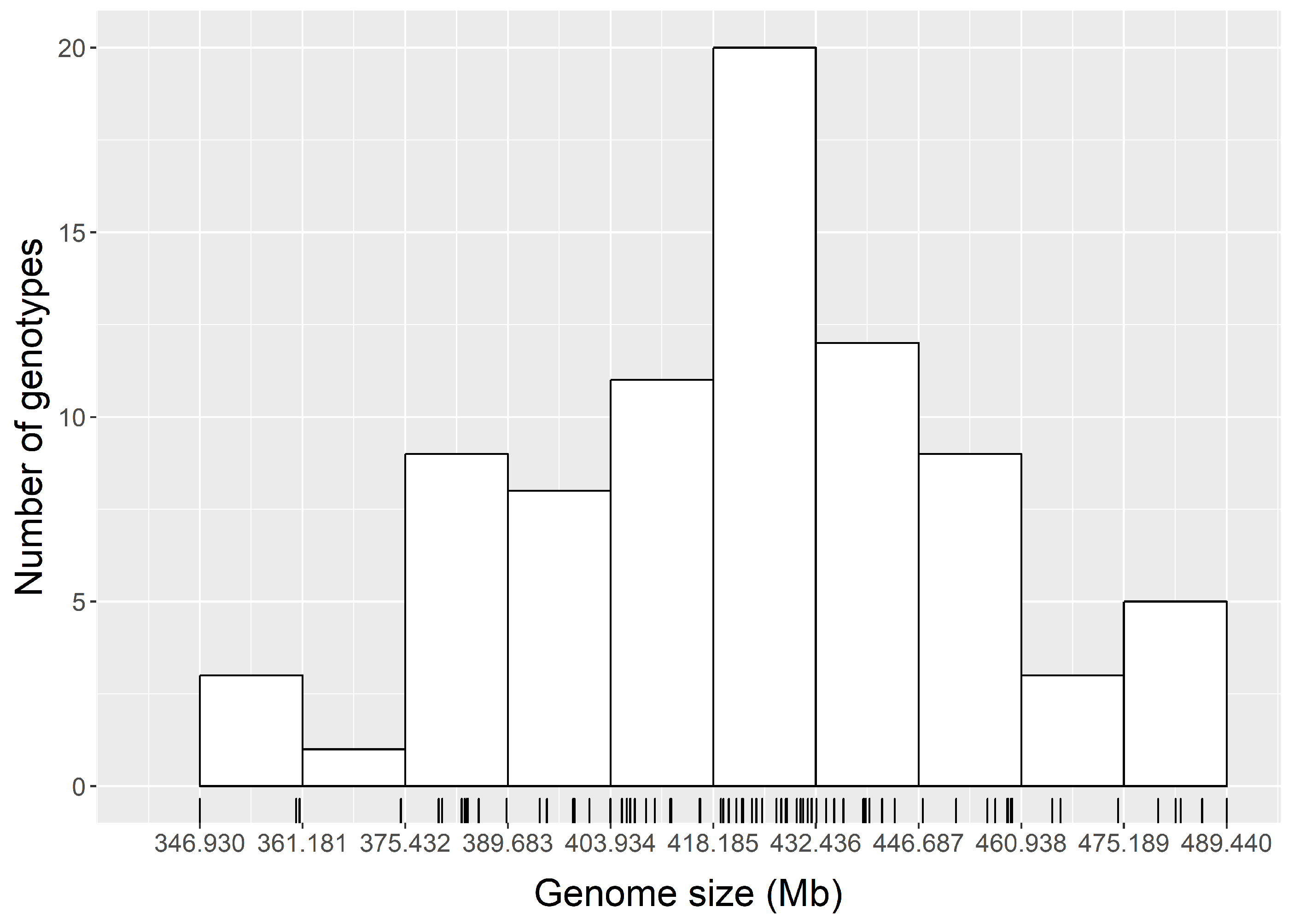
3.2. Genome Size Variation within Chinese Jujube
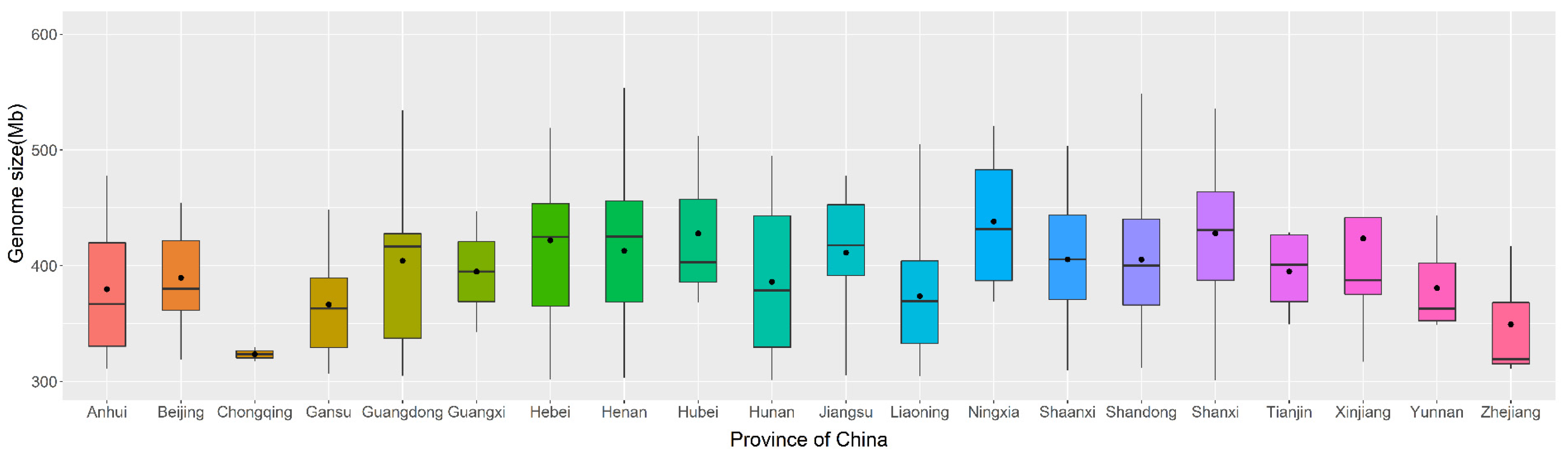
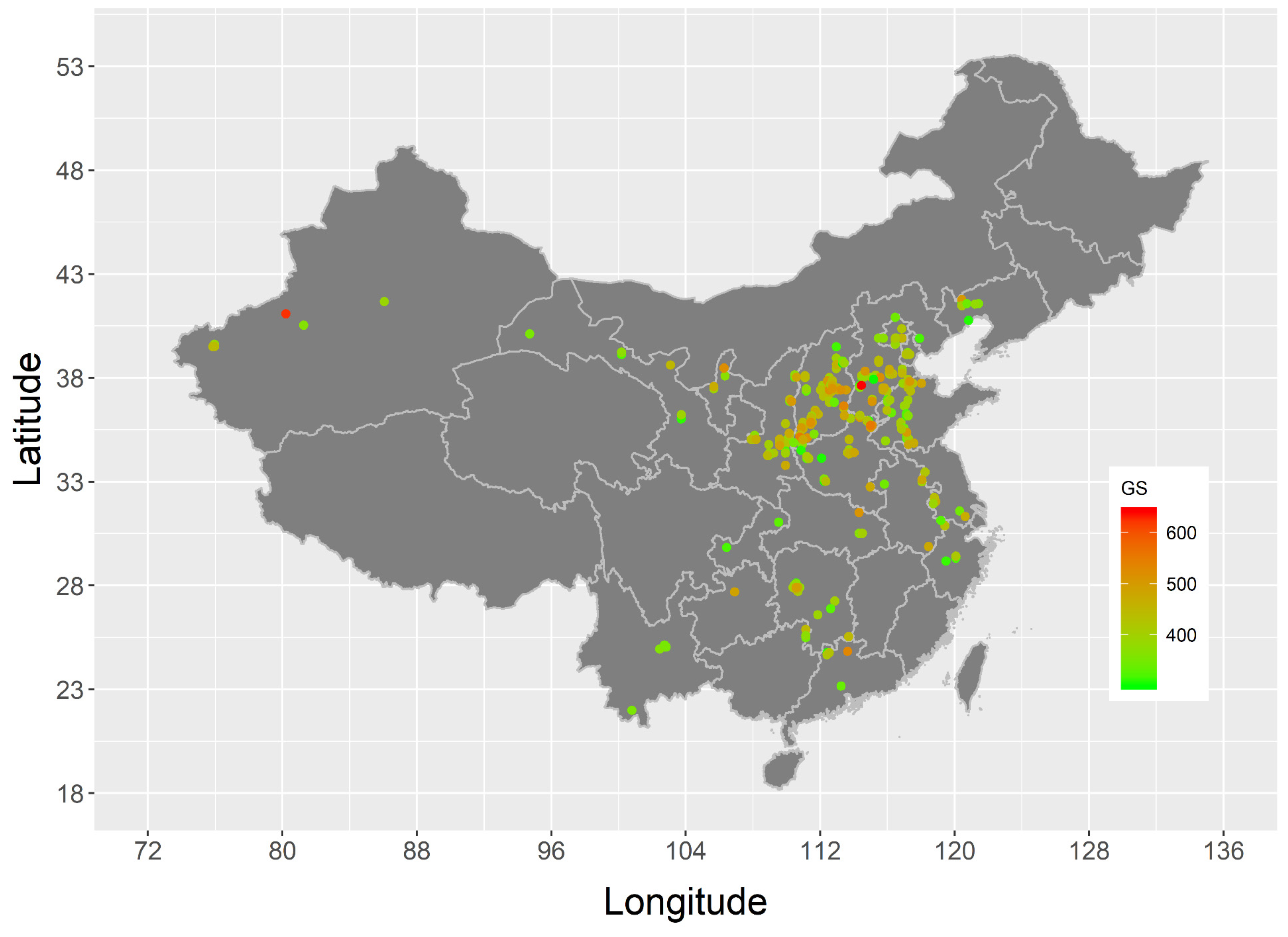
3.3. Geographical Variation in the Genome Size of Chinese Jujube
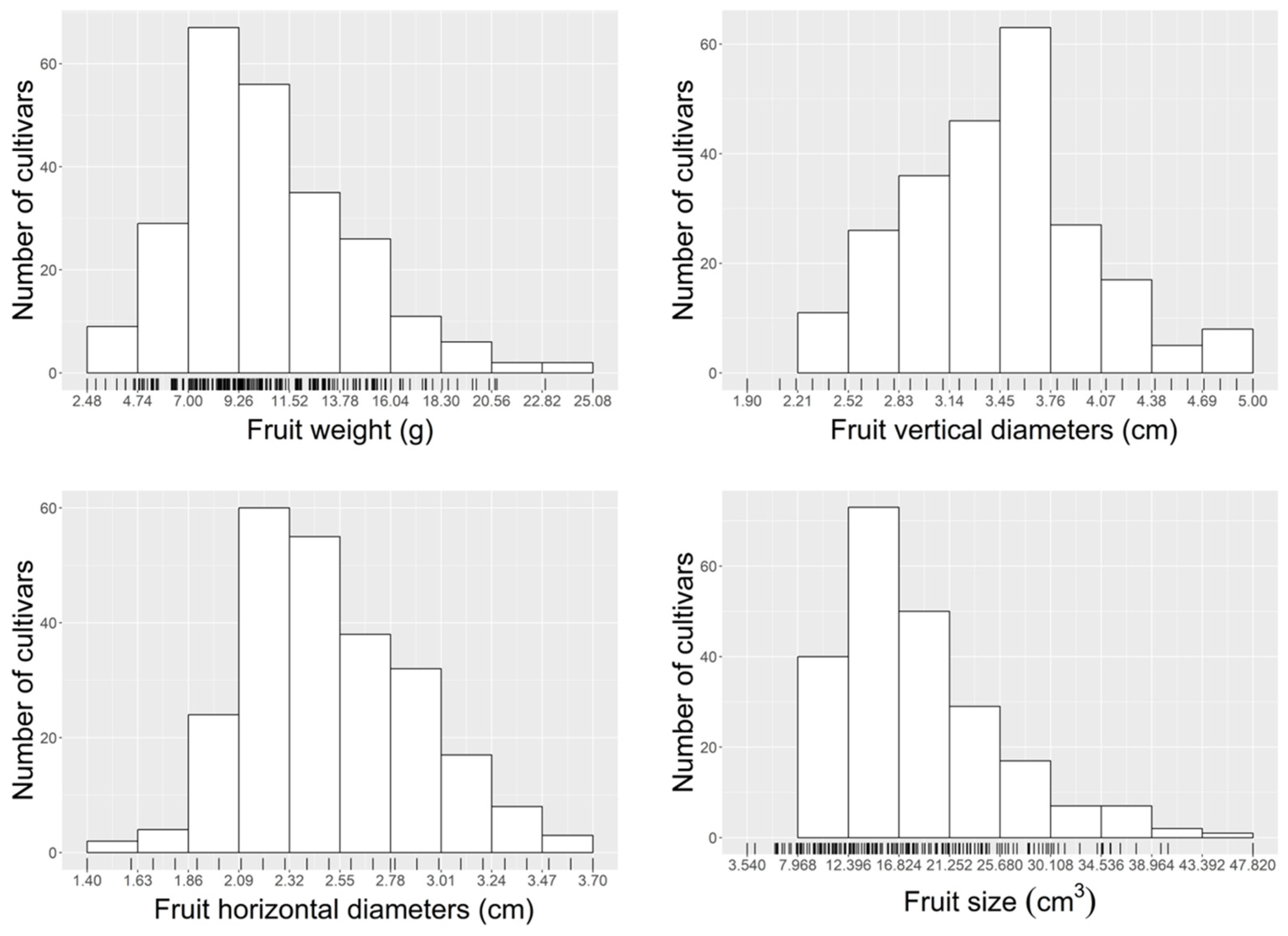
3.4. Morphological and Chemical Variation in Fruit among Cultivars of Chinese Jujube
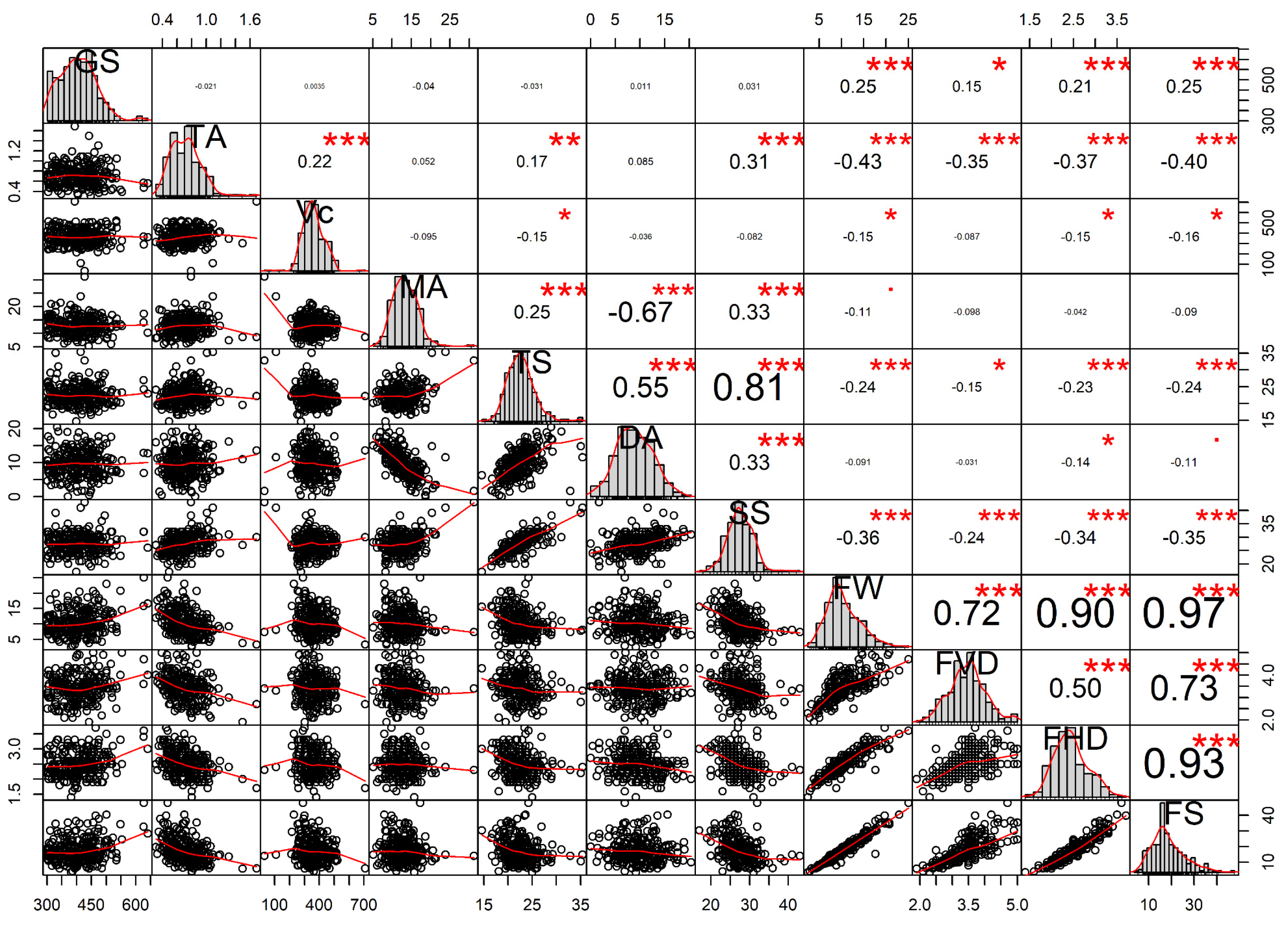
3.5. Correlation Analysis between Genome Sizes and Fruit Traits
3.6. Genome Size Variation in Sour Jujube
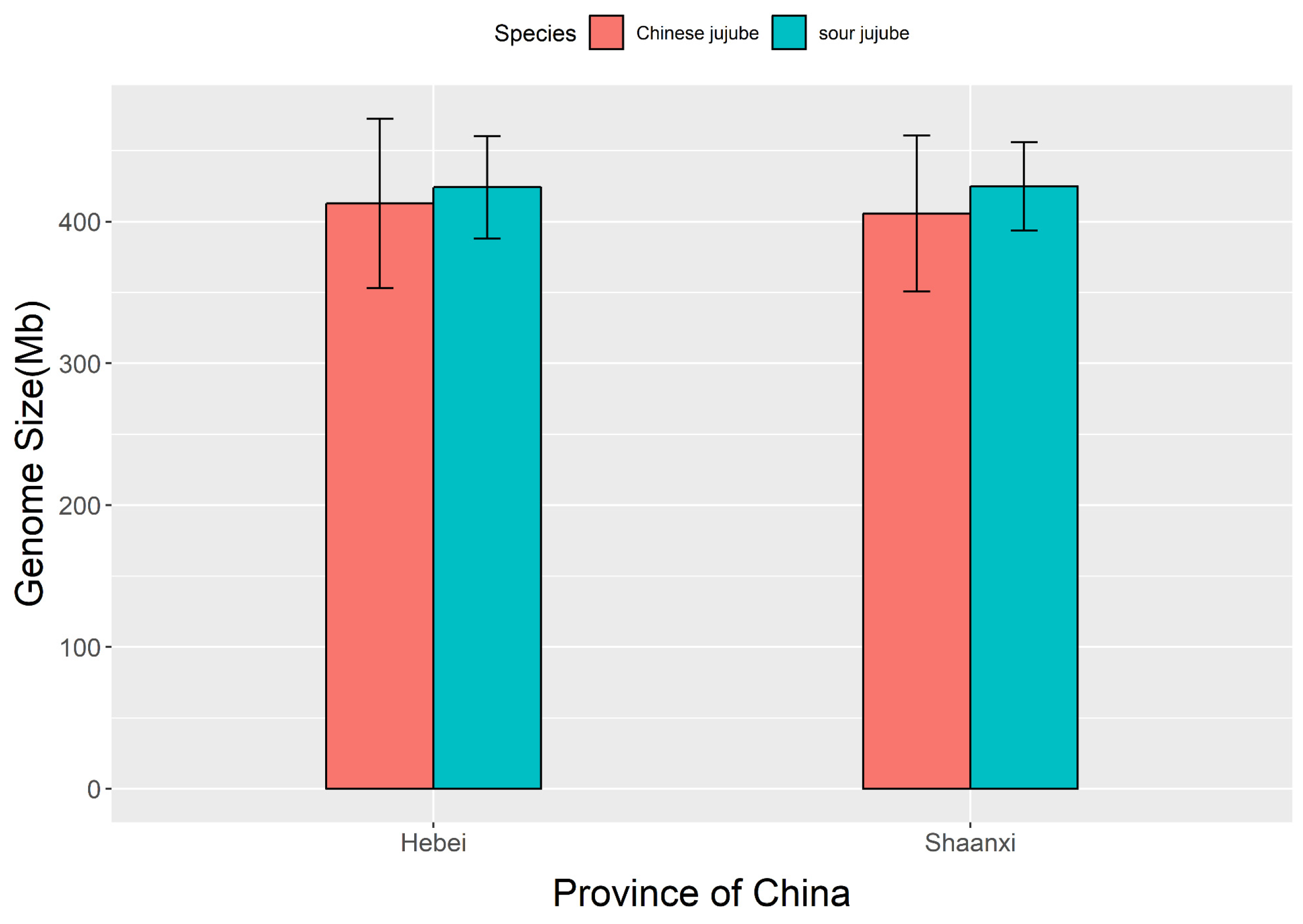
3.7. Comparison of Genome Sizes between Chinese Jujube and Sour Jujube
4. Discussion
4.1. The Genome Size of Chinese Jujube and Sour Jujube
4.2. Association of Genome Size with Fruit Phenotype in Chinese Jujube
4.3. Genome Downsizing during Domestication of Chinese Jujube
4.4. The Possible Mechanism of Large Genome Size Formation in Chinese Jujube
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Suda, J.; Krahulcova, A.; Travnicek, P.; Rosenbaumova, R.; Peckert, T.; Krahulec, F. Genome size variation and species relationships in Hieracium sub-genus Pilosella (Asteraceae) as inferred by flow cytometry. Ann. Bot. 2007, 100, 1323–1335. [Google Scholar] [CrossRef]
- Yan, H.; Martin, S.L.; Bekele, W.A.; Latta, R.G.; Diederichsen, A.; Peng, Y.; Tinker, N.A. Genome size variation in the genus Avena. Genome 2016, 59, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Suda, J.; Meyerson, L.A.; Leitch, I.J.; Pysek, P. The hidden side of plant invasions: the role of genome size. New Phytol. 2015, 205, 994–1007. [Google Scholar] [CrossRef]
- Pellicer, J.; Hidalgo, O.; Dodsworth, S.; Leitch, I.J. Genome size diversity and its impact on the evolution of land plants. Genes 2018, 9, 88. [Google Scholar] [CrossRef]
- Liu, Z.X.; Guo, S.; Xu, J.; Zhang, Y.J.; Dong, L.L.; Xiao, S.M.; Bai, R.; Liao, B.S.; Su, H.; Cheng, R.Y. Genome size estimation of Chinese cultured Artemisia annua L. Plant Biol. Crop. Res. 2018, 1, 1002. [Google Scholar] [CrossRef]
- Leitch, I.J.; Kahandawala, I.; Suda, J.; Hanson, L.; Ingrouille, M.J.; Chase, M.W.; Fay, M.F. Genome size diversity in orchids: consequences and evolution. Ann. Bot. 2009, 104, 469–481. [Google Scholar] [CrossRef]
- Frajman, B.; Resetnik, I.; Weiss-Schneeweiss, H.; Ehrendorfer, F.; Schonswetter, P. Cytotype diversity and genome size variation in Knautia (Caprifoliaceae, Dipsacoideae). BMC Evol. Biol. 2015, 15, 140. [Google Scholar] [CrossRef]
- Guignard, M.S.; Nichols, R.A.; Knell, R.J.; Macdonald, A.; Romila, C.A.; Trimmer, M.; Leitch, I.J.; Leitch, A.R. Genome size and ploidy influence angiosperm species biomass under nitrogen and phosphorus limitation. New Phytol. 2016, 210, 1195–1206. [Google Scholar] [CrossRef]
- Dolezel, J.; Bartos, J. Plant DNA flow cytometry and estimation of nuclear genome size. Ann. Bot. 2005, 95, 99–110. [Google Scholar] [CrossRef] [PubMed]
- A, R.; G, L.; A, K. In vitro tetraploid induction and generation of tetraploids from mixoploids in hop (Humulus lupulus L.). Plant Cell Rep. 2001, 20, 489–495. [Google Scholar] [CrossRef]
- Nadler, J.D.; Pooler, M.; Olsen, R.T.; Coleman, G.D. In vitro induction of polyploidy in Cercis glabra Pamp. Sci. Hort. 2012, 148, 126–130. [Google Scholar] [CrossRef]
- Podwyszyńska, M.; Gabryszewska, E.; Dyki, B.; Stępowska, A.A.; Kowalski, A.; Jasiński, A. Phenotypic and genome size changes (variation) in synthetic tetraploids of daylily (Hemerocallis) in relation to their diploid counterparts. Euphytica 2014, 203, 1–16. [Google Scholar] [CrossRef]
- Li, Q.Q.; Zhang, J.; Liu, J.H.; Yu, B.Y. Morphological and chemical studies of artificial Andrographis paniculata polyploids. Chin. J. Nat. Med. 2018, 16, 81–89. [Google Scholar] [CrossRef]
- Ebrahimi, S.; Mollaei, H.; Hoshyar, R. Ziziphus Jujuba: a review study of its anticancer effects in various tumor models invitro and in vivo. Cell. Mol. Biol. 2017, 63, 122–127. [Google Scholar] [CrossRef]
- Shin, M.; Lee, B.M.; Kim, O.; Tran, H.N.K.; Lee, S.; Cheol, H.; Min, B.S.; Lee, J.-H. Triterpenoids from Ziziphus jujuba induce apoptotic cell death in human cancer cells through mitochondrial reactive oxygen species production. Food Funct. 2018, 8, 3895–3905. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.H.; Wu, C.S.; Wang, M. The jujube (Ziziphus jujuba Mill.) fruit: A review of current knowledge of fruit composition and health benefits. J. Agric. Food Chem. 2013, 61, 3351–3363. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.H.; Liu, P.; Liu, M.J.; Wang, J.R.; Xu, J. A novel method for rapid in vivo induction of homogeneous polyploids via calluses in a woody fruit tree (Ziziphus jujuba Mill.). Plant Cell Tiss Organ Cult. 2015, 121, 423–433. [Google Scholar] [CrossRef]
- Zhang, C.M.; Yin, X.; Li, X.G.; Huang, J.; Wang, C.Z.; Lian, C.L. Genetic diversity of sour jujube along the Yellow River. J. Northwest A F Univ.-Nat. Sci. Ed. 2013, 12, 107–112. [Google Scholar] [CrossRef]
- Liu, M.J.; Wang, M. Germplasm Resources of Chinese Jujube; China Forestry Publishing House: Beijing, China, 2009; ISBN 7503857420. [Google Scholar]
- Wang, L.H.; Lv, Y.; Luo, Z.; Liu, P.; Liu, M.J. Establishment and application of a method for chromosome ploidy identification and genome size estimation using flow cytometry in Ziziphus jujuba. J. Agri. Biot. 2018, 26, 511–520. [Google Scholar] [CrossRef]
- Liu, M.J.; Zhao, J.; Cai, Q.L.; Liu, G.C.; Wang, J.R.; Zhao, Z.H.; Liu, P.; Dai, L.; Yan, G.; Wang, W.J.; et al. The complex jujube genome provides insights into fruit tree biology. Nat. Commun. 2014, 5, 5315. [Google Scholar] [CrossRef] [Green Version]
- Zhao, A.L.; Wang, Y.K.; Sui, C.L.; Ren, H.Y.; Li, D.K. The sugars and organic acids composition in fruits of different Chinese Jujube cultivars of different development stages. Acta Hort. Sin. 2016, 43, 1175–1185. [Google Scholar] [CrossRef]
- Wu, L.P.; Tang, Y.; Li, Y.Y.; Yin, D.N.; Pang, X.M. Estimation of genome size of Ziziphus jujuba and Z.acdiojujuba. J. Beijing For. Univ. 2013, 35, 77–83. [Google Scholar] [CrossRef]
- Shen, H.; Huang, J.; Tong, Y.; LI, X.G. Comparative analysis of genome sizes of Chinese jujube. J. Northwest For. Univ. 2016, 31, 138–142. [Google Scholar] [CrossRef]
- Li, S.Y.; Xu, J.R.; Sun, H.Y. RAPD marker analysis of dongzao jujube in different sites. J. Northwest For. Univ. 2006, 21, 89–93. [Google Scholar] [CrossRef]
- Huang, J.; Zhang, C.; Zhao, X.; Fei, Z.; Wan, K.; Zhang, Z.; Pang, X.; Yin, X.; Bai, Y.; Sun, X. The jujube genome provides insights into genome evolution and the domestication of sweetness/acidity taste in fruit trees. PLoS Genet. 2016, 12, e1006433. [Google Scholar] [CrossRef] [PubMed]
- Steven, D.; Mark, W.C.; Andrew, R.L. Is post-polyploidization diploidization the key to the evolutionary success of angiosperms? Bot. J. Linn Soc. 2016, 180, 1–5. [Google Scholar] [CrossRef]
- Wendel, J.F.; Lisch, D.; Hu, G.; Mason, A.S. The long and short of doubling down: polyploidy, epigenetics, and the temporal dynamics of genome fractionation. Curr. Opin. Genet. Dev. 2018, 49, 1–7. [Google Scholar] [CrossRef]
- Terezie, M.; Zheng, L.; Michael, S.B.; Martin, A.L. Diverse genome organization following 13 independent mesopolyploid events in Brassicaceae contrasts with convergent patterns of gene retention. Plant J. 2017, 91, 3–21. [Google Scholar] [CrossRef] [Green Version]
- Riet, D.S.; Keith, L.A.; Klaas, V.; Marc, C.E.; Van, M.; Steven, M.; Yves, V.P. Convergent gene loss following gene and genome duplications creates single-copy families in flowering plants. Proc Natl Acad Sci. 2011, 108, 4069–4074. [Google Scholar] [CrossRef]
- Mandakova, T.; Lysak, M.A. Post-polyploid diploidization and diversification through dysploid changes. Curr Opin Plant Biol. 2018, 42, 55–65. [Google Scholar] [CrossRef]
- Jordan, G.J.; Carpenter, R.J.; Koutoulis, A.; Price, A.; Brodribb, T.J. Environmental adaptation in stomatal size independent of the effects of genome size. New Phytol. 2015, 205, 608–617. [Google Scholar] [CrossRef]
- Lee, C.E. Evolutionary genetics of invasive species. Trends Ecol. Evol. 2002, 17, 386–391. [Google Scholar] [CrossRef]
- Pandit, M.K.; White, S.M.; Pocock, M.J. The contrasting effects of genome size, chromosome number and ploidy level on plant invasiveness: a global analysis. New Phytol. 2014, 203, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Martinez, M.A.; Baack, E.J.; Hovick, S.M.; Whitney, K.D. A reassessment of the genome size-invasiveness relationship in reed canarygrass (Phalaris arundinacea). Ann. Bot. 2018, 121, 1309–1318. [Google Scholar] [CrossRef]
- Cavalier-Smith, T. Economy, speed and size matter: evolutionary forces driving nuclear genome miniaturization and expansion. Ann. Bot. 2005, 95, 147–175. [Google Scholar] [CrossRef]
- Clark, J.; Hidalgo, O.; Pellicer, J.; Liu, H.; Marquardt, J.; Robert, Y.; Christenhusz, M.; Zhang, S.; Gibby, M.; Leitch, I.J.; et al. Genome evolution of ferns: evidence for relative stasis of genome size across the fern phylogeny. New Phytol. 2016, 210, 1072–1082. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Tong, Y.; Zhang, Q.J.; Gao, L.Z. Genome size variation among and within Camellia species by using flow cytometric analysis. PLoS One 2013, 8, e64981. [Google Scholar] [CrossRef]
- Doerks, T.; Copley, R.R.; Schultz, J.; Ponting, C.P.; Bork, P. Systematic identification of novel protein domain families associated with nuclear functions. Genome Res. 2002, 12, 47–56. [Google Scholar] [CrossRef]
- Chu, Z.F.; Wen, J.; Yang, Y.P.; Nie, Z.L.; Meng, Y. Genome size variation and evolution in the grape family Vitaceae. J. Syst. Evol. 2018, 56, 273–282. [Google Scholar] [CrossRef] [Green Version]
- Qin, C.; Yu, C.; Shen, Y.; Fang, X.; Chen, L.; Min, J.; Cheng, J.; Zhao, S.; Xu, M.; Luo, Y.; et al. Whole-genome sequencing of cultivated and wild peppers provides insights into Capsicum domestication and specialization. PANS 2014, 111, 5135–5140. [Google Scholar] [CrossRef]
- Loureiro, J.; Rodriguez, E.; Costa, A.; Santos, C. Nuclear DNA content estimations in wild olive (Olea europaea L. ssp. europaea var. sylvestris Brot.) and Portuguese cultivars of O. europaea using flow cytometry. Genet. Resour. Crop Evol. 2006, 54, 21–25. [Google Scholar] [CrossRef]
- Mei, W.; Stetter, M.G.; Gates, D.J.; Stitzer, M.C.; Ross-Ibarra, J. Adaptation in plant genomes: bigger is different. Am. J. Bot. 2018, 105, 16–19. [Google Scholar] [CrossRef]
- Simonin, K.A.; Roddy, A.B. Genome downsizing, physiological novelty, and the global dominance of flowering plants. PLoS Biol. 2018, 16, e2003706. [Google Scholar] [CrossRef]
- Marfil, C.F.; Duarte, P.F.; Masuelli, R.W. Phenotypic and epigenetic variation induced in newly synthesized allopolyploids and autopolyploids of potato. Sci. Hort. 2018, 234, 101–109. [Google Scholar] [CrossRef]
- Qiao, X.; Yin, H.; Li, L.; Wang, R.; Wu, J.; Wu, J.; Zhang, S. Different Modes of Gene Duplication Show Divergent Evolutionary Patterns and Contribute Differently to the Expansion of Gene Families Involved in Important Fruit Traits in Pear (Pyrus bretschneideri). Front. Plant Sci. 2018, 9, 161. [Google Scholar] [CrossRef]
- Guo, S.; Zhang, J.; Sun, H.; Salse, J.; Lucas, W.J.; Zhang, H.; Zheng, Y.; Mao, L.; Ren, Y.; Wang, Z.; et al. The draft genome of watermelon (Citrullus lanatus) and resequencing of 20 diverse accessions. Nat. Genet. 2013, 45, 51–58. [Google Scholar] [CrossRef]
- Sutherland, B.L.; Quarles, B.M.; Galloway, L.F. Intercontinental dispersal and whole-genome duplication contribute to loss of self-incompatibility in a polyploid complex. Am. J. Bot. 2018, 105, 249–256. [Google Scholar] [CrossRef]
- Wu, J.; Wang, Z.; Shi, Z.; Zhang, S.; Ming, R.; Zhu, S.; Khan, M.A.; Tao, S.; Korban, S.S.; Wang, H.; et al. The genome of the pear (Pyrus bretschneideri Rehd.). Genome Res. 2013, 23, 396–408. [Google Scholar] [CrossRef]
- Parisod, C.; Holderegger, R.; Brochmann, C. Evolutionary consequences of autopolyploidy. New Phytol. 2010, 186, 5–17. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar/Species | Relative Fluorescence Density | Coefficient of Variation (%) | Predicted Genome Size (Mb) |
|---|---|---|---|
| Populus trichocarpa | 3238.96 ± 96.89 | 3.25 | 480 (reference) |
| Dongzao | 2997.07 ± 50.44 | 4.03 | 444.27 ± 6.31 |
| Genotypes | Genome Size (Mb) | Genotypes | Genome Size (Mb) | Genotypes | Genome Size (Mb) | |||
|---|---|---|---|---|---|---|---|---|
| Mean | SD | Mean | SD | Mean | SD | |||
| Xingtai0608 | 346.93 | 9.60 | Suanzao28 | 410.09 | 4.27 | ZiA | 434.95 | 21.93 |
| Suanzao10 | 360.30 | 3.99 | Xingtai0604 | 412.21 | 1.12 | Xingtai0614 | 436.24 | 4.22 |
| Suanzao37 | 360.78 | 8.90 | Xingtai0613 | 412.34 | 2.90 | ZiU | 439.00 | 4.08 |
| Xingtai0601 | 374.83 | 4.05 | Suanzao49 | 416.26 | 6.53 | Zi16 | 439.11 | 7.47 |
| Zi2 | 380.06 | 4.98 | Zi3 | 416.38 | 7.87 | Beikedi2 | 439.36 | 5.26 |
| Suanzao36 | 380.06 | 5.42 | Xingtai0634 | 419.24 | 3.28 | Xingtai10 | 439.84 | 17.91 |
| Xingtai0605 | 380.56 | 0.12 | ZiT | 419.58 | 5.44 | Xingtai0629 | 439.88 | 7.46 |
| Suanzao17 | 383.26 | 11.49 | Xingtai6 | 420.33 | 2.02 | Zi25 | 441.62 | 5.23 |
| Xingtai0619 | 383.70 | 1.34 | Zi4 | 421.39 | 4.69 | Taigudasuanzao | 441.63 | 8.73 |
| ZiY | 383.97 | 9.23 | Xianxiansuanzao | 422.16 | 6.41 | Suanzao40 | 443.34 | 1.76 |
| Xingtai0602 | 384.15 | 5.64 | ZiZ | 422.34 | 12.72 | Beikedi1 | 447.27 | 4.57 |
| Xingtai0618 | 385.63 | 4.80 | Zi23 | 423.55 | 5.87 | GaoVcsuanzao | 451.89 | 4.97 |
| Suanzao21 | 389.51 | 4.09 | ZiX | 424.14 | 16.42 | Suanzao11 | 456.24 | 7.06 |
| Beikedi9 | 394.09 | 3.73 | Suanzao32 | 424.18 | 3.94 | Suanzao3 | 457.33 | 1.59 |
| Zi28 | 395.12 | 1.41 | Suanzao4 | 424.96 | 7.30 | Xingtai17 | 458.94 | 3.86 |
| Suanzao27 | 398.72 | 3.52 | Suanzao45 | 426.95 | 0.80 | Suanzao6 | 458.98 | 0.39 |
| Suanzao41 | 398.91 | 2.63 | Suanzao30 | 427.61 | 1.71 | Suanzao1 | 459.11 | 3.32 |
| Jiaochengtiansuanzao | 398.93 | 3.67 | Suanzao44 | 428.25 | 9.48 | Suanzaowang | 459.47 | 8.66 |
| Xingtai0603 | 398.95 | 2.65 | Suanzao9 | 428.43 | 2.52 | Beiqi6 | 459.67 | 6.00 |
| ZiO | 400.97 | 3.77 | Wumingsuanzao | 429.79 | 0.68 | Xingtai27 | 465.21 | 6.91 |
| Zi24 | 403.92 | 9.60 | Suanzao12 | 430.30 | 1.83 | Suanzao20 | 466.38 | 6.59 |
| Zi15 | 405.53 | 9.89 | ZiV | 430.66 | 2.54 | Suanzao24 | 474.38 | 3.47 |
| Xingtai0648 | 406.17 | 3.94 | Suanzao8 | 431.31 | 2.82 | Xingtai0609 | 479.91 | 7.97 |
| Suanzao23 | 406.63 | 8.65 | Suanzao34 | 431.79 | 2.55 | Xingtai16 | 482.34 | 4.22 |
| Beikedi3 | 406.69 | 3.94 | Suanzao14 | 431.87 | 2.78 | Suanzao25 | 483.04 | 7.81 |
| ZiH | 407.28 | 5.05 | Suanzao47 | 432.51 | 1.86 | Suanzao43 | 486.04 | 4.89 |
| Suanzao31 | 408.87 | 3.12 | Suanzao48 | 433.84 | 5.36 | Xingtai0610 | 489.44 | 1.29 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Luo, Z.; Liu, Z.; Zhao, J.; Deng, W.; Wei, H.; Liu, P.; Liu, M. Genome Size Variation within Species of Chinese Jujube (Ziziphus jujuba Mill.) and Its Wild Ancestor Sour Jujube (Z. acidojujuba Cheng et Liu). Forests 2019, 10, 460. https://doi.org/10.3390/f10050460
Wang L, Luo Z, Liu Z, Zhao J, Deng W, Wei H, Liu P, Liu M. Genome Size Variation within Species of Chinese Jujube (Ziziphus jujuba Mill.) and Its Wild Ancestor Sour Jujube (Z. acidojujuba Cheng et Liu). Forests. 2019; 10(5):460. https://doi.org/10.3390/f10050460
Chicago/Turabian StyleWang, Lihu, Zhi Luo, Zhiguo Liu, Jin Zhao, Wenping Deng, Hairong Wei, Ping Liu, and Mengjun Liu. 2019. "Genome Size Variation within Species of Chinese Jujube (Ziziphus jujuba Mill.) and Its Wild Ancestor Sour Jujube (Z. acidojujuba Cheng et Liu)" Forests 10, no. 5: 460. https://doi.org/10.3390/f10050460
APA StyleWang, L., Luo, Z., Liu, Z., Zhao, J., Deng, W., Wei, H., Liu, P., & Liu, M. (2019). Genome Size Variation within Species of Chinese Jujube (Ziziphus jujuba Mill.) and Its Wild Ancestor Sour Jujube (Z. acidojujuba Cheng et Liu). Forests, 10(5), 460. https://doi.org/10.3390/f10050460