Predicting the Bioclimatic Habitat Suitability of Ginkgo biloba L. in China with Field-Test Validations



Abstract
:1. Introduction
2. Materials and Methods
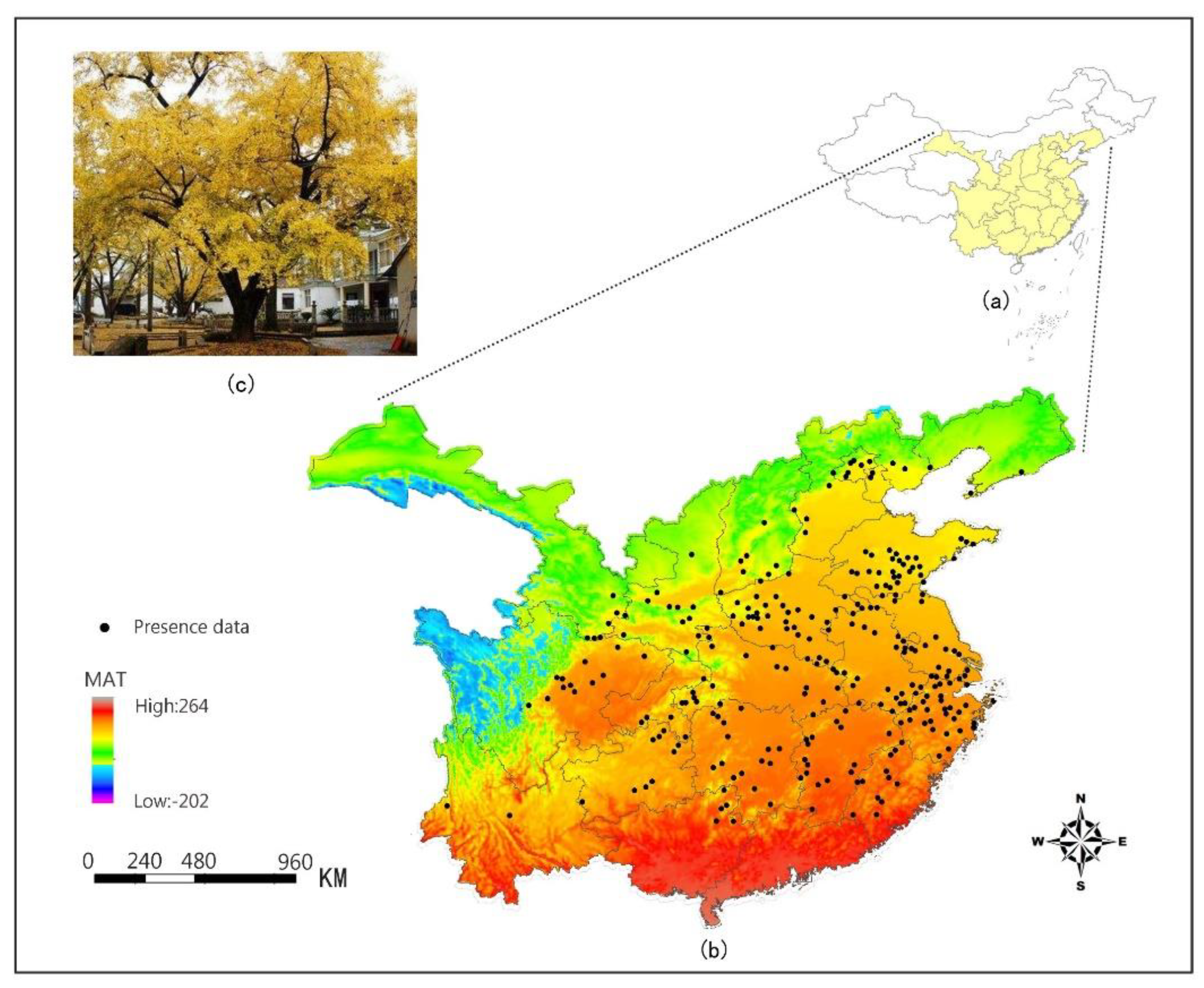
2.1. Occurrence Data
2.2. Climate Data
2.3. Model Development
2.4. Model Validation Test
3. Results
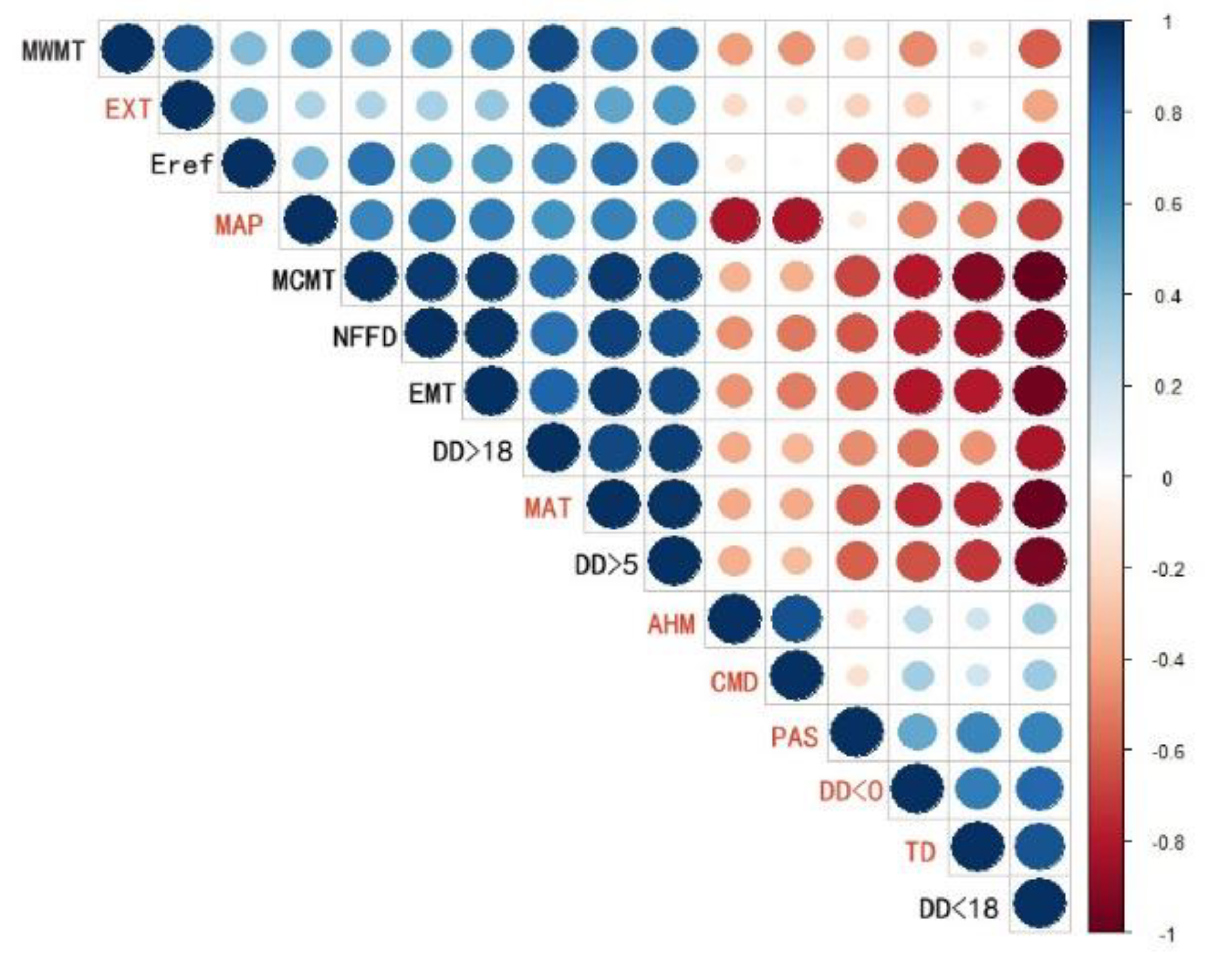
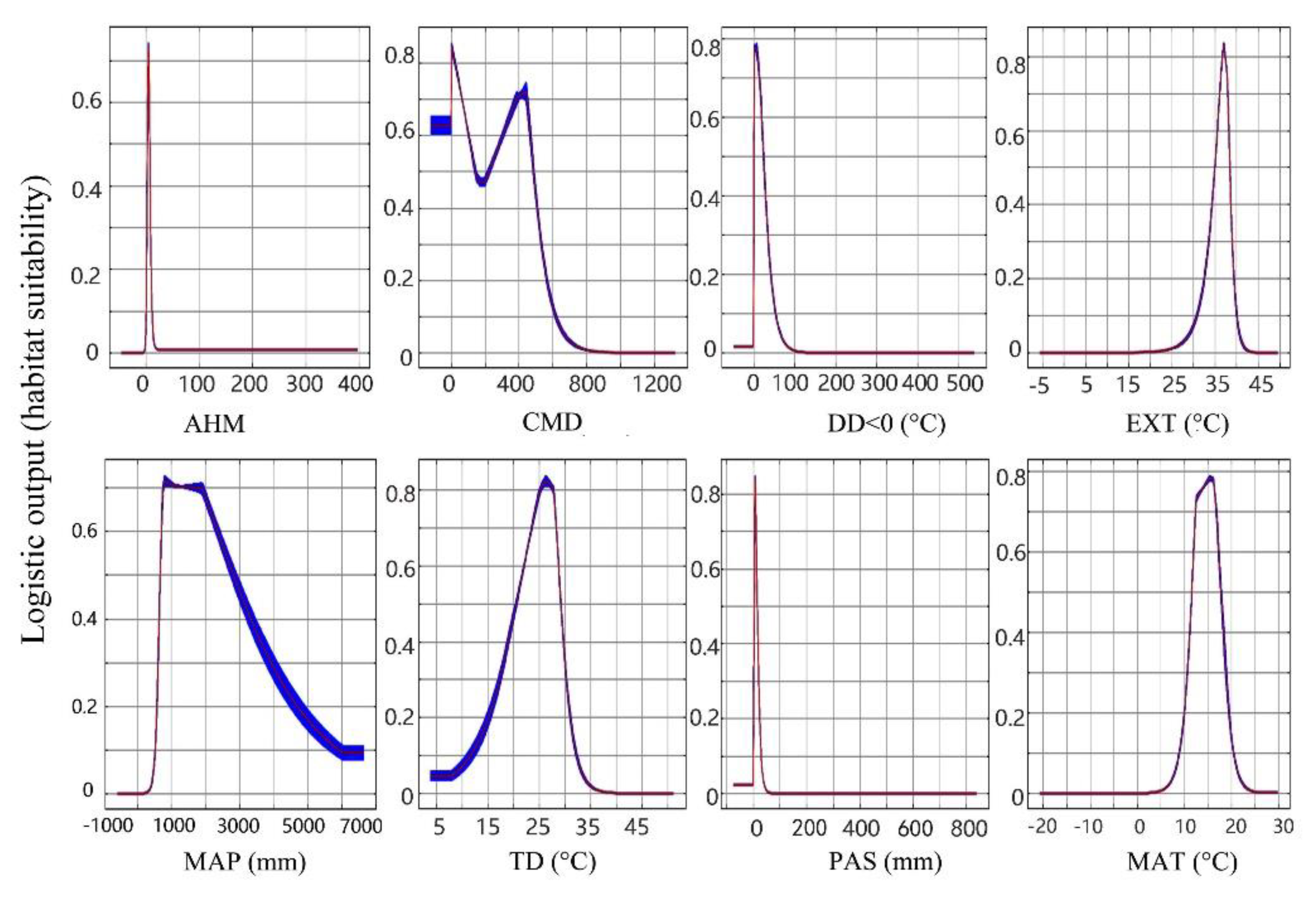
3.1. Model Performance and Climate Variables’ Contribution
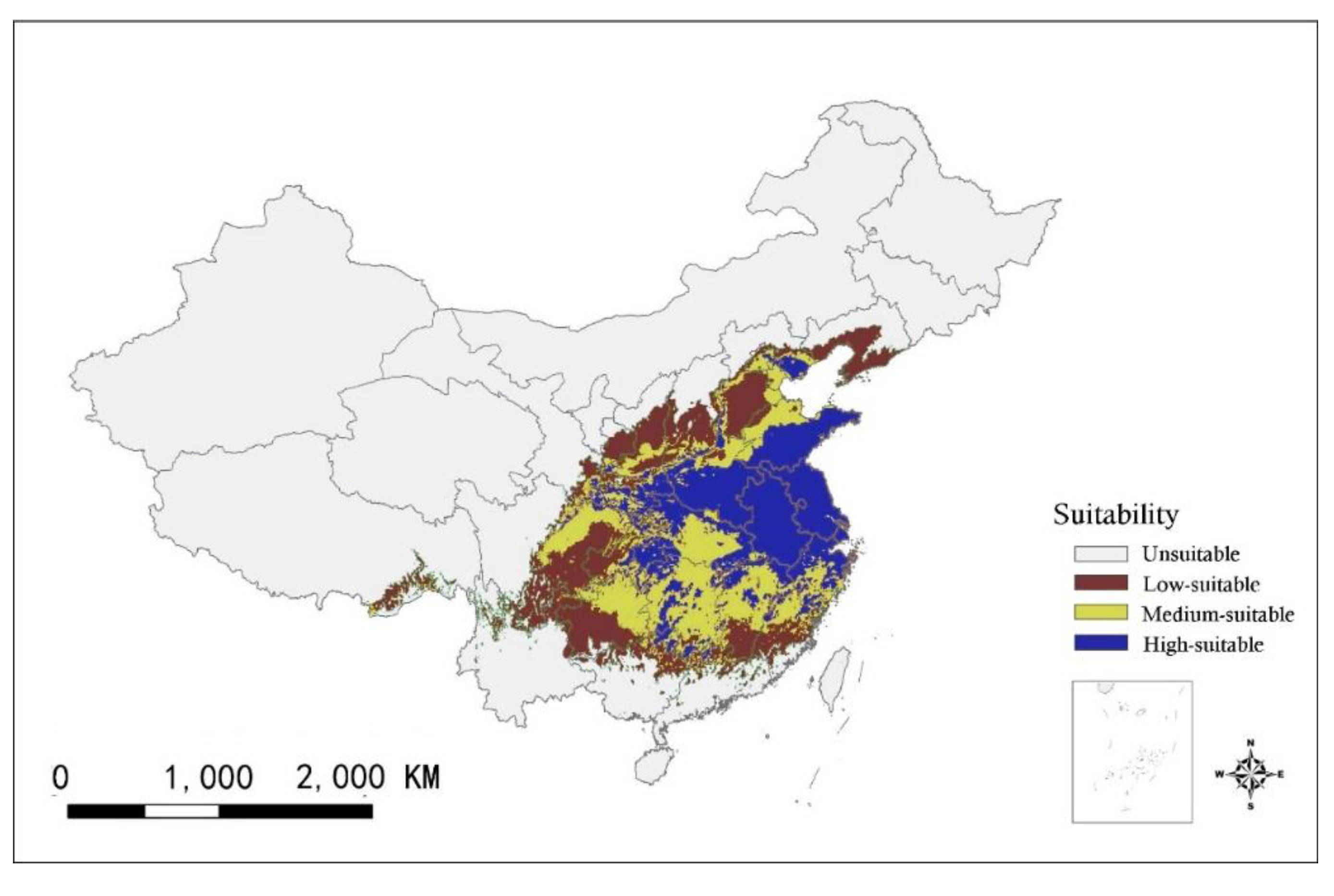
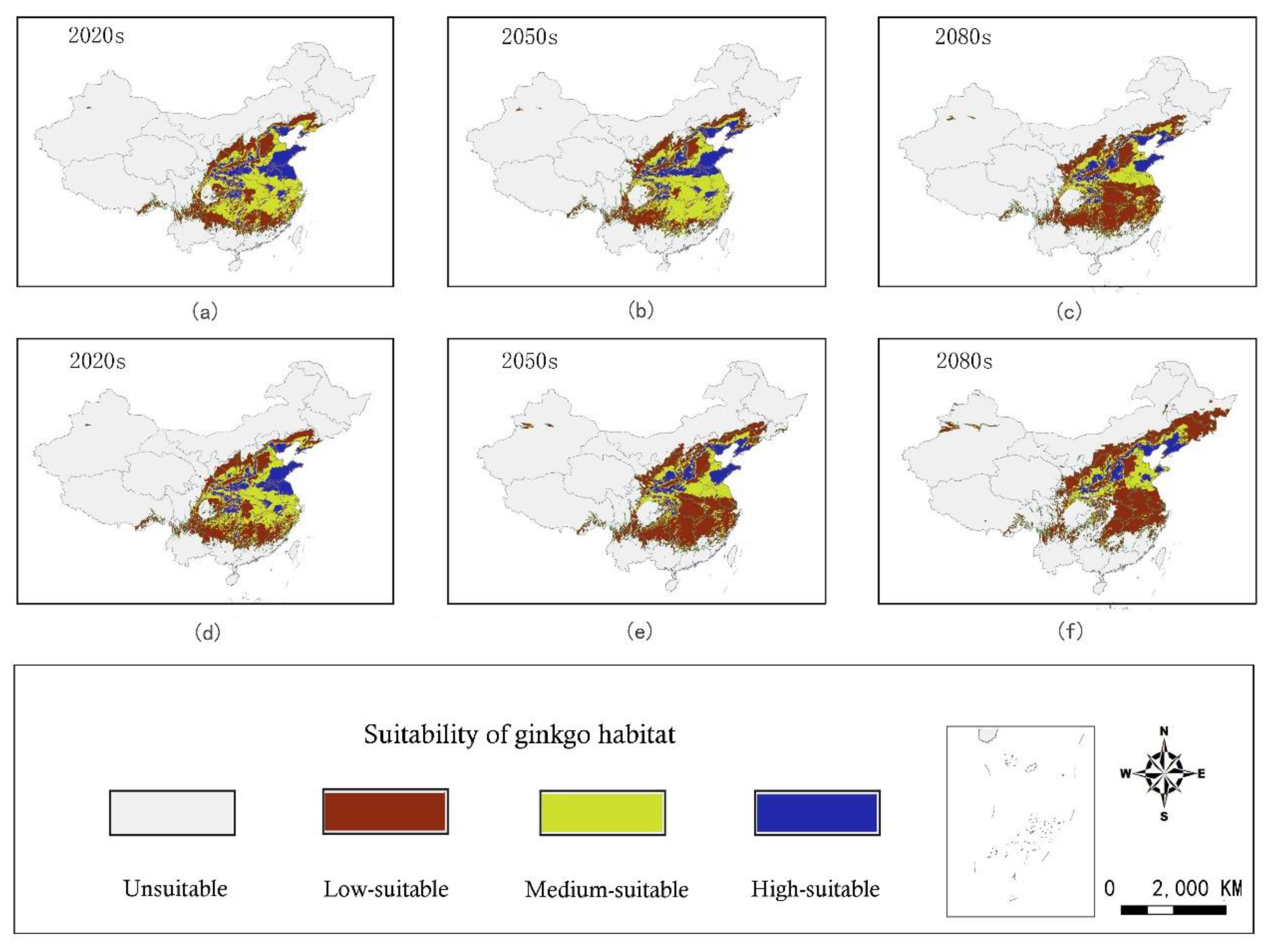
3.2. Distribution of Current Habitat Suitability
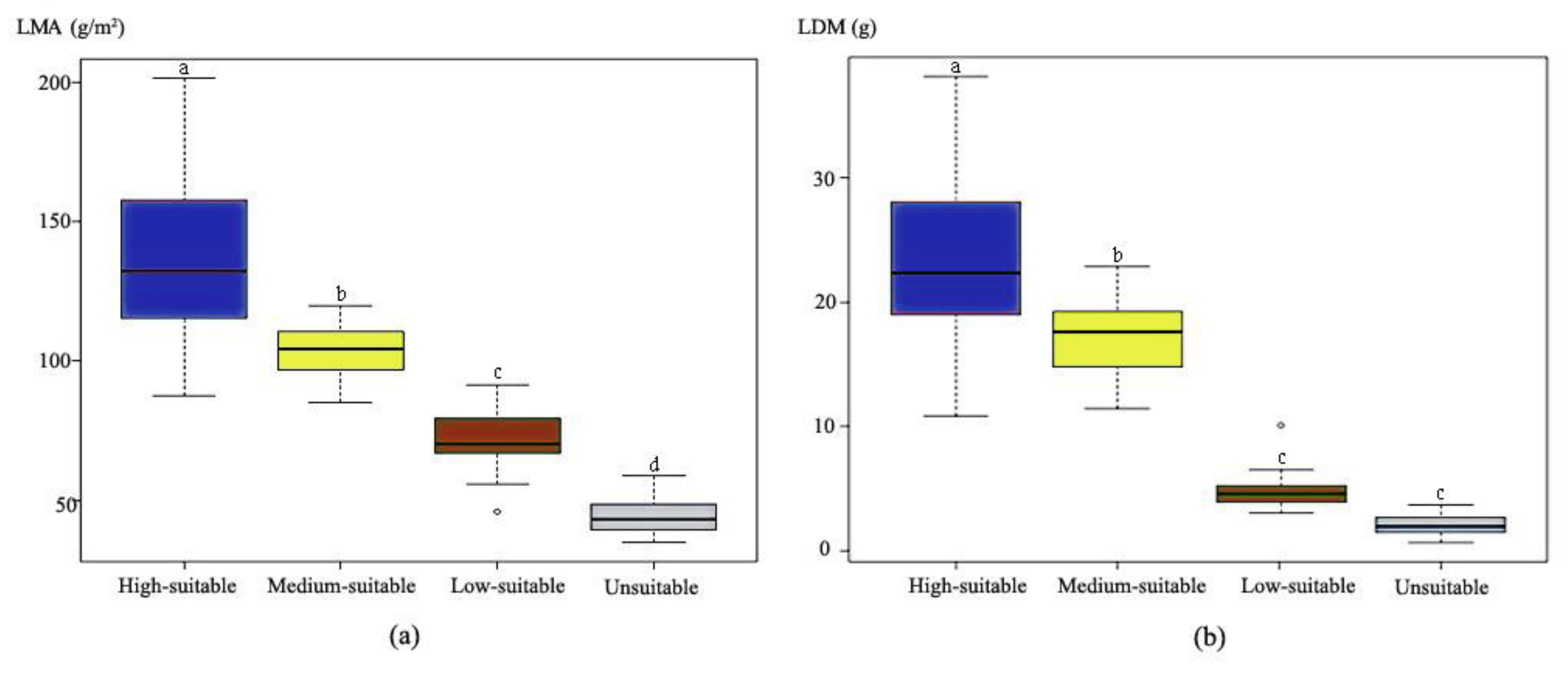
3.3. Validation of the Bioclimatic Model
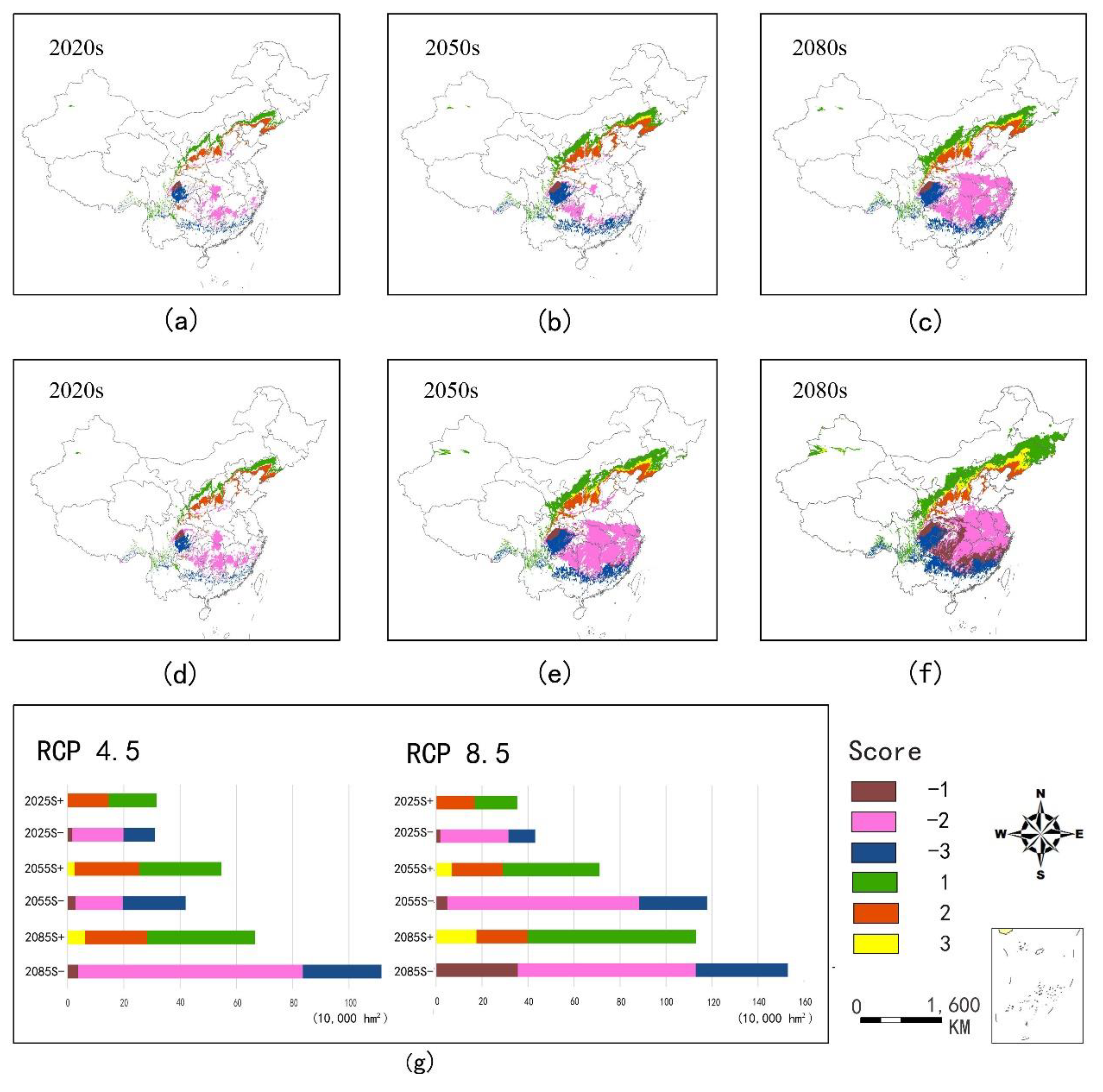
3.4. Distribution of Future Habitat Suitability
4. Discussion
4.1. Key Climate Factors Determining the Ginkgo Distribution
4.2. Distributions of Bioclimatic Habitat Categories
4.3. Impacts of Climate Change on the Habitat Suitability of Ginkgo in the Future
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Fu, L.K.; Jin, J.M. China Plant Red Data Book—Rare and Endangered Plants; Science Press: Beijing, China, 1992. (in Chinese) [Google Scholar]
- Mckeage, K.; Lyseng-Williamson, K.A. Ginkgo biloba extract egb 761 ® in the symptomatic treatment of mild-to-moderate dementia: A profile of its use. Drugs Ther. Perspect. 2018, 34, 358–366. [Google Scholar] [CrossRef] [PubMed]
- Qi, Z.; Mu, K.; Meng, X.; Ma, X.; Ni, Z.; Wang, J.; Xu, L.A. Variation in the concentrations of major secondary metabolites in ginkgo leaves from different geographical populations. Forests 2017, 8, 266–280. [Google Scholar]
- Wu, Y.; Guo, J.; Zhou, Q.; Xin, Y.; Xu, L.A. De novo transcriptome analysis revealed genes involved in flavonoid biosynthesis, transport and regulation in ginkgo biloba. Ind. Crop. Prod. 2018, 124, 226–235. [Google Scholar] [CrossRef]
- Liu, K.; Shen, X.; Cao, L.; Wang, G.; Cao, F. Estimating forest structural attributes using uav-lidar data in ginkgo plantations. ISPRS J. Photogramm. Remote Sens. 2018, 164, 465–482. [Google Scholar] [CrossRef]
- Cao, F.L. Chinese Ginkgo; Jiangsu Science and Technology Press: Hong Kong, China, 2002. (in Chinese) [Google Scholar]
- Deb, J.C.; Phinn, S.; Butt, N.; Mcalpine, C.A. The impact of climate change on the distribution of two threatened dipterocarp trees. Ecol. Evol. 2017, 7, 2238–2248. [Google Scholar] [CrossRef] [PubMed]
- Thuiller, W.; Lavorel, S.; Sykes, M.T.; Araújo, M.B. Using niche-based modelling to assess the impact of climate change on tree functional diversity in europe. Divers. Distrib. 2010, 12, 49–60. [Google Scholar] [CrossRef]
- Mckenney, D.W.; Pedlar, J.H.; Lawrence, K.; Campbell, K.; Hutchinson, M.F. Potential impacts of climate change on the distribution of north american trees. Bioscience 2007, 57, 939–948. [Google Scholar] [CrossRef]
- Diffenbaugh, N.S.; Field, C.B. Changes in ecologically critical terrestrial climate conditions. Science 2013, 341, 486–492. [Google Scholar] [CrossRef]
- Craig, M.; Rosa, A. The future of species under climate change: Resilience or decline? Science 2013, 341, 504–508. [Google Scholar]
- Liñán, I.D.; Heinrich, I.; Andreu-Hayles, L.; Muntán, E.; Campelo, F.; Helle, G. Age effects and climate response in trees: A multi-proxy tree-ring test in old-growth life stages. Eur. J. For. Res. 2012, 131, 933–944. [Google Scholar] [CrossRef]
- IPCC. Climate Change 2014: The Physical Science Basis. Contribution of Working Group i to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014. [Google Scholar]
- Aitken, S.N.; Yeaman, S.; Holliday, J.A.; Wang, T.; Curtis-Mclane, S. Adaptation, migration or extirpation: Climate change outcomes for tree populations. Evol. Appl. 2010, 1, 95–111. [Google Scholar] [CrossRef] [PubMed]
- Jump, A.; Penuelas, J. Running to stand still: Adaptation and the response of plants to rapid climate change. Ecol. Lett. 2010, 8, 1010–1020. [Google Scholar] [CrossRef]
- Santamour, F.S.; Ewert, T.E. Growth, survival and sex expression in ginkgo. J. Arboric. 1983, 9, 170–171. [Google Scholar]
- He, S.A.; Gu, Y.; Pang, Z.J. Resources and prospects of ginkgo biloba in china. In Ginkgo Biloba—A Global Treasure; Hori, R.W., Ridge, W., Tulecke, P., Del Tredici, J., Tremouillaux-Guiller, T.H., Eds.; Springer-Verlag: Tokyo, Japan, 1997; pp. 373–383. [Google Scholar]
- Hohmann, N.; Wolf, E.M.; Rigault, P.; Zhou, W.; Kiefer, M.; Zhao, Y.; Fu, C.X.; Koch, M.A. Ginkgo biloba’s footprint of dynamic pleistocene history dates back only 390,000 years ago. BMC Genom. 2018, 19, 299–316. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Sun, L.; Wu, Q.; Men, X.; Yao, L.; Xing, S. Transcriptome profile analysis reveals the ontogenesis of rooted chichi in ginkgo biloba l. Gene 2018, 669, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Peterson, A.T.; Soberón, J.; Pearson, R.G.; Anderson, R.P.; Martínez-Meyer, E.; Nakamura, M.; Araújo, M.B. Ecological Niches and Geographic Distributions (Mpb-49); Princeton University Press: Princeton, NJ, USA, 2011. [Google Scholar]
- Berry, P.M.; Dawson, T.P.; Harrison, P.A.; Pearson, R.G. Modelling potential impacts of climate change on the bioclimatic envelope of species in britain and ireland. Glob. Ecol. Biogeogr. 2010, 11, 453–462. [Google Scholar] [CrossRef]
- Pearson, R.G.; Dawson, T.P.; Berry, P.M.; Harrison, P.A. Species: A spatial evaluation of climate impact on the envelope of species. Ecol. Model. 2002, 154, 289–300. [Google Scholar] [CrossRef]
- Qin, A.; Liu, B.; Guo, Q.; Bussmann, R.W.; Ma, F.; Jian, Z.; Xu, G.; Pei, S. Maxent modeling for predicting impacts of climate change on the potential distribution of thuja sutchuenensis franch., an extremely endangered conifer from southwestern china. Glob. Ecol. Conserv. 2017, 10, 139–146. [Google Scholar] [CrossRef]
- Yu, J.; Wang, C.; Wan, J.; Han, S.; Wang, Q.; Nie, S. A model-based method to evaluate the ability of nature reserves to protect endangered tree species in the context of climate change. For. Ecol. Manag. 2014, 327, 48–54. [Google Scholar] [CrossRef]
- Dudík, M.; Phillips, S.J.; Schapire, R.E. Performance guarantees for regularized maximum entropy density estimation. Lect. Notes Comput. Sci. 2004, 3120, 472–486. [Google Scholar]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of maxent for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Pearson, R.G.; Raxworthy, C.J.; Nakamura, M.; Peterson, A.T. Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in madagascar. J. Biogeogr. 2010, 34, 102–117. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Yi, Y.J.; Cheng, X.; Yang, Z.F.; Zhang, S.H. Maxent modeling for predicting the potential distribution of endangered medicinal plant (h. Riparia lour) in yunnan, china. Ecol. Eng. 2016, 92, 260–269. [Google Scholar] [CrossRef]
- Yang, X.Q.; Kushwaha, S.P.S.; Saran, S.; XU, J.; ROY, P.S. Maxent modeling for predicting the potential distribution of medicinal plant, justicia adhatoda l. In lesser himalayan foothills. Reg. Environ. Chang. 2013, 51, 83–87. [Google Scholar] [CrossRef]
- Chemura, A.; Kutywayo, D.; Chidoko, P.; Mahoya, C. Bioclimatic modelling of current and projected climatic suitability of coffee (coffea arabica) production in zimbabwe. Reg. Environ. Change 2016, 16, 1–13. [Google Scholar] [CrossRef]
- Pearson, R.G.; Dawson, T.P. Predicting the impacts of climate change on the distribution of species: Are bioclimate envelope models useful? Glob. Ecol. Biogeogr. 2003, 12, 361–371. [Google Scholar] [CrossRef]
- Nicotra, A.B.; Atkin, O.K.; Bonser, S.P.; Davidson, A.M.; Finnegan, E.J.; Mathesius, U.; Poot, P.; Purugganan, M.D.; Richards, C.L.; Valladares, F. Plant phenotypic plasticity in a changing climate. Trends Plant Sci. 2010, 15, 684–692. [Google Scholar] [CrossRef] [PubMed]
- Vendramini, F.; Díaz, S.; Gurvich, D.E.; Wilson, P.J.; Thompson, K.; Hodgson, J.G. Leaf traits as indicators of resource-use strategy in flora with succulent species. New Phytol. 2010, 154, 147–157. [Google Scholar] [CrossRef]
- Westoby, M. A leaf-height-seed (lhs) plant ecology strategy scheme. Plant Soil 1998, 199, 213–227. [Google Scholar] [CrossRef]
- Poorter, H.; Niinemets, U.; Poorter, H.; Wright, I.J.; Rafael, V. Causes and consequences of variation in leaf mass per area (lma): A meta-analysis. New Phytol. 2010, 182, 565–588. [Google Scholar] [CrossRef]
- Soudzilovskaia, N.A.; Elumeeva, T.G.; Onipchenko, V.G.; Shidakov, I.I.; Salpagarova, F.S.; Khubiev, A.B.; Tekeev, D.K.; Cornelissen, J.H.C. Functional traits predict relationship between plant abundance dynamic and long-term climate warming. Proc. Natl. Acad. Sci. USA 2013, 110, 18180–18184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, W.; Chai, H.; Lin, P.H.; Lumsden, A.B.; Yao, Q.; Chen, C. Clinical use and molecular mechanisms of action of extract of ginkgo biloba leaves in cardiovascular diseases. Cardiovasc. Drug Rev. 2010, 22, 309–319. [Google Scholar] [CrossRef]
- Radosavljevic, A.; Anderson, R.P. Making better maxent models of species distributions: Complexity, overfitting and evaluation. J. Biogeogr. 2014, 41, 629–643. [Google Scholar] [CrossRef]
- Peterson, A.T.; Papeş, M.; Eaton, M. Transferability and model evaluation in ecological niche modeling: A comparison of garp and maxent. Ecography 2010, 30, 550–560. [Google Scholar] [CrossRef]
- Climate models | Climate Change Adaptation of Asia-Pacific Forests. Available online: http://asiapacific.forestry.ubc.ca/research-approaches/climate-modeling (accessed on 8 October 2018).
- Wang, T.; Wang, G.; Innes, J.L.; Seely, B.; Chen, B. Climateap: An application for dynamic local downscaling of historical and future climate data in asia pacific. Front. Agric. Sci. Eng. 2017, 4, 448–458. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.; Anderson, R.; Dudik, M.; Ferrier, S.; Guisan, A.; Hijmans, R.; Huettmann, F.; Leathwick, J.; Lehmann, A. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2010, 29, 129–151. [Google Scholar] [CrossRef]
- Remya, K.; Ramachandran, A.; Jayakumar, S. Predicting the current and future suitable habitat distribution of myristica dactyloides gaertn. Using maxent model in the eastern ghats, india. Ecol. Eng. 2015, 82, 184–188. [Google Scholar] [CrossRef]
- Kuloba, B.; Gils, H.V.; Duren, I.V.; Ngene, S. Modeling cheetah acinonyx jubatus fundamental niche in kenya. Int. J. Environ. Monit. Anal. 2015, 3, 317–330. [Google Scholar]
- Swets, J.A. Measuring the accuracy of diagnostic systems. Science 1988, 240, 1285–1293. [Google Scholar] [CrossRef]
- Crawford, P.H.C.; Hoagland, B.W. Using species distribution models to guide conservation at the state level: The endangered american burying beetle (nicrophorus americanus) in oklahoma. J. Insect Conserv. 2010, 14, 511–521. [Google Scholar] [CrossRef]
- Beerling, D.J.; Huntley, B.; Bailey, J.P. Climate and the distribution of fallopia japonica: Use of an introduced species to test the predictive capacity of response surfaces. J. Veg. Sci. 1995, 6, 269–282. [Google Scholar] [CrossRef]
- Callaghan, T.; Björn, L.; Chernov, Y.; Chapin, T.; Christensen, T.R.; Huntley, B.; Ims, R.; Johansson, M.; Jolly, D.; Jonasson, S.; et al. Biodiversity, distributions and adaptations of arctic species in the context of environmental change. Ambio 2004, 33, 448–458. [Google Scholar] [CrossRef] [PubMed]
- Julian, C.; Herrero, M.; Rodrigo, J. Anther meiosis time is related to winter cold temperatures in apricot (prunus armeniaca l.). Environ. Exp. Bot. 2014, 100, 20–25. [Google Scholar] [CrossRef]
- Cao, F.L. Chinese Ginkgo Record; China Forestry Publishing House: Beijing, China, 2007. (in Chinese) [Google Scholar]
- Wilson, J.C.; Altland, J.E.; Sibley, J.L.; Tilt, K.M. Effects of chilling and heat on growth of ginkgo biloba l. J. Arboric. 2004, 30, 45–51. [Google Scholar]
- Lambers, H.; Poorter, H. Inherent variation in growth rate between higher plants: A search for physiological causes and ecological consequences. Adv. Ecol. Res. 1992, 23, 187–261. [Google Scholar]
- Shipley, B. Structured interspecific determinants of specificl leaf area in 34 species of herbaceous angiosperms. Funct. Ecol. 1995, 9, 312–319. [Google Scholar] [CrossRef]
- Niinemets, U.; Kull, O.; Tenhunen, J.D. Within-canopy variation in the rate of development of photosynthetic capacity is proportional to integrated quantum flux density in temperate deciduous trees. Plant Cell Environ. 2010, 27, 293–313. [Google Scholar] [CrossRef]
- Sims, D.A.; Seemann, J.R.; Luo, Y. Elevated co2 concentration has independent effects on expansion rates and thickness of soybean leaves across light and nitrogen gradients. J. Exp. Bot. 1998, 49, 583–591. [Google Scholar] [CrossRef]
- Lambers, H.; Chapin, F.; Pons, T. Plant Physiological Ecology; Springer Verlag: Newyork, NY, USA, 2008. [Google Scholar]
- Ball, M.C.; Wolfe, J.; Canny, M.; Hofmann, M.; Nicotra, A.B.; Hughes, D. Space and time dependence of temperature and freezing in evergreen leaves. Funct. Plant Biol. 2002, 29, 1259–1272. [Google Scholar] [CrossRef]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Xuhui, W.; Shilong, P.; Philippe, C.; Pierre, F.; Myneni, R.B.; Peter, C.; Martin, H.; John, M.; Shushi, P.; Tao, W. A two-fold increase of carbon cycle sensitivity to tropical temperature variations. Nature 2014, 506, 212–215. [Google Scholar]
- Cunningham, S.; Read, J. Comparison of temperate and tropical rainforest tree species: Photosynthetic responses to growth temperature. Oecologia 2003, 30, 143–153. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.P.; Michaelsen, J.; Leavitt, S.W.; Still, C.J. Using tree rings to predict the response of tree growth to climate change in the continental united states during the twenty-first century. Earth Interact. 2010, 14, 256–261. [Google Scholar] [CrossRef]
- Boisvert-Marsh, L.; Périé, C.; Blois, S.D. Shifting with climate? Evidence for recent changes in tree species distribution at high latitudes. Ecosphere 2016, 5, 1–33. [Google Scholar] [CrossRef]
- Berger, S.; Walther, G.R.; Pott, R. Bioclimatic limits and range shifts of cold-hardy evergreen broad-leaved species at their northern distributional limit in europe. Phytocoenologia 2007, 37, 523–539. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Full Name (Units) | Code | Full Name (Units) |
|---|---|---|---|
| MAT | Mean annual temperature (°C) | DD > 5 | Degree-days above 5 °C, growing degree-days (°C-days) |
| MWMT | Mean warmest month temperature (°C) | DD < 0 | Degree-days below 0 °C, chilling degree-days (°C-days) |
| MCMT | Mean coldest month temperature (°C) | NFFD | The number of frost-free days (day) |
| TD | Temperature difference between MWMT and MCMT, or continentality (°C) | PAS | Precipitation as snow between August in previous year and July in the current year (mm) |
| MAP | Mean annual precipitation (mm) | EMT | Extreme minimum temperature over 30 years (°C) |
| EXT | Extreme maximum temperature over 30 years (°C) | Eref | Hargreaves reference evaporation |
| AHM | Annual heat moisture index (MAT + 10)/(MAP/1000) | CMD | Hargreaves climatic moisture deficit |
| Suitable Category | Site | Latitude (°N) | Longitude (°E) | Altitude (m) | MAT (°C) | MAP (mm) | N (g/kg) | P (g/kg) | C (g/kg) | K (g/kg) |
|---|---|---|---|---|---|---|---|---|---|---|
| Unsuitable habitat | 1 | 22.49 | 112.50 | 143 | 22.4 | 1955 | 1.37 | 0.50 | 3.31 | 11.05 |
| 2 | 43.41 | 81.11 | 820 | 5.2 | 331 | 1.19 | 0.46 | 2.14 | 9.92 | |
| 3 | 40.59 | 123.53 | 30 | 7.6 | 872 | 0.50 | 0.49 | 1.00 | 14.08 | |
| Low-suitable habitat | 4 | 25.52 | 103.58 | 2160 | 14.1 | 1067 | 1.17 | 0.37 | 1.39 | 22.23 |
| 5 | 25.17 | 110.36 | 247 | 19.5 | 1725 | 1.01 | 0.43 | 2.82 | 24.82 | |
| 6 | 25.89 | 114.52 | 131 | 19.2 | 1486 | 1.25 | 0.45 | 1.25 | 21.08 | |
| Medium-suitable habitat | 7 | 31.97 | 107.43 | 442 | 14.1 | 1205 | 0.87 | 0.50 | 1.25 | 16.85 |
| 8 | 39.34 | 117.91 | 35 | 11.5 | 627 | 1.18 | 0.43 | 1.70 | 14.12 | |
| 9 | 38.32 | 113.96 | 61 | 13.0 | 532 | 0.50 | 0.41 | 1.21 | 12.66 | |
| High-suitable habitat | 10 | 34.21 | 117.58 | 44 | 14.5 | 845 | 0.75 | 0.48 | 1.26 | 18.60 |
| 11 | 32.12 | 120.51 | 10 | 15.4 | 1046 | 0.33 | 0.29 | 1.07 | 19.26 | |
| 12 | 32.90 | 113.38 | 93 | 15.4 | 905 | 1.28 | 0.61 | 2.02 | 23.51 |
| Variable | Percent Contribution (%) | Permutation Importance (%) | Jackknife Test AUC | Appropriate Interval (Logistic Output > 0.5) |
|---|---|---|---|---|
| DD < 0 | 63.5 | 0.8 | 0.886 | 0–25 °C |
| PAS | 12.0 | 4.9 | 0.806 | 0–10 mm |
| MAP | 10.3 | 50.8 | 0.804 | 700–2900 mm |
| TD | 8.2 | 7.0 | 0.772 | 21–28.5 °C |
| CMD | 2.2 | 3.8 | 0.698 | 225–510 |
| MAT | 2.1 | 21.1 | 0.876 | 12–17 °C |
| EXT | 1.7 | 11.7 | 0.782 | 35–38 °C |
| AHM | 0 | 0 | 0.703 | 7.5–38.5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Y.; Guo, J.; Shen, X.; Wang, G.; Wang, T. Predicting the Bioclimatic Habitat Suitability of Ginkgo biloba L. in China with Field-Test Validations. Forests 2019, 10, 705. https://doi.org/10.3390/f10080705
Guo Y, Guo J, Shen X, Wang G, Wang T. Predicting the Bioclimatic Habitat Suitability of Ginkgo biloba L. in China with Field-Test Validations. Forests. 2019; 10(8):705. https://doi.org/10.3390/f10080705
Chicago/Turabian StyleGuo, Ying, Jing Guo, Xin Shen, Guibin Wang, and Tongli Wang. 2019. "Predicting the Bioclimatic Habitat Suitability of Ginkgo biloba L. in China with Field-Test Validations" Forests 10, no. 8: 705. https://doi.org/10.3390/f10080705
APA StyleGuo, Y., Guo, J., Shen, X., Wang, G., & Wang, T. (2019). Predicting the Bioclimatic Habitat Suitability of Ginkgo biloba L. in China with Field-Test Validations. Forests, 10(8), 705. https://doi.org/10.3390/f10080705