Relationship between Pine Wilt Disease Outbreaks and Climatic Variables in the Three Gorges Reservoir Region

Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Location
2.2. Pine Wilt Disease Datasets
2.3. Climatic Datasets
2.4. Data Analysis
3. Results
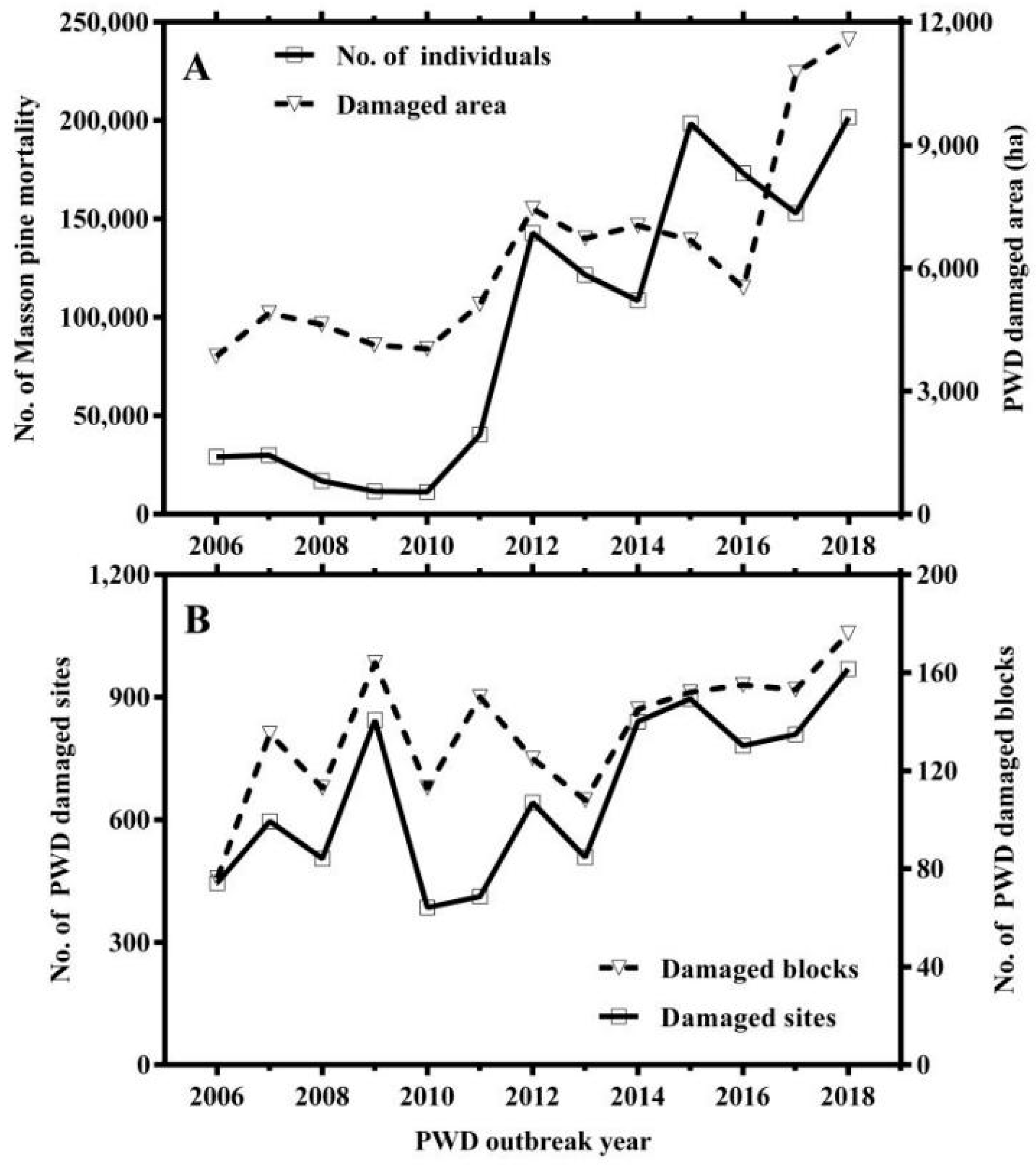
3.1. Overview of Pine Wilt Disease Outbreak
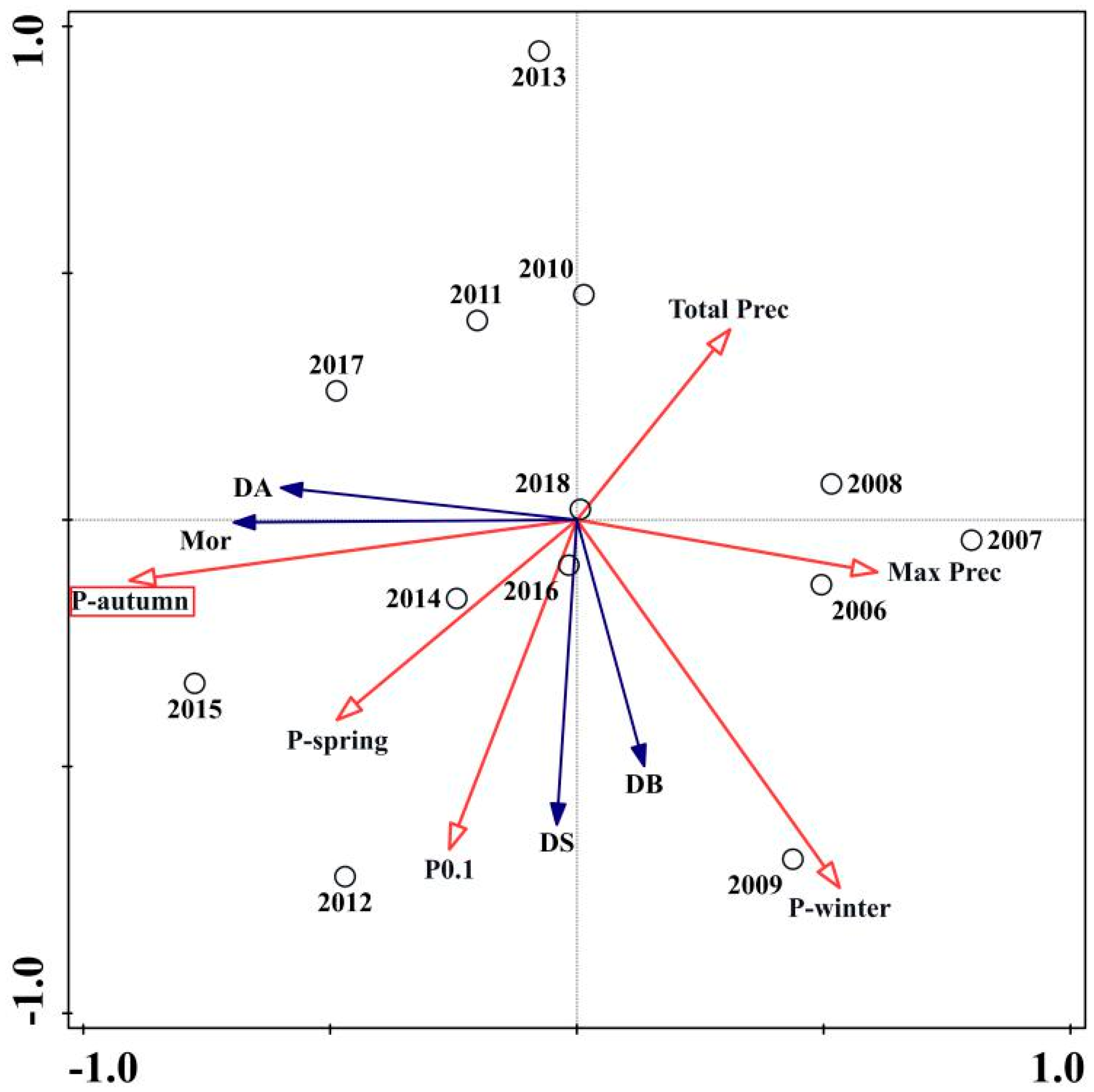
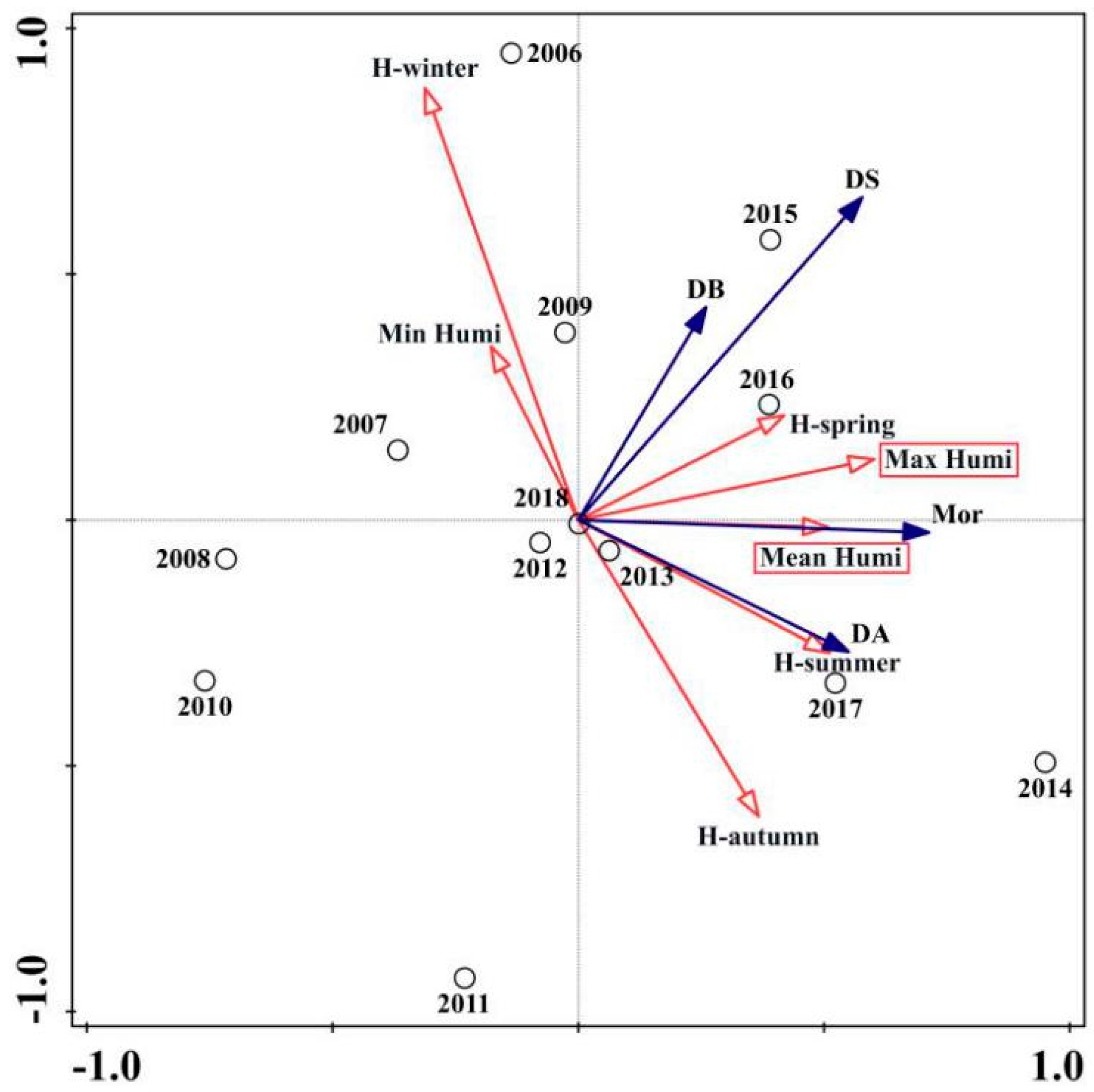
3.2. Ordination of PWD Variables and Climatic Variables
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Climate Change 2013. The Physical Science Basis: Working Group I Contribution to the 5th Assessment of the Intergovernmental Panel on Climate Change; Stocker, T.F., Qin, D., Plattner, G.K., Tignor, M.M.B., Allen, S.K., Boschung, J., Nauels, A., Xia, Y., Bex, V., Midgley, P.M., Eds.; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2014. [Google Scholar]
- Walther, G.R.; Roques, A.; Hulme, P.E.; Sykes, M.T.; Pysek, P.; Fuhn, I.; Zobel, M.; Bacher, S.; Dukat, Z.B.; Bugmann, H.; et al. Alien species in a warmer world: Risks and opportunities. Trends Ecol. Evol. 2009, 24, 686–693. [Google Scholar] [CrossRef] [PubMed]
- Jepsen, J.U.; Kapari, L.; Hagen, S.B.; Schott, T.; Vindstad, O.L.; Nilssen, A.C.; Ims, R.A. Rapid northwards expansion of a forest insect pest attributed to spring phenology matching with sub-Arctic birch. Glob. Chang. Biol. 2011, 17, 2071–2083. [Google Scholar] [CrossRef]
- Ikegami, M.; Jenkins, T.A. Estimate global risks of a forest disease under current and future climates using species distribution model and simple thermal model–pine wilt disease as a model case. For. Ecol. Manag. 2018, 409, 343–352. [Google Scholar] [CrossRef]
- Bale, J.S.; Masters, G.J.; Hodkinson, I.D.; Awmack, C.; Bezemer, T.M.; Brown, V.K.; Butterfield, J.; Buse, A.; Coulson, J.C.; Farrar, J.; et al. Herbivory in global climate change research: Direct effects of rising temperature on insect herbivores. Glob. Chang. Biol. 2002, 8, 1–16. [Google Scholar] [CrossRef]
- Tobin, P.C.; Nagarkatti, S.; Loeb, G.; Saunders, M.C. Historical and projected interactions between climate change and insect voltinism in a multivoltine species. Glob. Chang. Biol. 2008, 14, 951–957. [Google Scholar] [CrossRef]
- Kingsolver, J.G.; Ragland, G.J.; Diamond, S.E. Evolution in a constant environment: Thermal fluctuations and thermal sensitivity of laboratory and field populations of Manduca Sexta. Evolution 2009, 63, 537–541. [Google Scholar] [CrossRef]
- Sambaraju, K.R.; Carroll, A.L.; Zhu, J.; Stahl, K.; Moore, R.D.; Aukema, B.H. Climate change could alter the distribution of mountain pine beetle outbreaks in western Canada. Ecography 2012, 35, 211–223. [Google Scholar] [CrossRef]
- Paaijmans, K.P.; Heinig, R.L.; Seliga, R.A.; Blanford, J.I.; Blanford, S.; Murdock, C.C.; Thomas, M.B. Temperature variation makes ectotherms more sensitive to climate change. Glob. Chang. Biol. 2013, 19, 2373–2380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roques, A.; Zhao, L.L.; Sun, J.H.; Robinet, C. Pine wood nematode, pine wilt disease, vector beetle and pine tree: How a multiplayer system could reply to climate change. In Climate Change and Insect Pests; Björkman, C., Niemelä, P., Eds.; CABI publishing: Oxfordshire, UK, 2015; pp. 220–234. ISBN 9781780643786. [Google Scholar]
- David, G.; Giffard, B.; Piou, D.; Roques, A.; Jactel, H. Potential effects of climate warming on the survivorship of adult Monochamus galloprovincialis. Agric. For. Entomol. 2017, 19, 192–199. [Google Scholar] [CrossRef]
- Lindner, M.; Maroschek, M.; Netherer, S.; Kremer, A.; Barbati, A.; Gonzalo, J.G.; Seidl, R.; Delzon, S.; Corona, P.; Kolstrom, M.; et al. Climate change impacts, adaptive capacity, and vulnerability of European forest ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Calvão, T.; Duarte, C.M.; Pimentel, C.S. Climate and landscape patterns of pine forest decline after invasion by the pinewood nematode. For. Ecol. Manag. 2019, 433, 43–51. [Google Scholar] [CrossRef]
- Adams, H.D.; Guardiola-Claramonte, M.; Barron-Gafford, G.A.; Villegas, J.C.; Breshears, D.D.; Zou, C.B.; Troch, P.A.; Huxman, T.E. Temperature sensitivity of drought-induced tree mortality portends increased regional die-off under global-change-type drought. Proc. Natl. Acad. Sci. USA 2009, 106, 7063–7066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, C.D.; Macalady, A.K.; Chenchouni, H.; Bacgelet, D.; McDowell, N.; Vennetier, M.; Kitzbergerg, T.; Rigling, A.; Breshears, D.D.; Hogg, E.H.; et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 2010, 259, 660–684. [Google Scholar] [CrossRef] [Green Version]
- McDowell, N.G.; Beerling, D.J.; Breshears, D.D.; Fisher, R.A.; Raffa, K.; Stitt, M. The interdependence of mechanisms underlying climate-driven vegetation mortality. Trends Ecol. Evol. 2011, 26, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Pautasso, M.; Schlegel, M.; Holdenrieder, O. Forest health in a changing world. Microb. Ecol. 2015, 69, 826–842. [Google Scholar] [CrossRef] [PubMed]
- Garrett, K.A.; Dobson, A.D.M.; Kroschel, J.; Natarajan, B.; Orlandini, S.; Tonnang, H.E.Z.; Valdivia, C. The effects of climate variability and the color of weather time series on agricultural diseases and pests, and on decisions for their management. Agric. For. Meteorol. 2013, 170, 216–227. [Google Scholar] [CrossRef] [Green Version]
- Giblin-Davis, R.M.; Davies, K.A.; Morris, K.; Thomas, W.K. Evolution of parasitism in insect transmitted plant nematodes. J. Nematol. 2003, 35, 133–141. [Google Scholar]
- Zhao, B.G. Pine wilt disease in China. In Pine Wilt Disease; Zhao, B.G., Futai, K., Sutherland, J.R., Takeuchi, Y., Eds.; Springer: Tokyo, Japan, 2008; pp. 18–25. ISBN 978-4-431-75655-2. [Google Scholar]
- Abelleira, A.; Picoaga, A.; Mansilla, J.P.; Aguin, O. Detection of Bursaphelenchus xylophilus, causal agent of pine wilt disease on Pinus pinaster in Northwestern Spain. Plant Dis. 2011, 95, 776. [Google Scholar] [CrossRef]
- Dropkin, V.; Foudin, A.; Kondo, E.; Linit, M.; Smith, M. Pinewood nematode: A threat to US forest? Plant Dis. 1981, 65, 1022–1027. [Google Scholar] [CrossRef]
- Wingfield, M.J.; Blanchette, R.A.; Nicholls, T.H. Is the pine wood nematode an important pathogen in the United States? J. For. 1984, 82, 232–235. [Google Scholar]
- Pimentel, C.S.; Ayres, M.P.; Vallery, E.; Young, C.; Streett, D.A. Geographical variation in seasonality and life history of pine sawyer beetles Monochamus: Its relationship with phoresy by the pinewood nematode Bursaphelenchus xylophilus. Agric. For. Entomol. 2014, 16, 196–206. [Google Scholar] [CrossRef]
- Shi, J.; Luo, Y.Q.; Song, J.Y.; Wu, H.W.; Wang, L.; Wang, G.Z. Traits of Masson pine affecting attack of pine wood nematode. J. Integr. Plant Biol. 2007, 49, 1763–1771. [Google Scholar] [CrossRef]
- Gao, R.; Shi, J.; Huang, R.F.; Wang, Z.; Luo, Y.Q. Effects of pine wilt disease invasion on soil properties and Masson pine forest communities in the Three Gorges reservoir region, China. Ecol. Evol. 2015, 5, 1702–1716. [Google Scholar] [CrossRef] [PubMed]
- The 2019 No.4 Bulletin of National Forestry and Grassland Administration—The Epidemic Area of Pine Wilt Disease. Available online: http://www.forestry.gov.cn/sites/main/main/index.jsp (accessed on 1 February 2019).
- An, H.; Lee, S.; Cho, S.J. The effects of climate change on pine wilt disease in South Korea: Challenges and Prospects. Forests 2019, 10, 486. [Google Scholar] [CrossRef]
- Firmino, P.N.; Calvão, T.; Ayres, M.P.; Pimentel, C.S. Monochamus galloprovincialis and Bursaphelenchus xylophilus life history in an area severely affected by pine wilt disease: Implications for forest management. For. Ecol. Manag. 2017, 389, 105–115. [Google Scholar] [CrossRef]
- Pimentel, C.S.; Ayres, M.P. Latitudinal patterns in temperature-dependent growth rates of a forest pathogen. J. Biol. 2018, 72, 39–43. [Google Scholar] [CrossRef] [PubMed]
- Pasho, E.; Camarero, J.J.; Luis, M.; Vicente-Serrano, S.M. Spatial variability in large-scale and regional atmospheric drivers of Pinus halepensis growth in eastern Spain. Agric. For. Meteorol. 2011, 151, 1106–1119. [Google Scholar] [CrossRef]
- Dorman, M.; Perevolotsky, A.; Sarris, D.; Svoray, T. The effect of rainfall and competition intensity on forest response to drought: Lessons learned from a dry extreme. Oecologia 2015, 177, 1025–1038. [Google Scholar] [CrossRef]
- Kurz-Besson, C.B.; Lousada, J.L.; Gaspar, M.J.; Correia, I.E.; David, T.S.; Soares, P.M.M.; Cardoso, R.M.; Russo, A.; Varino, F.; Meriaux, C.; et al. Effects of recent minimum temperature and water deficit increases on Pinus pinaster radial growth and wood density in southern Portugal. Front. Plant Sci. 2016, 7, 1170. [Google Scholar] [CrossRef]
- Robinet, C.; Roques, A.; Pan, H.Y.; Fang, G.F.; Ye, J.R.; Zhang, Y.Z.; Sun, J.H. Role of human-mediated dispersal in the spread of the pinewood nematode in China. PLoS ONE 2009, 4, e4646. [Google Scholar] [CrossRef]
- Gruffudd, H.R.; Jenkins, T.A.R.; Evans, H.F. Using an evapo-transpiration model (ETpN) to predict the risk and expression of symptoms of pine wilt disease (PWD) across Europe. Biol. Invasions 2016, 18, 2823–2840. [Google Scholar] [CrossRef]
- Rutherford, T.A.; Webster, J.M. Distribution of pine wilt disease with respect to temperature in North America, Japan, and Europe. Can. J. For. Res. 1987, 17, 1050–1059. [Google Scholar] [CrossRef]
- Rutherford, T.A.; Mamiya, Y.; Webster, J.M. Nematode-induced pine wilt disease: Factors influencing its occurrence and distribution. For. Sci. 1990, 36, 145–155. [Google Scholar]
- Pérez, G.; Díez, J.J.; Ibeas, F.; Pajares, J.A. Assessing pine wilt disease risk under a climate change scenario in Northwestern Spain. In Managing Forest Ecosystems: The Challenge of Climate Change; Bravo, F., Jandl, R., LeMay, V., von Gadow, K., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 269–282. ISBN 978-1-4020-8343-3. [Google Scholar]
- Evans, S.; Evans, H.; Ikegami, M. Modeling PWN-induced wilt expression: A mechanistic approach. In Pine Wilt Disease: A Worldwide Threat to Forest Ecosystems; Mota, M.M., Vieira, P., Eds.; Springer: Dordrecht, The Netherland, 2008; pp. 259–278. ISBN 978-1-4020-8455-3. [Google Scholar]
- Mamiya, Y. Pathology of the pine wilt disease caused by Bursaphelenchus xylophilus. Annu. Rev. Phytopathol. 1983, 21, 201–220. [Google Scholar] [CrossRef] [PubMed]
- Takemoto, S. Population ecology of Bursaphelenchus xylophilus. In Pine Wilt Disease; Zhao, B.G., Futai, K., Sutherland, J.R., Takeuchi, Y., Eds.; Springer: Tokyo, Japan, 2008; pp. 105–122. ISBN 978-4-431-75655-2. [Google Scholar]
- Xi, Y.; Niu, S.K. The effect of climatic factors on pine wilt disease. For. Resour. Manag. 2008, 8, 70–76. [Google Scholar]
- Kong, W.N. The effects of temperature and humidity on the longevity of Monochamus Aalternatus Hope (Coleoptera: Cerambycidae). J. Shanxi Agric. Univ. 2006, 26, 294–295. [Google Scholar]
- Legendre, P.; Legendre, L. Numerical Ecology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2012; pp. 65–78. ISBN 9780444538680. [Google Scholar]
- Sturrock, R.N. Climate change and forest diseases: Using today’s knowledge to address future challenges. For. Syst. 2012, 21, 329–336. [Google Scholar] [CrossRef]
- Gao, R.H.; Luo, Y.Q.; Wang, Z.; Yu, H.J.; Shi, J. Patterns of biomass, carbon, and nitrogen storage distribution dynamics after the invasion of pine forests by Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae) in the three Gorges Reservoir Region. J. For. Res. 2018, 29, 459–470. [Google Scholar] [CrossRef]
- Naves, P.; Sousa, E. Threshold temperatures and degree-day estimates for development of post-dormancy larvae of Monochamus galloprovincialis (Coleoptera: Cerambycidae). J. Pest Sci. 2009, 82, 1–6. [Google Scholar] [CrossRef]
- Gao, R.H.; Ji, W.R.; Li, H.; Shi, J. The relationship between pine wilt disease variables and climatic variables. J. Shanxi Agric. Univ. 2019, 39, 32–40. [Google Scholar]
- Elad, Y.; Pertot, I. Climate change impacts on plant pathogens and plant diseases. J. Crop Improv. 2014, 28, 99–139. [Google Scholar] [CrossRef]
- Sternberg, E.D.; Thomas, M.B. Local adaptation to temperature and the implications for vector-borne diseases. Trends Parasitol. 2014, 30, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Hirata, A.; Nakamura, K.; Nakao, K.; Kominami, Y.; Tanaka, N.; Ohashi, H.; Takano, K.T.; Takeuchi, W.; Matsui, T. Potential distribution of pine wilt disease under future climate change scenarios. PLoS ONE 2017, 12, e0182837. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, K.; Maehara, N.; Aikawa, T.; Ichihara, Y. Research project to develop strategic action plan in the pine wilt disease unaffected area in Northern Japan. In Proceedings of the Book of Abstract of the Pine Wilt Disease Conference 2013, Braunschweig, Germany, 15–18 October 2013; Julius Kühn Institute, Federal Research Centre for Cultivated Plants: Quedlinburg, Germany, 2013; p. 66. [Google Scholar]
- Shinya, R.; Morisaka, H.; Takeuchi, Y.; Futai, K.; Ueda, M. Making headway in understanding pine wilt disease: What do we perceive in the postgenomic era? J. Biosci. Bioeng. 2013, 116, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Lv, Q.; Feng, Y.M.; Li, Y.X.; Wang, Y.L.; Zhang, X.Y. Temporal and spatial dynamic pattern of PWD distribution in China predicted under climate change scenario. Sci. Silv. Sin. 2015, 51, 119–126. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Description | Rationale |
|---|---|---|
| Max temp | Maximum temperature during pine wilt disease (PWD) life cycle | Pine wilt disease has been observed mainly in areas where the mean daily summer temperature exceed 20 °C for several weeks [36,37,38]. |
| Mean temp | Mean temperature during PWD life cycle | |
| Min temp | Minimum temperature during PWD life cycle | |
| Mtspring | Mean temperature in spring quarter (3–5) | |
| MTsummer | Mean temperature in summer quarter (6–8) | High temperature and seasonal drought causing water deficit drive potential tree evaporation and weaken the tree’s defense capacity against the pine wood nematode, thus favored pathogen and vector development and likely to trigger an outbreak of pine wilt disease [10,20,36,39]. |
| MTautumn | Mean temperature in autumn quarter (9–11) | |
| MTwinter | Mean temperature in winter quarter (12, 1, 2) | |
| T20 | Number of days with temperature at or above 20 °C | |
| MT20 | Mean temperature of days with temperature at or above 20 °C | |
| TOpt | Number of days with optimum temperature | |
| MTOpt | Mean temperature of days with optimum temperature | Pine wood nematode cannot get enough effective accumulated temperature when the mean temperature is lower than 10 °C [20]. |
| TUnfav | Number of days with unfavorable temperature | |
| T10 | Number of days with temperature below 10 °C | Temperature directly affect the development rate of B. xylophilus, the optimum temperature range for pine wilt disease development is between 10–25 °C [10,20,40]. |
| MT10 | Mean temperature of days with temperature below 10 °C | |
| T25 | Number of days with temperature above 25 °C | |
| MT25 | Mean temperature of days with temperature above 25 °C | |
| T28 | Number of days with temperature above 28 °C | High temperature could have negative effects on nematode development (above 28 °C) as well as on nematode reproductive process (above 35 °C) [10,40,41]. |
| MT28 | Mean temperature of days with temperature above 28 °C | |
| T35 | Number of days with temperature above 35 °C | The distribution of Monnchamus species is constrained by thermal barriers, especially by low winter temperatures that regulate the survival of the overwintering fifth-instar larvae [10]. |
| MT35 | Mean temperature of days with temperature above 35 °C | |
| PSB-ddegg | Degree-day accumulation for 50% egg hatch from June through September | A relatively cold condition, which the temperature range between 10–15 °C, is necessary for the growth and development of pine sawyer beetle (PSB) larva from October to December [20]. |
| PSB-ddadult | Degree-day accumulation for adult emergence from March through May | |
| PSB-dd | Degree-day accumulation for one generation during PSB’s life cycle | |
| T10–15 | Number of days with temperature between 10–15 °C from October to December | In China, at least 1200 degree-days were required for the development of PSB generation from egg to adult, with 528 degree-days for adult emergence [20]. |
| MT10–15 | Mean temperature of days with temperature between 10–15 °C from October to December | |
| T19–28 | Number of days with temperature between 19–28 °C from June to September | |
| MT19–28 | Mean temperature of days with temperature between 19–28 °C from June to September | The optimum temperature range for PSB hatching is between 19–28 °C. As to 50% egg hatch, at least 350 degree-days are required through the growing season [10,20]. |
| T0 | Number of days with minimum temperature at or below 0 °C | |
| MT0 | Mean temperature of days with minimum temperature at or below 0 °C | |
| Total prec | Total precipitation during PWD life cycle | |
| Max prec | Maximum daily precipitation during PWD life cycle | |
| P0.1 | Number of days with precipitation at or above 0.1 mm per day | Too much precipitation have a significantly effects on PSB’s flight performance and feeding capacity [10,11,20,42,43]. |
| Pspring | Precipitation in spring quarter (3–5) | |
| Psummer | Precipitation in summer quarter (6–8) | |
| Pautumn | Precipitation in autumn quarter (9–11) | |
| Pwinter | Precipitation in winter quarter (12,1,2) | |
| Max humi | Maximum relative humidity during PWD life cycle | |
| Mean humi | Mean relative humidity during PWD life cycle | |
| Min humi | Minimum relative humidity during PWD life cycle | Relative humidity is prevalent from March to May, being correlated positively with pine wilt disease epidemic degree [20,42]. |
| Hspring | Relative humidity in spring quarter (3–5) | |
| Hsummer | Relative humidity in summer quarter (6–8) | |
| Hautumn | Relative humidity in autumn quarter (9–11) | |
| Hwinter | Relative humidity in winter quarter (12,1,2) | |
| Ext wind | Extreme wind speed from May to September | The flight capacity of PSB can significantly affected by wind speed and higher monthly wind speed facilitates the spread and diffusion of PSB over long distance [20,42]. |
| Max wind | Maximum wind speed from May to September | |
| Mean wind | Mean wind speed from May to September |
| PWD Damaged Blocks | PWD Damaged Sites | Masson Pine Mortality | PWD Damaged Area | |
|---|---|---|---|---|
| PWD damaged blocks | 1 | 0.811 * | 0.255 | 0.253 |
| PWD damaged sites | 1 | 0.502 | 0.411 | |
| Masson pine mortality | 1 | 0.772 * | ||
| PWD damaged area | 1 |
| Variables | Contribution% | F-Ratio | p-Value | |
|---|---|---|---|---|
| Temperature | MTautumn | 40.5 | 7.5 * | 0.018 |
| MTsummer | 14.9 | 3.9 * | 0.032 | |
| MTOpt | 9.6 | 4.4 * | 0.024 | |
| T10–15 | 5.7 | 3.5 * | 0.044 | |
| TOpt | 4.7 | 4.8 * | 0.03 | |
| T35 | 9.8 | 2 | 0.152 | |
| MT19–28 | 9.9 | 3.2 | 0.088 | |
| T0 | 2.4 | 3.8 | 0.082 | |
| Min-T | 1.1 | 2.3 | 0.144 | |
| TUnfav | 0.8 | 2.6 | 0.14 | |
| T28 | 0.4 | 1.8 | 0.33 | |
| MT10–15 | 0.2 | <0.1 | 1 | |
| Precipitation | Pautumn | 75.5 | 6.8 * | 0.026 |
| Pwinter | 6.7 | 0.6 | 0.534 | |
| Pspring | 5.8 | 0.5 | 0.588 | |
| P0.1 | 10.1 | 0.8 | 0.402 | |
| Max prec | 1.8 | 0.1 | 0.888 | |
| Total prec | <0.1 | <0.1 | 1 | |
| Relative humidity | Max humi | 33.1 | 0.9 ** | 0.008 |
| Mean humi | 14.1 | 3.7 * | 0.038 | |
| Hautumn | 13.5 | 2.4 | 0.13 | |
| Min humi | 15.5 | 1.2 | 0.308 | |
| Hsummer | 7.3 | 0.5 | 0.544 | |
| Hwinter | 6 | 0.4 | 0.612 | |
| Hspring | 10.6 | 0.6 | 0.504 | |
| Mean wind | 63.7 | 5.9 * | 0.036 | |
| Wind speed | Ext wind | 21.2 | 2.2 | 0.176 |
| Max wind | 15.1 | 1.7 | 0.188 | |
| Variables | Canonical Axes | Eigenvalues | Cumulative Explained Variation (%) | Pseudo Canonical Correlation | Cumulative Explained Fitted Variation (%) | Sum of All Eigenvalues | Sum of All Canonical Eigenvalues |
|---|---|---|---|---|---|---|---|
| Temperature | RDA1 | 0.901 | 90.05 | 1 | 90.44 | 1 | 0.996 |
| RDA2 | 0.073 | 97.36 | 0.978 | 97.79 | |||
| RDA3 | 0.018 | 99.16 | 1 | 99.6 | |||
| RDA4 | 0.004 | 99.56 | 0.964 | 100 | |||
| Precipitation | RDA1 | 0.496 | 49.58 | 0.744 | 92.57 | 1 | 0.536 |
| RDA2 | 0.036 | 53.16 | 0.672 | 99.26 | |||
| RDA3 | 0.003 | 53.42 | 0.689 | 99.74 | |||
| RDA4 | 0.001 | 53.56 | 0.265 | 100 | |||
| Relative humidity | RDA1 | 0.538 | 53.83 | 0.78 | 90.59 | 1 | 0.594 |
| RDA2 | 0.045 | 58.29 | 0.71 | 98.09 | |||
| RDA3 | 0.009 | 59.26 | 0.687 | 99.72 | |||
| RDA4 | 0.002 | 59.43 | 0.492 | 100 | |||
| Wind speed | RDA1 | 0.596 | 59.63 | 0.814 | 97.88 | 1 | 0.609 |
| RDA2 | 0.012 | 60.83 | 0.419 | 99.85 | |||
| RDA3 | 0.001 | 60.92 | 0.223 | 100 | |||
| RDA4 | 0.304 | 91.27 | 0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, R.; Wang, Z.; Wang, H.; Hao, Y.; Shi, J. Relationship between Pine Wilt Disease Outbreaks and Climatic Variables in the Three Gorges Reservoir Region. Forests 2019, 10, 816. https://doi.org/10.3390/f10090816
Gao R, Wang Z, Wang H, Hao Y, Shi J. Relationship between Pine Wilt Disease Outbreaks and Climatic Variables in the Three Gorges Reservoir Region. Forests. 2019; 10(9):816. https://doi.org/10.3390/f10090816
Chicago/Turabian StyleGao, Ruihe, Zhuang Wang, Haixiang Wang, Yanping Hao, and Juan Shi. 2019. "Relationship between Pine Wilt Disease Outbreaks and Climatic Variables in the Three Gorges Reservoir Region" Forests 10, no. 9: 816. https://doi.org/10.3390/f10090816
APA StyleGao, R., Wang, Z., Wang, H., Hao, Y., & Shi, J. (2019). Relationship between Pine Wilt Disease Outbreaks and Climatic Variables in the Three Gorges Reservoir Region. Forests, 10(9), 816. https://doi.org/10.3390/f10090816