Interregional Crown Width Models for Individual Trees Growing in Pure and Mixed Stands in Austria

Abstract
:1. Introduction
2. Materials and Methods
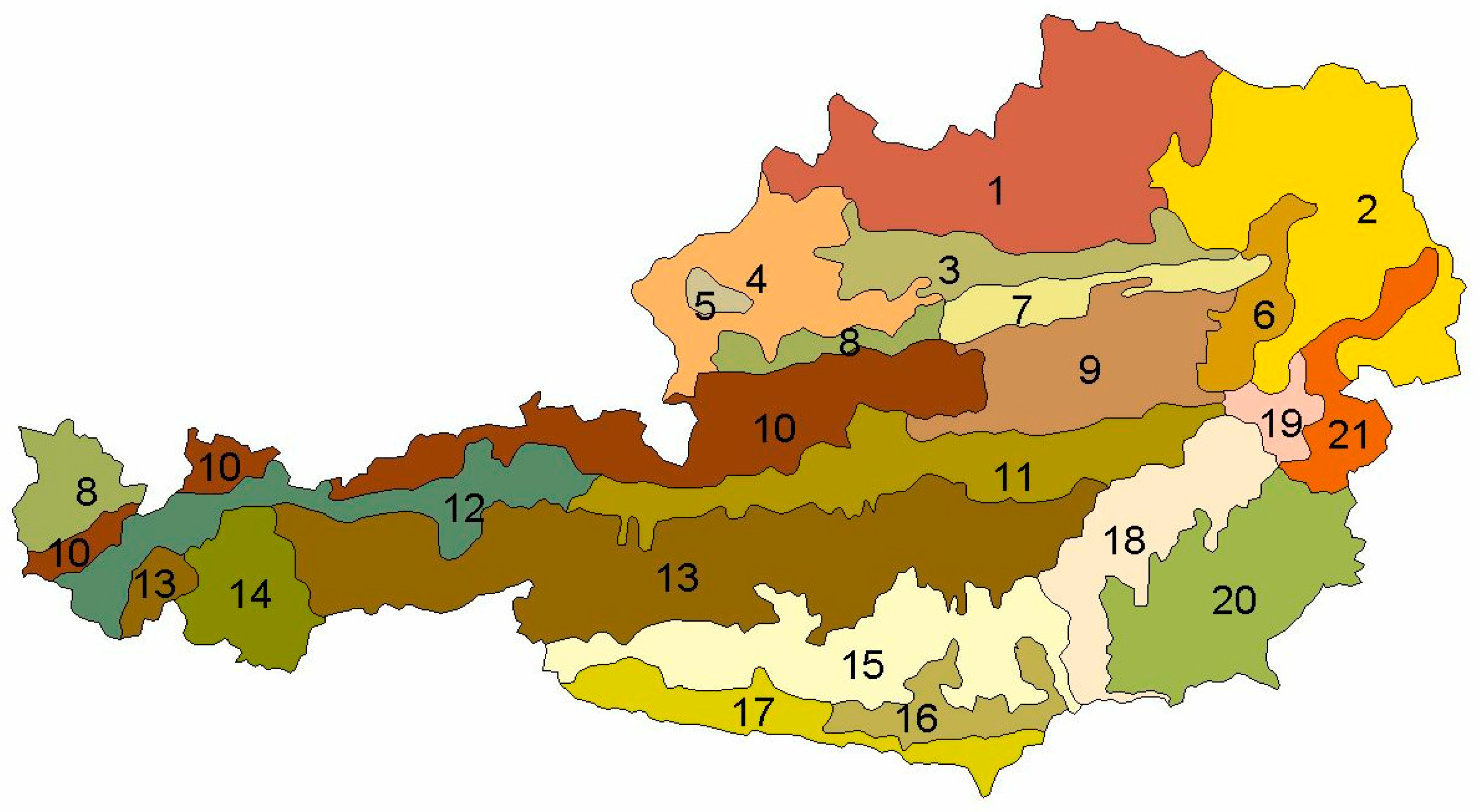
2.1. Material
2.2. Statistical Analysis
3. Results
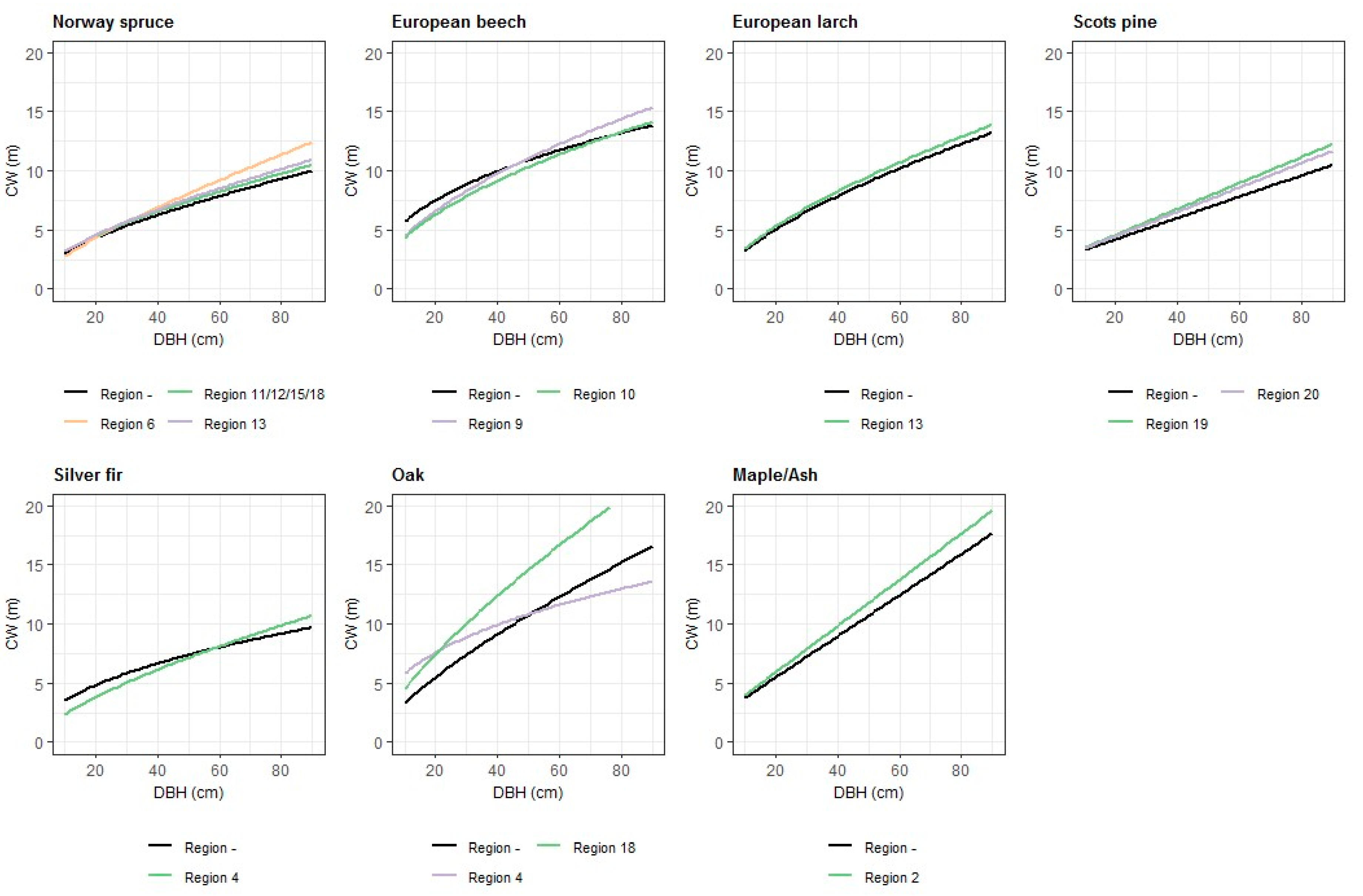
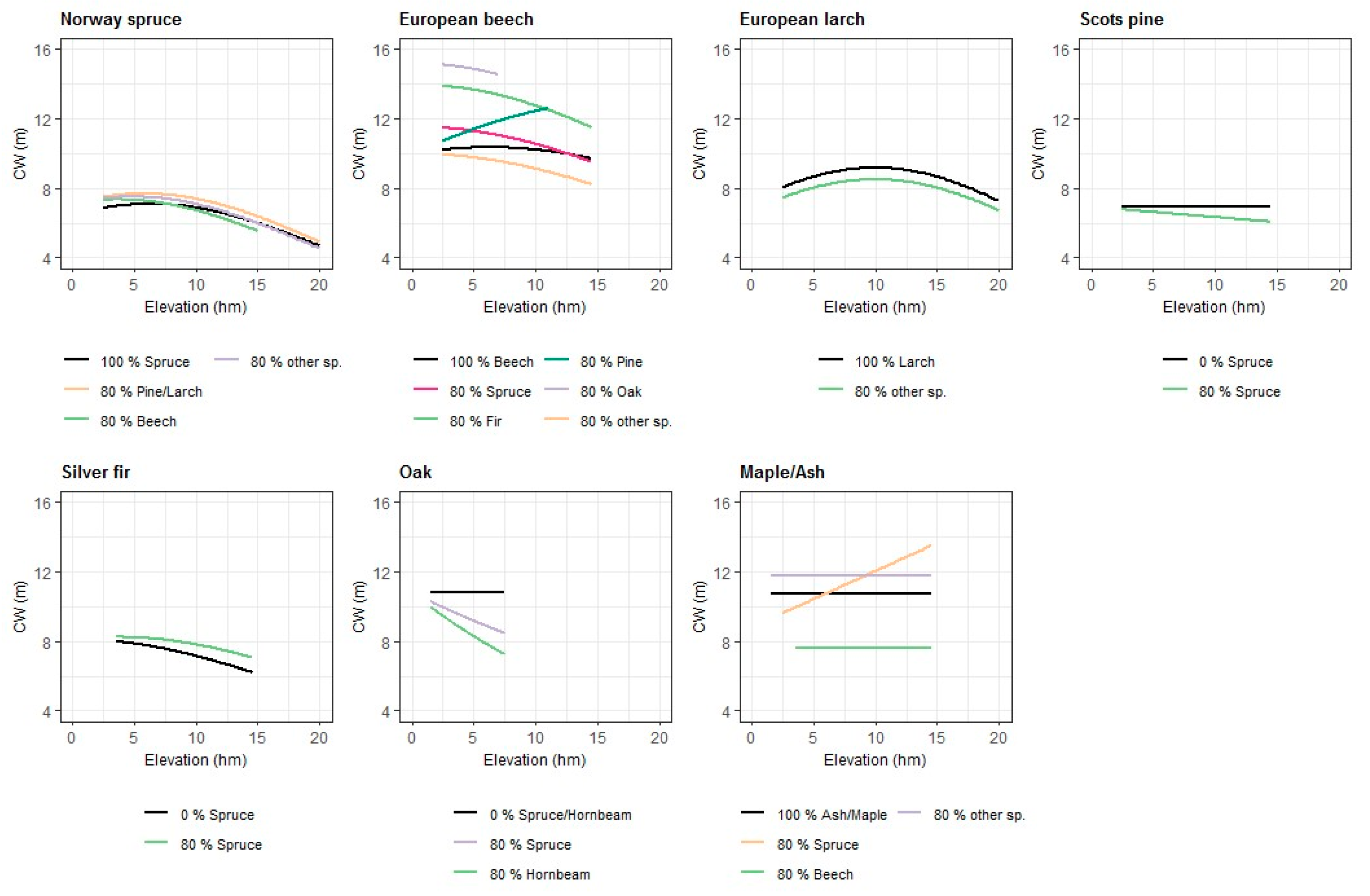
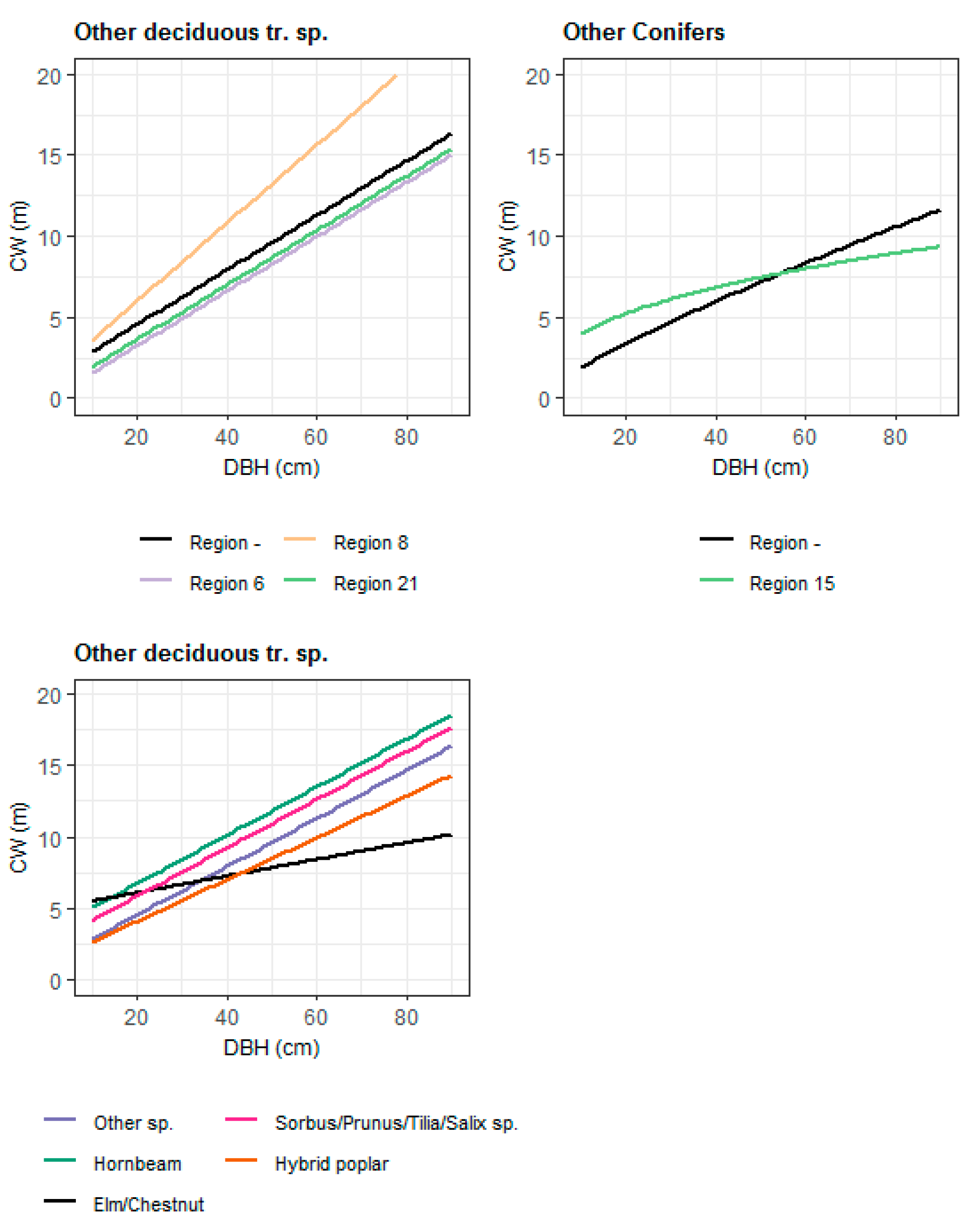
3.1. Crown Width Models for Conifer Species
3.2. Crown Width Models for Broadleaf Species
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

References
- Reinhardt, E.; Crookston, N.L.; Beukema, S.J.; Kurz, W.A.; Greenough, J.A.; Robinson, D.C.E.; Lutes, D.C. Purpose and Applications. The Fire and Fuels Extension to the Forest Vegetation Simulator: Updated Model Documentation; Reinhardt, E., Crookston, N.L., Eds.; U.S. Department of Agriculture, Forest Service and Rocky Mountain Research Station: Fort Collings, CO, USA, 2010; pp. 1–8.
- Crookston, N.L.; Dixon, E.D. The forest vegetation simulator: A review of its structure, content, and applications. Comput. Electron. Agric. 2005, 49, 60–80. [Google Scholar] [CrossRef]
- Castaldi, C.; Vacchiano, G.; Marchi, M.; Corona, P. Projecting nonnative douglas fir plantations in Southern Europe with the forest vegetation simulator. For. Sci. 2017, 63, 101–110. [Google Scholar] [CrossRef] [Green Version]
- Crookston, N.L.; Rehfeldt, G.E.; Dixon, G.E.; Weiskittel, A.R. Addressing climate change in the forest vegetation simulator to assess impacts on landscape forest dynamics. For. Ecol. Manag. 2010, 260, 1198–1211. [Google Scholar] [CrossRef]
- Hasenauer, H.; Monserud, R.A. Biased predictions for tree height increment models developed from smoothed ‘data’. Ecol. Model. 1997, 98, 13–22. [Google Scholar] [CrossRef]
- Biging, G.S.; Dobbertin, M. Evaluation of competition indices in individual tree growth models. For. Sci. 1995, 41, 360–370. [Google Scholar]
- Fu, L.; Sharma, H.; Hao, K.; Tang, S. A generalized interregional nonlinear mixed-effects crown width model for Prince Rupprecht larch in northern China. For. Ecol. Manag. 2017, 389, 364–373. [Google Scholar] [CrossRef]
- Monserud, R.A.; Sterba, H. A basal area increment model for individual trees growning in even- and uneven-aged forest stands in Austria. For. Ecol. Manag. 1996, 80, 57–80. [Google Scholar] [CrossRef]
- Ledermann, T.; Neumann, M. Biomass equations from data of old long-term experimental plots. Austrian J. For. Sci. 2006, 1, 47–64. [Google Scholar]
- Carbalho, J.P.; Parresol, B.R. Additivity in tree biomass components of Pyrenean oak (Quercus pyrenaica Willd.). For. Ecol. Manag. 2003, 179, 269–276. [Google Scholar] [CrossRef]
- Krajicek, C.L.; Brinkman, K.A.; Gingrich, S.F. Crown competition—A measure of density. For. Sci. 1961, 7, 35–42. [Google Scholar]
- Paulo, J.A.; Faias, S.P.; Ventrua-Giroux, C.; Tome, M. Estimation of stand crown cover using a generalized crown diameter model: Application for the analysis of Portuguese cork oak stands stocking evolution. iForest 2015, 9, 437–444. [Google Scholar] [CrossRef] [Green Version]
- Shaw, J.D. Models for Estimation and Simulation of Crown and Canopy Cover. In Proceedings of the Fifth Annual Forest Inventory and Analysis Symposium, New Orleans, LA, USA, 18–20 November 2003; U.S. Department of Agriculture Forest Service: Washington, DC, USA; pp. 183–191. [Google Scholar]
- Pukkala, T.; Becker, P.; Kuuluvainen, T.; Oker-Blom, P. Predicting spatial distribution of direct radiation below forest canopies. Agric. For. Meteorol. 1991, 55, 295–307. [Google Scholar] [CrossRef]
- Elmungheira, M.I.; Elmamoun, H.O. Diameter at Breast Height—Crown Width Prediction Models for Anogeissus Leiocarpus (DC.) Guill & Perr and Combretum Hartmannianum Schweinf. J. For. Prod. Ind. 2014, 3, 191–197. [Google Scholar]
- Sharma, R.P.; Vacek, Z.; Vacek, S. Individual tree crown width models for Norway spruce and European beech in Czech Republic. For. Ecol. Manag. 2016, 366, 208–220. [Google Scholar] [CrossRef]
- Fu, L.; Sun, H.; Sharma, R.P.; Lei, Y.; Zhang, H.; Tang, S. Nonlinear mixed-effects crown width models for individual trees of Chines fir (Cunninghamia lanceolata) in south-central China. For. Ecol. Manag. 2013, 302, 210–220. [Google Scholar] [CrossRef]
- Davies, O.; Pommerening, A. The contribution of structural indices to the modelling of Sitka spruce (Picea stichensis) and birch (Betula spp.) crowns. For. Ecol. Manag. 2008, 256, 68–77. [Google Scholar] [CrossRef]
- Xu, H.; Sun, Y.; Wang, X.; Wang, J.; Fu, Y. Linear mixed-effects models to describe individual tree crown width for China-fir in Fujian Province, Southeast China. PLoS ONE 2015, 10, e0122257. [Google Scholar]
- Jucker, T.; Bourlaud, C.; Coomes, D.A. Crown plasticity enables trees to optimize canopy packing in mixed species forests. Funct. Ecol. 2015, 29, 1078–1086. [Google Scholar] [CrossRef] [Green Version]
- Pretzsch, H. The effect of tree crown allometry on community dynamics in mixed-species stands versus monocultures. A review and perspectives for modeling and silvicultural regulation. Forests 2019, 10, 810. [Google Scholar] [CrossRef] [Green Version]
- Sterba, H.; Dirnberger, G.; Ritter, T. Vertical distribution of leaf area of European larch (Larix decidua Mill. And Norway Spruce (Picea abies (L.) Karst.) in pure and mixed stands. Forests 2019, 10, 570. [Google Scholar] [CrossRef] [Green Version]
- Barbeito, I.; Dassot, M.; Bayer, D.; Collet, C.; Drössler, L.; Löf, M.; Rio, M.; Ruiz-Peinado, R.; Forrester, D.I.; Bravo-Oviedo, A.; et al. Terrestrial laser scanning reveals differences in crown structure of Fagus sylvatica in mixed vs. pure European forests. For. Ecol. Manag. 2017, 405, 381–390. [Google Scholar] [CrossRef]
- Thorpe, H.C.; Astrup, A.; Trowbridge, A.; Coates, K.D. Competition and tree crowns: A neighbourhood analysis of three boreal tree species. For. Ecol. Manag. 2010, 259, 1586–1596. [Google Scholar] [CrossRef]
- Yoshida, T.; Kamitani, T. Interspecific competition among three canopy-tree species in a mixed-species even-aged forest of central Japan. For. Ecol. Manag. 2000, 137, 221–230. [Google Scholar] [CrossRef]
- Ledermann, T.; Kindermann, G.; Gschwantner, T. National woody biomass projection systems based on forest inventory in Austria. In Forest Inventory-Based Projection Systems for Wood and Biomass Availability; Barreiro, S., Schelhaas, M.J., McRoberts, R.E., Kändler, G., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 79–95. [Google Scholar]
- Braun, M.; Fritz, D.; Weiss, P.; Braschel, N.; Büchsenmeister, R.; Freudenschuß, A.; Gschwantner, T.; Jandl, R.; Ledermann, T.; Neumann, M.; et al. A holistic assessment of greenhouse gas dynamics from forests to the effects of wood products use in Austria. Carbon Manag. 2016, 7, 271–283. [Google Scholar] [CrossRef]
- Jelem, H.; Kilian, W. Standortsaufnahme im rahmen der österr. Forstinventur–eine forstpolitische entscheidungshilfe. Allg. Forstztg. 1972, 83, 295–297. [Google Scholar]
- Bitterlich, W. Die Winkelzählprobe. Allg. Forst-u. Holzwirtaschtsztg 1948, 59, 4–5. [Google Scholar] [CrossRef]
- Pretzsch, H. Zum einfluß des baumverteilungsmusters auf den bestandeszuwachs. Allg. Forst-u. J. Ztg. 1995, 166, 190–201. [Google Scholar]
- Pinheiro, J.C.; Bates, D.M. Mixed-Effects Models in S and S-PLUS; Spring: New York, NY, USA, 2000; pp. 3–56. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Kunetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest Package: Tests in linear mixed effects models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar]
- Wonn, H.T.; O’Hara, K.L. Height: diameter ratios and stability relationships for four Northern Rocky Mountain tree species. West. J. Appl. For. 2001, 16, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Ledermann, T. A non-linear model to predict crown recession of Norway spruce (Picea abies [L.] Karst.) in Austria. Eur. J. For. Res. 2011, 130, 521–531. [Google Scholar] [CrossRef]
- Stage, A.R. An expression for the effect of slope, aspect and habitat type on tree growth. For. Sci. 1976, 22, 457–460. [Google Scholar]
- Sönmez, T. Diameter at breast height-crown diameter prediction models for Picea orientalis. Afr. J. Agric. 2009, 4, 215–219. [Google Scholar]
- Bragg, D.C. A local basal area adjustment for crown width prediction. North. J. Appl. For. 2001, 18, 22–28. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.; Sharma, R.P.; Wang, G.; Tang, S. Modelling a system of nonlinear additive crown width models applying seemingly unrelated regression for Prince Rupprecht larch in northern China. For. Ecol. Manag. 2017, 386, 71–80. [Google Scholar] [CrossRef]
- Lei, Y.; Fu, L.; Affleck, D.L.R.; Nelson, A.S.; Shen, C.; Wang, M.; Zheng, J.; Ye, Q.; Yang, G. Additivity of nonlinear tree crown width models: Aggregated and disaggregated model structures using nonlinear simultaneous equations. For. Ecol. Manag. 2018, 427, 372–382. [Google Scholar] [CrossRef]
- Foli, E.G.; Alder, D.; Miller, D.G.; Swaine, M.D. Modelling growing space requirements for some tropical tree species. For. Ecol. Manag. 2003, 173, 79–88. [Google Scholar] [CrossRef]
- Grote, R. Estimation of crown radii and crown projection area from stem size and tree position. Ann. For. Sci. 2003, 60, 393–402. [Google Scholar] [CrossRef] [Green Version]
- Hegyi, F. A simulation model for managing jack-pine stands. In Growth Models for Tree and Stand Simulation; Fries, J., Ed.; Royal College of Forestry: Stockholm, Sweden, 1974; pp. 74–90. [Google Scholar]
- Reineke, L.H. Perfecting a Stand density index for even-aged forests. J. Agric. Res. 1933, 46, 627–638. [Google Scholar]
- Hasenauer, H. Dimensional relationships of open-grown trees in Austria. For. Ecol. Manag. 1997, 96, 197–206. [Google Scholar] [CrossRef]
- Mayer, H.; Eckerhart, G.; Nather, G.; Rachoy, J.; Zuckrigl, K. Die Waldgebiete und Wuchsbezirke Österreichs. Cbl. f. d. ges. Forstw. 1971, 88, 129–164. [Google Scholar]
- Ledermann, T. Description of PrognAus for Windows 2.2. In Sustainable Forest Management—Growth Models for Europe; Hasenauer, H., Ed.; Springer: New York, NY, USA, 2006; pp. 71–78. [Google Scholar]
- Krucek, M.; Trochta, J.; Cibulka, M.; Kral, K. Beyond the cones: How crown shape plasticity alters aboveground competition for space and light—Evidence from terrestrial laser scanning. Agric. For. Meteorol. 2019, 264, 188–199. [Google Scholar] [CrossRef]
- Hanewinkel, M.; Cullmann, D.A.; Schelhaas, M.J.; Nabuurs, G.J.; Zimmermann, N.E. Climate change may cause severe loss in the economic value of European forest land. Nat. Clim. Chang. 2013, 3, 203–207. [Google Scholar] [CrossRef]
- Dyderski, M.K.; Paz, S.; Frelich, L.E.; Jagodzinski, A.M. How much does climate change threaten European forest tree species distributions? Glob. Chang. Biol. 2018, 24, 1150–1163. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Variable/Definition | Unit |
|---|---|---|
| CW | Crown width | m |
| DBH | Diameter at breast height (1.3 m) | cm |
| H | Height of a tree | m |
| HCB | Height of a trees’s crown base | m |
| CR | Crown ratio = 1 − HCB/H | |
| H/D | Height − diameter ratio = H [cm]/DBH [cm] | |
| ELEV | Sea level of the sample plot | hectometre |
| EXP | Azimuth of the aspect of the sample area | radiant |
| SL | Slope gradient of the sample area | percentage |
| Region_x | Growth region [28] in which the sample plot is located | |
| PS_x | Relative proportion of basal area of species x (x … species code) | |
| PSL_x | Relative proportion of basal area of species x (x … species code) considering only trees of the same crown layer | |
| Species | Code | N | DBH in cm | H in m | CW in m | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Min | Mean | Max | Min | Mean | Max | Min | Mean | Max | |||
| Picea Abies Norway spruce | 1 | 5436 | 10.5 | 34.8 | 115.3 | 4.8 | 24.5 | 53.3 | 1.2 | 5.6 | 16.4 |
| Abies Alba Silver fir | 2 | 272 | 10.7 | 40.5 | 89.0 | 7.3 | 27.5 | 43.0 | 2.6 | 6.9 | 13.9 |
| Larix deciduas European larch | 3 | 627 | 10.9 | 42.5 | 92.9 | 8.1 | 27.2 | 42.7 | 1.5 | 7.5 | 16.6 |
| Pinus Sylvestris Scots pine | 4 | 612 | 10.6 | 33.6 | 68.5 | 6.0 | 23.6 | 38.6 | 1.3 | 5.4 | 10.9 |
| Pinus Nigra Austrian pine | 5 | 97 | 11.3 | 37.0 | 79.6 | 6.6 | 18.5 | 34.3 | 2.0 | 5.9 | 12.1 |
| Pinus Cembra Stone pine | 6 | 48 | 19.4 | 42.4 | 80.0 | 9.0 | 16.5 | 30.3 | 2.6 | 5.7 | 8.7 |
| Pinus strobus Weymouth pine | 7 | 3 | 34.2 | 36.5 | 37.6 | 24.0 | 26.2 | 28.6 | 5.4 | 6.3 | 6.8 |
| Pseudotsuga Menziesii Douglas fir | 8 | 5 | 18.4 | 36.8 | 70.8 | 12.6 | 22.4 | 42.1 | 4.6 | 6.6 | 9.2 |
| Fagus Sylvatica European beech | 10 | 811 | 10.7 | 35.3 | 106.7 | 6.1 | 23.8 | 39.9 | 2.2 | 9.3 | 22.4 |
| Quercus sp. Oak | 11 | 238 | 10.7 | 36.3 | 90.3 | 8.7 | 21.3 | 36.8 | 2.1 | 8.1 | 21.0 |
| Carpinus betulus Hornbeam | 12 | 68 | 11.1 | 23.1 | 65.1 | 10.8 | 17.1 | 33.3 | 0.2 | 7.7 | 19.4 |
| Fraxinus sp. Ash | 13 | 192 | 10.9 | 28.3 | 71.7 | 6.9 | 23.5 | 38.8 | 1.8 | 7.0 | 20.5 |
| Acer sp. Maple | 14 | 96 | 11.4 | 29.7 | 110.4 | 8.8 | 20.0 | 32.3 | 3.0 | 7.8 | 19.7 |
| Ulmus sp. Elm | 15 | 8 | 13.3 | 36.2 | 60.8 | 13.5 | 21.5 | 32.0 | 6.7 | 9.5 | 16.3 |
| Castanea sativa Spanish chestnut | 16 | 15 | 10.5 | 25.8 | 51.5 | 11.1 | 17.5 | 25.5 | 4.3 | 6.6 | 10.7 |
| Robinia pseudoacacia Robinia | 17 | 27 | 10.6 | 20.0 | 37.6 | 9.6 | 17.7 | 29.8 | 2.4 | 5.5 | 10.4 |
| Sorbus sp., Prunus sp. | 18 | 26 | 10.7 | 22.1 | 48.7 | 6.5 | 15.2 | 25.9 | 1.6 | 6.2 | 13.2 |
| Betula sp. Birch | 20 | 86 | 10.5 | 26.2 | 52.2 | 8.8 | 19.1 | 33.0 | 2.0 | 5.7 | 11.2 |
| Alnus glutinosa Black alder | 21 | 94 | 11.4 | 26.0 | 46.1 | 9.9 | 21.7 | 29.9 | 2.3 | 5.7 | 10.9 |
| Alnus incana White alder | 22 | 25 | 11.3 | 17.7 | 30.9 | 9.0 | 13.8 | 24.9 | 0.1 | 3.7 | 7.1 |
| Tilia sp. | 23 | 29 | 11.6 | 28.5 | 79.9 | 10.4 | 19.1 | 34.3 | 3.6 | 7.3 | 18.3 |
| Populus tremula, Populus Alba Trembling and white poplar | 24 | 22 | 11.1 | 29.9 | 67.9 | 11.2 | 19.8 | 31.0 | 3.3 | 7.2 | 14.3 |
| Populus Nigra Black poplar | 25 | 8 | 25.5 | 47.9 | 77.5 | 17.9 | 26.1 | 32.4 | 3.8 | 9.2 | 15.1 |
| Populus sp. X Hybrid poplar | 26 | 15 | 17.7 | 46.9 | 86.2 | 14.8 | 23.7 | 31.5 | 3.8 | 8.9 | 12.9 |
| Salix sp. Willow | 27 | 22 | 12.8 | 24.3 | 56.5 | 7.8 | 13.1 | 30.3 | 5.9 | 7.9 | 12.5 |
| Tree Species | Picea abies | Larix decidua | Pinus sylvestris | Abies alba | Other sp. |
|---|---|---|---|---|---|
| Depend Variable | ln(CW) | ln(CW) | CW | ln(CW) | ln(CW) |
| Intercept | 0.052386 | −0.4307885 | 3.7155099 | 1.4180761 | −0.8317729 |
| Region_4 | - | - | - | −0.9031416 | - |
| Region_6 | −0.4215087 | - | - | - | - |
| Region_13 | - | 0.0482368 | - | - | - |
| Region_15 | - | - | - | - | 1.7352668 |
| Region_11/12/15/18 | 0.046185 | - | - | - | - |
| DBH | - | - | 0.0898499 | - | - |
| Region_19 * DBH | - | - | 0.0194008 | - | - |
| Region_20 * DBH | - | - | 0.0130925 | - | - |
| ln (DBH) | 0.486831 | 0.6384635 | - | 0.4629727 | 0.8211466 |
| Region_4 * ln(DBH) | - | - | - | 0.2227205 | - |
| Region_6 * ln(DBH) | 0.1416563 | - | - | - | - |
| Region_13 * ln(DBH) | 0.0193874 | - | - | - | - |
| Region_15 * ln(DBH) | - | - | - | - | −0.4343612 |
| H/D | −0.0031566 | - | - | - | - |
| (H/D)² | - | −0.0000207 | - | - | - |
| sqrt (H/D) | - | - | −0.3717171 | - | - |
| ln (H/D) | - | - | - | −0.2695577 | - |
| CR | 0.6153011 | - | - | - | - |
| CR² | −0.2123078 | - | - | - | - |
| ln (CR) | - | 0.0987259 | - | - | 0.1951676 |
| sqrt (CR) | - | - | 3.0042626 | - | - |
| ELEV | - | 0.046124 | - | - | −0.0275227 |
| ELEV * DBH | 0.0003257 | - | - | - | - |
| ELEV² | −0.0014439 | −0.0023145 | - | −0.0012659 | - |
| ELEV² * DBH | −0.0000151 | - | - | - | - |
| cos (EXP) * SL | −0.0001333 | - | - | - | 0.0003318 |
| sin (EXP) * SL | 0.0002676 | - | - | - | 0.0023244 |
| sqrt (PS_1) | −0.1657646 | - | - | - | - |
| PS_3² | - | 0.0791549 | - | - | - |
| PS_1 * ELEV | - | - | - | 0.0110309 | - |
| sqrt (PS_1) * ELEV | 0.0112108 | - | - | - | - |
| sqrt (PS_3/4) * ELEV | 0.0046689 | - | - | - | - |
| sqrt (PS_10) * ELEV | −0.0058753 | - | - | - | - |
| PSL_1² * ELEV | - | - | −0.0911981 | - | - |
| Picea abies | Fagus sylvatica | Larix decidua | Pinus sylvestris | Abies alba | Quercus sp. | Acer/Fraxinus sp. | Other Conifers | Other Broadleaf Sp. | |
|---|---|---|---|---|---|---|---|---|---|
| SD Plot () | 0.102 | 0.136 | 0.113 | 0.707 | 0.098 | 0.110 | 1.024 | 0.139 | 0.883 |
| SD Residual () | 0.147 | 0.184 | 0.156 | 0.878 | 0.148 | 0.162 | 1.357 | 0.172 | 1.263 |
| ICC | 0.325 | 0.351 | 0.344 | 0.393 | 0.305 | 0.318 | 0.363 | 0.397 | 0.328 |
| AIC | −3952 | −69 | −260 | 1815 | −132 | −79 | 1115 | 5 | 1704 |
| Marginal R² | 0.74 | 0.64 | 0.69 | 0.54 | 0.68 | 0.79 | 0.68 | 0.61 | 0.70 |
| Conditional R² | 0.83 | 0.76 | 0.80 | 0.72 | 0.78 | 0.86 | 0.80 | 0.76 | 0.80 |
| Tree Species | Fagus sylvatica | Quercus sp. | Acer/Fraxinus sp. | Other sp. |
|---|---|---|---|---|
| Depend Variable | ln(CW) | ln(CW) | CW | CW |
| Intercept | 1.0910323 | −1.0471094 | 1.1865967 | 0.4423783 |
| Region_4 | - | 1.3611911 | - | - |
| Region_6 | - | - | - | −1.3161855 |
| Region_9 | −0.5924605 | - | - | - |
| Region_10 | −0.5657388 | - | - | - |
| Region_21 | - | - | - | −0.9284775 |
| Region_18 | - | 0.304054 | - | - |
| SP_12 | - | - | - | 2.1919433 |
| SP_15/16 | - | - | - | 3.7354244 |
| SP_18/23/27 | - | - | - | 1.3092625 |
| DBH | - | - | 0.1735585 | 0.1683018 |
| Region_2 *DBH | - | - | 0.021643 | - |
| Region_8 * DBH | - | - | - | 0.0722409 |
| SP_15/16 * DBH | - | - | - | −0.1101649 |
| SP_26 * DBH | - | - | - | −0.0224963 |
| ln(DBH) | 0.4036294 | 0.7315097 | - | - |
| Region_4 * ln(DBH) | - | -0.3467259 | - | - |
| Region_9 * ln(DBH) | 0.1551168 | - | - | - |
| Region_10 * ln(DBH) | 0.1305651 | - | - | - |
| H/D | −0.0029587 | - | - | - |
| CR | - | - | 3.3979624 | 2.3711534 |
| ln (CR) | 0.1684201 | - | - | - |
| sqrt (CR) | - | 0.7273862 | - | - |
| ELEV² | −0.000946 | - | - | −0.0059434 |
| cos (EXP) * SL | 0.00073 | - | - | −0.0026763 |
| sin (EXP) * SL | 0.0005842 | - | - | 0.0166171 |
| PSL_1 | 0.1809567 | - | - | - |
| PSL_1² | - | - | −4.662721 | - |
| PSL_2² | 0.5235613 | - | - | - |
| PSL_11² | 0.6546167 | - | - | - |
| PSL_13/14² | - | - | −1.1155706 | - |
| sqrt (PSL_10) | - | - | 2.4700418 | - |
| PS_1 * ELEV | - | −0.0405627 | - | - |
| PS_12 * ELEV | - | −0.0660866 | - | - |
| PSL_1² * ELEV | - | - | 0.5035608 | - |
| PSL_4² * ELEV | 0.0485298 | - | - | - |
| PSL_10² * ELEV | 0.0118516 | - | - | - |
| sqrt (PSL_10) * DBH | - | - | −0.1433859 | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buchacher, R.; Ledermann, T. Interregional Crown Width Models for Individual Trees Growing in Pure and Mixed Stands in Austria. Forests 2020, 11, 114. https://doi.org/10.3390/f11010114
Buchacher R, Ledermann T. Interregional Crown Width Models for Individual Trees Growing in Pure and Mixed Stands in Austria. Forests. 2020; 11(1):114. https://doi.org/10.3390/f11010114
Chicago/Turabian StyleBuchacher, Rafael, and Thomas Ledermann. 2020. "Interregional Crown Width Models for Individual Trees Growing in Pure and Mixed Stands in Austria" Forests 11, no. 1: 114. https://doi.org/10.3390/f11010114
APA StyleBuchacher, R., & Ledermann, T. (2020). Interregional Crown Width Models for Individual Trees Growing in Pure and Mixed Stands in Austria. Forests, 11(1), 114. https://doi.org/10.3390/f11010114